
Productive Performance and Cecum Microbiota Analysis of Broiler Chickens Supplemented with β-Mannanases and Bacteriophages—A Pilot Study

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Facilities and Care of Experimental Animals
2.2. Experimental Design and Animal Management
2.3. Productive Performance
2.4. Systemic Humoral Immune Response
2.5. Quantification of Intestinal Immunoglobulin A (IgA) Antibodies
2.6. Morphometric Index
2.7. Evaluation of Gut Morphology
2.8. DNA Extraction, PCR, and Library Preparation for Sequencing
2.9. Amplicon Sequence Analysis
2.10. Statistical Analysis
3. Results
3.1. Productive Performance
3.2. Systemic Humoral Immune Response
3.3. Morphometric Index
3.4. Evaluation of Gut Morphology
3.5. Cecum Microbiota Analysis
3.5.1. Summary of DNA Sequence Data
3.5.2. Alpha Diversity Analysis
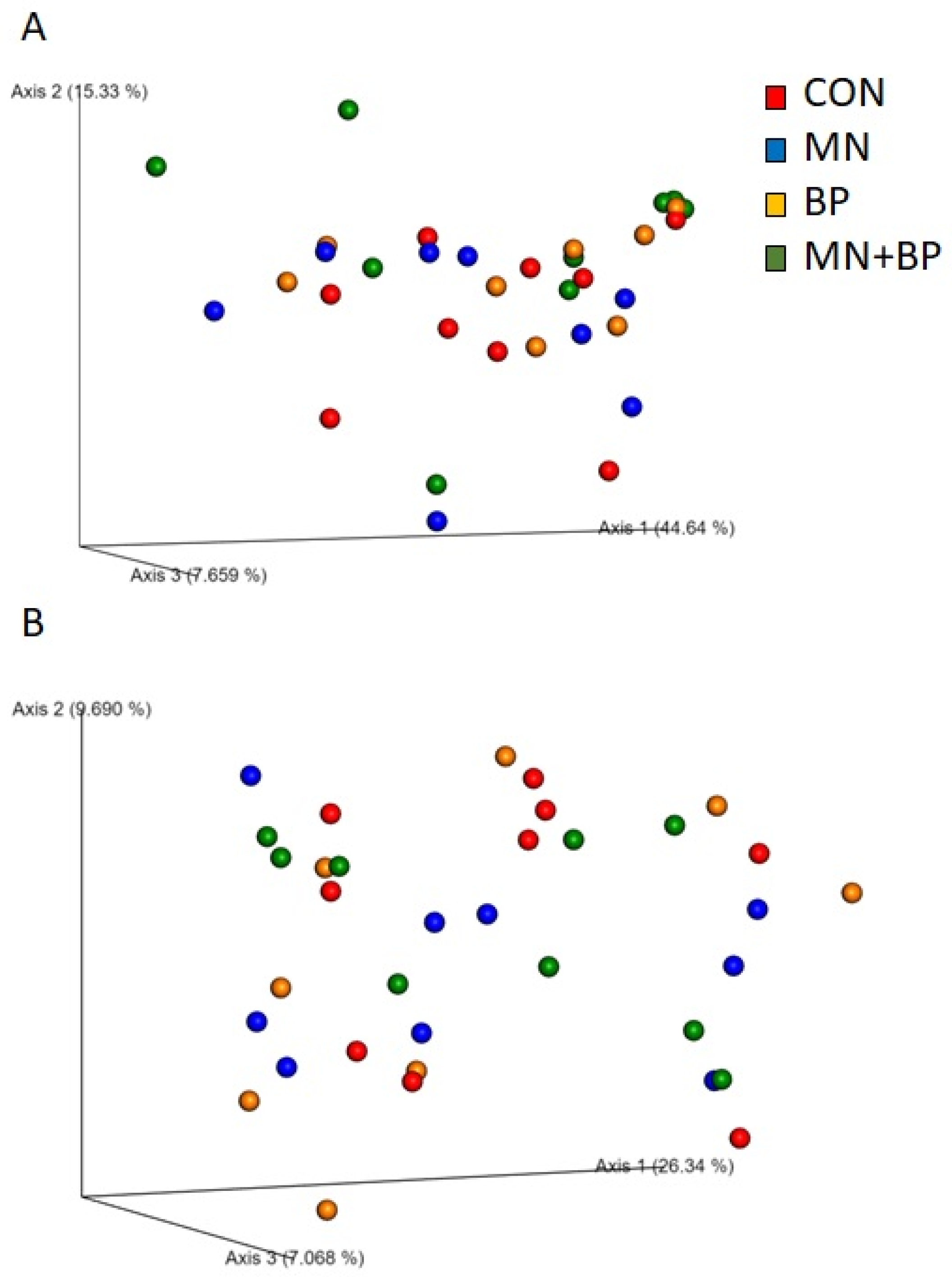
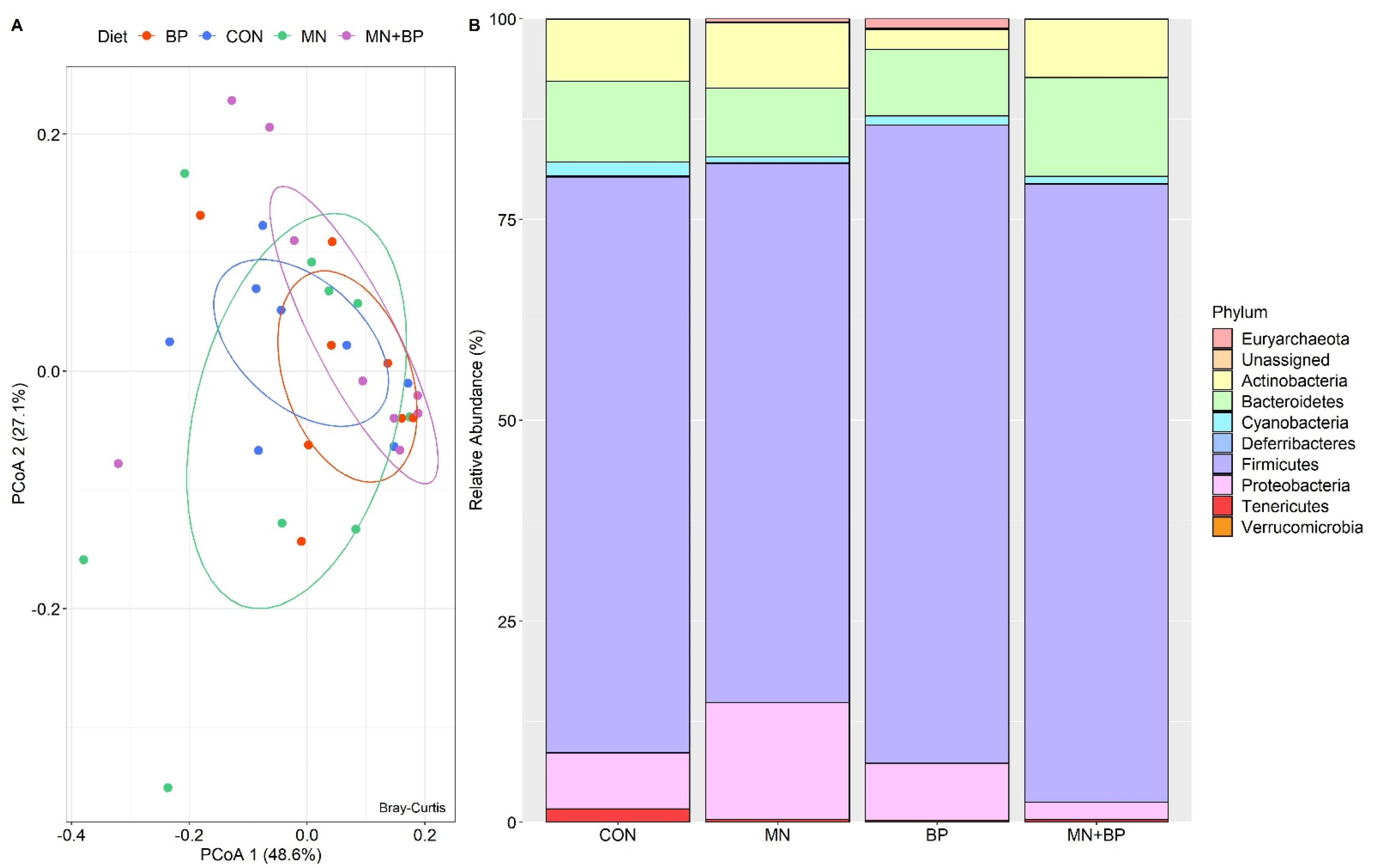
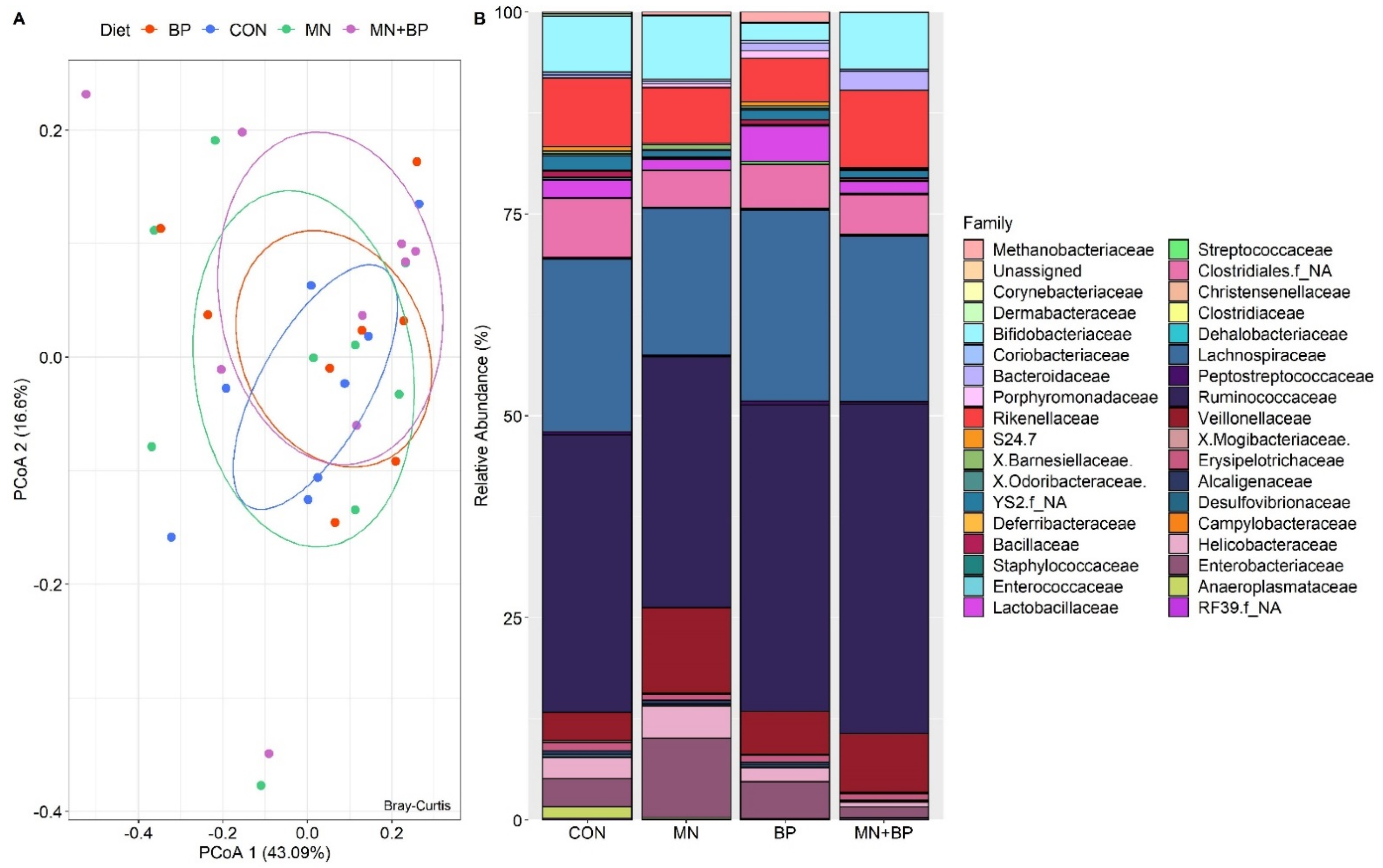
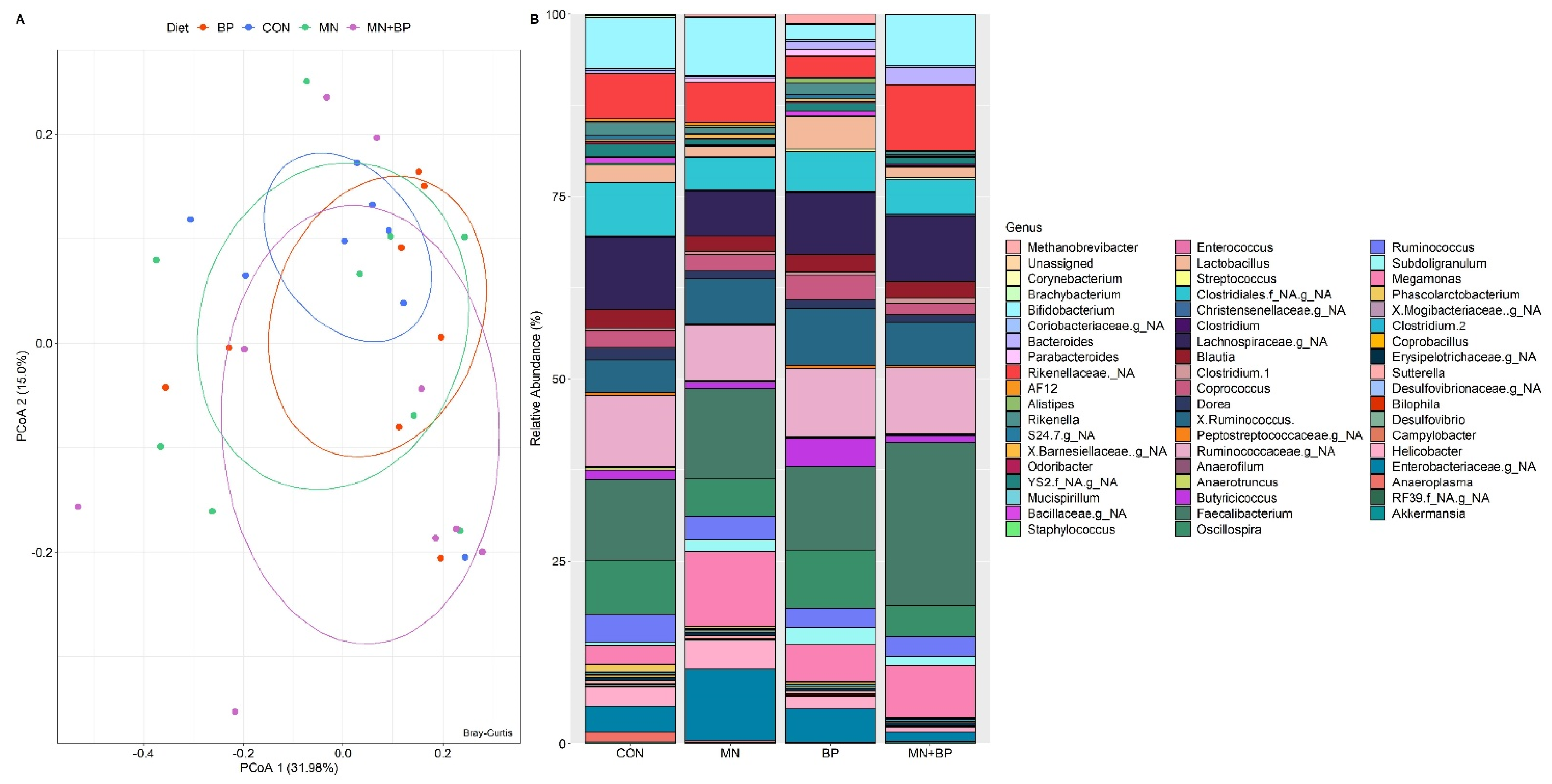
3.5.3. Beta Diversity Analysis
3.5.4. Taxonomic Assignments
- Taxonomic Assignment at the Phylum Level
- Taxonomic Assignment at the Family Level
- Taxonomic Assignment at the Genus Level
4. Discussion
4.1. Productive Performance
4.2. Humoral Immune Response
4.3. Cecum Microbiota Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caly, D.L.; D’Inca, R.; Auclair, E.; Drider, D. Alternatives to antibiotics to prevent necrotic enteritis in broiler chickens: A microbiologist’s perspective. Front. Microbiol. 2015, 6, 1336. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S. Role of nutraceuticals in gut health and growth performance of poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Apajalahti, J.; Vienola, K. Interaction between chicken intestinal microbiota and protein digestion. Anim. Feed. Sci. Technol. 