
Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets



1
Dipartimento di Scienze Mediche Veterinarie (DIMEVET), Università di Bologna, via Tolara di Sopra 50, 40064 Ozzano dell’Emilia (BO), Italy
2
Vetagro S.p.A., via Porro 2, 42124 Reggio Emilia, Italy
3
Vetagro Inc., 116 W. Jackson Blvd., Suite #320, Chicago, IL 60604, USA
*
Author to whom correspondence should be addressed.
Animals 2021, 11(3), 642; https://doi.org/10.3390/ani11030642
Submission received: 22 January 2021
/
Revised: 19 February 2021
/
Accepted: 23 February 2021
/
Published: 28 February 2021
(This article belongs to the Special Issue Feeding Strategies to Minimize the Use of Antimicrobials and Zinc Oxide in Pig Production)
Abstract
:Simple Summary
Zinc oxide (ZnO) supplementation at pharmacological doses in post-weaning piglets is a consolidated practice that allows efficient control of post-weaning diarrhea (PWD), a condition exacerbated by Escherichia coli F4 (K88) infections. Far from being completely elucidated, the multifactorial ZnO mechanism of action is in all likelihood exerted at the gastrointestinal level. However, increasing environmental concerns are arising from prolonged ZnO use. This article reviews the utilization of ZnO in piglets, the biological rationale behind its powerful activity, and the emerging threats that are leading towards a significant reduction in its use. Finally, a wide analysis of the strengths and weaknesses of innovative alternative strategies to manage PWD at the nutritional level is given.
Abstract
Zinc oxide (ZnO) at pharmacological doses is extensively employed in the pig industry as an effective tool to manage post-weaning diarrhea (PWD), a condition that causes huge economic losses because of its impact on the most pivotal phase of a piglet’s production cycle. In a multifactorial way, ZnO exerts a variety of positive effects along the entire gastrointestinal tract by targeting intestinal architecture, digestive secretions, antioxidant systems, and immune cells. ZnO also has a moderate antibacterial effect against Escherichia coli F4 (K88), the main causative agent of PWD. However, the environmental impact of ZnO and new emerging threats are posing serious questions to the sustainability of its extensive utilization. To work towards a future free from pharmacological ZnO, novel nutritional approaches are necessary, and many strategies have been investigated. This review article provides a comprehensive framework for ZnO utilization and its broad mode of action. Moreover, all the risks related to pharmacological ZnO levels are presented; we focus on European institutions’ decisions subsequently. The identification of a novel, complete solution against PWD should be accompanied by the adoption of holistic strategies, thereby combining good management practices to feeding approaches capable of mitigating Escherichia coli F4 (K88) infections and/or lowering ZnO utilization. Promising results can be obtained by adjusting diet composition or employing organic acids, natural identical compounds, polyphenol-rich extracts, prebiotics, and probiotics.
Keywords:
zinc oxide; post-weaning diarrhea; Escherichia coli F4; K88; ETEC; piglet; pig nutrition; weaning1. Introduction
Pigs are one of the most important livestock species, being the world’s primary meat resource for human nutrition along with poultry [1]. According to OECD-FAO 2019–2028 projections, pork will remain a pivotal nutritional source of animal protein in both developed and developing countries [2]. To face the expected continuous consumption, which will mirror the population growth and the increasing access of developing countries to newer meat varieties, attention to production cycles will be more and more necessary, to identify critical phases and recognize all the essential strategies to improve animal performance.
Concerning pigs, the most pivotal stage of their breeding is weaning, a sudden and extremely stressful moment [3]. In the modern pig industry, weaning generally occurs 21–28 days after birth [4,5]. Several aspects contribute to the consideration of weaning as a challenging state, ranging from psychosocial factors to dietary changes and morpho-functional alterations along the digestive tract [6,7,8]. The switch from a liquid to a solid diet damages intestinal villi, with an impairment in intestinal absorption and secretion of digestive enzymes, thereby leading to a reduced growth rate, starvation periods, and severe weaning anorexia [9,10]. Noteworthy effects also occur in the stomach, where the secretion of HCl is not sufficient to ensure proteolysis and protection against pathogens, which finally reach the hindgut [10,11,12]. Moreover, because of the sudden lack of passive immunity provided through maternal milk and the underdevelopment of the piglet’s immune system at weaning, acute immunological changes occur and an inflammatory status is established [10,13].
All these interconnected circumstances exert detrimental effects on the overall health status of animals, providing the ideal environment for the onset of post-weaning diarrhea (PWD), one of the most economically-relevant diseases in pig husbandry due to the costs of therapies, slower growth, and increased mortality [14]. The main causative agent is enterotoxigenic Escherichia coli (ETEC), hence the second name of PWD: post-weaning colibacillosis. The most common strain associated with PWD occurrence in piglets worldwide is Escherichia coli F4 (E. coli F4, also known as E. coli K88), which infects animals via the oral route [15]. Bacterial ancillary plasmids are responsible for the pathogenicity of these strains: they harbor not only antimicrobial resistance genes, but also sequences encoding fimbrial adhesins, essential for ETEC pathogenicity and capable of interacting with enterocytes’ receptors [16]. However, the onset of diarrhea is primarily due to the heat-labile (LT) and the two heat-stable (STa and STb) toxins that E. coli K88 can secrete to target intestinal epithelial cells. Although triggering different pathways, all the toxins disrupt tight-junctions, activate massive luminal secretions of electrolytes and water, and finally bring about diarrhea [17,18].
For a long time, zinc oxide (ZnO) has been widely used at high doses as an effective means to prevent diarrhea and control E. coli F4 infections. However, starting from June 2022, medicinal doses of ZnO will no longer be authorized in the European Union, so novel strategies to manage PWD are urgently needed. Starting from this general framework, the aim of this review is to investigate the rationale behind the large-scale utilization of ZnO in post-weaning piglets, its precise mechanism of action, and the reasons that are causing researchers, industries, and political institutions to reduce its employment at high levels. Additionally, the present work provides an overview of all the main in-feed alternatives to the use of medicinal levels of ZnO and strategies to control E. coli F4; their strengthens and limitations are described as well.
2. Zinc Oxide
Zinc (Zn) is one of the most important trace elements in animal nutrition because it ensures the activity of several enzymes involved in cellular signaling, digestion, cellular respiration, and nucleic acid metabolism [19,20]. Zinc deficiency has been correlated with growth retardation, lower appetite, worsening of feed conversion rate, and skin complications [21].
In diets, Zn can be supplemented in many different forms. The addition of Zn in the form of ZnO represents the commonly recommended way to prevent and reduce the incidence of PWD. The administration of ZnO in piglets usually lasts 14 days, starting immediately after weaning, the time during which its highest beneficial effects are observed [22,23]. In vivo studies proved that—to exert its positive action—ZnO inclusion in feeds should reach a concentration greater than 1000 ppm, with an optimum of 2500 ppm of Zn (equivalent to approximately 3100 ppm of ZnO), a dose generally defined as “pharmacological” [24]. Davin and colleagues proved that weaning causes a Zn deficiency (low plasma Zn concentration) that can be counteracted by pharmacological ZnO [25]. As demonstrated in rats, increasing levels of dietary ZnO, although ensuring an incremental absorption of Zn, do not implicate a surge in Zn retention, thereby proving that pharmacological doses of ZnO overcome physiological needs and that beneficial effects do not strictly depend on the fulfilment of nutritional Zn requirements [26].
The supplementation of ZnO not only results in better fecal scores at weaning [27,28] and a lower appearance of PWD symptoms and mortality [29], but also improves piglets’ growth performance [22,30], digestion [31], and feed intake [32]. Hahn and Baker demonstrated that the best result in daily weight gain was obtained for plasma Zn levels between 1.5 and 3 mg/L, an ideal concentration reached by supplementing exactly 3000 ppm of ZnO [33]. Despite increasing plasma Zn concentration, other Zn forms provided at the same level did not report significant enhancements in growth parameters, thereby suggesting that Zn’s efficacy is likely independent from its effective absorption, but it should exert a primary action within the gut [33].
2.1. Mechanism of Action of ZnO
The precise mechanism of action of ZnO against PWD has not yet been fully elucidated. However, many studies have hypothesized and demonstrated several beneficial effects of ZnO on different targets, as summarized in Figure 1. Rather than the fulfilment of Zn daily nutritional requirements, the main mechanism of action of ZnO seems to be related to a substantial enhancement in nutrient absorption and intestinal morphology [34]. Indeed, further studies confirmed that the supplementation of pharmacological ZnO improves performance by ensuring higher villi:crypt ratio, greater occludin protein levels, and a considerable proliferative effect on enterocytes [35,36]. The increase in tight-junction expression is a clear marker of ZnO ability to reduce intestinal permeability in a very delicate phase for piglets [37], with a marked influence on the appearance of diarrheic symptoms.
The positive effect of ZnO on intestinal mucosa can be also related to its antioxidant properties. Zn is an essential ion for the catalytic action of superoxide dismutase (SOD), whose level is increased in the intestine of ZnO-supplemented piglets [36], and for the expression of metallothioneins, which possess an antioxidant action for their metal-binding capacity [38,39] and that may play a role in cell growth and development [40]. For what concerns bacteria, ZnO causes the onset of oxidative stress in microbial cells: in aqueous solutions, high doses of ZnO generate reactive oxygen species that, thanks to their direct action on cell walls, can damage bacterial cells [39,41]. This mechanism of action could partially explain the antimicrobial activity that many Zn forms exert mostly at concentrations equivalent to the medicinal doses used in animal diets [42,43].
Despite this potential mechanism, the actual effect of ZnO against E. coli F4, the main causative agent of PWD, is moderate. Indeed, ZnO antimicrobial activity seems to be mainly addressed against Gram-positive rather than Gram-negative bacteria [42], as also confirmed by microbiota analysis, with ZnO supplemented animals reporting higher coliforms and enterococci but lower lactic acid bacteria [44]. Furthermore, pig diets added with 2500 ppm of ZnO show no differences in the number of fecal coliforms if compared to controls [45]. Considering that the low diarrhea occurrence in ZnO-supplemented piglets is recurrently associated with a persistent fecal excretion of E. coli, it is realistic that mechanisms of action different from a direct antimicrobial activity do exist for ZnO [46].
An in vitro study from Roselli et al. confirmed the limited direct antimicrobial activity of ZnO against E. coli K88, emphasizing instead the capacity of ZnO to protect cultured enterocytes from ETEC-induced damages. Indeed, it is demonstrated that ZnO inhibits bacterial adhesion to cells, thereby preventing the disruption of the intestinal tight-junctions, as proved by the higher trans-epithelial electrical resistance (TEER) values for infected cells when supplemented with ZnO [47].
Cytokines are major inflammation players that influence the tight-junction functioning of epithelial cells [48]. During in vitro infections of intestinal epithelial cells with E. coli F4, ZnO significantly reduced the expression of several pro-inflammatory cytokines and improved the transcription of the anti-inflammatory cytokine TGF-β [47], together with a significant downregulation of genes related to the innate immune response [49]. The same modulatory effect on intestinal inflammation was demonstrated by many in vivo studies, which reported lower IFN-γ, TNF-α, and IL-6 in ZnO-supplemented piglets [35,36,50].
Not only is ZnO capable to modulate cytokines expression, but also it is an essential trace element with a key action on the entire immune system, for both its development and its correct function [50]. Deficiency of Zn brings to a significant decrease of macrophage capacity to perform phagocytosis [51] and to kill parasites [52]. Moreover, Zn seems fundamental to enhance the killing activity of natural-killer lymphocytes [53]. A disturbed Zn homeostasis has been correlated with an increased risk for infections, due to an impaired balance of T helper cells and to an increased apoptosis of immature T cell precursors [54], therefore proving the key role of Zn for T helper cell differentiation [55]. In weaned piglets, pharmacological doses of ZnO showed the ability to increase the number of regulatory T cells, which are pivotal for modulating the immune response and maintaining homeostasis [56]. A study from Ou and colleagues highlighted the role of pharmacological doses of ZnO in reducing the expression of stem cell factor in the small intestine of weanling pigs, an effect that could not only repress maturation and proliferation of intestinal mucosal mast cells, but also decrease their release of histamine, a key molecule for the pathogenesis of diarrhea and one of the main players for the onset of an inflammatory status [57].
Researchers proved that pharmacological supplementation of ZnO also participates to the stimulation of ghrelin secretion at the stomach level of young pigs. Ghrelin action promotes the release of insulin-like growth factor 1 (IGF-1) and cholecystokinin (CCK) by the liver and the GI tract, respectively, thereby enhancing muscle protein synthesis, cell proliferation, and feed intake [58,59]. Moreover, Hedemann and colleagues verified that high dietary levels of ZnO in weaned pigs increase the activity of several pancreatic digestive enzymes such as carboxypeptidases, trypsin, and chymotrypsin [60], guaranteeing a higher nutrient digestibility. All these multiple effects can together contribute to the systematic performance improvement observed in weaning piglets fed with medicinal ZnO doses.
2.2. Risks Related to Pharmacological Levels of ZnO
The large and prolonged employment of ZnO at pharmacological levels in pig farming has raised several concerns, as depicted in Figure 2. Too high or too much protracted ZnO supplementations lead to the loss of ZnO benefits with the potential onset of toxic effects [22,61,62], because of the excessive accumulation of Zn in animal tissues such as kidney, liver, and pancreas [63,64], which can experience Zn overload [62].
