
Field Evaluation of CRISPR-Driven Jointless Pedicel Fresh-Market Tomatoes



Horticultural Sciences Department, University of Florida, Gainesville, FL 32611, USA
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(10), 1957; https://doi.org/10.3390/agronomy11101957
Submission received: 10 September 2021
/
Accepted: 27 September 2021
/
Published: 29 September 2021
(This article belongs to the Special Issue Shaping the Agronomy of Tomorrow: GMOs, CRISPR and New Breeding Techniques for Crop Improvement)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Clustered regularly interspaced short palindromic repeats (CRISPR)-driven traits are commonly introduced into tomato (Solanum lycopersicum) in less than a year. Although CRISPR technology is highly suitable for rapid incorporation of new traits into plants with superior breeding backgrounds, the usefulness of these traits must be demonstrated prior to large-scale distribution of any new variety possessing them. The jointless pedicel trait is crucial for stem-free harvest of tomato fruits and is therefore an essential requirement for developing low-labor tomato varieties. We aimed to evaluate the effect of CRISPR-driven jointless pedicel trait on the yield and horticultural traits of fresh-market tomatoes. We conducted field trials during three consecutive seasons using three related genotypes of two elite fresh-market tomatoes, namely a jointed pedicel tomato, a CRISPR-driven jointless pedicel mutant of the jointed pedicel tomato, and a conventionally-bred jointless pedicel near isogenic line (NIL) to the jointed pedicel tomato. Field evaluations confirmed that the total and medium-, large-, or extra-large-size fruit yields of the CRISPR-driven mutants were not statistically different for those corresponding to their backgrounds and that the fruit yield stratified by fruit size might be varied under different growth conditions. We did not observe any negative correlation between the genotype and the fruit/abscission joint detachment force for any of the genotypes under study.
1. Introduction
Field production of U.S. fresh-market tomatoes continues to rely heavily on manual labor, especially for several production practices including harvesting and tying [1,2,3]. However, rapidly increasing labor costs and recent trends regarding the uncertainty associated with trained workers [2,4,5] have forced the fresh-market tomato industry to seek a change toward to mechanical harvest in order to reduce dependence on labor. A transition to tomato varieties specifically designed for ground culture is a potential solution to this challenge. Ultimately, the development of machine-harvested fresh-market tomatoes is one of the few viable solutions available for the achievement for long-term economic viability in the fresh-market tomato industry.
One essential aspect of plant architecture to consider before such low-labor tomatoes can be cultivated is that the fruit must be removed such that no stem is attached (i.e., without allowing the calyx and part of the pedicel to remain attached to the fruit). An attached stem may cause scratches/punctures in the fruit skin, thereby rendering the fruit unmarketable. To achieve a stem-free fruit harvest, the tomato jointless2 gene (j2), which creates the jointless pedicel trait [6], has been already utilized extensively by tomato breeders, especially for the processing tomato market; almost all contemporary processing tomato varieties in the U.S. have jointless pedicels and are mechanically harvested. Mechanical harvesting of fresh-market tomatoes will necessitate jointless pedicel varieties but little progress has been made to date in this regard [7].
The CRISPR-CRISPR-associated protein 9 (Cas9) gene-editing system (hereafter, CRISPR) provides a useful tool for generating loss-of-function mutations in specific genes [8]. The ability to introgress a trait of interest into an elite tomato breeding line in a cost- or time-effective manner is the main advantage of CRISPR over existing conventional breeding methods [9]. CRISPR-driven traits can typically be introduced into tomato rather quickly, in less than a year. Although CRISPR-driven traits can be highly beneficial for rapidly incorporating new traits into breeding backgrounds, the usefulness of these traits must be demonstrated prior to their large-scale dissemination of such varieties to growers. Soyk et al. developed a CRISPR for targeting J2 [10]. This system has been used to generate transgene-free jointless pedicel mutants of fresh-market tomatoes, but the yields and horticultural traits of such CRISPR-driven jointless pedicel tomatoes, which are directly relevant to tomato variety-development programs, have not been evaluated.
Therefore, our objective was to evaluate the effect of the CRISPR-driven jointless pedicel trait on the yield and horticultural traits for two different types of elite fresh-market tomatoes. We performed this analysis to evaluate the effect of loss-of-function mutations in a single gene J2 (i.e., CRISPR-driven J2 gene-edited mutant) on field performance. If field performance is not adversely affected by the CRISPR-driven j2 gene, this approach may be widely adopted for the rapid conversion of fresh-market tomato varieties from jointed pedicel to jointless pedicel.
2. Materials and Methods
2.1. Plant Material
Fla. 7946 [11] and Fla. 8735, used in this study are elite modern tomato germplasm releases that were registered and stored by the tomato breeding program of the Institute of Food and Agricultural Sciences (IFAS) at the University of Florida (UF) (UF/IFAS [12]). Both tomatoes were jointed (J2/J2; [6]), determinate (sp/sp, homozygous at the self-pruning locus [13]) fresh-market inbreds. Fla. 7946 and Fla. 8735 are the flat and globe fruit types, respectively [11,14]. In addition, the CRISPR-driven jointless pedicel mutant (homozygous transgenic tomato line developed by CRISPR targeting the J2 gene, Solyc12g038510) and conventionally-bred (i.e., via backcross) NIL of both Fla. 7946 and Fla. 8735 were obtained from the UF/IFAS tomato breeding program; hereafter, CRISPR-driven j2 mutants are referred to as Fla. 7946j2cr and Fla. 8735j2cr and conventionally-bred NILs as Fla. 7946j2nil and Fla. 8735j2nil. Both of these mutants have been reported previously [15].