2016, 221, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiome in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Schokker, D.; Jansman, A.J.M.; Veninga, G.; De Bruin, N.; Vastenhouw, S.A.; De Bree, F.M.; Bossers, A.; Rebel, J.M.J.; Smits, M.A. Perturbation of microbiota in one-day old broiler chickens with antibiotic for 24 hours negatively affects intestinal immune development. BMC Genom. 2017, 18, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudabos, A.M.; Al-Atiyat, R.; Albatshan, H.A.; AlJassim, R.; Aljumaah, M.R.; Alkhulaifi, M.M.; Stanley, D. Effects of concentration of corn distillers dried grains with solubles and enzyme supplementation on cecal microbiota and performance in broiler chickens. Appl. Microbiol. Biotechnol. 2017, 101, 7017–7026. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-B.; Stanley, D.; Rodgers, N.; Swick, R.; Moore, R. Two necrotic enteritis predisposing factors, dietary fishmeal and Eimeria infection, induce large changes in the caecal microbiota of broiler chickens. Vet. Microbiol. 2014, 169, 188–197. [Google Scholar] [CrossRef]
- Stanley, D.; Wu, S.-B.; Rodgers, N.; Swick, R.; Moore, R.J. Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens. PLoS ONE 2014, 9, e104739. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lilburn, M.; Yu, Z. Intestinal Microbiota of Broiler Chickens as Affected by Litter Management Regimens. Front. Microbiol. 2016, 7, 593. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Bessegatto, J.A.; Alfieri, A.A.; Weese, J.S.; Filho, J.A.B.; Oba, A. Different antibiotic growth promoters induce specific changes in the cecal microbiota membership of broiler chicken. PLoS ONE 2017, 12, e0171642. [Google Scholar] [CrossRef] [PubMed]
- Proctor, A.; Phillips, G.J. Differential Effects of Bacitracin Methylene Disalicylate (BMD) on the Distal Colon and Cecal Microbiota of Young Broiler Chickens. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.O.O.; Hugerth, L.W.; Bengtsson, C.; Alisjahbana, A.; Seifert, M.; Kamal, A.; Sjöling, Å.; Midtvedt, T.; Norin, E.; Du, J.; et al. Bacteriophages Synergize with the Gut Microbial Community to Combat Salmonella. mSystems 2018, 3, e00119-18. [Google Scholar] [CrossRef] [Green Version]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage therapy to combat bacterial infections in poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef]
- Yeh, Y.; Purushothaman, P.; Gupta, N.; Ragnone, M.; Verma, S.; de Mello, A. Bacteriophage application on red meats and poultry: Effects on Salmonella population in final ground products. Meat Sci. 2017, 127, 30–34. [Google Scholar] [CrossRef]
- Grant, A.; Parveen, S.; Schwarz, J.; Hashem, F.; Vimini, B. Reduction of Salmonella in ground chicken using a bacteriophage. Poult. Sci. 2017, 96, 2845–2852. [Google Scholar] [CrossRef]
- Borie, C.; Albala, I.; Sànchez, P.; Sánchez, M.L.; Ramírez, S.; Navarro, C.; Morales, M.A.; Retamales, J.; Robeson, J. Bacteriophage Treatment Reduces Salmonella Colonization of Infected Chickens. Avian Dis. 2008, 52, 64–67. [Google Scholar] [CrossRef]
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the impact of bacteriophages in the treatment of Salmonella in broiler chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Lee, J.T.; Latham, R.; Carter, B.; Kogut, M.H. Changes in immune and metabolic gut response in broilers fed β-mannanase in β-mannan-containing diets. Poult. Sci. 2017, 96, 4307–4316. [Google Scholar] [CrossRef]
- Caldas, J.V.; Vignale, K.; Boonsinchai, N.; Wang, J.; Putsakum, M.; England, J.A.; Coon, C.N. The effect of β-mannanase on nutrient utilization and blood parameters in chicks fed diets containing soybean meal and guar gum. Poult. Sci. 2018, 97, 2807–2817. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Jo, N.; Lee, J.-J.; Lee, J.-H.; Kam, D.-K.; Seo, J.; Kebreab, E.; Seo, S. Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves. Animals 2021, 11, 372. [Google Scholar] [CrossRef]