The main issue concerning ZnO is related to the considerable risks for the environment, deriving from the application of Zn-rich manure to land: manure coming from facilities employing pharmacological doses of ZnO is definitely abundant in Zn, considering that animals excrete all the excess of Zn that exceeds physiological requirements [22,65,66]. Due to the non-volatile and non-degradable physicochemical properties of Zn, the long-term continuous spread of manure on crops can progressively increase its concentration into soils and groundwaters, finally reaching dangerous levels for plant and animal life [67,68]. Bak et al. demonstrated that, over the period 1986–2014, the application of Zn-rich pig slurry led to a greater soil Zn concentration of 2–5%, with an average increase of 24% over the last sampling period (1998–2014), concluding that there might be significant risks for aquatic species because of Zn leaching from fertilized fields to groundwaters [69]. Although risk mitigation measures are more and more implemented, such as manure dilution and ensuring distance to surface waters, the European Medicines Agency (EMA) 2017 report about veterinary medicinal products containing ZnO concluded that these precautions only represent a delay for the unavoidable Zn accumulation in the environment [23].
Several studies proved that ZnO pharmacological supplementation in piglets might also contribute to the acquisition and spread of antibiotic resistance genes [70]. Slifierz et al. showed that therapeutic doses of ZnO can increase the persistence and prevalence of methicillin-resistant Staphylococcus aureus in weaning piglets, probably due to the colocalization of Zn and methicillin resistance genes [71]. The diffusion of resistance genes amongst E. coli inside the intestine of piglets weaned with high ZnO levels was also observed [72], together with a considerable increase of multi-resistant E. coli strains in feces, digesta, and colonic mucosa samples [73,74]. Piglets supplemented with high ZnO doses show a tendency into the selection of more heavy metal tolerant E. coli isolates such as strains harboring the czrC gene [75,76], thereby jeopardizing any potential antimicrobial activity of Zn.
The impact of ZnO on the intestinal microbiota is controversial. While some data only suggest minor or transient modifications to hindgut bacterial compositions [77,78,79], other studies reveal remarkable effects on porcine microbial populations [80]. It was demonstrated that pharmacological doses of ZnO can significantly increase the count of Enterobacteriaceae in piglets [81]. Moreover, ZnO can influence microbiota composition, by reducing Lactobacillus species and increasing Clostridiales and Enterobacteriales diversity [82,83]. The modulatory activity of ZnO on commensal microbiota probably resembles the activity of growth-promoting antibiotics: the suppression of Gram-positive species, without affecting Gram-negative strains, brings to lower bacterial activity and ATP concentration in the gut of piglets, which makes more energy available for the host [44], at the cost of losing, though, beneficial commensal bacteria.
Given all the risks linked to Zn, the European Food Safety Agency (EFSA) is constantly monitoring pollution levels and proposed a significant reduction in the use of Zn in feeds [84]. Moreover, to reduce the environmental and antimicrobial-resistance threats derived from the accumulation of Zn from medicated feeds, the EMA concluded that the benefit-risk balance for the utilization of medicinal ZnO containing products in pig breeding had a negative outcome. This resolution was dictated by the severe hazards for the environment, that took priority over the prevention of post-weaning diarrhea in piglets. The decision, issued on 26th June 2017, imposes a complete elimination of ZnO at medicinal levels in piglet feed, enforcing a five-year timeframe to phase these products out of the market [23]. No laws or restrictions are actually in force in other countries such as the United States, but breeders are always welcoming any new alternative strategy to manage PWD and E. coli F4 infections.
Since the European Commission has supported the EMA decision towards a future free from pharmacological levels of ZnO by 2022 [85], an urgent need for ZnO-reducing alternatives is arising. Indeed, following the EC regulation 1334/2003 and 2016/1095, ZnO will be only utilizable as a feed additive with the legal limit of 150 ppm of total Zn in complete feed [86,87]. This dose unlikely exerts the same effects of pharmacological levels of ZnO, so non-traditional approaches against PWD are urgently needed.
2.3. Why ZnO?
As an ion, Zn is contained inside a great number of compounds, some of which are of primary interest to fulfill Zn dietary requirements in animal nutrition. For this reason, many Zn-containing molecules have been tested as potential substitutes for pharmacological doses of ZnO in piglets, with the aim to supplement them at concentrations lower than the medicinal ones.
Together with ZnO, the most used source of Zn in animal husbandry is zinc sulphate. The reason why this alternative inorganic form is used relies on the higher bioavailability of Zn when bound to the sulphate group [88,89,90]. The investigation of the potential in vitro growth-inhibiting activity of Zn sulphate against a panel of intestinal pathogens (i.e., Salmonella spp., Enterobacter spp., Escherichia coli, Vibrio cholerae) showed that it exerts an antimicrobial action between 1.2 and 1.8 mg/mL, thereby suggesting a possible antidiarrheal activity not only linked to physiological effects, but also to a mild antibacterial action [43]. However, to the best of our knowledge, no comparative studies between pharmacological levels of ZnO and Zn sulphate have been published so far; thus, no reports support Zn sulphate as an alternative to ZnO.
Zinc chloride is another Zn inorganic compound employed in animal nutrition. Although being less studied, its beneficial effects are probably linked to the inhibition of certain bacterial populations and the improvement of intestinal health [91]. One of the most diffused forms of Zn chloride is tetrabasic zinc chloride (TBZC), an additive manufactured through reactive crystallization processes, with the final production of a water-insoluble colorless salt. This form is preferable because of the higher purity, the excellent palatability, the reduced reactivity with other feed nutrients, and the better bioavailability compared to ZnO [92,93]. In piglets, supplementation of 1000–1500 mg/kg of Zn chloride in the TBZC form can enhance fecal score and growth parameters as equally as pharmacological ZnO [94,95,96], however without being able to drastically reduce Zn doses below the limits imposed by the European Commission.
Effective Zn sources can also be organic Zn forms [97]. At pharmacological levels, Zn from Zn-methionine or Zn-lysine can increase plasma Zn concentration much more than ZnO or other inorganic Zn forms [33,98,99]. Bouwhuis and colleagues verified that the inclusion of 500 ppm of Zn-methionine in post-weaning piglets did not result in improved fecal scores and intestinal architecture at the same extent of 3300 ppm of ZnO [100]. Other research groups proved that 250–500 ppm of various organic Zn forms could increase to a certain extent the growth parameters of weaning piglets, however without reaching the performance of pharmacological levels of ZnO [32,101,102]. These results further prove that the mechanism of action of ZnO against PWD is not likely related to Zn absorption, so any utilization of organic Zn forms to improve Zn bioavailability would not exert desirable positive effects on PWD symptoms.
As a whole, all the previously presented alternatives do support the idea that the activity of Zn in preventing PWD or controlling E. coli F4 infections is not merely related to the supply of Zn, but rather it is strictly dependent to its inclusion in the form of ZnO. Considering that other Zn forms do not represent feasible alternative ways to reduce pharmacological doses of ZnO under the limit established by the European authorities, several approaches were explored to maintain the ZnO molecule by minimizing its levels with the aim to decrease its environmental footprint. The rationale behind the reduction of ZnO doses lies in its solubility at acidic pH [103]: in the low pH gastric environment, the majority of ZnO is dissolved as Zn2+ and eventually absorbed, with a limited amount flowing into the small intestine, where all the beneficial effects of ZnO are exerted [104]. Moreover, it is likely that Zn2+ dissolved ions, once entered the intestine, could react with NaOH forming Zn(OH)X, an insoluble and inactive Zn form. If ZnO is industrially processed, such as miniaturized, incorporated in minerals, or microencapsulated, a higher dose can reach the small intestine without being transformed and lost along the gastric tract, thereby allowing a significant reduction of ZnO concentration in animal feed.
Thanks to the great strides of nanotechnologies, an attractive form of ZnO is represented by ZnO nanoparticles (nZnO). Nanoparticles of ZnO possess better chemical stability and reactivity because of their smaller size, that brings to an increased number of particles and surface area per unit mass [105,106]. Moreover, nZnO show an antimicrobial activity against pathogenic bacteria such as E. coli, Salmonella, Staphylococcus, and Listeria [107]. Their mechanism of action is related to nZnO enhanced capacity to permeate into bacterial cell membranes, causing oxidative damages and lipid peroxidation [108]. In 2019, a study by Pei et al. proved that dietary supplementation of 450 ppm of ZnO nanoparticles in piglets diets could exert similar effects to the common ZnO pharmacological dose of 3000 ppm, maintaining improvement in growth and intestinal morphology parameters, with a significant reduction of fecal Zn excretion [109]. Additional findings proved other positive effects such as fecal E. coli count reduction and intestinal tight-junctions expression improvement [110]. However, other research groups did not confirm the enhanced performance parameters, or reported contrasting results on diarrhea rate when pharmacological ZnO was replaced with 500–1200 ppm of nZnO [106,110,111]. Furthermore, all the available studies report the use of nZnO at doses far from the strict limits imposed by European authorities, thereby making it difficult to suggest nZnO as an effective low-dose alternative to medicinal ZnO levels.
Several carrier minerals have been employed to control the release of ZnO along the gastrointestinal tract. The supplementation of 600–900 ppm of zeolite-supported ZnO proved to be as efficacious as pharmacological doses of ZnO in enhancing growth performance, maintaining intestinal barrier function, and alleviating diarrheic symptoms of post-weaned piglets [28]. The same beneficial effects, together with a significant improvement of tight-junction protein expression, was reported for 500 ppm of ZnO incorporated inside diosmectite [27]. Moreover, piglets fed with 500 ppm of montmorillonite-ZnO hybrid reported—as equally as pharmacological doses of ZnO—a noteworthy enhancement in growth performance, lower diarrhea, improvement of intestinal morphology, and higher activity of pancreatic and intestinal digestive enzymes [31,112].
One of the most efficient ways to diminish ZnO concentration originates from the microencapsulation technology, which was shown to allow for a reduced amount of ZnO requested to maintain health and performance of piglets after weaning [113]. As firstly demonstrated by Grilli et al., 150 and 400 ppm of microencapsulated ZnO were as effective as the free pharmacological dose of 3000 ppm, with beneficial effects on intestinal morphology, cell proliferation, gut inflammatory cytokines and tight-junction protein expression, and production parameters [35]. Subsequent studies proved similar effects: coated ZnO doses ranging from 380 to 500 ppm exerted positive actions on diarrhea incidence, intestinal development, mucosal immune system, and nutrient digestibility [104,114]. Moreover, low doses of coated ZnO could enhance intestinal antioxidant capacity and reduce intestinal cell apoptotic index [115], together with a significant decrease in fecal Zn [116] and environmental pollution.
Although the above presented approaches look promising in reducing the ZnO bioactive concentration and in controlling its environmental impact, all the employed doses are far from complying with the requirements imposed by the European Commission of 150 ppm of total Zn in complete feed from 2022. While pursuing a future without pharmacological ZnO, novel cutting-edge approaches are currently being investigated as effective pioneering ZnO substitutes to control PWD and E. coli F4 infections.
3. Alternatives to ZnO
Many non-feed strategies have been investigated to control PWD and E. coli F4 infections. Poor environmental conditions are considered contributing factors to the establishment of PWD [14]. Sanitary aspects deeply affect the overall health status of piglets: a deterioration in husbandry hygiene levels can trigger a background inflammation status that may intensify other stressors [117], finally leading to depressed growth performance and diarrhea [118]. Moreover, considering that some animals are not susceptible to ETEC infections because of genetic reasons [119,120], the breeding selection of animals resistant to E. coli F4 was considered as a solution to erase PWD, but several difficulties occurred [121,122].
Even if non-feed approaches all represent noteworthy factors to PWD onset and environmental enterotoxigenic E. coli persistence [123], their analysis is beyond the scope of this review. Since the mechanism of action of ZnO is primarily related to a wide-spectrum action on the gastrointestinal tract of weaning piglets, only feed-related strategies different from pharmacological ZnO levels will be discussed. For each proposed alternative approach, only studies including a clear comparison with high ZnO doses and/or investigating novel methods to control E. coli F4—the main etiological agent of PWD—were considered.
A prospect of all the different strategies to PWD and E. coli F4 is reported in Table 1, which summarizes all the advantages and limitations of each analyzed approach.
3.1. Adjusting Diet Composition: Protein and Fiber
At weaning, the high buffering capacity of the solid feed and the low HCl production of the piglet stomach lead to low protein digestion [124]. For this reason, a large amount of undigested protein flows into the large intestine, where it is easily fermented by the host microbiota causing an overgrowth of proteolytic bacteria—such as pathogenic E. coli—and their toxic fermentation by-products [141,142]. Moreover, undigested proteins can facilitate the onset of osmotic diarrhea. An effective way to control the protein amount arriving into the hindgut is the reduction of the dietary crude protein (CP) level simultaneously to the supplementation of essential amino acids (AA) [143]. Low CP and AA supplemented diets proved useful to enhance fecal consistency and decrease PWD incidence even in E. coli F4 experimentally challenged piglets without the aid of pharmacological ZnO up to 14 days after weaning [125,144]. Researchers also showed that weaned pigs fed low CP diets (16% instead of 20% CP) can reduce up to 30% the daily treatments for diarrheic symptoms in the 6–15 kg phase [145]. Furthermore, a low CP diet (15.5% instead of 20% CP) helps in lowering IL-1, TNF-α, IL-2, IL-6 and increasing IL-10 and IL-13 concentrations in the colonic mucosa of weanling piglets, probably due to a reduction of the inflammation elicited by the ingestion of antinutritional factors and feed antigens usually occurring with common protein sources such as soybean meal [146].
