
Endophytic Fungi Accelerate Leaf Physiological Activity and Resveratrol Accumulation in Polygonum cuspidatum by Up-Regulating Expression of Associated Genes

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Culture and Endophytic Fungal Inoculation
2.2. Experimental Design
2.3. Determination of Root Fungal Colonization and Biomass Production
2.4. Determination of Chlorophyll, Soluble Protein, and Gas Exchange
2.5. Determination of Medicinal Components in Leaves
2.6. Determination of Relative Expression Levels of Resveratrol-Synthesis-Related Enzyme Genes
2.7. Statistical Analysis
3. Results
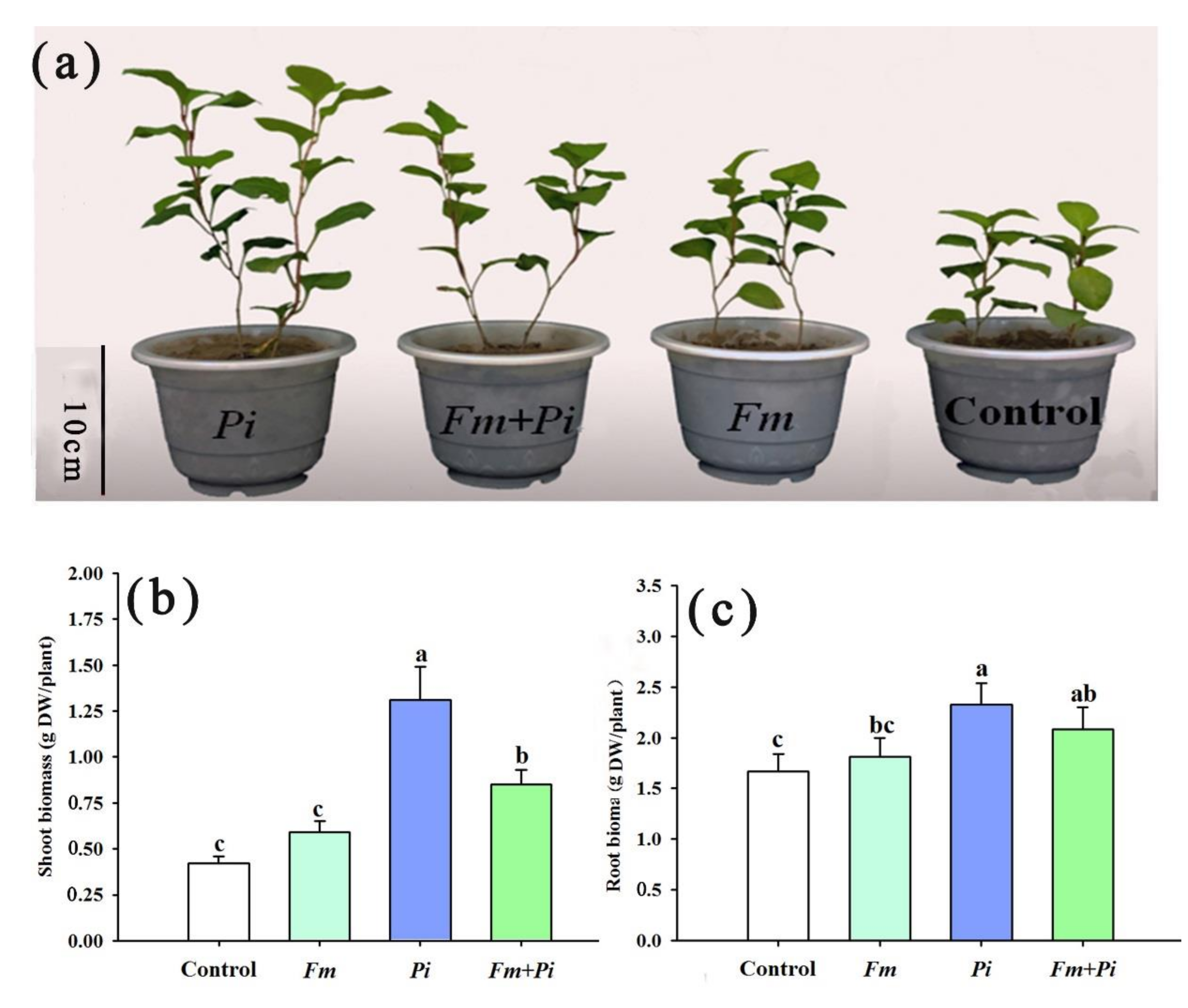
3.1. Changes in Root Fungal Colonization Frequency and Biomass Production
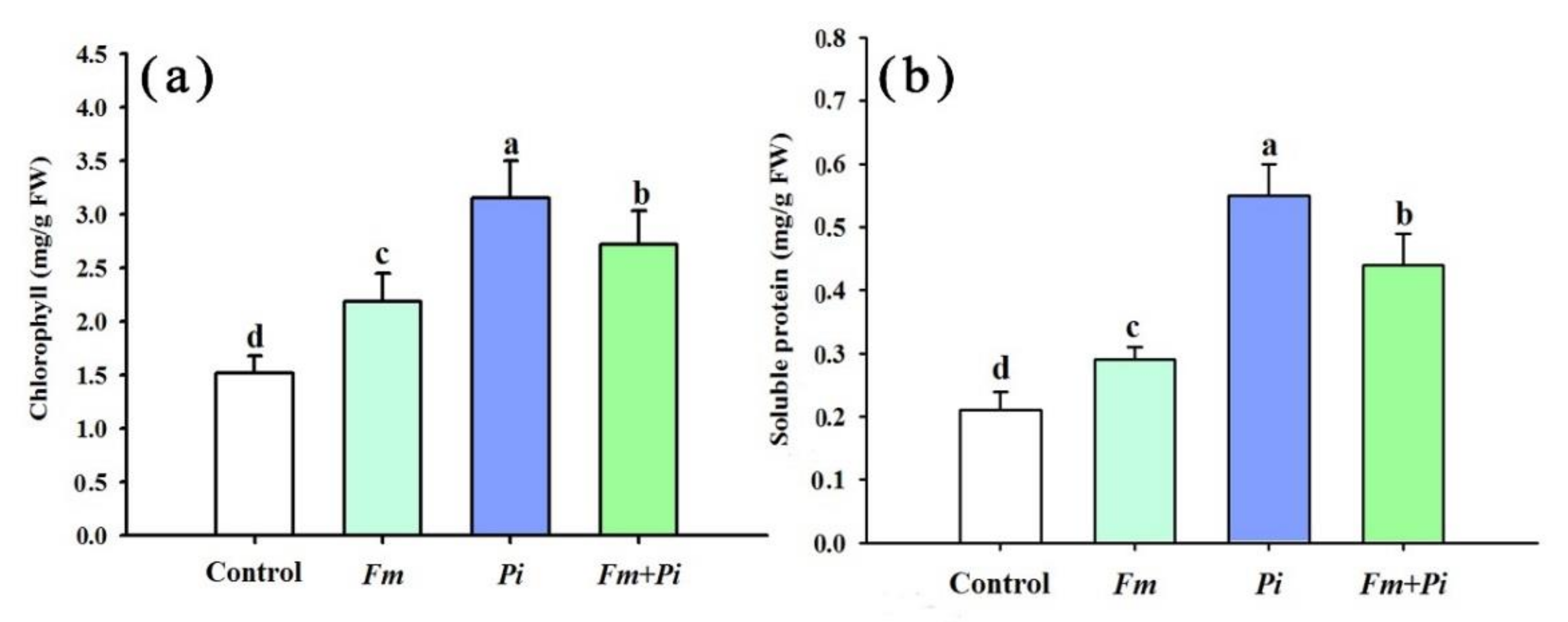
3.2. Changes in Leaf Chlorophyll and Soluble-Protein Concentrations
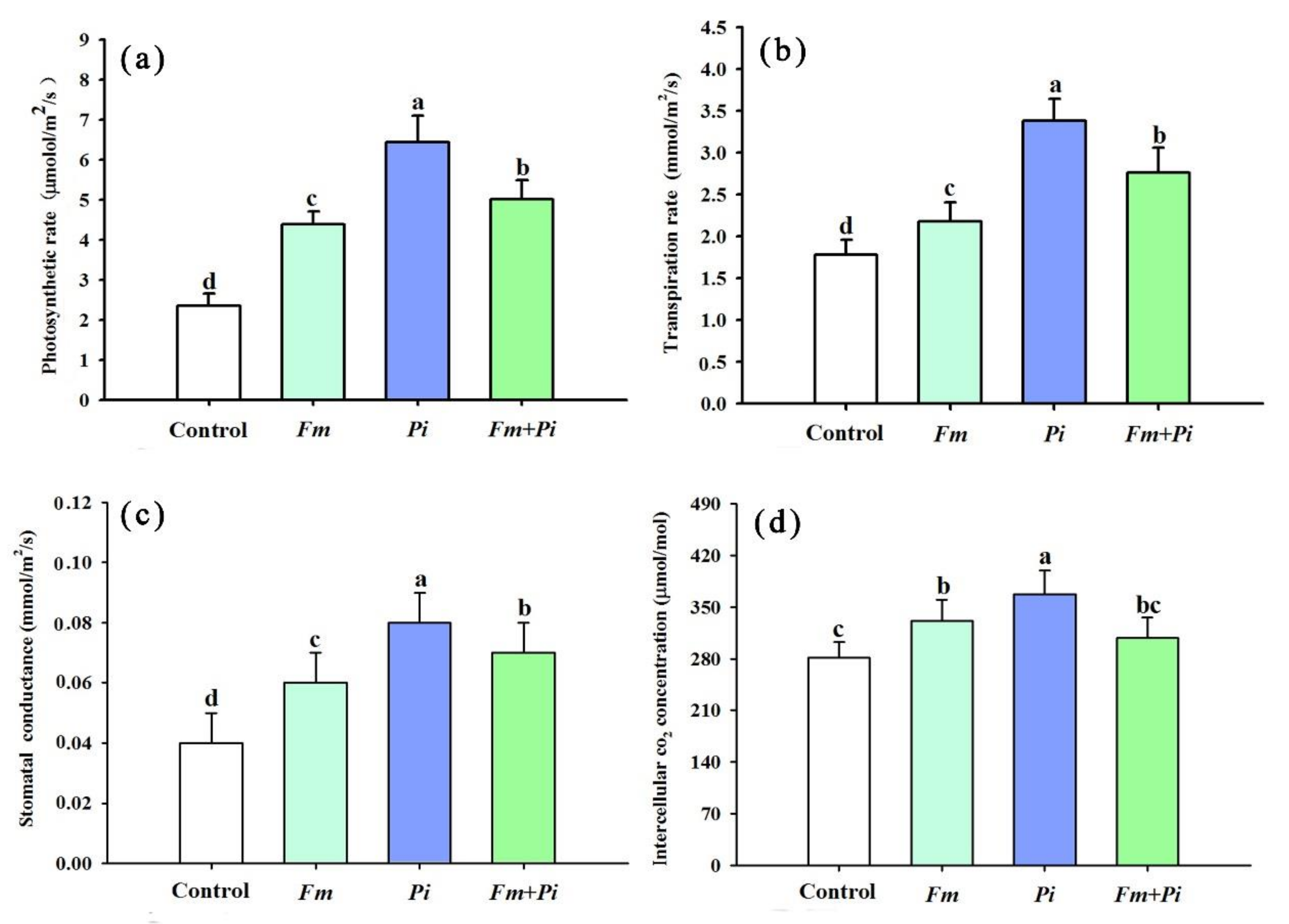
3.3. Changes in Leaf Gas Exchange
3.4. Changes in Leaf Medicinal Components
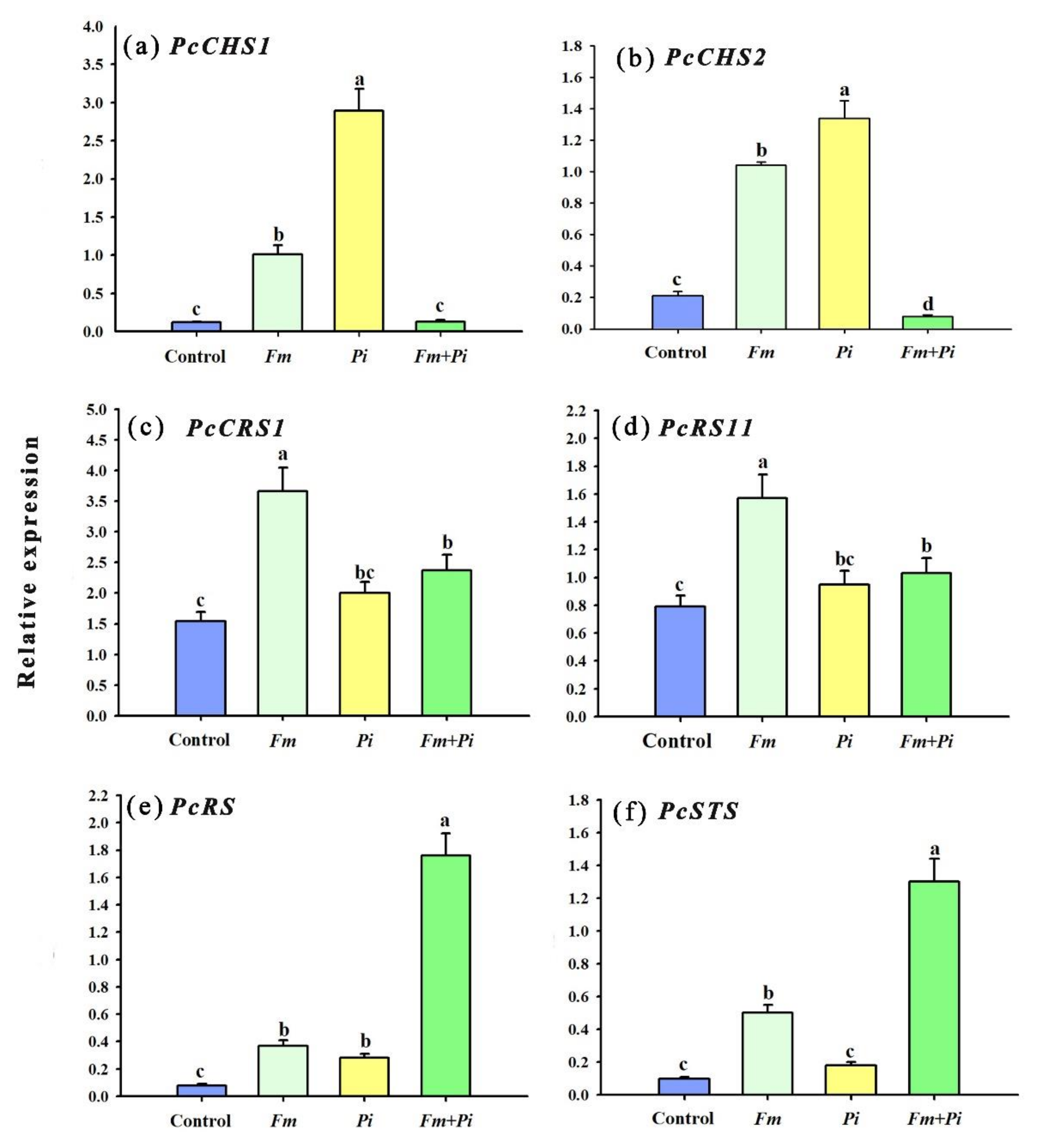
3.5. Changes in Expression of Associated Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, H.T.; Ding, S.L.; Lin, H.S. Pharmacological research progress of Polygonum cuspidatum. China J. Chin. Mat. Med. 2013, 38, 651–653. [Google Scholar]
- Pei, W.; Qin, R.X.; Li, X.L.; Zhou, H. Botany, phytochemistry, pharmacology, and potential application of Polygonum cuspidatum Sieb. et Zucc.: A review. J. Ethnopharmacol. 2013, 148, 729–745. [Google Scholar]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.; Namkoong, K.; Shin, M.; Park, J.; Yang, E.; Ihm, J. Cardiovascular protective effects and clinical applications of resveratrol. J. Med. Food. 2017, 20, 323. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.X.; Wang, S.S.; Chen, S.J.; Wang, Y.; Li, J.; Chang, Y.X. Research development on chemical composition and pharmacology of Polygoni cuspidati Rhizoma et Radix. Chin. Tradit. Herb. Drugs 2022, 53, 1264–1276. [Google Scholar]