- Gómez-Verduzco, G.; Cortes-Cuevas, A.; López-Coello, C.; Ávila-González, E.; Nava, G.M. Dietary supplementation of mannan-oligosaccharide enhances neonatal immune responses in chickens during natural exposure to Eimeria spp. Acta Vet. Scand. 2009, 51, 11. [Google Scholar] [CrossRef] [Green Version]
- Perozo-Marín, F.; Nava, J.; Mavárez, Y.; Arenas, E.; Serje, P.; Briceño, M. Caracterización morfométrica de los órganos linfoides en pollos de engorde de la línea ross criados bajo condiciones de campo en el estado Zulia, Venezuela. Rev. Cient. Fac. Cienc. Vet. Univ. Zulia 2004, 14, 1–18. [Google Scholar]
- Mikel, U.V.; Armed Forces Institute of Pathology (US). Advanced Laboratory Methods in Histology and Pathology; American Registry of Pathology: Silver Spring, MD, USA, 1994; ISBN 1881041131. [Google Scholar]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2016, 71, 672–685. [Google Scholar] [CrossRef] [Green Version]
- Latham, E.A.; Weldon, K.K.; Wickersham, T.; Coverdale, J.A.; Pinchak, W.E. Responses in the rumen microbiome of Bos taurus and indicus steers fed a low-quality rice straw diet and supplemented protein. J. Anim. Sci. 2018, 96, 1032–1044. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Buttigieg, P.L.; Ramette, A. A guide to statistical analysis in microbial ecology: A community-focused, living review of multivariate data analyses. FEMS Microbiol. Ecol. 2014, 90, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Lu, P.; Choi, J.; Yang, C.; Mogire, M.; Liu, S.; Lahaye, L.; Adewole, D.; Rodas-Gonzalez, A.; Yang, C. Effects of antibiotic growth promoter and dietary protease on growth performance, apparent ileal digestibility, intestinal morphology, meat quality, and intestinal gene expression in broiler chickens: A comparison. J. Anim. Sci. 2020, 98, skaa254. [Google Scholar] [CrossRef]