However, pig performance typically declines when dietary CP percent is heavily reduced, so lower productivity should be taken into account when opting for this alternative strategy to phase out medicinal levels of ZnO [144]. Even if later additions of dietary high CP can counterbalance for the initial lower performance [145], some compensatory strategies are currently under investigation, such as management precautions and supplementation of both essential and branched-chain AA [146,147,148]. Overall, these insights show that dietary CP plays a role in the PWD onset and that its concentration should be carefully determined to avoid an easier E. coli F4 gut colonization at weaning.
If acting with CP can be risky for the maintenance of piglets performance, several authors suggested the possibility to use insoluble fibers to control PWD symptoms. Fibers improve intestinal health by promoting the establishment of a healthy bacterial community in the hindgut [149]. Furthermore, fiber improves intestinal morphology [150] and prevents diarrhea formation by decreasing E. coli counts in feces [128].
Fibers such as insoluble non-starch polysaccharides (iNSP) have been employed with success in several studies. Fernandes et al. demonstrated that the combination of fibers with low doses of ZnO (500–800 ppm) allowed the same growth performance of pharmacological ZnO in piglets experimentally challenged with E. coli F4 [127]. Furthermore, researchers proved how supplementation of fibers could help in control PWD by reducing E. coli F4 faecal counts, and by improving short-chain fatty acids (SCFA) levels in the digesta [126,151]. The mechanism of action of iNSP is related to their ability to diminish the adhesion of pathogens such as E. coli F4 to intestinal epithelial cells, as demonstrated by in vitro studies on IPEC-J2 cells [152,153], and decrease the retention time of digesta [116]. The two mechanisms likely play a synergistic role because an increased passage rate can lower the attachment of pathogens to the intestinal surface.
3.2. Organic Acids
Organic acids (OA) are one of the most employed groups of feed additives in pig nutrition. In their classic role as acidifiers, OA can reduce gastric pH of weanling piglets counteracting the buffering capacity of the diet, ensuring a proper digestion of dietary proteins. Moreover, gastric acidification is also valuable as an antimicrobial barrier [12,154]. In addition, OA are also widely used for their direct antimicrobial activity and their ability to improve the overall health status of pigs gastrointestinal tract [129,155]. Several studies proved the beneficial effects of lactic, propionic, citric, formic, and caprylic acids supplementation in post-weaning piglets, which ensure a reduction in intestinal harmful coliform bacteria [156], a modulation of the immune response [157,158], and an improvement in diet digestibility [159], laying the ground for better performance and reduced post-weaning diarrhea incidence [130]. Moreover, it was demonstrated that butyrate can improve growth parameters in post-weaning piglets, also thanks to its capacity to modulate intestinal inflammation and integrity [160,161,162]. An in vitro study by our research group highlighted how selected OA can reduce the expression of many E. coli K88 virulence-related genes, ranging from toxins secretion to motility and quorum sensing [163]. A few studies in piglets experimentally challenged with E. coli F4 demonstrated that mixtures of short chain fatty acids (SCFA) can decrease diarrhea severity, improve growth performance, and reduce fecal E. coli counts [130,164]. Moreover, it was also demonstrated that a complex of SCFA with capric acid (a medium chain fatty acid) can significantly alleviate the inflammatory response in piglets challenged with E. coli F4 [165], remarkably reducing TNF-α and IFN-γ plasma levels. These findings ascribe OA as powerful alternatives to manage PWD, even if further studies are required to fully understand the effectiveness of OA to completely replace pharmacological doses of Zn.
3.3. Essential Oils and Nature Identical Compounds
Following the complete ban of antibiotics as growth promoters in the European Union, essential oils (EO) have immediately gained attention as potential alternatives in animal diets. Their usefulness is related to their strong antimicrobial, antioxidant, and anti-inflammatory activity [131,132,133]. The main etiological agent of PWD, E. coli F4, proved to be susceptible to several EO, including cinnamon, clove, and thyme oils [166,167,168]. Many in vivo studies confirmed positive effects deriving from the supplementation of EO blends in weanling piglets, with a reduction in coliform proliferation [156], a lower fecal score and diarrhea appearance [169], an improvement in intestinal morphology and feed digestibility [170], and an increase in total antioxidant activity [171]. However, studies providing EO supplementation sometimes report contrasting results or inconclusive outcomes [172,173], probably due to different EO inclusions and compositions, which could be significantly influenced by factors such as soil composition, plant species, harvesting season, extraction method, and stability [168,174].
For these reasons, nature identical compounds (NIC)—pure chemicals reflecting active principles of EO—are more and more employed to overcome variabilities and potentiate EO effects [133]. Thymol, carvacrol, and eugenol are three of the most effective NIC against E. coli F4, as demonstrated by their in vitro antimicrobial activity [153,159,168]. Moreover, we previously demonstrated that these three monoterpenes significantly downregulate mRNA levels of E. coli K88 genes involved in toxins production, bacterial adhesion to enterocytes, motility, and quorum sensing [163]. As regards in vivo trials, feeding a blend of thymol, vanillin, and OA for two weeks in post-weaning piglets resulted in a modulation of several inflammatory markers and an enhancement in growth parameters, while in vitro experiments showed a reduction in tight-junction permeability [175]. Moreover, the same blend significantly lowered the E. coli shedding and tended to contribute to the reduction of Zn excretion in feces of weanling pigs supplemented with low doses of microencapsulated ZnO [113]. Therefore, these data suggest NIC as promising candidates to serve as effective ZnO alternatives to manage PWD and E. coli F4.
3.4. Polyphenol-Rich Extracts
Plant and fruit extracts—referred as natural extracts (NE)—are widely used in traditional medicine to treat various diseases, including gastrointestinal conditions such as diarrhea. Several studies proved the effectiveness of watery or alcoholic NE extracts against enteric pathogens, with polyphenols being the class of molecules with the highest activity [176,177]. Due to their accumulation into the hindgut, polyphenols such as resveratrol, quercetin, and phenolic acids have a broad antimicrobial activity against microorganisms such as S. aureus, Campylobacter, Salmonella, and E. coli [135,178].
The antidiarrhoeic activity of polyphenols is probably related to their capacity to bind proteins. Historically, this property has labelled them as antinutritional factors, but research is now exploring their usefulness against pathogenic E. coli [136]. Several NE, together with their polyphenolic content, were shown to be active against E. coli F4 pathogenicity at different levels, ranging from a moderate antimicrobial activity [179] to the inhibition of LT toxin and the repression of enterocytic ionic channels [134,180,181]. Thus, targeting virulence instead of bacterial growth and survival represents a novel and pioneering approach to the pathogenicity of microorganisms, with the key advantage to preserve host microflora from unwanted harmful modifications and reduce selective pressure against the development of bacterial resistance [182,183].
In addition, polyphenols possess the capacity to bind metals such as iron, aluminum, and copper [184]. This ion chelating capacity, although not desirable in deficient individuals, can be considered one of the reasons why polyphenols exert antidiarrheal properties [134], even if no specific actions have been confirmed with E. coli F4 so far.
Polyphenolic-rich extracts and their bioactive components are primarily recognized for their great antioxidant activity thanks to their hydroxyl groups, which can be oxidized by many molecules, including reactive oxygen species (ROS) [185]. The increased inflammation status of piglets caused by weaning leads to an increase in blood flow which produces a surge in the gastrointestinal ROS, a condition which has been coupled with an expansion of the E. coli gut population [186,187]. Moreover, as a response to inflammation, the production of NO provides a source of nitrogen that can be transformed into nitrate and used as a growth fuel for E. coli strains [188,189]. The detoxification of these reactive species by polyphenols could be an additional mechanism of action for their antimicrobial, antidiarrhoeic, and intestinal villi protective action against oxidative stress and morphological damages at weaning [190].
A study from Liu and colleagues compared the effect of hydrolysable tannins (HT) to pharmacological doses of ZnO in post-weaning piglets. The utilization of HT could reduce diarrhea incidence at the same extent of 2000 ppm of ZnO, and showed a synergistic tendency in the group supplemented with both HT and ZnO [191]. The same synergy was proved between HT and ZnO for diet digestibility, an effect probably exerted by the increased activity of enzymes such as trypsin, lipase, and α-amylase, the improvement of intestinal villi structure, and the enhanced antioxidative enzymes activity of the small intestine [191]. To the best of our knowledge, no other studies comparing polyphenol-rich extracts to pharmacological doses of ZnO or exploring their effects in piglets challenged with E. coli F4 are available, so further investigations will be required to elucidate their effectiveness as alternative strategies to manage PWD.
3.5. Prebiotics, Probiotics, and Symbiotics
Prebiotics are nondigestible feed ingredients that positively affect the host by stimulating the growth and/or the activity of a limited number of bacteria of the gastrointestinal microbiota [192]. A few studies have elucidated the advantages of supplementing different prebiotics to prevent damages exerted by the PWD main causative agent, E. coli F4. An in vitro study conducted on Caco-2 cells proved the ability of several milk-derived oligosaccharides fractions to avoid the adhesion of pathogenic E. coli to the cellular monolayer [193]. Selected galacto- and mannan-oligosaccharides reduce the in vitro adherence of E. coli F4 to intestinal mucins or porcine IPEC-J2 cells [194,195]. Moreover, orally-administered β-glucans decrease E. coli F4 colonization and diarrheic symptoms in post-weaning piglets [196]. The supplementation of an alginate-derived oligosaccharide could alleviate damages exerted by E. coli F4 infection by decreasing apoptosis and stimulating proliferation of enterocytes in challenged weanling piglets [197].
Probiotics are described as viable microorganisms that, if administered in sufficient amount, reach the intestine and exert positive effects for the host via direct inhibition of pathogens and/or stimulation of host defense systems [198]. The most common probiotics are lactic acid bacteria and yeasts. Roselli et al. demonstrated that in vitro addition of probiotic strains such as Lactobacillus sobrius, Lactobacillus rhamnosus, and Bifidobacterium animalis prevent E. coli F4 adhesion and tight-junction damages to intestinal epithelial cells, with a significant modulation of inflammatory cytokines [199,200]. Post-weaning piglets experimentally infected with E. coli F4 showed a significant reduction of diarrhea incidence when supplemented with Lactobacillus rhamnosus thanks to the modulation of intestinal microflora and inflammatory cytokines [201]. The addition of Bacillus species to weanling piglet diets after infection with E. coli F4 proved useful to reduce fecal score and increase the villi mitotic index through the enrichment of genes related to immune response [202]. A study by Nordeste and colleagues highlighted that probiotics beneficial effects are largely obtained by their ability to produce bioactive molecules: piglets challenged with E. coli F4 and supplemented with bioactive media conditioned with Lactobacillus acidophilus exhibited less PWD symptoms and better performance parameters because of an improvement in gut health and a lower E. coli F4 colonization [203]. Finally, feeding Saccharomyces cerevisiae in E. coli F4 infected piglets can lead to a reduction in diarrhea scores and duration, with a positive influence on animal growth capacity after weaning [204].
Some studies also explored the beneficial influences of combining both probiotics and prebiotics to improve the effective implantation and survival of health-promoting bacteria in the gastrointestinal tract [192]. The inclusion of a combination of 14% raw potato starch and E. coli probiotics could increase growth performance and reduce diarrhea incidence in piglets challenged with E. coli F4 [205]. A combination of Lactobacillus plantarum and lactulose proved effective to improve daily gain and intestinal morphology in post-weaning piglets orally challenged with E. coli F4, possibly through an increase in colonic trophic SCFA such as butyric acid [206]. In vitro administration of Saccharomyces cerevisiae and β-galactomannan oligosaccharides on porcine intestinal epithelial cells co-cultured with dendritic cells reduced the expression of ETEC-induced pro-inflammatory cytokines [207]. Moreover, the addition of Lactobacillus rhamnosus and fructooligosaccharides to E. coli F4 cultures could inactivate ETEC pathogenicity by reducing LT toxin expression [208].
Prebiotics and probiotics might represent an interesting approach to manage PWD and E. coli F4, thanks to their ability to secrete antibacterial molecules, inhibit pathogen virulence, compete for adherence sites, and modulate the animal immune system. However, the lack of consensus about the composition of the healthy piglet microbiota and the considerable differences between the attempted approaches still require further investigations [187].
3.6. Others
Researchers are always working on finding pioneering strategies against many diseases, including PWD and its etiological agents. The outcomes of several in vitro and in vivo trials provide promising results that could suggest other supplementary approaches to replace ZnO or against ETEC strains responsible of PWD. However, some of the alternatives which are being investigated remain at a research level, and have still not found a sustainable high-scale application and/or the approval of regulatory authorities.