- Ludwig-Müller, J. Plants and endophytes: Equal partners in secondary metabolite production? Biotechnol. Lett. 2015, 37, 1325–1334. [Google Scholar] [CrossRef]
- Cheng, S.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Elucidating the mechanisms underlying enhanced drought tolerance in plants mediated by arbuscular mycorrhizal fungi. Front. Microbiol. 2021, 12, 809473. [Google Scholar] [CrossRef]
- Wu, Q.S.; He, J.D.; Srivastava, A.K.; Zou, Y.N.; Kuča, K. Mycorrhizas enhance drought tolerance of citrus by altering root fatty acid compositions and their saturation levels. Tree Physiol. 2019, 39, 1149–1158. [Google Scholar] [CrossRef]
- Zhou, N.; Ding, B.; Feng, Y.; Qi, W.H.; Zhang, H.; Guo, D.Q.; Xiang, J. Effects of mycorrhizal colonization and medicine quality of Paris polyphylla var. yunnanensis inoculated by different foreign AM fungi species. China J. Chin. Mat. Med. 2015, 40, 3158–3167. [Google Scholar]
- Mandal, S.; Upadhyay, S.; Wajid, S.; Ram, M.; Jain, D.C.; Singh, V.P.; Abdin, M.Z.; Kapoor, R. Arbuscular mycorrhiza increase artemisinin accumulation in Artemisia annua by higher expression of key biosynthesis genes via enhanced jasmonic acid levels. Mycorrhiza 2015, 25, 345–357. [Google Scholar] [CrossRef]
- Khaosaad, T.; Vierheilig, H.; Nell, M. Arbuscular mycorrhiza alter the concentration of essential oils in oregano (Origanum sp., Lamiaceae). Mycorrhiza 2006, 16, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Hao, Z.; Zhou, X. Arbuscular mycorrhiza facilitates the accumulation of glycyrrhizin and liquiritin in Glycyrrhiza uralensis under drought stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.F.; Xie, M.M.; Li, Y.; Liu, B.Y.; Liu, C.Y.; Wu, Q.S.; Kuča, K. Effects of field inoculation with arbuscular mycorrhizal fungi and endophytic fungi on fruit quality and soil properties of Newhall navel orange. Appl. Soil Ecol. 2022, 170, 104308. [Google Scholar] [CrossRef]