- Latham, R.; Williams, M.; Walters, H.; Carter, B.; Lee, J. Efficacy of β-mannanase on broiler growth performance and energy utilization in the presence of increasing dietary galactomannan. Poult. Sci. 2018, 97, 549–556. [Google Scholar] [CrossRef]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- Xu, B.; Fu, J.; Zhu, L.; Li, Z.; Jin, M.; Wang, Y. Overall assessment of antibiotic substitutes for pigs: A set of meta-analyses. J. Anim. Sci. Biotechnol. 2021, 12, 3. [Google Scholar] [CrossRef]
- Wang, J.P.; Yan, L.; Lee, J.H.; Kim, I.H. Evaluation of Bacteriophage Supplementation on Growth Performance, Blood Characteristics, Relative Organ Weight, Breast Muscle Characteristics and Excreta Microbial Shedding in Broilers. Asian-Australas. J. Anim. Sci. 2013, 26, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Shashidhara, R.; Devegowda, G. Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult. Sci. 2003, 82, 1319–1325. [Google Scholar] [CrossRef]
- Halas, V.; Nochta, I. Mannan Oligosaccharides in Nursery Pig Nutrition and Their Potential Mode of Action. Animals 2012, 2, 261–274. [Google Scholar] [CrossRef] [Green Version]
- De Barros, V.R.S.M.; Lana, G.R.Q.; Lana, S.R.V.; Lana, A.; Cunha, F.S.A.; Neto, J.V.E. β-mannanase and mannan oligosaccharides in broiler chicken feed. Ciência Rural 2014, 45, 111–117. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Chen, Y.; Li, Z.; Cao, Y. Effects of β-mannanase expressed by Pichia pastoris in corn–soybean meal diets on broiler performance, nutrient digestibility, energy utilization and immunoglobulin levels. Anim. Feed. Sci. Technol. 2010, 159, 59–67. [Google Scholar] [CrossRef]
- Saleem, K.; Rahman, A.; Pasha, T.N.; Mahmud, A.; Hayat, Z. Effects of dietary organic acids on performance, cecal microbiota, and gut morphology in broilers. Trop. Anim. Health Prod. 2020, 52, 3589–3596. [Google Scholar] [CrossRef]
- Chacher, M.; Kamran, Z.; Ahsan, U.; Ahmad, S.; Koutoulis, K.; Din, H.Q.U.; Cengiz, O. Use of mannan oligosaccharide in broiler diets: An overview of underlying mechanisms. World’s Poult. Sci. J. 2017, 73, 831–844. [Google Scholar] [CrossRef]