3.6.1. Antimicrobial Peptides
All the organisms naturally produce antimicrobial peptides (AMP), a heterogeneous class of small molecules that play a role in innate immunity providing immediate defense against bacterial, fungal, viral, and parasitic infections [138]. Moreover, it is suggested that they exert a low bacterial resistance acquisition [209]. In weanling piglets, other than directly targeting bacteria through many mechanisms of action, AMP can also modulate the immune system, improve growth performance, ensure an optimal intestinal structure, and suppress microbiota microorganisms such as harmful E. coli and Salmonella strains [137]. Even if no studies compared AMP to pharmacological doses of ZnO, a few research groups investigated the effects of two AMP in piglets challenged with ETEC. Wu et al. demonstrated that the supplementation of cecropin AD can improve intestinal morphology, reduce the count of E. coli, and increase the Lactobacillus count in the digesta, with a restoration of piglet growth parameters at the same extent of the group supplemented with colistin [210]. Some commensal E. coli strain can produce colicin, an AMP effective against pathogenic E. coli; Cutler and colleagues proved that its addition in diets of weanling piglet challenged with E. coli decreases PWD incidence and reduces the expression of pro-inflammatory cytokines [211]. Finally, it has been demonstrated that the in vitro overexpression of the BPI antimicrobial peptide can help in reducing ETEC adhesion to target cells and modulating the expression of several inflammatory cytokines [212].
3.6.2. Bacteriophages
Bacteriophages are a large group of viruses that infects and replicates within bacterial cells. With the global concern generated by antimicrobial resistance, bacteriophages represent an interesting alternative against bacterial pathogens [213]. The utilization of bacteriophages active against E. coli F4 resulted in lower onset of diarrheic symptoms and reduced ETEC count in feces [214]. Moreover, bacteriophage supplementation ensured better growth parameters, fecal consistency score, villi:crypt ratio, and goblet cell density in E. coli F4 experimentally challenged weanling piglets [215]. To overcome their narrow spectrum of activity, some studies have explored with success the administration of a combination of more phages, with a significant improvement of many parameters, without affecting the total E. coli population of the gut microflora [216]. However, bacteriophages can elicit an immune response in the animal body, together with the possibility to induce bacterial resistance [217]. Finally, it has also been demonstrated that bacteriophages can carry antibiotic resistance genes, thereby representing dangerous horizontal gene transfer vectors [218].
3.6.3. Egg Yolk Antibodies and Spray-Dried Plasma
Laying hens have attracted attention because of their capacity, after immunization, to concentrate specific antibodies (IgY) into the egg yolk, from which the IgY can be isolated to treat enteric pathogens [139]. In piglets experimentally challenged with E. coli F4, egg yolk antibodies significantly reduced the incidence of diarrhea, ensuring a positive weight gain and maximum survival [219]. The proposed mechanism of action against E. coli F4 is probably related to their ability to inhibit bacterial adhesion to mucus and intestinal cells [220,221]. However, no recent studies employing egg yolk antibodies are currently available, probably because of huge variabilities in IgY research studies due to their stability issues in the gastrointestinal tract and their cost of production [220,222].
Another immunoglobulin-related approach is represented by spray-dried plasma (SDP), a biologically active product obtained by industrial fractionation of animal blood. Studies have proved its effectiveness to control gastrointestinal infections, probably thanks to the direct activity of its immunoglobulin-rich fraction against pathogens or its supporting function for the immune system [140]. In post-weaning piglets orally challenged with E. coli F4, SDP supplementation ensured good levels of feed intake and weight gain [223], improved scours score and E. coli shedding in feces [224], provided better villi height:crypt depth ratio in the small intestine [225], and reduced the expression of inflammatory cytokines such as IL-8 and TNF-α [226]. However, the high production costs and the risks related to the potential presence of pathogens in SDP are endangering its effective implementation as a ZnO alternative strategy to prevent PWD [227].
4. Conclusions
Despite recent advances, the worldwide pig industry is still suffering from PWD and its main etiological agent, E. coli F4. At weaning, ZnO represents one of the major strategies for managing detrimental symptoms and bacterial infections, but severe threats arise from ZnO environmental pollution and its contribution to the spread of bacterial resistance. For a future free from medicinal ZnO doses, novel alternatives are necessary against pig related pathogens.
Numerous approaches have been attempted to treat PWD, but the proposed alternatives all show a significant gap with respect to ZnO’s efficacy. Moreover, the majority of all the analyzed studies lack a meticulous comparison between beneficial effects of pharmacological levels of ZnO and any of the newly-proposed methods, so it is often difficult to understand the real therapeutic or preventive actions of novel tools against PWD. The complexity in finding a unique effective replacement to ZnO probably lies in the supposedly multi-factorial and multi-target action of ZnO itself. Finding innovative alternatives to pharmacological ZnO for weaning pigs will probably require the combination of several of the strategies and tools presented across this review, to complement their different modes of action and targets synergistically.
In conclusion, the establishment of a novel approach against PWD will demand broad-spectrum interventions. Not only is it of crucial importance to find the optimal method to control growth, pathogenicity, and the virulence of PWD’s main etiological agent—E. coli F4—across all the pathogenic stages, but we must also figure out the best procedures to manage the weaning phase for piglets with pioneering environmentally-friendly molecules other than ZnO, along with the association of animal welfare with biosafety and nutritional measures, thereby adopting a holistic strategy.
Author Contributions
Conceptualization, E.G., B.T., and A.B.; data collection, A.B.; writing—original draft preparation, A.B.; writing—review and editing, B.T. and E.G.; supervision, E.G. and A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Images were created with BioRender.com.
Conflicts of Interest
Andrea Piva serves as a professor at the University of Bologna and is a member of the board of directors of Vetagro S.p.A. (Reggio Emilia, Italy). Ester Grilli serves as an advisor of Vetagro S.p.A.
References
- FAO’s Animal Production and Health Division Meat & Meat Products. Available online: http://www.fao.org/ag/againfo/themes/en/meat/backgr_sources.html (accessed on 22 December 2020).
- FAO; OECD Meat. OECD-FAO Agricultural Outlook 2020–2028; OECD Publishing: Paris, France, 2020; pp. 162–173. ISBN 9789264582958. [Google Scholar]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 2–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, P.; Recén, B. When to wean—Observations from free-ranging domestic pigs. Appl. Anim. Behav. Sci. 1989, 23, 49–60. [Google Scholar] [CrossRef]
- Bøe, K. The process of weaning in pigs: When the sow decides. Appl. Anim. Behav. Sci. 1991, 30, 47–59. [Google Scholar] [CrossRef]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Colson, V.; Martin, E.; Orgeur, P.; Prunier, A. Influence of housing and social changes on growth, behaviour and cortisol in piglets at weaning. Physiol. Behav. 2012, 107, 59–64. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional management of gut health in pigs around weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning Anorexia May Contribute to Local Inflammation in the Piglet Small Intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Castillo, M.; Martín-Orúe, S.M.; Nofrarías, M.; Manzanilla, E.G.; Gasa, J. Changes in caecal microbiota and mucosal morphology of weaned pigs. Vet. Microbiol. 2007, 124, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. (Berl.) 2013, 97, 207–237. [Google Scholar] [CrossRef] [PubMed]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning Is Associated with an Upregulation of Expression of Inflammatory Cytokines in the Intestine of Piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roussel, C.; Cordonnier, C.; Livrelli, V.; Van de Wiele, T.; Blanquet-Diot, S. Enterotoxigenic and Enterohemorrhagic Escherichia coli: Survival and Modulation of Virulence in the Human Gastrointestinal Tract. In Escherichia coli—Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; Amidou, S., Ed.; InTech: London, UK, 2017; pp. 3–24. [Google Scholar]
- Dubreuil, J.D. Enterotoxigenic Escherichia coli targeting intestinal epithelial tight junctions: An effective way to alter the barrier integrity. Microb. Pathog. 2017, 113, 129–134. [Google Scholar] [CrossRef]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Hill, G.M.; Shannon, M.C. Copper and Zinc Nutritional Issues for Agricultural Animal Production. Biol. Trace Elem. Res. 2019, 188, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Sloup, V.; Jankovská, I.; Nechybová, S.; Peřinková, P.; Langrová, I. Zinc in the Animal Organism: A Review. Sci. Agric. Bohem. 2017, 48, 13–21. [Google Scholar] [CrossRef] [Green Version]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, R.G. Animal Nutrition, 7th ed.; Benjamin Cummings: San Francisco, CA, USA, 2011; ISBN 9781408204238. [Google Scholar]
- Poulsen, H.D. Zinc oxide for weanling piglets. Acta Agric. Scand. A Anim. Sci. 1995, 45, 159–167. [Google Scholar] [CrossRef]
- EMA; CVMP. EMA—Zinc Oxide—Annex II—Scientific Conclusions and Grounds for the Refusal of the Marketing Authorisation and for Withdrawal of the Existing Marketing Authorisations; EMEA: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Hill, G.M.; Mahan, D.C.; Carter, S.D.; Cromwell, G.L.; Ewan, R.C.; Harrold, R.L.; Lewis, A.J.; Miller, P.S.; Shurson, G.C.; Veum, T.L.; et al. Effect of pharmacological concentrations of zinc oxide with or without the inclusion of an antibacterial agent on nursery pig performance. J. Anim. Sci. 2001, 79, 934–941. [Google Scholar] [CrossRef]
- Davin, R.; Manzanilla, E.G.; Klasing, K.C.; Pérez, J.F. Effect of weaning and in-feed high doses of zinc oxide on zinc levels in different body compartments of piglets. J. Anim. Physiol. Anim. Nutr. (Berl.) 2013, 97, 6–12. [Google Scholar] [CrossRef]
- King, J.C.; Shames, D.M.; Woodhouse, L.R. Zinc Homeostasis in Humans. J. Nutr. 2000, 130, 1360S–1366S. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Song, J.; Li, Y.; Luan, Z.; Zhu, K. Diosmectite-zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br. J. Nutr. 2013, 110, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Xiao, K.; Song, J.; Luan, Z.S. Effects of zinc oxide supported on zeolite on growth performance, intestinal microflora and permeability, and cytokines expression of weaned pigs. Anim. Feed Sci. Technol. 2013, 181, 65–71. [Google Scholar] [CrossRef]
- Johansen, M.; Jørgensen, L.; Schultz, M.S. Effect of Zinc and Organic Acids on Diarrhoea in the Weaner Period; SEGES: Copenhagen, Denmark, 2007. [Google Scholar]
- Poulsen, H. Zinc and copper as feed additives, growth factors or unwanted environmental factors. J. Anim. Feed Sci. 1998, 7, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Song, J.; You, Z.; Luan, Z.; Li, W. Zinc Oxide–Montmorillonite Hybrid Influences Diarrhea, Intestinal Mucosal Integrity, and Digestive Enzyme Activity in Weaned Pigs. Biol. Trace Elem. Res. 2012, 149, 190–196. [Google Scholar] [CrossRef]