2.2. Phenotypic Data Collection
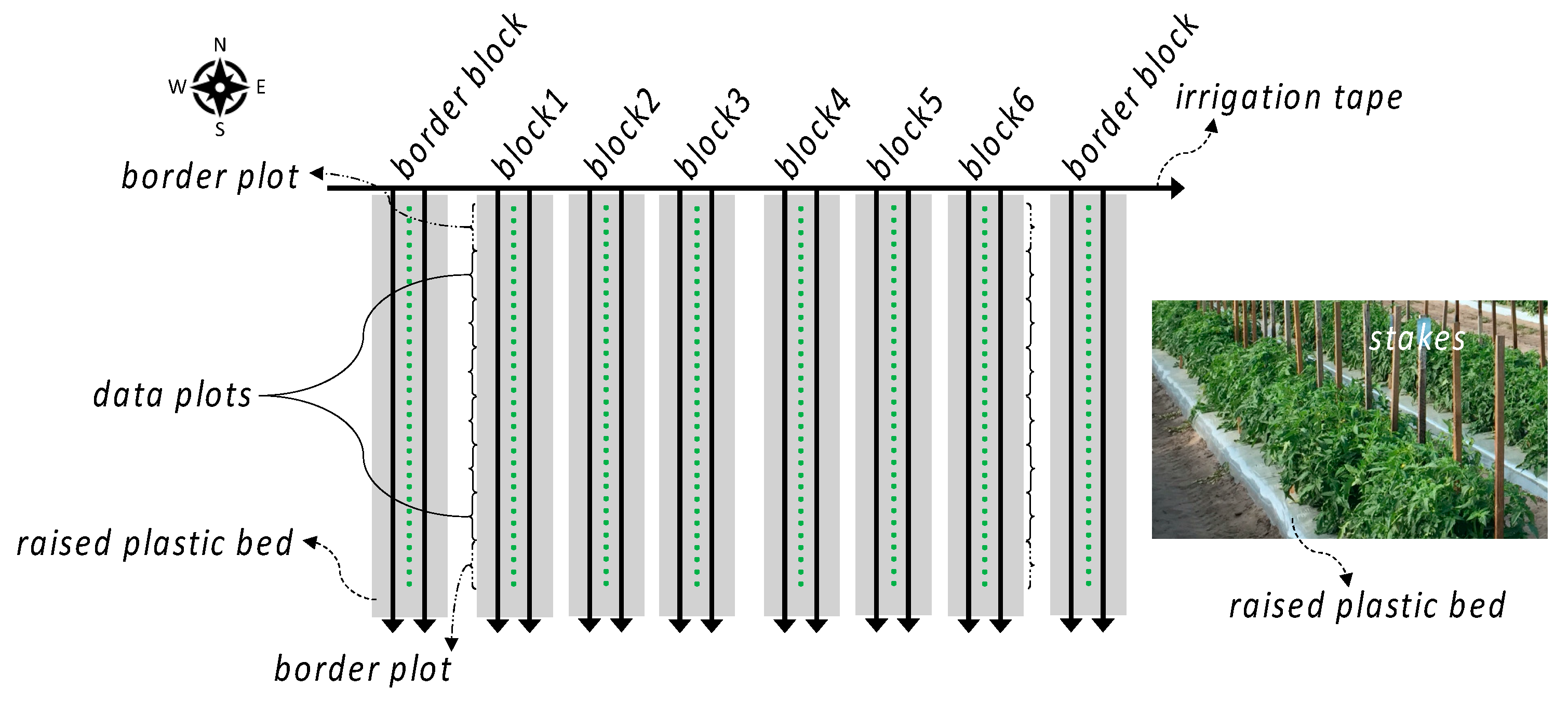
Plants were grown as previously described (“Phenotype analysis” section in Lee et al. [16]. Field trials were conducted in three consecutive growing seasons during 2020 and 2021 at the Gulf Coast Research and Education Center (Wimauma, FL, USA) of the University of Florida, where conditions are representative of typical field fresh-market tomato production environments in the Southeastern U.S. During the spring of 2020, seed sowing in the greenhouse (S), seedling transplanting to the field (T), and fruit harvest (H) were performed on 22 January, 4 March, and 23 May, respectively. For the second growing cycle during the fall of 2020, S, T, and H were performed on 3 August, 16 September, and 8 December. Finally, during the spring of 2021, S was performed on 15 January, T on 1 March, and H on 1 June. In each growing season, the experiment was established according to a randomized complete block design (Figure 1). Two rectilinear irrigation tapes buried underneath the plastic supplied water and nutrients to each test field (each block). Six blocks were included in each season. Each block was split into eight plots; border plants were assigned to the first and last plots (border plots), and plants from which data were to be collected were assigned to the remaining six inner plots (data plots). 36 plots of six different genotypes (each plot of four plants) were randomly assigned, and six different genotypes were assigned to each block.