- Sinovec, Z.; Markovic, R.; Gledic, D. Influence of Bio-Mos on broilers performances and gut morphology. In Proceedings of the 15th European Symposium on Poultry Nutrition, Balatonfüred, Hungary, 25–29 September 2005; pp. 353–355. [Google Scholar]
- Mesa, D.; Lammel, D.; Balsanelli, E.; Sena, C.; Noseda, M.; Caron, L.F.; Cruz, L.M.; Pedrosa, F.O.; Souza, E.M. Cecal Microbiota in Broilers Fed with Prebiotics. Front. Genet. 2017, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides. Nutrients 2018, 10, 1055. [Google Scholar] [CrossRef] [Green Version]
- Torok, V.A.; Allison, G.E.; Percy, N.J.; Ophel-Keller, K.; Hughes, R.J. Influence of Antimicrobial Feed Additives on Broiler Commensal Posthatch Gut Microbiota Development and Performance. Appl. Environ. Microbiol. 2011, 77, 3380–3390. [Google Scholar] [CrossRef] [Green Version]
- Tamai, K.; Akashi, Y.; Yoshimoto, Y.; Yaguchi, Y.; Takeuchi, Y.; Shiigai, M.; Igarashi, J.; Hirose, Y.; Suzuki, H.; Ohkusu, K. First case of a bloodstream infection caused by the genus Brachybacterium. J. Infect. Chemother. 2018, 24, 998–1003. [Google Scholar] [CrossRef]
- Angelopoulou, A.; Warda, A.K.; Hill, C.; Ross, R.P. Non-antibiotic microbial solutions for bovine mastitis–live biotherapeutics, bacteriophage, and phage lysins. Crit. Rev. Microbiol. 2019, 45, 564–580. [Google Scholar] [CrossRef]
- Bindari, Y.R.; Moore, R.J.; Van, T.T.H.; Walkden-Brown, S.W.; Gerber, P.F. Microbial taxa in dust and excreta associated with the productive performance of commercial meat chicken flocks. Anim. Microbiome 2021, 3, 66. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, e1987783. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Lv, Y.-W.; Long, J.; Chen, J.-M.; He, J.-M.; Ruan, X.-Z.; Zhu, H.-B. Butyrate Improves the Metabolic Disorder and Gut Microbiome Dysbiosis in Mice Induced by a High-Fat Diet. Front. Pharmacol. 