- Case, C.L.; Carlson, M.S. Effect of feeding organic and inorganic sources of additional zinc on growth performance and zinc balance in nursery pigs. J. Anim. Sci. 2002, 80, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.D.; Baker, D.H. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci. 1993, 71, 3020–3024. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.C.; Sanz Fernandez, M.-V.; Torrison, J.; Wilson, M.E.; Baumgard, L.H.; Gabler, N.K. Dietary organic zinc attenuates heat stress–induced changes in pig intestinal integrity and metabolism. J. Anim. Sci. 2015, 93, 4702–4713. [Google Scholar] [CrossRef] [PubMed]
- Grilli, E.; Tugnoli, B.; Vitari, F.; Domeneghini, C.; Morlacchini, M.; Piva, A.; Prandini, A. Low doses of microencapsulated zinc oxide improve performance and modulate the ileum architecture, inflammatory cytokines and tight junctions expression of weaned pigs. Animal 2015, 9, 1760–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Lv, H.; Chen, Z.; Wang, L.; Wu, X.; Chen, Z.; Zhang, W.; Liang, R.; Jiang, Z. Dietary Zinc Oxide Modulates Antioxidant Capacity, Small Intestine Development, and Jejunal Gene Expression in Weaned Piglets. Biol. Trace Elem. Res. 2017, 175, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, S.R. The Antioxidant Properties of Zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.R. Critical Role of Zinc as Either an Antioxidant or a Prooxidant in Cellular Systems. Oxid. Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Carlson, M.S.; Hill, G.M.; Link, J.E. Early- and traditionally weaned nursery pigs benefit from phase-feeding pharmacological concentrations of zinc oxide: Effect on metallothionein and mineral concentrations. J. Anim. Sci. 1999, 77, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surfaces A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Söderberg, T.A.; Sunzel, B.; Holm, S.; Elmros, T.; Hallmans, G.; Sjöberg, S. Antibacterial effect of zinc oxide in vitro. Scand. J. Plast. Reconstr. Surg. Hand Surg. 1990, 24, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Surjawidjaja, J.E.; Hidayat, A.; Lesmana, M. Growth inhibition of enteric pathogens by zinc sulfate: An in vitro study. Med. Princ. Pract. 2004, 13, 286–289. [Google Scholar] [CrossRef]
- Højberg, O.; Canibe, N.; Poulsen, H.D.; Hedemann, M.S.; Jensen, B.B. Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Appl. Environ. Microbiol. 2005, 71, 2267–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katouli, M.; Melin, L.; Jensen-Waern, M.; Wallgren, P.; Möllby, R. The effect of zinc oxide supplementation on the stability of the intestinal flora with special reference to composition of coliforms in weaned pigs. J. Appl. Microbiol. 1999, 87, 564–573. [Google Scholar] [CrossRef] [Green Version]
- Mores, N.; Cristani, J.; Piffer, I.A.; Barioni, W.J.; Lima, G.M.M. Effects of zinc oxide on postweaning diarrhea control in pigs experimentally infected with E. coli. Arq. Bras. Med. Veterinária e Zootec. 1998, 50, 513–523. [Google Scholar]
- Roselli, M.; Finamore, A.; Garaguso, I.; Britti, M.S.; Mengheri, E. Zinc Oxide Protects Cultured Enterocytes from the Damage Induced by Escherichia coli. J. Nutr. 2003, 133, 4077–4082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sargeant, H.R.; Miller, H.M.; Shaw, M.-A. Inflammatory response of porcine epithelial IPEC J2 cells to enterotoxigenic E. coli infection is modulated by zinc supplementation. Mol. Immunol. 2011, 48, 2113–2121. [Google Scholar] [CrossRef]
- Gammoh, N.Z.; Rink, L. Zinc and the Immune System. In Nutrition and Immunity; Springer International Publishing: Cham, Switzerland, 2019; pp. 127–158. [Google Scholar]
- Ercan, M.T.; Bor, N.M. Phagocytosis by macrophages in zinc-deficient rats. Int. J. Radiat. Appl. Instrum. 1991, 18, 765–768. [Google Scholar] [CrossRef]
- Wirth, J.J.; Fraker, P.J.; Kierszenbaum, F. Zinc requirement for macrophage function: Effect of zinc deficiency on uptake and killing of a protozoan parasite. Immunology 1989, 68, 114–119. [Google Scholar]
- Rolles, B.; Maywald, M.; Rink, L. Influence of zinc deficiency and supplementation on NK cell cytotoxicity. J. Funct. Foods 2018, 48, 322–328. [Google Scholar] [CrossRef]
- Honscheid, A.; Rink, L.; Haase, H. T-Lymphocytes: A Target for Stimulatory and Inhibitory Effects of Zinc Ions. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 132–144. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014, 28, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Kloubert, V.; Blaabjerg, K.; Dalgaard, T.S.; Poulsen, H.D.; Rink, L.; Wessels, I. Influence of zinc supplementation on immune parameters in weaned pigs. J. Trace Elem. Med. Biol. 2018, 49, 231–240. [Google Scholar] [CrossRef]
- Ou, D.; Li, D.; Cao, Y.; Li, X.; Yin, J.; Qiao, S.; Wu, G. Dietary supplementation with zinc oxide decreases expression of the stem cell factor in the small intestine of weanling pigs. J. Nutr. Biochem. 2007, 18, 820–826. [Google Scholar] [CrossRef]
- Yin, J.; Li, X.; Li, D.; Yue, T.; Fang, Q.; Ni, J.; Zhou, X.; Wu, G. Dietary supplementation with zinc oxide stimulates ghrelin secretion from the stomach of young pigs. J. Nutr. Biochem. 2009, 20, 783–790. [Google Scholar] [CrossRef]
- MacDonald, R.S. The Role of Zinc in Growth and Cell Proliferation. J. Nutr. 2000, 130, 1500S–1508S. [Google Scholar] [CrossRef] [Green Version]
- Hedemann, M.S.; Jensen, B.B.; Poulsen, H.D. Influence of dietary zinc and copper on digestive enzyme activity and intestinal morphology in weaned pigs. J. Anim. Sci. 2006, 84, 3310–3320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brink, M.F.; Becker, D.E.; Terrill, S.W.; Jensen, A.H. Zinc Toxicity in the Weanling Pig. J. Anim. Sci. 1959, 18, 836–842. [Google Scholar] [CrossRef]
- Burrough, E.R.; De Mille, C.; Gabler, N.K. Zinc overload in weaned pigs: Tissue accumulation, pathology, and growth impacts. J. Vet. Diagnostic Investig. 2019, 31, 537–545. [Google Scholar] [CrossRef]
- Martin, L.; Pieper, R.; Schunter, N.; Vahjen, W.; Zentek, J. Performance, organ zinc concentration, jejunal brush border membrane enzyme activities and mRNA expression in piglets fed with different levels of dietary zinc. Arch. Anim. Nutr. 2013, 67, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Sugie, K.; Inukai, N.; Eguchi, O.; Oyamada, T.; Sawada, H.; Yamanaka, N.; Shibahara, T. Chronic pancreatitis in farmed pigs fed excessive zinc oxide. J. Vet. Diagn. Investig. 2020, 32, 689–694. [Google Scholar] [CrossRef]
- Poulsen, H.D.; Larsen, T. Zinc excretion and retention in growing pigs fed increasing levels of zinc oxide. Livest. Prod. Sci. 1995, 43, 235–242. [Google Scholar] [CrossRef]
- Meyer, T.A.; Lindemann, M.D.; Cromwell, G.L.; Monegue, H.J.; Inocencio, N. Effects of Pharmacological Levels of Zinc as Zinc Oxide on Fecal Zinc and Mineral Excretion in Weanling Pigs. Prof. Anim. Sci. 2002, 18, 162–168. [Google Scholar] [CrossRef]
- Monteiro, S.C.; Lofts, S.; Boxall, A.B.A. Pre-assessment of environmental impact of zinc and copper used in animal nutrition. EFSA Support. Publ. 2010, 7, 1–138. [Google Scholar] [CrossRef] [Green Version]
- Gräber, I.; Hansen, J.F.; Olesen, S.E.; Petersen, J.; Øtergaard, H.S.; Krogh, L. Accumulation of copper and zinc in Danish agricultural soils in intensive pig production areas. Geogr. Tidsskr. 2005, 105, 15–22. [Google Scholar] [CrossRef]
- Bak, J.L.; Jensen, J.; Larsen, M.M. Belysning af Kobber- og Zinkindholdet i Jord—Indhold og Udvikling i Kvadratnettet og Måling på Udvalgte Brugstyper; Aarhus Universitet, Institut for Bioscience: Aarhus, Denmark, 2015; Volume 159, ISBN 9788771560350. [Google Scholar]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and copper in animal feed—Development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb. Ecol. Health Dis. 2014, 25. [Google Scholar] [CrossRef] [Green Version]
- Slifierz, M.J.; Friendship, R.; Weese, J.S. Zinc oxide therapy increases prevalence and persistence of methicillin-resistant staphylococcus aureus in pigs: A randomized controlled trial. Zoonoses Public Health 2015, 62, 301–308. [Google Scholar] [CrossRef]
- Vahjen, W.; Pietruszyńska, D.; Starke, I.C.; Zentek, J. High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog. 2015, 7, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Bednorz, C.; Oelgeschläger, K.; Kinnemann, B.; Hartmann, S.; Neumann, K.; Pieper, R.; Bethe, A.; Semmler, T.; Tedin, K.; Schierack, P.; et al. The broader context of antibiotic resistance: Zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med. Microbiol. 2013, 303, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Ciesinski, L.; Guenther, S.; Pieper, R.; Kalisch, M.; Bednorz, C.; Wieler, L.H. High dietary zinc feeding promotes persistence of multi-resistant E. coli in the swine gut. PLoS ONE 2018, 13, e0191660. [Google Scholar] [CrossRef] [PubMed]
- Johanns, V.C.; Ghazisaeedi, F.; Epping, L.; Semmler, T.; Lübke-Becker, A.; Pfeifer, Y.; Bethe, A.; Eichhorn, I.; Merle, R.; Walther, B.; et al. Effects of a Four-Week High-Dosage Zinc Oxide Supplemented Diet on Commensal Escherichia coli of Weaned Pigs. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Slifierz, M.J.; Park, J.; Friendship, R.M.; Weese, J.S. Zinc-resistance gene CzrC identified in methicillin-resistant Staphylococcus hyicus isolated from pigs with exudative epidermitis. Can. Vet. J. La Rev. Vet. Can. 2014, 55, 489–490. [Google Scholar]
- Yu, T.; Zhu, C.; Chen, S.; Gao, L.; Lv, H.; Feng, R.; Zhu, Q.; Xu, J.; Chen, Z.; Jiang, Z. Dietary high zinc oxide modulates the microbiome of ileum and colon in weaned piglets. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Starke, I.C.; Pieper, R.; Neumann, K.; Zentek, J.; Vahjen, W. The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol. Ecol. 2014, 87, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Li, B.T.; Van Kessel, A.G.; Caine, W.R.; Huang, S.X.; Kirkwood, R.N. Small intestinal morphology and bacterial populations in ileal digesta and feces of newly weaned pigs receiving a high dietary level of zinc oxide. Can. J. Anim. Sci. 2001, 81, 511–516. [Google Scholar] [CrossRef]
- Vahjen, W.; Pieper, R.; Zentek, J. Bar-Coded Pyrosequencing of 16S rRNA Gene Amplicons Reveals Changes in Ileal Porcine Bacterial Communities Due to High Dietary Zinc Intake. Appl. Environ. Microbiol. 2010, 76, 6689–6691. [Google Scholar] [CrossRef] [Green Version]
- Pieper, R.; Vahjen, W.; Neumann, K.; Van Kessel, A.G.; Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J. Anim. Physiol. Anim. Nutr. (Berl.) 2012, 96, 825–833. [Google Scholar] [CrossRef]
- Vahjen, W.; Pieper, R.; Zentek, J. Increased dietary zinc oxide changes the bacterial core and enterobacterial composition in the ileum of piglets. J. Anim. Sci. 2011, 89, 2430–2439. [Google Scholar] [CrossRef] [Green Version]
- Starke, I.C.; Pieper, R.; Vahjen, W.; Zentek, J. The impact of dietary zinc oxide on the bacterial diversity of the small intestinal microbiota of weaned piglets. J. Vet. Sci. Technol. 2014, 5. [Google Scholar] [CrossRef]
- EFSA; FEEDAP. Scientific Opinion on the potential reduction of the currently authorised maximum zinc content in complete feed. EFSA J. 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Commission Implementing Decision of 26.6.2017 Concerning, in the Framework of Article 35 of Directive 2001/82/EC of the European Parliament and of the Council, the Marketing Authorisations for Veterinary Medicinal Products Containing “Zinc Oxide” to be Ad; Official Journal of the European Union: Brussels, Belgium, 2017. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1334/2003 of 25 July 2003 Amending the Conditions for Authorisation of a Number of Additives in Feedingstuffs Belonging to the Group of Trace Elements; Official Journal of the European Union: Brussels, Belgium, 2003. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2016/1095 of 6 July 2016; Official Journal of the European Union: Brussels, Belgium, 2016. [Google Scholar]