All fruits (>0.5 cm in diameter) developed in each seasonal trial were harvested on a single harvesting date, regardless of fruit weight, quality (e.g., irrespective of whether the fruits had defects such as cracks), or color. Separate preliminary trials showed that variations associated with fruit collection and fruit color/size during manual harvest (i.e., harvest of fruits on the vine with the support of stakes and ties, data not shown) were primarily because of the typical morphology of the U.S. fresh-market tomatoes (i.e., presence of dense vegetative tissues and woody lateral branches at harvest) and the subjectivity of the individual(s) conducting the harvest, as they were not blinded to allocation (knowledge of plant materials) or to the outcome assessment during experiments. For this reason, no information was provided to avoid bias resulting from predetermining plant materials during harvest; hence, multiple harvest steps were conducted. First, the stakes and ties were removed from the field before harvest. The base of the main stem in each plant was cut once the plants were stake/tie-free. Each plant was then delivered to a separate place, where at least two individuals collected fruits from each plant.
Fruit color (green vs. breakers [17]) was visually determined on harvest day. Fruits were sorted by size (four size classes; any fruit smaller than medium size, medium, large, and extra-large sizes) using the USDA Tomato Sizer [17], and descriptions of medium, large, and extra-large sizes were in accordance with the shipping point and market inspection instructions for tomatoes [18]. For each genotype per plot, the fruit weight and number were calculated from the average values for weight and number of fruits from the plants in each plot, respectively. Days to first flower has been described as the number of days from sowing to the first full bloom (i.e., when petals create a 180° angle). Days to first flower and fruit color data were collected by the same individual throughout the three seasons.