- Waller, F.; Achatz, B.; Baltruschat, H. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagde, U.S.; Prasad, R.; Varma, A. Impact of culture filtrate of Piriformospora indica on biomass and biosynthesis of active ingredient aristolochic acid in Aristolochia elegans Mart. Int. J. Biol. 2014, 6, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Agrawal, V. Marked enhancement in the artemisinin content and biomass productivity in Artemisia annua L. shoots co-cultivated with Piriformospora indica. World J. Microbiol. Biotechnol. 2013, 29, 1133–1138. [Google Scholar] [CrossRef]
- Bagde, U.S.; Prasad, R.; Varma, A. Influence of culture filtrate of Piriformospora indica on growth and yield of seed oil in Helianthus annus. Symbiosis 2011, 53, 83–88. [Google Scholar] [CrossRef]
- Shayan, M.; Jagriti, S.; Kushwaha, A.S.; KaPil, M.; Jai, S.; Nidhi, A. Endophytic fungi Piriformospora indica mediated protection of host from arsenic toxicity. Front. Microbiol. 2017, 8, 754. [Google Scholar]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Meng, L.L.; Liu, R.C.; Yang, L.; Zou, Y.N.; Srivastava, A.K.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Giri, B.; Wu, Q.S. The change in fatty acids and sugars reveals the association between trifoliate orange and endophytic fungi. J. Fungi 2021, 7, 716. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophyll a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of proteins utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sun, R.-T.; Feng, X.-C.; Zhang, Z.Z.; Zhou, N.; Feng, H.D.; Liu, Y.M.; Hashem, A.; Al-Arjani, A.B.F.; Abd_Allah, E.F.; Wu, Q.S. Root endophytic fungi regulate changes in sugar and medicinal compositions of Polygonum cuspidatum. Front. Plant Sci. 2022, 13, 818909. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schuck, S.; Camehl, I.; Gilardoni, P.A. HSPRO controls early Nicotiana attenuata seedling growth during interaction with the fungus Piriformospora indica. Plant Physiol. 2012, 160, 929–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltruschat, H.; Fodor, J.; Harrach, B.D. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Eugui, D.; Abril-Urías, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Yaghoubian, Y.; Goltapeh, E.M.; Pirdashti, H.; Esfandiari, E.; Feiziasl, V.; Dolatabadi, H.K.; Varma, A.; Hassim, M.H. Effect of Glomus mosseae and Piriformospora indica on growth and antioxidant defense responses of wheat plants under drought stress. Agric. Res. 2014, 3, 229–245. [Google Scholar] [CrossRef]
- Sun, R.T.; Zhang, Z.Z.; Zhou, N.; Srivastava, A.K.; Kuča, K.; Abd_Allah, E.F.; Hashem, A.; Wu, Q.S. A review of the interaction of medicinal plants and arbuscular mycorrhizal fungi in the rhizosphere. Not. Bot. Horti Agrobot. 2021, 49, 12454. [Google Scholar] [CrossRef]
- Khalvandi, M.; Amerian, M.; Pirdashti, H.; Keramati, S. Does co-inoculation of mycorrhiza and Piriformospora indica fungi enhance the efficiency of chlorophyll fluorescence and essential oil composition in peppermint under irrigation with saline water from the Caspian Sea? PLoS ONE 2021, 16, e0254076. [Google Scholar]
- Sharma, P.; Kharkwal, A.C.; Abdin, M.Z.; Verma, A. Piriformospora indica improves micropropagation, growth and phytochemical content of Aloe vera L. plants. Symbiosis 2014, 64, 11–23. [Google Scholar] [CrossRef]
- Pandey, D.K.; Banik, R.M. The influence of dual inoculation with Glomus mossae and Azotobacter on growth and barbaloin content of Aloe vera. Am. Eurasian J. Sustain. Agric. 2009, 3, 703–714. [Google Scholar]