- Wedekind, K.J.; Baker, D.H. Zinc bioavailability in feed-grade sources of zinc. J. Anim. Sci. 1990, 68, 684–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scientific Committee for Animal Nutrition of the European Commission. Opinion of the Scientific Committee for Animal Nutrition on the Use of Zinc in Feedingstuffs; European Commission—Health & Consumer Protection Directorate General: Brussels, Belgium, 2003. [Google Scholar]
- Schell, T.C.; Kornegay, E.T. Zinc concentration in tissues and performance of weanling pigs fed pharmacological levels of zinc from ZnO, Zn-methionine, Zn-lysine, or ZnSO4. J. Anim. Sci. 1996, 74, 1584. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Hasman, H. Susceptibility of different bacterial species isolated from food animals to copper sulphate, zinc chloride and antimicrobial substances used for disinfection. Vet. Microbiol. 2004, 100, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Henry, P.R.; Ammerman, C.B.; Miles, R.D.; Littell, R.C. Relative bioavailability of basic zinc sulfate and basic zinc chloride for chicks. J. Appl. Poult. Res. 2000, 9, 513–517. [Google Scholar] [CrossRef]
- Batal, A.B.; Parr, T.M.; Baker, D.H. Zinc bioavailability in tetrabasic zinc chloride and the dietary zinc requirement of young chicks fed a soy concentrate diet. Poult. Sci. 2001, 80, 87–90. [Google Scholar] [CrossRef]
- Mavromichalis, I.; Webel, D.M.; Parr, E.N.; Baker, D.H. Growth-promoting efficacy of pharmacological doses of tetrabasic zinc chloride in diets for nursery pigs. Can. J. Anim. Sci. 2001, 81, 387–391. [Google Scholar] [CrossRef]
- Zhang, B.; Guo, Y. Beneficial effects of tetrabasic zinc chloride for weanling piglets and the bioavailability of zinc in tetrabasic form relative to ZnO. Anim. Feed Sci. Technol. 2007, 135, 75–85. [Google Scholar] [CrossRef]
- Zhang, G.; Xia, T.; Zhao, J.; Liu, L.; He, P.; Zhang, S.; Zhang, L. Moderate tetrabasic zinc chloride supplementation improves growth performance and reduces diarrhea incidence in weaned pigs. Asian-Australas. J. Anim. Sci. 2020, 33, 264–276. [Google Scholar] [CrossRef]
- Nitrayova, S.; Windisch, W.; von Heimendahl, E.; Müller, A.; Bartelt, J. Bioavailability of zinc from different sources in pigs. J. Anim. Sci. 2012, 90, 185–187. [Google Scholar] [CrossRef]
- Zhang, Y.; Ward, T.L.; Ji, F.; Peng, C.; Zhu, L.; Gong, L.; Dong, B. Effects of zinc sources and levels of zinc amino acid complex on growth performance, hematological and biochemical parameters in weanling pigs. Asian-Australas. J. Anim. Sci. 2018, 31, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, K.J.; Hortin, A.E.; Baker, D.H. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J. Anim. Sci. 1992, 70, 178–187. [Google Scholar] [CrossRef]
- Bouwhuis, M.A.; Sweeney, T.; Mukhopadhya, A.; Thornton, K.; McAlpine, P.O.; O’Doherty, J.V. Zinc methionine and laminarin have growth-enhancing properties in newly weaned pigs influencing both intestinal health and diarrhoea occurrence. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, 1273–1285. [Google Scholar] [CrossRef]
- Woodworth, J.C.; Tokach, M.D.; Nelssen, J.L.; Goodband, R.D.; Quinn, P.R.O.; Fakler, T.M. Interactive Effects of Diet Complexity, Zinc Source and Feed-grade Antibiotics on Weanling Pig Growth Performance. J. Anim. Vet. Adv. 2005, 4, 688–693. [Google Scholar]
- Hollis, G.R.; Carter, S.D.; Cline, T.R.; Crenshaw, T.D.; Cromwell, G.L.; Hill, G.M.; Kim, S.W.; Lewis, A.J.; Mahan, D.C.; Miller, P.S.; et al. Effects of replacing pharmacological levels of dietary zinc oxide with lower dietary levels of various organic zinc sources for weanling pigs. J. Anim. Sci. 2005, 83, 2123–2129. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Van Noten, N.; Degroote, J.; Romeo, A.; Vermeir, P.; Michiels, J. Effect of zinc oxide sources and dosages on gut microbiota and integrity of weaned piglets. J. Anim. Physiol. Anim. Nutr. (Berl.) 2019, 103, 231–241. [Google Scholar] [CrossRef]
- Shen, J.; Chen, Y.; Wang, Z.; Zhou, A.; He, M.; Mao, L.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; et al. Coated zinc oxide improves intestinal immunity function and regulates microbiota composition in weaned piglets. Br. J. Nutr. 2014, 111, 2123–2134. [Google Scholar] [CrossRef]
- Singh, S. Zinc oxide nanoparticles impacts: Cytotoxicity, genotoxicity, developmental toxicity, and neurotoxicity. Toxicol. Mech. Methods 2019, 29, 300–311. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, L.; Ying, Z.; He, J.; Zhou, L.; Zhang, L.; Zhong, X.; Wang, T. Effects of Dietary Zinc Oxide Nanoparticles on Growth, Diarrhea, Mineral Deposition, Intestinal Morphology, and Barrier of Weaned Piglets. Biol. Trace Elem. Res. 2018, 185, 364–374. [Google Scholar] [CrossRef]
- Ma, H.; Williams, P.L.; Diamond, S.A. Ecotoxicity of manufactured ZnO nanoparticles—A review. Environ. Pollut. 2013, 172, 76–85. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; ur Rahman, A.; Husen, A. Properties of Zinc Oxide Nanoparticles and Their Activity Against Microbes. Nanoscale Res. Lett. 2018, 13, 141. [Google Scholar] [CrossRef]
- Pei, X.; Xiao, Z.; Liu, L.; Wang, G.; Tao, W.; Wang, M.; Zou, J.; Leng, D. Effects of dietary zinc oxide nanoparticles supplementation on growth performance, zinc status, intestinal morphology, microflora population, and immune response in weaned pigs. J. Sci. Food Agric. 2019, 99, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, L.; Su, W.; Ying, Z.; He, J.; Zhang, L.; Zhong, X.; Wang, T. Zinc oxide nanoparticles as a substitute for zinc oxide or colistin sulfate: Effects on growth, serum enzymes, zinc deposition, intestinal morphology and epithelial barrier in weaned piglets. PLoS ONE 2017, 12, e0181136. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Chen, J.; Zhang, Y.; Liang, X.; Ni, H.; Zhang, B.; Yin, Y. Comparison of porous and nano zinc oxide for replacing high-dose dietary regular zinc oxide in weaning piglets. PLoS ONE 2017, 12, e0182550. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Gu, L.Y.; Luan, Z.S.; Song, J.; Zhu, K. Effects of montmorillonite-zinc oxide hybrid on performance, diarrhea, intestinal permeability and morphology of weanling pigs. Anim. Feed Sci. Technol. 2012, 177, 108–115. [Google Scholar] [CrossRef]
- Cho, J.H.; Liu, S.D.; Yun, W.; Kim, K.S.; Kim, I.H. Effect of supplemented microencapsulated zinc oxide and organic acids and pure botanicals on growth performance, nutrient digestibility, blood profiles, feces microflora, and zinc level of feces in weanling pigs. Can. J. Anim. Sci. 2019, 99, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.J.; Kim, I.H. Low dose of coated zinc oxide is as effective as pharmacological zinc oxide in promoting growth performance, reducing fecal scores, and improving nutrient digestibility and intestinal morphology in weaned pigs. Anim. Feed Sci. Technol. 2018, 245, 117–125. [Google Scholar] [CrossRef]
- Dong, X.; Xu, Q.; Wang, C.; Zou, X.; Lu, J. Supplemental-coated zinc oxide relieves diarrhoea by decreasing intestinal permeability in weanling pigs. J. Appl. Anim. Res. 2019, 47, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Pluske, J.R. Nutrition and pathology of weaner pigs: Nutritional strategies to support barrier function in the gastrointestinal tract. Anim. Feed Sci. Technol. 2012, 173, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Le Floc’h, N.; Jondreville, C.; Matte, J.J.; Seve, B. Importance of sanitary environment for growth performance and plasma nutrient homeostasis during the post-weaning period in piglets. Arch. Anim. Nutr. 2006, 60, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef]
- Gibbons, R.A.; Sellwood, R.; Burrows, M.; Hunter, P.A. Inheritance of resistance to neonatal E. coli diarrhoea in the pig: Examination of the genetic system. Theor. Appl. Genet. 1977, 51, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, I.G.W.; Bouw, J. Inheritance of K88-mediated adhesion of Escherichia coli to jejunal brush borders in pigs: A genetic analysis. Vet. Res. Commun. 1987, 11, 509–518. [Google Scholar] [CrossRef]
- Fontanesi, L.; Bertolini, F.; Dall’Olio, S.; Buttazzoni, L.; Gallo, M.; Russo, V. Analysis of Association between the MUC4 g.8227C>G Polymorphism and Production Traits in Italian Heavy Pigs Using a Selective Genotyping Approach. Anim. Biotechnol. 2012, 23, 147–155. [Google Scholar] [CrossRef]
- Rasschaert, K.; Verdonck, F.; Goddeeris, B.M.; Duchateau, L.; Cox, E. Screening of pigs resistant to F4 enterotoxigenic Escherichia coli (ETEC) infection. Vet. Microbiol. 2007, 123, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Nicolai, R.W.; Sørensen, T.; Bækbo, P. Weaning without Zinc Oxide—Field Experiences; SEGES: Copenhagen, Denmark, 2019. [Google Scholar]
- Halas, D.; Heo, J.M.; Hansen, C.F.; Kim, J.C.; Hampson, D.J.; Mullan, B.P.; Pluske, J.R. Organic acids, prebiotics and protein level as dietary tools to control the weaning transition and reduce post-weaning diarrhoea in piglets. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Maribo, H.; Kjeldsen, N.; Pluske, J.R. Effects of dietary protein level and zinc oxide supplementation on the incidence of post-weaning diarrhoea in weaner pigs challenged with an enterotoxigenic strain of Escherichia coli. Livest. Sci. 2010, 133, 210–213. [Google Scholar] [CrossRef]
- Molist, F.; Gómez de Segura, A.; Pérez, J.F.; Bhandari, S.K.; Krause, D.O.; Nyachoti, C.M. Effect of wheat bran on the health and performance of weaned pigs challenged with Escherichia coli K88+. Livest. Sci. 2010, 133, 214–217. [Google Scholar] [CrossRef]
- Fernandes, C.D.; Resende, M.; Rodrigues, L.M.; Garbossa, C.A.P.; Costa, L.B.; Ferreira, R.A.; de Abreu, M.L.T.; Cantarelli, V.S. Dietary fiber and zinc additives on performance and intestinal health of Escherichia coli challenged piglets. Sci. Agric. 2020, 77. [Google Scholar] [CrossRef] [Green Version]
- Molist, F.; Hermes, R.G.; De Segura, A.G.; Martí-N-Orúe, S.M.; Gasa, J.; Manzanilla, E.G.; Pérez, J.F. Effect and interaction between wheat bran and zinc oxide on productive performance and intestinal health in post-weaning piglets. Br. J. Nutr. 2011, 105, 1592–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tugnoli, B.; Giovagnoni, G.; Piva, A.; Grilli, E. From acidifiers to intestinal health enhancers: How organic acids can improve growth efficiency of pigs. Animals 2020, 10, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiloyiannis, V.K.; Kyriakis, S.C.; Vlemmas, J.; Sarris, K. The effect of organic acids on the control of porcine post-weaning diarrhoea. Res. Vet. Sci. 2001, 70, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single components of botanicals and nature-identical compounds as a non-antibiotic strategy to ameliorate health status and improve performance in poultry and pigs. Nutr. Res. Rev. 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, J.D. Antibacterial and antidiarrheal activities of plant products against enterotoxinogenic Escherichia coli. Toxins (Basel) 2013, 5, 2009–2041. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Verhelst, R.; Schroyen, M.; Buys, N.; Niewold, T. The effects of plant polyphenols on enterotoxigenic Escherichia coli adhesion and toxin binding. Livest. Sci. 2010, 133, 101–103. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef]
- Xiao, H.; Shao, F.; Wu, M.; Ren, W.; Xiong, X.; Tan, B.; Yin, Y. The application of antimicrobial peptides as growth and health promoters for swine. J. Anim. Sci. Biotechnol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mine, Y.; Kovacs-Nolan, J. Chicken egg yolk antibodies as therapeutics in enteric infectious disease: A review. J. Med. Food 2002, 5, 159–169. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Polo, J.; Torrallardona, D. Spray dried plasma as an alternative to antibiotics in piglet feeds, mode of action and biosafety. Porc. Health Manag. 2016, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Pieper, R.; Villodre Tudela, C.; Taciak, M.; Bindelle, J.; Pérez, J.F.; Zentek, J. Health relevance of intestinal protein fermentation in young pigs. Anim. Health Res. Rev. 2016, 17, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. Effects of dietary protein supply, weaning age and experimental enterotoxigenic Escherichia coli infection on newly weaned pigs: Health. Animal 2008, 2, 834–842. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Yue, L.Y.; Qiao, S.Y. Effects of low-protein diets supplemented with crystalline amino acids on performance and intestinal development in piglets over the first 2 weeks after weaning. Livest. Sci. 2008, 115, 144–152. [Google Scholar] [CrossRef]