Fruit/abscission joint detachment force (Newton, N) was measured on harvest day using a FORCE ONE™ FDIX 100 (Wagner, Greenwich, CT, USA) according to the manufacturer instructions. The force required to detach green or red fruits (Tomato Color Chart, [17]) from the abscission joint of the pedicel (hereafter, abscission joint detachment force), and the force required to detach fruits from the fruit stalk (hereafter, fruit detachment force) were measured for genotypes with the jointed pedicel. Only the fruit detachment force was measured for genotypes with the jointless pedicel. The average weights of fruits collected from recurrent parents in 2020 were 165.0 ± 17.6 g (mean ± 95% confidence interval) and 214.6 ± 14.0 g of green or red Fla. 7946 fruit, respectively, and 217.8 ± 39.6 g and 232.4 ± 37.5 g of green or red Fla. 8735 fruit, respectively. In 2021, the average weights of fruits from recurrent parents were 205.0 ± 21.3 g and 193.3 ± 24.5 g of green or red Fla. 7946 fruit, respectively, and 225.3 ± 31.5 g and 213.1 ± 18.4 g of green or red Fla. 8735 fruit, respectively.
The statistical significance level (* p < 0.05, ** p < 0.01, *** p < 0.001) of any two genotype mean values was based on the results of one-way analysis of variance (ANOVA) in conjunction with a two-tailed Tukey’s HSD multiple comparison test, or the Welch’s test. Error bars indicate the 95% confidence intervals for the six blocks.
3. Results and Discussion
Understanding the relationship at the phenotypic level between a causative gene and the corresponding horticultural trait(s) is of critical importance to applied tomato geneticists and breeders. The inflorescence architectures of jointed pedicel fresh-market tomatoes and their CRISPR-driven jointless pedicel mutants have been previously analyzed [15]. However, the effect of CRISPR-driven jointless pedicel mutation(s) on the