2019, 10, 1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridgeman, S.C.; Northrop, W.; Melton, P.E.; Ellison, G.C.; Newsholme, P.; Mamotte, C.D. Butyrate generated by gut microbiota and its therapeutic role in metabolic syndrome. Pharmacol. Res. 2020, 160, 105174. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg) | Starter (1–21 d) | Finisher (22–49 d) |
|---|---|---|
| Corn | 570.405 | 539.30 |
| Soybean meal | 370.925 | 289 |
| Vegetable oil | 18.425 | 20.4 |
| Calcium carbonate | 14.75 | 14.5 |
| Orthophosphate | 10.55 | 10.1 |
| Salt | 3.5 | 3.5 |
| DL-methionine | 3.12 | 3.5 |
| L-lysine HCl | 2.85 | 4 |
| L-threonine | 0.725 | - |
| Mineral and Vitamin premix * | 3.5 | 3.5 |
| Phytase | 0.55 | 0.55 |
| Avelut® (pigment) | - | 6.0 |
| Coccidiostat | 0.5 | 5.0 |
| Choline chloride 60% | 0.05 | 0.5 |
| Antioxidant ** | 0.15 | 0.15 |
| Total | 1000 | 1000 |
| Calculated composition | ||
| Metabolizable energy Kcal/kg | 3010 | 3200 |
| Crude protein (%) | 22 | 19 |
| Digestible lysine (%) | 1.44 | 0.94 |
| Digestible Met + Cys (%) | 0.9 | 0.73 |
| Total calcium (%) | 0.96 | 0.85 |
| Available phosphorus (%) | 0.48 | 0.42 |
| Treatment | Weight Gain (g) | Feed Consumption (g) | Feed Conversion Ratio (kg/kg) | Carcass Yield (%) | Mortality (%) |
|---|---|---|---|---|---|
| Growth Promoter | |||||
| AGP | 2873 | 5410 | 1.89 | 69.7 a | 5.1 a |
| BF | 2860 | 5459 | 1.91 | 69.2 a | 4.7 a |
| β-Mannanases | |||||
| - | 2898 | 5471 | 1.90 | 69.7 a | 4.1 a |
| + | 2835 | 5398 | 1.91 | 69.3 a | 5.6 a |
| p-Value | |||||
| Growth Promoter | 0.91 | 0.67 | 0.60 | 0.14 | 0.9 |
| β-Mannanases | 0.58 | 0.52 | 0.81 | 0.18 | 0.4 |
| Interaction | 0.80 | 0.77 | 0.54 | 0.97 | - |
| SEM | 270 | 2750 | 0.10 | 1.87 | 3.4 |
| Treatment | Morphometric Index Day 49 | Intestinal IgA (ng/mL) d 35 | |
|---|---|---|---|
| Liver | Thymus | ||
| Growth Promoter | |||
| AGP | 1.89 b | 0.18 b | 136.5 b |