- Zahedi, H.S.; Jazayeri, S.; Ghiasvand, R.; Djalali, M.; Eshraghian, M.R. Effects of Polygonum cuspidatum containing resveratrol on inflammation in male professional basketball players. Int. J. Prev. Med. 2013, 4, 1–4. [Google Scholar]
- Bruisson, S.; Maillot, P.; Schellenbaum, P.; Walter, B.; Gindro, K.; Deglene-Benbrahim, L. Arbuscular mycorrhizal symbiosis stimulates key genes of the phenylpropanoid biosynthesis and stilbenoid production in grapevine leaves in response to downy mildew and grey mould infection. Phytochemistry 2016, 131, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Kovářová, M.; Frantík, T.; Koblihová, H.; Bartůňková, K.; Vosátka, M. Effect of clone selection, nitrogen supply, leaf damage and mycorrhizal fungi on stilbene and emodin production in knotweed. BMC Plant Biol. 2011, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Wang, D. Efficiency of Inoculation with Arbuscular Mycorrhizal Fungi to Rumex gmelini Turcz Seedlings. Master’s Thesis, Heilongjiang University of Chinese Medicine, Harbin, China, 2008. [Google Scholar]
- Burns, J.; Yokota, T.; Ashihara, H. Plant foods and herbal sources of resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Park, E.J.; Pezzuto, J.M. The pharmacology of resveratrol in animals and humans. Biochim. Biophys. Acta 2015, 1852, 1071–1113. [Google Scholar] [CrossRef] [Green Version]
- De Maria, S.; Scognamiglio, I.; Lombardi, A.; Amodio, N.; Caraglia, M.; Cartenì, M.; Ravagnan, G.; Stiuso, P. Polydatin, a natural precursor of resveratrol, inducescell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med. 2013, 11, 264. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.J.; Shao, K.H.; Chan, M.T.; Cheng, C.P.; Yeh, K.W.; Oelmuller, R.; Wang, S.J. Piriformospora indica symbiosis improves water stress tolerance of rice through regulating stomata behavior and ROS scavenging systems. Plant Signal. Behav. 