yield and horticultural traits, such as the fruit/abscission joint detachment force, has not been reported to date. In this study, three related genotypes—a jointed pedicel tomato (parent), a CRISPR-driven mutant of the jointed tomato, a jointless pedicel NIL to the jointed pedicel recurrent parent—were evaluated using two different types of fresh-market tomatoes in an open field located in the Southeastern U.S., where these tomatoes were originally bred.
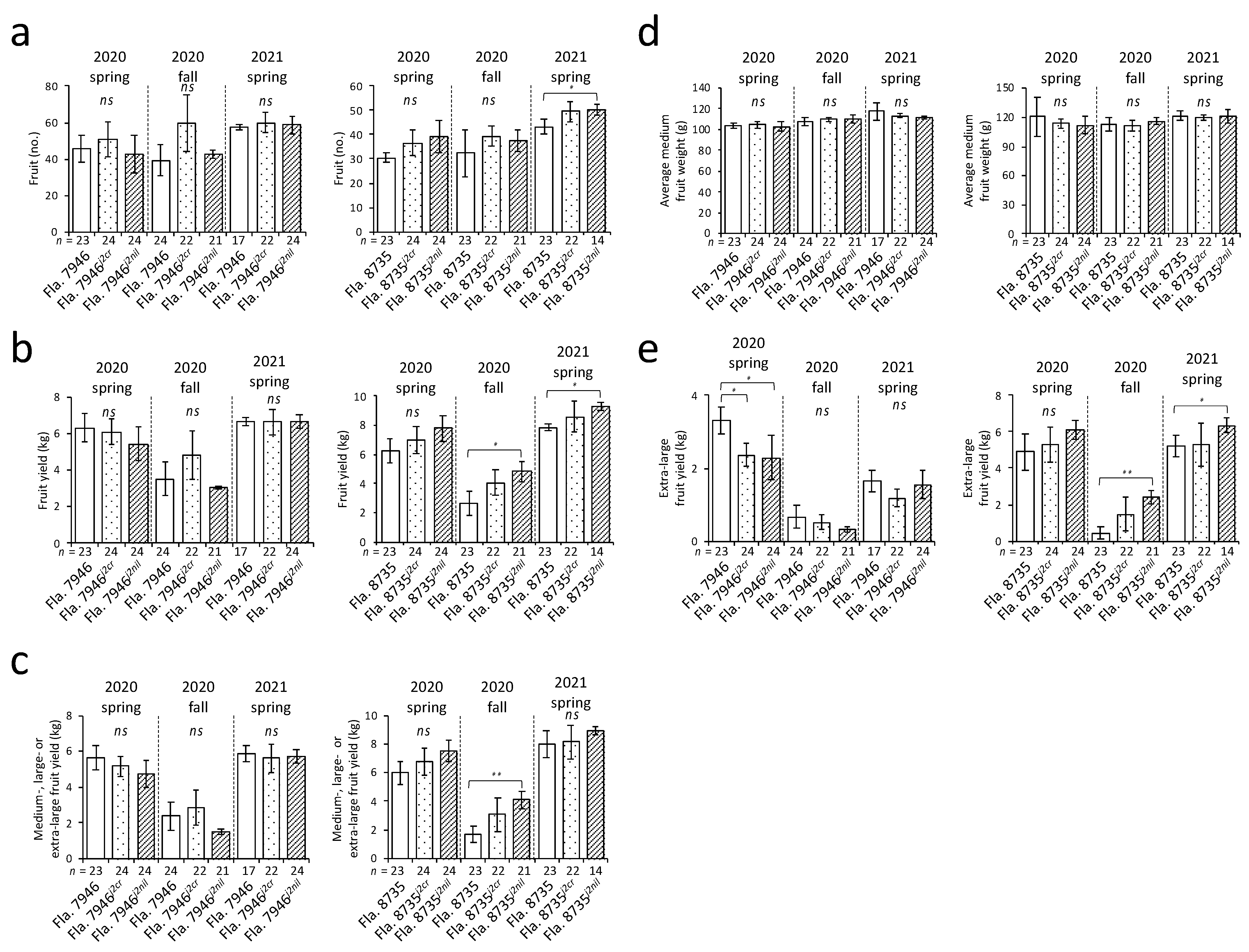
There were no significant differences in the total number of fruits among genotypes for either tomato fruit type (flat or globe) across the three seasons over which this study was conducted (except a comparison of Fla. 8735 and Fla. 8735j2nil in 2021; Figure 2a). However, during yield evaluation, genotype effects on yield were not clear (i.e., lack of consistency in statistical significance across three seasons), especially on fruit yield based on fruit-size classification. Importantly, the differences were less clear in NILs than in other genotypes. However, in the absence of a completely known pedigree for tomato materials, results remain speculative whether potential background genes that will impact yield could cause the effect observed or compensate for the effect of the j2 gene. The results showed that there were no differences in the total fruit yield across the three seasons between the jointed pedicel tomato and its CRISPR-driven mutant in either fruit type (Figure 2b). Furthermore, there were no differences in the medium-, large-, or extra-large size fruit yield between the jointed pedicel tomato and the CRISPR-driven mutant in either fruit type (Figure 2c). We did not observe differences in the average total weight of medium-size fruits among all the genotypes tested in either fruit type (Figure 2d). Fruit size, especially large fruit size, is a particularly important trait for fresh-market tomato growers, especially in the