| BF | 2.01 a | 0.22 a | 146.0 a |
| β-Mannanases | |||
| - | 1.97 a | 0.19 b | 122.5 a |
| + | 1.93 a | 0.21 b | 160.4 b |
| p-Value | |||
| Growth Promoter | 0.002 | 0.002 | 0.47 |
| β-Mannanases | 0.26 | 0.10 | 0.01 |
| Interaction | 0.17 | 0.12 | 0.08 |
| SEM | 0.24 | 0.08 | 51.60 |
| Treatment | Day 35 | ||
|---|---|---|---|
| VL | CD | VL/CD Ratio | |
| Growth Promoter | |||
| AGP | 2187.5 a | 286.6 a | 8.0 a |
| BF | 2304.0 b | 276.7 a | 8.3 a |
| β-Mannanases | |||
| - | 2230.6 b | 291.6 a | 8.0 a |
| + | 2266.0 b | 271.3 b | 8.4 b |
| p-Value | |||
| Growth Promoter | <0.0001 | 0.13 | 0.17 |
| β-Mannanases | 0.23 | 0.002 | 0.04 |
| Interaction | 0.8 | <0.0001 | 0.0002 |
| SEM | 146.1 | 37.5 | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantoja-Don Juan, C.A.; Gómez-Verduzco, G.; Márquez-Mota, C.C.; Téllez-Isaías, G.; Kwon, Y.M.; Cortés-Cuevas, A.; Arce-Menocal, J.; Martínez-Gómez, D.; Ávila-González, E. Productive Performance and Cecum Microbiota Analysis of Broiler Chickens Supplemented with β-Mannanases and Bacteriophages—A Pilot Study. Animals 2022, 12, 169. https://doi.org/10.3390/ani12020169
Pantoja-Don Juan CA, Gómez-Verduzco G, Márquez-Mota CC, Téllez-Isaías G, Kwon YM, Cortés-Cuevas A, Arce-Menocal J, Martínez-Gómez D, Ávila-González E. Productive Performance and Cecum Microbiota Analysis of Broiler Chickens Supplemented with β-Mannanases and Bacteriophages—A Pilot Study. Animals. 2022; 12(2):169. https://doi.org/10.3390/ani12020169
Chicago/Turabian StylePantoja-Don Juan, Carlos A., Gabriela Gómez-Verduzco, Claudia C. Márquez-Mota, Guillermo Téllez-Isaías, Young M. Kwon, Arturo Cortés-Cuevas, José Arce-Menocal, Daniel Martínez-Gómez, and Ernesto Ávila-González. 2022. "Productive Performance and Cecum Microbiota Analysis of Broiler Chickens Supplemented with β-Mannanases and Bacteriophages—A Pilot Study" Animals 12, no. 2: 169. https://doi.org/10.3390/ani12020169