- Kjeldsen, N.J.; Lynegaard, J.C.; Bache, J.K. Low Protein for Weaned Pigs Can Reduce Diarrhoea; SEGES: Copenhagen, Denmark, 2019. [Google Scholar]
- Wan, K.; Li, Y.; Sun, W.; An, R.; Tang, Z.; Wu, L.; Chen, H.; Sun, Z. Effects of dietary calcium pyruvate on gastrointestinal tract development, intestinal health and growth performance of newly weaned piglets fed low-protein diets. J. Appl. Microbiol. 2020, 128, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Effects of feeding low protein diets to piglets on plasma urea nitrogen, faecal ammonia nitrogen, the incidence of diarrhoea and performance after weaning. Arch. Anim. Nutr. 2008, 62, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Zhang, S.H.; Zeng, X.F.; Liu, H.; Qiao, S.Y. Branched-chain amino acids are beneficial to maintain growth performance and intestinal immune-related function in weaned piglets fed protein restricted diet. Asian-Australas. J. Anim. Sci. 2015, 28, 1742–1750. [Google Scholar] [CrossRef] [Green Version]
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascoal, L.A.F.; Thomaz, M.C.; Watanabe, P.H.; Ruiz, U.D.S.; Amorim, A.B.; Daniel, E.; Silva, S.Z.D. Purified cellulose, soybean hulls and citrus pulp as a source of fiber for weaned piglets. Sci. Agric. 2015, 72, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Molist, F.; de Segura, A.G.; Gasa, J.; Hermes, R.G.; Manzanilla, E.G.; Anguita, M.; Pérez, J.F. Effects of the insoluble and soluble dietary fibre on the physicochemical properties of digesta and the microbial activity in early weaned piglets. Anim. Feed Sci. Technol. 2009, 149, 346–353. [Google Scholar] [CrossRef]
- González-Ortiz, G.; Pérez, J.F.; Hermes, R.G.; Molist, F.; Jiménez-Díaz, R.; Martín-Orúe, S.M. Screening the ability of natural feed ingredients to interfere with the adherence of enterotoxigenic Escherichia coli (ETEC) K88 to the porcine intestinal mucus. Br. J. Nutr. 2014, 111, 633–642. [Google Scholar] [CrossRef] [Green Version]
- González-Ortiz, G.; Hermes, R.G.; Jiménez-Díaz, R.; Pérez, J.F.; Martín-Orúe, S.M. Screening of extracts from natural feed ingredients for their ability to reduce enterotoxigenic Escherichia coli (ETEC) K88 adhesion to porcine intestinal epithelial cell-line IPEC-J2. Vet. Microbiol. 2013, 167, 494–499. [Google Scholar] [CrossRef]
- Hansen, C.F.; Riis, A.L.; Bresson, S.; Højbjerg, O.; Jensen, B.B. Feeding organic acids enhances the barrier function against pathogenic bacteria of the piglet stomach. Livest. Sci. 2007, 108, 206–209. [Google Scholar] [CrossRef]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-chain fatty acids and monoglycerides as feed additives for pig production: Towards gut health improvement and feed pathogen mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Namkung, H.; Li, M.; Gong, J.; Yu, H.; Cottrill, M.; De Lange, C.F.M. Impact of feeding blends of organic acids and herbal extracts on growth performance, gut microbiota and digestive function in newly weaned pigs. Can. J. Anim. Sci. 2004, 84, 697–704. [Google Scholar] [CrossRef]
- Roselli, M.; Finamore, A.; Britti, M.S.; Bosi, P.; Oswald, I.; Mengheri, E. Alternatives to in-feed antibiotics in pigs: Evaluation of probiotics, zinc or organic acids as protective agents for the intestinal mucosa. A comparison of in vitro and in vivo results. Anim. Res. 2005, 54, 203–218. [Google Scholar] [CrossRef]
- Ferrara, F.; Tedin, L.; Pieper, R.; Meyer, W.; Zentek, J. Influence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Hanczakowska, E.; Szewczyk, A.; Okoñ, K. Effects of dietary caprylic and capric acids on piglet performance and mucosal epithelium structure of the ileum. J. Anim. Feed Sci. 2011, 20, 556–565. [Google Scholar] [CrossRef]
- Piva, A.; Morlacchini, M.; Casadei, G.; Gatta, P.P.; Biagi, G.; Prandini, A. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 2002, 1, 35–41. [Google Scholar] [CrossRef]
- Grilli, E.; Tugnoli, B.; Foerster, C.J.; Piva, A. Butyrate modulates inflammatory cytokines and tight junctions components along the gut of weaned pigs. J. Anim. Sci. 2016, 94, 433–436. [Google Scholar] [CrossRef]
- Xiong, H.; Guo, B.; Gan, Z.; Song, D.; Lu, Z.; Yi, H.; Wu, Y.; Wang, Y.; Du, H. Butyrate upregulates endogenous host defense peptides to enhance disease resistance in piglets via histone deacetylase inhibition. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonetti, A.; Tugnoli, B.; Rossi, B.; Giovagnoni, G.; Piva, A.; Grilli, E. Nature-Identical Compounds and Organic Acids Reduce E. coli K88 Growth and Virulence Gene Expression In Vitro. Toxins (Basel) 2020, 12, 468. [Google Scholar] [CrossRef]
- Bosi, P.; Sarli, G.; Casini, L.; De Filippi, S.; Trevisi, P.; Mazzoni, M.; Merialdi, G. The influence of fat protection of calcium formate on growth and intestinal defence in Escherichia coli K88-challenged weanling pigs. Anim. Feed Sci. Technol. 2007, 139, 170–185. [Google Scholar] [CrossRef]
- Ren, C.; Zhou, Q.; Guan, W.; Lin, X.; Wang, Y.; Song, H.; Zhang, Y. Immune response of piglets receiving mixture of formic and propionic acid alone or with either capric acid or bacillus licheniformis after Escherichia coli challenge. Biomed Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Zeghib, A.; Kabouche, A.; Laggoune, S.; Calliste, C.A.; Simon, A.; Bressolier, P.; Aouni, M.; Duroux, J.L.; Kabouche, Z. Antibacterial, antiviral, antioxidant and antiproliferative activities of thymus guyonii essential oil. Nat. Prod. Commun. 2017, 12, 1651–1654. [Google Scholar] [CrossRef] [Green Version]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yoo, J.S.; Kim, H.J.; Wang, Y.; Chen, Y.J.; Cho, J.H.; Kim, I.H. Effects of dietary supplementation with blended essential oils on growth performance, nutrient digestibility, blood profiles and fecal characteristics in weanling Pigs. Asian-Australas. J. Anim. Sci. 2010, 23, 607–613. [Google Scholar] [CrossRef]
- Li, P.; Piao, X.; Ru, Y.; Han, X.; Xue, L.; Zhang, H. Effects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian-Australas. J. Anim. Sci. 2012, 25, 1617–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.Y.; Piao, X.S. Essential oil blend could decrease diarrhea prevalence by improving antioxidative capability for weaned pigs. Animals 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.R.; Li, X.L.; Awati, A.; Bento, H.; Zhang, H.J.; Bontempo, V. Effect of an essential oils blend on growth performance, and selected parameters of oxidative stress and antioxidant defence of Escherichia coli challenged piglets. J. Anim. Feed Sci. 2017, 26, 38–43. [Google Scholar] [CrossRef]
- Thacker, P.A. Alternatives to antibiotics as growth promoters for use in swine production: A review. J. Anim. Sci. Biotechnol. 2013, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nezhadali, A.; Nabavi, M.; Rajabian, M.; Akbarpour, M.; Pourali, P.; Amini, F. Chemical variation of leaf essential oil at different stages of plant growth and in vitro antibacterial activity of Thymus vulgaris Lamiaceae, from Iran. Beni-Suef Univ. J. Basic Appl. Sci. 2014, 3, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Grilli, E.; Tugnoli, B.; Passey, J.L.; Stahl, C.H.; Piva, A.; Moeser, A.J. Impact of dietary organic acids and botanicals on intestinal integrity and inflammation in weaned pigs. BMC Vet. Res. 2015, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.C.; Paiva de Sousa, C.; Fernandez-Prada, C.; Harel, J.; Dubreuil, J.D.; de Souza, E.L. A review of the current evidence of fruit phenolic compounds as potential antimicrobials against pathogenic bacteria. Microb. Pathog. 2019, 130, 259–270. [Google Scholar] [CrossRef]
- Dubreuil, J.D. Fruit extracts to control pathogenic Escherichia coli: A sweet solution. Heliyon 2020, 6, e03410. [Google Scholar] [CrossRef] [PubMed]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In Vitro Digestion of Chestnut and Quebracho Tannin Extracts: Antimicrobial Effect, Antioxidant Capacity and Cytomodulatory Activity in Swine Intestinal IPEC-J2 Cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkung, W.; Thiagarajah, J.R.; Phuan, P.; Verkman, A.S. Inhibition of Ca2+-activated Cl- channels by gallotannins as a possible molecular basis for health benefits of red wine and green tea. FASEB J. 2010, 24, 4178–4186. [Google Scholar] [CrossRef] [Green Version]
- Verhelst, R.; Schroyen, M.; Buys, N.; Niewold, T. Selection of Escherichia coli heat-labile toxin (LT) inhibitors using both the gm1-elisa and the cAMP Vero cell assay. Foodborne Pathog. Dis. 2013, 10, 603–607. [Google Scholar] [CrossRef]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compound-nature, occurrence, dietary intake and effects. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M. Functional components of grape pomace: Their composition, biological properties and potential applications. Int. J. Food Sci. Technol. 2013, 48, 221–237. [Google Scholar] [CrossRef]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol-thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Spees, A.M.; Wangdi, T.; Lopez, C.A.; Kingsbury, D.D.; Xavier, M.N.; Winter, S.E.; Tsolis, R.M.; Bäumler, A.J. Streptomycin-Induced Inflammation Enhances Escherichia coli Gut Colonization Through Nitrate Respiration. MBio 2013, 4, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.H.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, J.; Mahfuz, S.; Piao, X. Effects of Hydrolysable Tannins as Zinc Oxide Substitutes on Antioxidant Status, Immune Function, Intestinal Morphology, and Digestive Enzyme Activities in Weaned Piglets. Animals 2020, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Facinelli, B.; Ferrante, L.; Capretti, R.; Orazio, G. Human Milk Oligosaccharides Inhibit the Adhesion to Caco-2 Cells of Diarrheal Pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr. Res. 2006, 59, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Sarabia-Sainz, H.M.; Armenta-Ruiz, C.; Sarabia-Sainz, J.A.I.; Guzmán-Partida, A.M.; Ledesma-Osuna, A.I.; Vázquez-Moreno, L.; Montfort, G.R.C. Adhesion of enterotoxigenic Escherichia coli strains to neoglycans synthesised with prebiotic galactooligosaccharides. Food Chem. 2013, 141, 2727–2734. [Google Scholar] [CrossRef]
- Hermes, R.G.; Manzanilla, E.G.; Martín-Orúe, S.M.; Pérez, J.F.; Klasing, K.C. Influence of dietary ingredients on in vitro inflammatory response of intestinal porcine epithelial cells challenged by an enterotoxigenic Escherichia coli (K88). Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 479–488. [Google Scholar] [CrossRef]
- Stuyven, E.; Cox, E.; Vancaeneghem, S.; Arnouts, S.; Deprez, P.; Goddeeris, B.M. Effect of β-glucans on an ETEC infection in piglets. Vet. Immunol. Immunopathol. 2009, 128, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Zhang, J.; Chen, D.; Yu, B.; Mao, X.; Zheng, P.; Yu, J.; Huang, Z.; Luo, J.; Luo, Y.; et al. Alginate oligosaccharide alleviates enterotoxigenic: Escherichia coli-induced intestinal mucosal disruption in weaned pigs. Food Funct. 2018, 9, 6401–6413. [Google Scholar] [CrossRef]
- Dubreuil, J.D. Enterotoxigenic Escherichia coli and probiotics in swine: What the bleep do we know? Biosci. Microbiota Food Health 2017, 36, 75–90. [Google Scholar] [CrossRef] [Green Version]
- Roselli, M.; Finamore, A.; Britti, M.S.; Mengheri, E. Probiotic bacteria Bifidobacterium animalis MB5 and Lactobacillus rhamnosus GG protect intestinal Caco-2 cells from the inflammation-associated response induced by enterotoxigenic Escherichia coli K88. Br. J. Nutr. 2006, 95, 1177. [Google Scholar] [CrossRef]