U.S., because of the market demand (fruit weight and size are analogous to yield, since fresh-market tomato fruit can be sold in packages that meet a net standard weight and fruit-size requirements as USDA market standards) [7,18]. Furthermore, there were no differences in yield of Fla. 8735j2cr and the corresponding jointed pedicel background Fla. 8735 (globe fruit type); however, Fla. 7946j2cr (flat fruit type) showed a reduction in extra-large-size fruit yield in the spring of 2020 (Figure 2e). Because the yield of tomatoes with the globe fruit type is often higher than that of tomatoes with the flat fruit type [14] and the environmental effects on fruit quality under many growth conditions are observed (J.W. Scott, personnel communication), it is possible that changes occurring during fruit development in the flat fruit type tomatoes may result in variations in fruit yield.
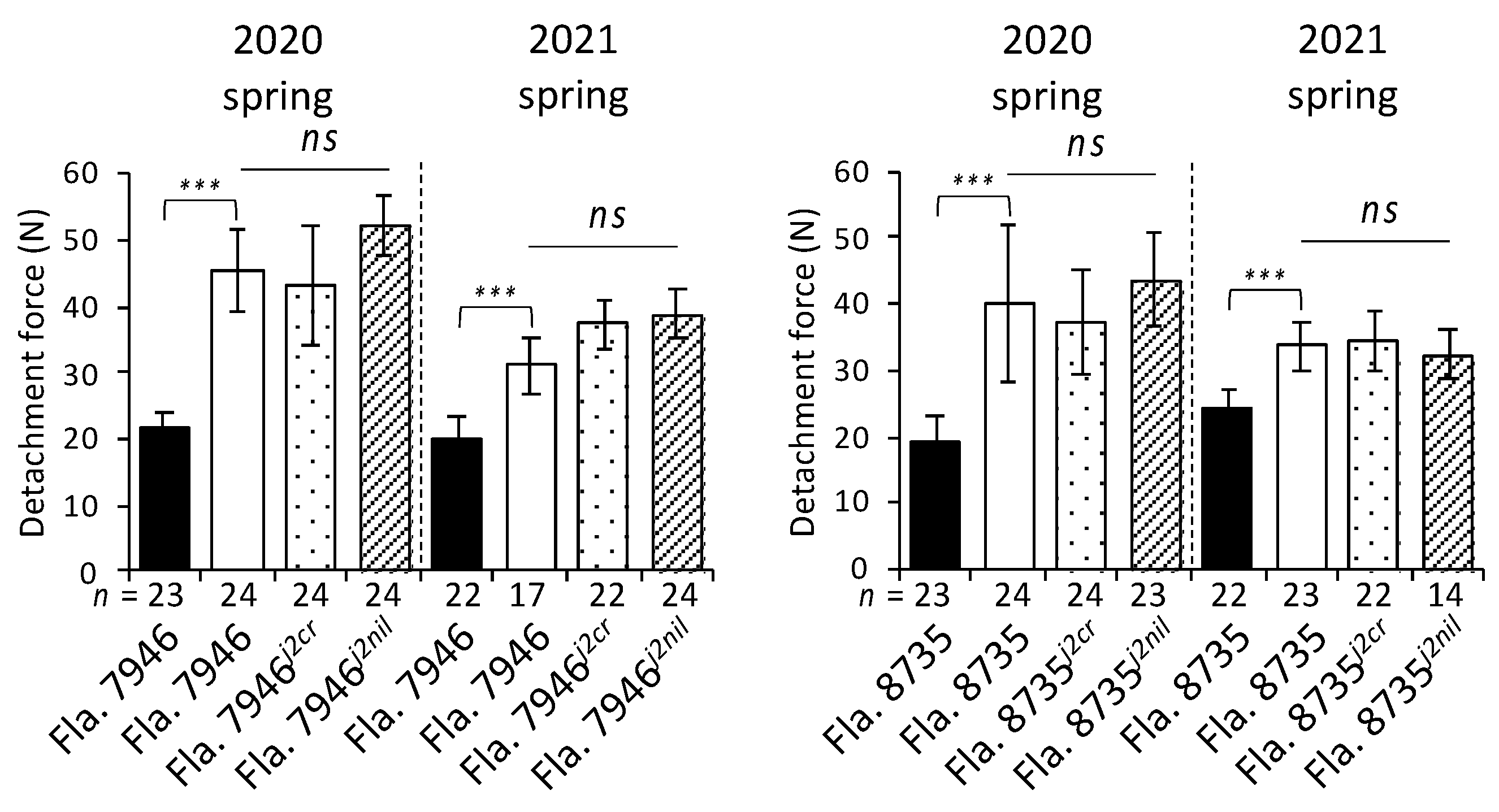
The fruit detachment force was higher than the abscission joint detachment force at all instances (Figure 3), indicating that the stem (part of the pedicel) was mostly attached at the time of harvest, unless the stem was manually removed during the harvesting process. No negative correlation (i.e., higher detachment force required in CRISPR-driven mutants) was observed between the genotype and fruit/abscission joint detachment force.
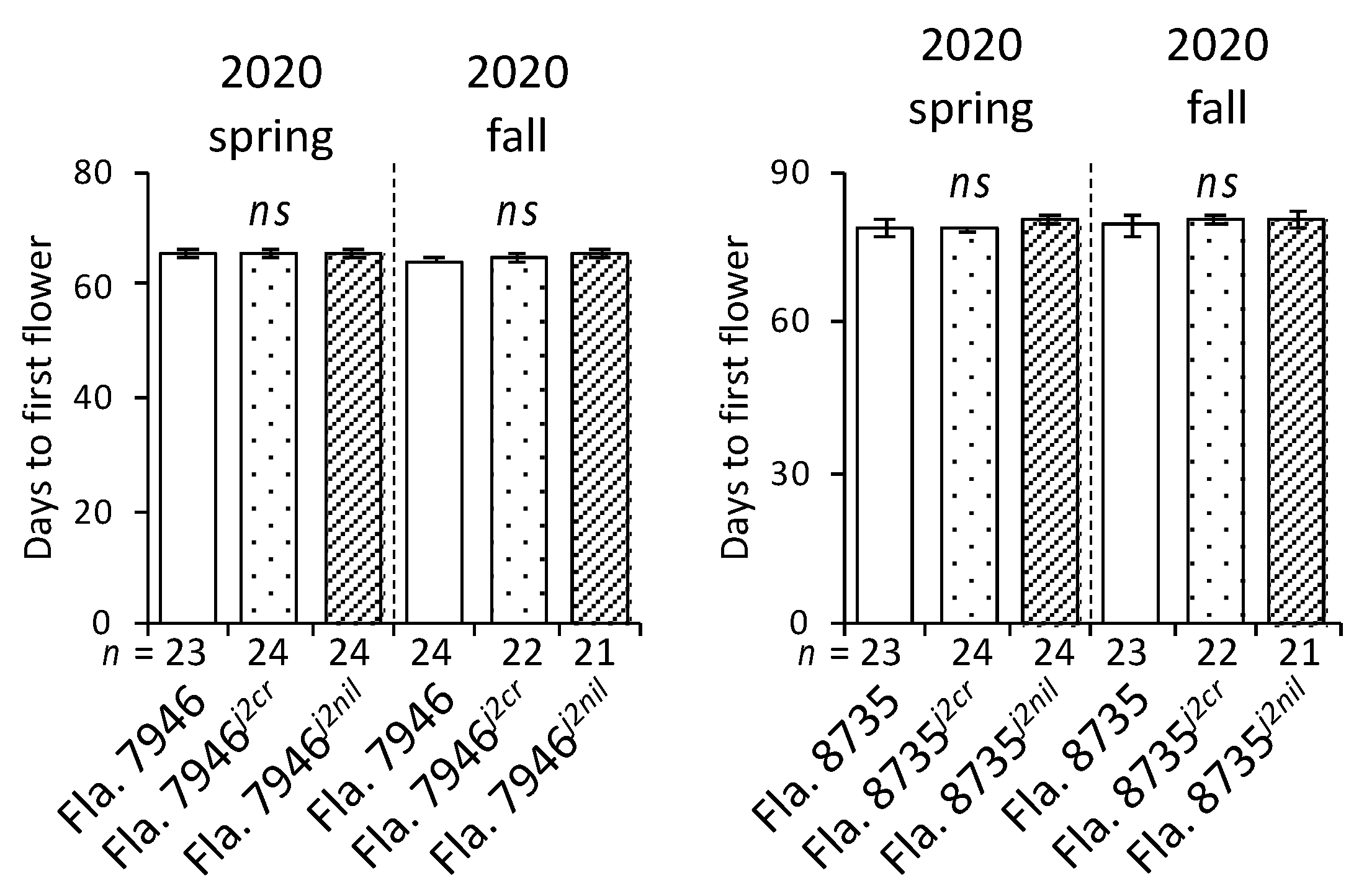
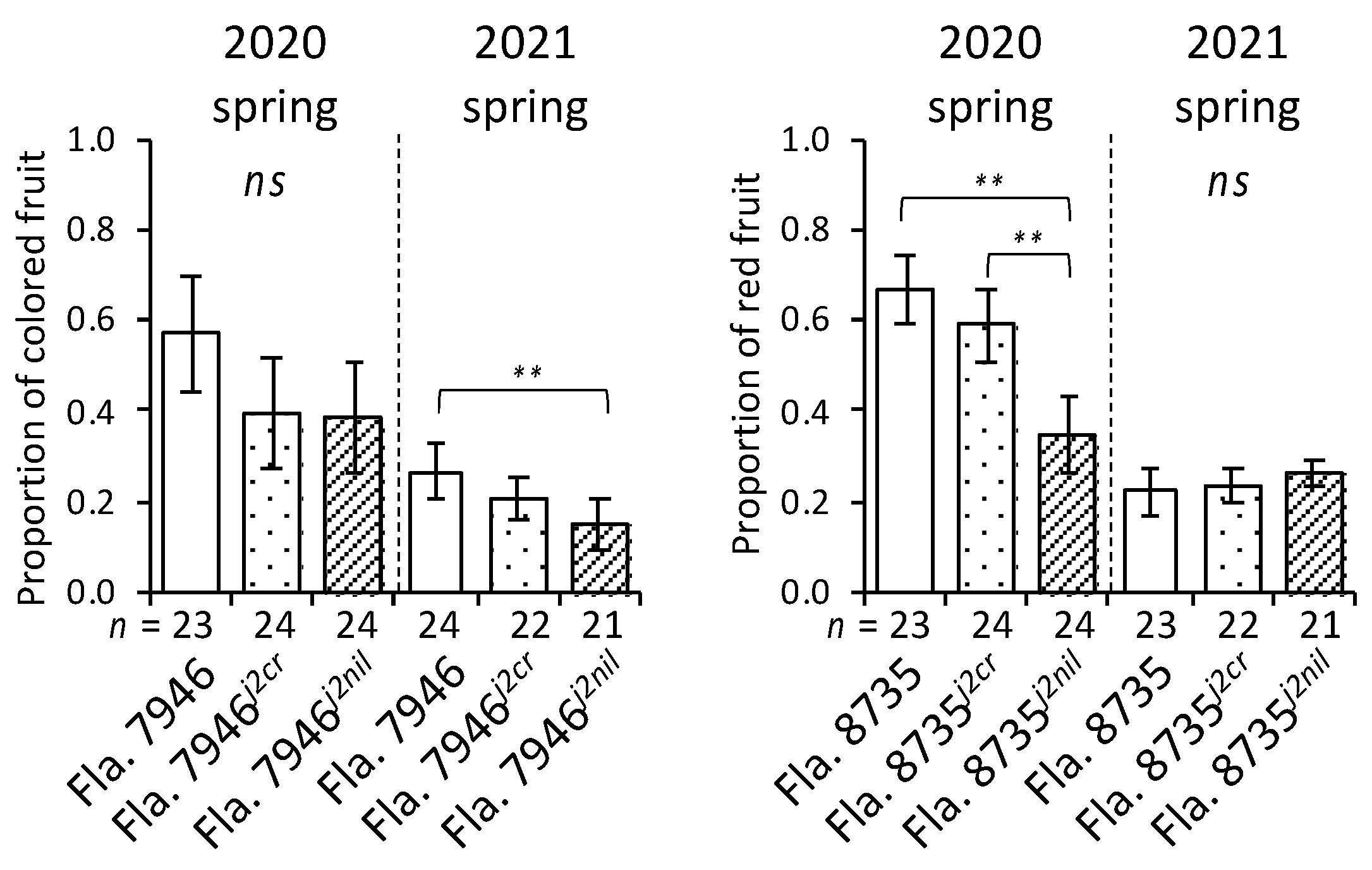
The flowering time and floral architecture of plants have been widely studied in plants [10,19,20], and multiple genes associated with fruit development have been identified [21,22]. Our results showed that there was no difference in days to first flower among the three genotypes in either fruit type (Figure 4), suggesting that j2 does not affect flowering time in this type of tomato. Additionally, the differences in the proportion of colored fruits were less clear in NILs than in other genotypes (Figure 5).