2020, 15, 2. [Google Scholar] [CrossRef]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Wang, S. Medical Plant Arbuscular Mycorrhizal Fungi Resource and Inoculation Effect of Arbuscular Mycorrhizal Fungi on Ophopogon japonicus. Master’s Thesis, Northwest A&F University, Yangling, China, 2008. [Google Scholar]
- Zhang, Y.T.; Huang, X.; Chen, Y.Z.; Li, J.D.; Yu, K. Chemical constituents and their biosynthesis mechanisms of Polygonum cuspidatum. China J. Chin. Mat. Med. 2020, 45, 4364–4372. [Google Scholar]
- Lei, Y.M. Cloning and Expression of the Resveratrol Synthase Gene from Polygonum cuspidatum. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2007. [Google Scholar]
- Li, H.L.; Xu, L.F.; Li, Z.W.; Zhao, S.X.; Guo, D.Q.; Rui, L.; Zhou, N. Mycorrhizas affect polyphyllin accumulation of Paris polyphylla var. yunnanensis through promoting PpSE expression. Phyton Int. J. Exp. Bot. 2021, 90, 1535–1547. [Google Scholar] [CrossRef]
- Kumar, V.; Sahai, V.; Bisaria, V.S. Effect of Piriformospora indica on enhanced biosynthesis of anticancer drug, po-dophyllotoxin, in plant cell cultures of Linum album. In Piriformospora Indica; Varma, A., Kost, G., Oelmüller, R., Eds.; Springer: Heidelberg/Berlin, Germany, 2013; pp. 119–137. [Google Scholar]
- Flores-Sanchez, I.J.; Verpoorte, R. Plant polyketide synthases: A fascinating group of enzymes. Plant Physiol. Biochem. 2009, 47, 167–174. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene ID | Forward Sequence (5′→3′) | Reverse Sequence (5′→3′) |
|---|---|---|---|
| PcCHS1 | AB019030. 1 | GTAGCTGCCGAATCTTCTACTG | TGTCGTAGCATCGTCCTTTG |
| PcCHS2 | EU647246. 1 | GAAGCTTAAGGCGACTAGACAA | CAACCGACTTCTTCCTCATCTC |
| PcCRS1 | DQ459350. 1 | TGAGCGAGTACGGGAATTTG | CCTTCTCCAGTCGTCTTCTTAC |
| PcRS11 | EF117977. 1 | GATGAGATGATGAAGGCACAAAC | GGAAGTAGAAGTCGGGAAAGTC |