- Roselli, M.; Finamore, A.; Britti, M.S.; Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Mengheri, E. The Novel Porcine Lactobacillus sobrius Strain Protects Intestinal Cells from Enterotoxigenic Escherichia coli K88 Infection and Prevents Membrane Barrier Damage. J. Nutr. 2007, 137, 2709–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xu, Y.Q.; Liu, H.Y.; Lai, T.; Ma, J.L.; Wang, J.F.; Zhu, Y.H. Evaluation of Lactobacillus rhamnosus GG using an Escherichia coli K88 model of piglet diarrhoea: Effects on diarrhoea incidence, faecal microflora and immune responses. Vet. Microbiol. 2010, 141, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Bertocchi, M.; Motta, V.; Salvarani, C.; Bosi, P.; Luppi, A.; Fanelli, F.; Mazzoni, M.; Archetti, I.; Maiorano, G.; et al. Bacillus sp. probiotic supplementation diminish the Escherichia coli F4ac infection in susceptible weaned pigs by influencing the intestinal immune response, intestinal microbiota and blood metabolomics. J. Anim. Sci. Biotechnol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Nordeste, R.; Tessema, A.; Sharma, S.; Kovač, Z.; Wang, C.; Morales, R.; Griffiths, M.W. Molecules produced by probiotics prevent enteric colibacillosis in pigs. BMC Vet. Res. 2017, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Trckova, M.; Faldyna, M.; Alexa, P.; Zajacova, Z.S.; Gopfert, E.; Kumprechtova, D.; Auclair, E.; D’Inca, R. The effects of live yeast Saccharomyces cerevisiae on postweaning diarrhea, immune response, and growth performance in weaned piglets. J. Anim. Sci. 2014, 92, 767–774. [Google Scholar] [CrossRef]
- Krause, D.O.; Bhandari, S.K.; House, J.D.; Nyachoti, C.M. Response of nursery pigs to a synbiotic preparation of starch and an anti-Escherichia coli K88 probiotic. Appl. Environ. Microbiol. 2010, 76, 8192–8200. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Ordaz, A.A.; González-Ortiz, G.; La Ragione, R.M.; Woodward, M.J.; Collins, J.W.; Pérez, J.F.; Martín-Orúe, S.M. Lactulose and Lactobacillus plantarum, a potential complementary synbiotic to control postweaning colibacillosis in piglets. Appl. Environ. Microbiol. 2014, 80, 4879–4886. [Google Scholar] [CrossRef] [Green Version]
- Badia, R.; Zanello, G.; Chevaleyre, C.; Lizardo, R.; Meurens, F.; Martínez, P.; Brufau, J.; Salmon, H. Effect of Saccharomyces cerevisiae var. Boulardii and β-galactomannan oligosaccharide on porcine intestinal epithelial and dendritic cells challenged in vitro with Escherichia coli F4 (K88). Vet. Res. 2012, 43, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, S.; Mandal, S.; Tomar, S.K. Effect of Lactobacillus rhamnosus NCDC 298 with FOS in Combination on Viability and Toxin Production of Enterotoxigenic Escherichia coli. Probiotics Antimicrob. Proteins 2019, 11, 23–29. [Google Scholar] [CrossRef]
- Bradshaw, J.P. Cationic Antimicrobial Peptides. BioDrugs 2003, 17, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, F.; Huang, Z.; Liu, H.; Xie, C.; Zhang, J.; Thacker, P.A.; Qiao, S. Effects of the antimicrobial peptide cecropin AD on performance and intestinal health in weaned piglets challenged with Escherichia coli. Peptides 2012, 35, 225–230. [Google Scholar] [CrossRef]
- Cutler, S.A.; Lonergan, S.M.; Cornick, N.; Johnson, A.K.; Stahl, C.H. Dietary inclusion of colicin E1 is effective in preventing postweaning diarrhea caused by F18-positive Escherichia coli in pigs. Antimicrob. Agents Chemother. 2007, 51, 3830–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Huang, Y.; Sun, S.; Wu, Z.; Wu, S.; Yin, Z.; Bao, W. The impact of BPI expression on Escherichia coli f18 infection in porcine kidney cells. Animals 2020, 10, 2118. [Google Scholar] [CrossRef]
- Johnson, R.P.; Gyles, C.L.; Huff, W.E.; Ojha, S.; Huff, G.R.; Rath, N.C.; Donoghue, A.M. Bacteriophages for prophylaxis and therapy in cattle, poultry and pigs. Anim. Health Res. Rev. 2008, 9, 201–215. [Google Scholar] [CrossRef]
- Cha, S.B.; Yoo, A.N.; Lee, W.J.; Shin, M.K.; Jung, M.H.; Shin, S.W.; Cho, Y.W.; Yoo, H.S. Effect of bacteriophage in enterotoxigenic Escherichia coli (ETEC) infected pigs. J. Vet. Med. Sci. 2012, 74, 1037–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.Y.; Kim, S.J.; Park, B.C.; Han, J.H. Effects of dietary supplementation of bacteriophages against enterotoxigenic Escherichia coli (ETEC) K88 on clinical symptoms of post-weaning pigs challenged with the ETEC pathogen. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, 88–95. [Google Scholar] [CrossRef]
- Jamalludeen, N.; Johnson, R.P.; Shewen, P.E.; Gyles, C.L. Evaluation of bacteriophages for prevention and treatment of diarrhea due to experimental enterotoxigenic Escherichia coli O149 infection of pigs. Vet. Microbiol. 2009, 136, 135–141. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Cao, Z.; Wang, L.; Li, X.; Li, S.; Xu, Y. Bacteriophages as antimicrobial agents against major pathogens in swine: A review. J. Anim. Sci. Biotechnol. 2015, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Colomer-Lluch, M.; Imamovic, L.; Jofre, J.; Muniesa, M. Bacteriophages carrying antibiotic resistance genes in fecal waste from cattle, pigs, and poultry. Antimicrob. Agents Chemother. 2011, 55, 4908–4911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquardt, R.R.; Jin, L.Z.; Kim, J.W.; Fang, L.; Frohlich, A.A.; Baidoo, S.K. Passive protective effect of egg-yolk antibodies against enterotoxigenic Escherichia coli K88+ infection in neonatal and early-weaned piglets. FEMS Immunol. Med. Microbiol. 1999, 23, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, L.; Zhen, Y.; Li, S.; Xu, Y. Chicken egg yolk antibodies (IgY) as non-antibiotic production enhancers for use in swine production: A review. J. Anim. Sci. Biotechnol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.Z.; Baidoo, S.K.; Marquardt, R.R.; Frohlich, A.A. In vitro inhibition of adhesion of enterotoxigenic Escherichia coli K88 to piglet intestinal mucus by egg-yolk antibodies. FEMS Immunol. Med. Microbiol. 1998, 21, 313–321. [Google Scholar] [CrossRef]
- Chernysheva, L.V.; Friendship, R.M.; Abvp, D.; Dewey, C.E.; Gyles, C.L. The effect of dietary chicken egg-yolk antibodies on the clinical response in weaned pigs challenged with a K88+ Escherichia coli isolate. J. Swine Health Prod. 2004, 12, 119–122. [Google Scholar]
- Owusu-Asiedu, A.; Nyachoti, C.M.; Baidoo, S.K.; Marquardt, R.R.; Yang, X. Response of early-weaned pigs to an enterotoxigenic Escherichia coli (K88) challenge when fed diets containing spray-dried porcine plasma or pea protein isolate plus egg yolk antibody. J. Anim. Sci. 2003, 81, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Asiedu, A.; Nyachoti, C.M.; Marquardt, R.R. Response of early-weaned pigs to an enterotoxigenic Escherichia coli (K88) challenge when fed diets containing spray-dried porcine plasma or pea protein isolate plus egg yolk antibody, zinc oxide, fumaric acid, or antibiotic. J. Anim. Sci. 2003, 81, 1790–1798. [Google Scholar] [CrossRef] [Green Version]
- Yi, G.F.; Carroll, J.A.; Allee, G.L.; Gaines, A.M.; Kendall, D.C.; Usry, J.L.; Toride, Y.; Izuru, S. Effect of glutamine and spray-dried plasma on growth performance, small intestinal morphology, and immune responses of Escherichia coli K88 +-challenged weaned pigs. J. Anim. Sci. 2005, 83, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Bosi, P.; Casini, L.; Finamore, A.; Cremokolini, C.; Merialdi, G.; Trevisi, P.; Nobili, F.; Mengheri, E. Spray-dried plasma improves growth performance and reduces inflammatory status of weaned pigs challenged with enterotoxigenic Escherichia coli K88. J. Anim. Sci. 2004, 82, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Adewole, D.I.; Kim, I.H.; Nyachoti, C.M. Gut Health of Pigs: Challenge Models and Response Criteria with a Critical Analysis of the Effectiveness of Selected Feed Additives—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 909–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Beneficial effects and mechanisms of action of zinc oxide in post-weaning piglets. For each parameter, the up arrow “↑” means an increase, while the down arrow “↓” identifies a reduction.
Figure 1.
Beneficial effects and mechanisms of action of zinc oxide in post-weaning piglets. For each parameter, the up arrow “↑” means an increase, while the down arrow “↓” identifies a reduction.

Figure 2.
Risks related to pharmacological zinc oxide utilization in post-weaning piglets.


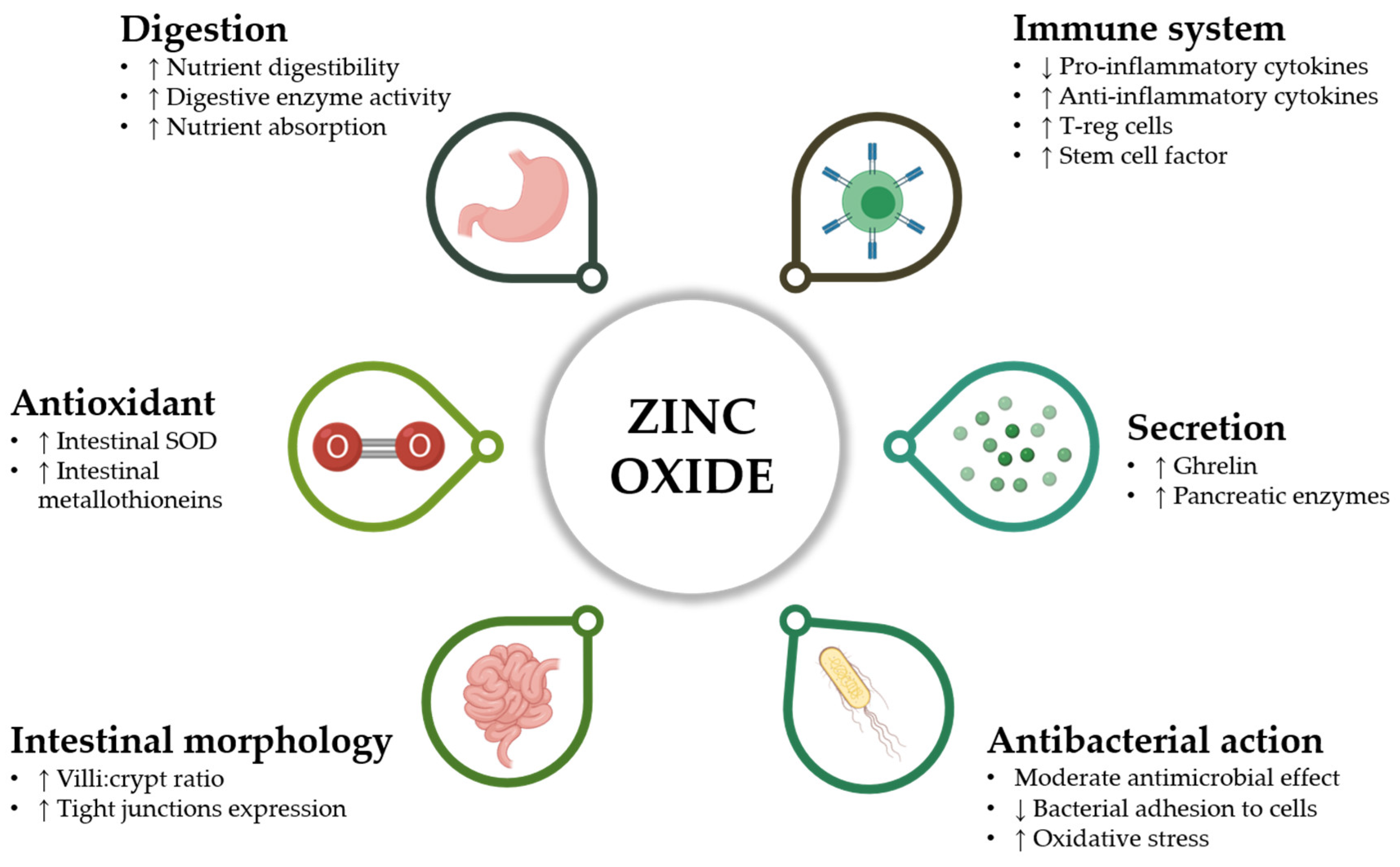{kind=link}
{kind=link}
Table 1.
Advantages and disadvantages of the main ZnO feeding alternatives for managing post-weaning diarrhea (PWD) and Escherichia coli F4 (K88) infections in weanling piglets. For each parameter, the up arrow “↑” means an increase, while the down arrow “↓” identifies a reduction.
Table 1.
Advantages and disadvantages of the main ZnO feeding alternatives for managing post-weaning diarrhea (PWD) and Escherichia coli F4 (K88) infections in weanling piglets. For each parameter, the up arrow “↑” means an increase, while the down arrow “↓” identifies a reduction.
| ZnO Feeding Alternatives | Advantages | Disadvantages | References |
|---|---|---|---|
| Low protein diets | ↓ proteolytic bacteria population ↓ pathogenic E. coli ↓ PWD symptoms ↓ pro-inflammatory cytokines | ↓ pig productivity | [124,125] |
| High fiber diets | ↓ PWD symptoms ↓ E. coli shedding ↓ E. coli adhesion ↑ SCFA production in digesta ↓ retention time of digesta | Few comparisons with pharmacological ZnO | [126,127,128] |
| Organic acids | Powerful antimicrobial activity Potential complete pharmacological Zn elimination ↑ gastric acidity ↑ nutrient digestibility ↑ growth performance ↓ PWD symptoms ↓ harmful coliforms ↓ inflammation ↑ intestinal morphology | Lack of comparisons with pharmacological ZnO Different activities between acids | [129,130] |
| Essential oils and nature identical compounds | Powerful antimicrobial, antioxidant, and anti-inflammatory activity ↑ growth performance ↑ diet digestibility ↓ PWD symptoms ↓ harmful coliforms ↓ bacterial virulence gene expression ↑ intestinal morphology | High variability in efficacy among different EO and molecules | [131,132,133] |
| Polyphenol-rich extracts | Considerable antimicrobial activity Ion chelating capacity High antioxidant activity ↓ PWD symptoms ↓ bacterial virulence gene expression ↓ bacterial adhesion to enterocytes ↓ bacterial toxin action ↑ intestinal morphology ↑ digestive enzymes activity | Few studies comparing polyphenols to pharmacological ZnO Polyphenols mechanism of action not yet fully elucidated | [134,135,136] |
| Antimicrobial peptides | Powerful antimicrobial activity ↓ bacterial resistance acquisition ↓ bacterial adhesion ↑ growth performance ↓ PWD symptoms ↑ intestinal morphology ↓ harmful coliforms ↑ immune response parameters | Lack of comparisons with pharmacological ZnO Need to investigate AMP pharmacokinetics | [137,138] |
| Egg yolk antibodies | ↑ growth performance ↓ PWD symptoms ↓ bacterial adhesion to enterocytes | Lack of comparisons with pharmacological ZnO High cost Stability issues in the gastrointestinal tract | [139] |
| Spray-dried plasma | ↑ growth performance ↓ PWD symptoms ↓ E. coli shedding ↑ intestinal morphology ↓ pro-inflammatory cytokines | Lack of comparisons with pharmacological ZnO High cost Complex production Potential presence of pathogens in SDP | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. https://doi.org/10.3390/ani11030642
AMA Style
Bonetti A, Tugnoli B, Piva A, Grilli E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals. 2021; 11(3):642. https://doi.org/10.3390/ani11030642
Chicago/Turabian StyleBonetti, Andrea, Benedetta Tugnoli, Andrea Piva, and Ester Grilli. 2021. "Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets" Animals 11, no. 3: 642. https://doi.org/10.3390/ani11030642
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.