The potential benefits of the jointless pedicel trait are well known, and linked molecular markers for the causative gene of the trait already exist [23]. However, it is difficult to develop jointless pedicel tomato varieties that are as good as existing jointed pedicel tomato varieties because defects such as fruit-cracking, rough blossom scars, and off-shapes, are often more severe in the former under many growth conditions [24]. Because of the challenges associated with the development of jointless pedicel fresh-market tomato varieties using conventional breeding methods, it can be hypothesized that an alternative loss-of-function mutation(s) in the J2 gene (i.e., CRISPR-driven J2 gene-edited mutant) may enable the achievement of a (similar or identical) jointless pedicel phenotype, while bypassing the negative effects on fruit quality. This would be a feasible objective, given that CRISPR-driven J2 gene-edited fresh-market tomato germplasm resources are available. Unfortunately, we did not evaluate fruit quality. However, the objective measurement of the quality of tomatoes, especially field-grown fresh-market tomatoes, has required much time and labor to accomplish (i.e., measuring the depth and length of defects in the fruit skin using the right tool; e.g., score as growth cracks damage when cracks are not well healed, more than 1/8 inch in depth, individual radial cracks are more than 1/2 inch in length, the aggregate length of all radial cracks more than 1 inch measured from edge of stem scar per §51.1877 Classification of Defects [18]). In addition, the scoring guides for defects [18] can often conflict with market demands. The significant advances in high-throughput plant phenotyping and the development of algorithmic and computing system approaches, such as artificial intelligence (AI), would accelerate analysis and decision-making process in the near future. Additionally, an expansion of standardized phenotyping trials, whose data can be shared to provide a common overall principle, would enable tomato researchers to overcome the current limitations associated with fruit quality measurement, for the rapid incorporation of the jointless pedicel trait to research programs dealing with development of new and improvement varieties.
Author Contributions
The T.G.L. laboratory designed and conducted all experiments and data analysis for all figures. Conceptualization, T.G.L. & S.F.H.; methodology, T.G.L.; formal analysis, T.G.L.; investigation, T.G.L.; resources, S.F.H.; validation, T.G.L.; visualization, T.G.L.; writing, T.G.L.; funding acquisition, T.G.L.; supervision, T.G.L.; project administration, T.G.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Florida Tomato Committee (T.G.L.) and USDA National Institute of Food and Agriculture Hatch project FLA-GCC-005550 (T.G.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank members of the T.G.L. laboratory, especially Claudia Jose and Katherine Brown, for the management of the field.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Davis, J.M.; Estes, E.A. Spacing and pruning affect growth, yield, and economic returns of staked fresh-market tomatoes. J. Am. Soc. Hortic. Sci. 1993, 118, 719–725. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture. Tomatoes. 2016. Available online: www.ers.usda.gov/topics/crops/vegetables-pulses/tomatoes (accessed on 28 July 2021).
- VanSickle, J.; Smith, S.; McAvoy, E. Production Budget for Tomatoes Grown in Southwest Florida; Electronic Data Info. Source PE818; Univ. Florida, Inst. Food Agr. Sci.: Gainesville, FL, USA, 2009. [Google Scholar]
- California Tomato Growers Association. 2015. Available online: http://www.ctga.org (accessed on 28 July 2021).
- Florida Tomato Exchange. 2020. Available online: https://www.floridatomatoes.org (accessed on 28 July 2021).
- Butler, L. Inherited characters in the tomato. II. Jointless pedicel. J. Hered. 1936, 27, 25–26. [Google Scholar] [CrossRef]
- Scott, J.W.; Myers, J.R.; Boches, P.S. Classical genetics and traditional breeding. In Genetics, Genomics and Breeding of Tomato; Liedl, B.E., Labate, J.A., Stommel, J.R., Slade, A., Kole, C., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 37–73. [Google Scholar]
- Wang, H.; La Russa, M.; Qi, L.S. CRISPR/Cas9 in genome editing and beyond. Annu. Rev. Biochem. 2016, 85, 227–264. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.G. CRISPR: A Technical Breakthrough for Tomato Research; Univ. Florida, Inst. Food Agr. Sci.: Gainesville, FL, USA, 2018; p. HS1314. [Google Scholar]
- Soyk, S.; Lemmon, Z.H.; Oved, M.; Fisher, J.; Liberatore, K.L.; Park, S.J.; Goren, A.; Jiang, K.; Ramos, A.; van der Knaap, E.; et al. Bypassing negative epistasis on yield in tomato imposed by a domestication gene. Cell 2017, 169, 1142–1155.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.W. Fla. 7946 tomato breeding line resistant to Fusarium oxysporum f.sp. lycopersici Races 1, 2, and 3. HortScience 2004, 39, 440–441. [Google Scholar] [CrossRef]
- University of Florida, Institute of Food and Agricultural Sciences. UF/IFAS Tomato Breeding Program. 2021. Available online: https://tombreeding.ifas.ufl.edu (accessed on 28 July 2021).