| PcRS | DQ900615. 1 | GAGATGACGAAGGCACTAACA | GGAAGTAGAAGTCGGGAAAGTC |
| PcSTS | EU647245. 1 | GAAGAGATGATGAAGGCACAAAC | GGAAGTAGAAGTCGGGAAAGTC |
| PcActin | MK288156. 1 | TACAATGAGCTTCGGGTTGC | GCTCTTTGCAGTTTCCAGCT |
| Treatments | Chrysophanol | Emodin | Physcion | Polydatin | Resveratrol |
|---|---|---|---|---|---|
| Control | 0.28 ± 0.03 ab | 0.62 ± 0.05 a | 0.10 ± 0.01 b | 1.01 ± 0.11 c | 0.30 ± 0.04 d |
| Fm | 0.32 ± 0.02 a | 0.55 ± 0.03 b | 0.07 ± 0.01 c | 1.54 ± 0.14 b | 0.96 ± 0.09 a |
| Pi | 0.26 ± 0.02 b | 0.57 ± 0.04 ab | 0.13 ± 0.01 a | 2.52 ± 0.25 a | 0.71 ± 0.08 b |
| Fm + Pi | 0.27 ± 0.02 b | 0.42 ± 0.04 c | 0.07 ± 0.01 c | 2.36 ± 0.20 a | 0.54 ± 0.06 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, R.-T.; Zhang, Z.-Z.; Feng, X.-C.; Zhou, N.; Feng, H.-D.; Liu, Y.-M.; Harsonowati, W.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Endophytic Fungi Accelerate Leaf Physiological Activity and Resveratrol Accumulation in Polygonum cuspidatum by Up-Regulating Expression of Associated Genes. Agronomy 2022, 12, 1220. https://doi.org/10.3390/agronomy12051220
Sun R-T, Zhang Z-Z, Feng X-C, Zhou N, Feng H-D, Liu Y-M, Harsonowati W, Hashem A, Abd_Allah EF, Wu Q-S. Endophytic Fungi Accelerate Leaf Physiological Activity and Resveratrol Accumulation in Polygonum cuspidatum by Up-Regulating Expression of Associated Genes. Agronomy. 2022; 12(5):1220. https://doi.org/10.3390/agronomy12051220
Chicago/Turabian StyleSun, Rui-Ting, Ze-Zhi Zhang, Xiang-Cao Feng, Nong Zhou, Hai-Dong Feng, Yi-Mei Liu, Wiwiek Harsonowati, Abeer Hashem, Elsayed Fathi Abd_Allah, and Qiang-Sheng Wu. 2022. "Endophytic Fungi Accelerate Leaf Physiological Activity and Resveratrol Accumulation in Polygonum cuspidatum by Up-Regulating Expression of Associated Genes" Agronomy 12, no. 5: 1220. https://doi.org/10.3390/agronomy12051220