- Barton, D.W.; Butler, L.; Jenkins, J.A.; Rick, C.M.; Young, P.A. Rules for nomenclature in tomato genetics (includes a list of known genes). J. Hered. 1955, 46, 22–76. [Google Scholar] [CrossRef]
- Sierra-Orozco, E.; Shekasteband, R.; Illa-Berenguer, E.; Snouffer, A.; van der Knaap, E.; Lee, T.G.; Hutton, S.F. Identification and characterization of GLOBE, a major gene controlling fruit shape and impacting fruit size and marketability in tomato. Hortic. Res. 2021, 8, 138. [Google Scholar] [CrossRef]
- Soyk, S.; Lemmon, Z.H.; Sedlazeck, F.J.; José, J.M.; Jiménez-Gómez, M.; Alonge, M.; Hutton, S.F.; Van Eck, J.; Schatz, M.C.; Lippman, Z.B. Duplication of a domestication locus neutralized a cryptic variant that caused a breeding barrier in tomato. Nat. Plants 2019, 5, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.G.; Hutton, S.F.; Shekasteband, R. Fine mapping of the brachytic locus on the tomato genome. J. Am. Soc. Hortic. Sci. 2018, 143, 239–247. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture. Equipment Catalog for Fresh and Processed Products Inspections. 2017. Available online: https://www.ams.usda.gov/sites/default/files/media/Fresh%20and%20Processed%20FV%20Products%20Inspections.pdf (accessed on 28 July 2021).
- U.S. Department of Agriculture. Shipping Point and Market Inspection Instructions for Tomatoes. 2005. Available online: https://www.ams.usda.gov/sites/default/files/media/Tomato_Inspection_Instructions%5B1%5D.pdf (accessed on 28 July 2021).
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Jung, C.; Pillen, K.; Staiger, D.; Coupland, G.; von Korff, M. Editorial: Recent advances in flowering time control. Front Plant Sci. 2011, 7, 2011. [Google Scholar] [CrossRef] [Green Version]
- Daccord, N.; Celton, J.M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Seymour, G.; Poole, M.; Manning, K.; King, G.J. Genetics and epigenetics of fruit development and ripening. Curr. Opin. Plant Biol. 2008, 11, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.G.; Shekasteband, R.; Hutton, S.F. Molecular markers to select for the j-2–mediated jointless pedicel in tomato. HortScience 2018, 53, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.W. Phenotyping of tomato for SolCAP and onward into the void. HortScience 2010, 45, 1314–1316. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
A depiction of block design. Black solid arrows indicate the direction of water/nutrients flow. Green dots represent tomato plants. Photo was taken on 9 April 2020 at the UF Gulf Coast Research and Education Center.
Figure 1.
A depiction of block design. Black solid arrows indicate the direction of water/nutrients flow. Green dots represent tomato plants. Photo was taken on 9 April 2020 at the UF Gulf Coast Research and Education Center.

Figure 2.
Evaluation of yield across three consecutive growing seasons in 2020 and 2021. (a) total number of fruits per plant; (b) fruit yield (kg) per plant; (c) yield of medium-, large-, or extra-large-size fruit per plant; (d) average weight (g) of medium-size fruit; (e) yield of extra-large fruit per plant. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistical difference (ANOVA, p > 0.05) among any of the genotypes. Statistically significant difference (* p < 0.05, ** p < 0.01 based on ANOVA in conjunction with a two-tailed Tukey’s HSD multiple comparison test) is indicated between genotypes. Error bars indicate 95% confidence intervals.
Figure 2.
Evaluation of yield across three consecutive growing seasons in 2020 and 2021. (a) total number of fruits per plant; (b) fruit yield (kg) per plant; (c) yield of medium-, large-, or extra-large-size fruit per plant; (d) average weight (g) of medium-size fruit; (e) yield of extra-large fruit per plant. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistical difference (ANOVA, p > 0.05) among any of the genotypes. Statistically significant difference (* p < 0.05, ** p < 0.01 based on ANOVA in conjunction with a two-tailed Tukey’s HSD multiple comparison test) is indicated between genotypes. Error bars indicate 95% confidence intervals.

Figure 3.
Evaluation of fruit/abscission joint detachment force during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The black filled and white bars indicate the values for abscission joint detachment force and fruit detachment force for jointed tomatoes, respectively; differences between means with *** p < 0.001 based on the Welch’s test are indicated. The n value represents the total number of fruits for each genotype evaluated during each season; ns indicates no statistically significant difference (ANOVA, p > 0.05) among any of the genotypes evaluated for the fruit detachment force. Error bars indicate 95% confidence intervals.
Figure 3.
Evaluation of fruit/abscission joint detachment force during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The black filled and white bars indicate the values for abscission joint detachment force and fruit detachment force for jointed tomatoes, respectively; differences between means with *** p < 0.001 based on the Welch’s test are indicated. The n value represents the total number of fruits for each genotype evaluated during each season; ns indicates no statistically significant difference (ANOVA, p > 0.05) among any of the genotypes evaluated for the fruit detachment force. Error bars indicate 95% confidence intervals.

Figure 4.
Evaluation of days to first flower during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistically significant difference (ANOVA, p > 0.05) among any of the genotypes. Error bars indicate 95% confidence intervals.
Figure 4.
Evaluation of days to first flower during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistically significant difference (ANOVA, p > 0.05) among any of the genotypes. Error bars indicate 95% confidence intervals.

Figure 5.
Evaluation of proportion of colored fruits during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistical difference (ANOVA, p > 0.05) among any of the genotypes. Statistically significant difference (** p < 0.01 based on ANOVA in conjunction with a two-tailed Tukey’s HSD multiple comparison test) is indicated between genotypes. Error bars indicate 95% confidence intervals.
Figure 5.
Evaluation of proportion of colored fruits during two seasons. Fla. 7946j2cr and Fla. 8735j2cr are CRISPR-driven mutants, whereas Fla. 7946j2nil and Fla. 8735j2nil are conventionally-bred near-isogenic lines to the corresponding recurrent parents. The n value represents the total number of plants for each genotype evaluated during each season; ns indicates no statistical difference (ANOVA, p > 0.05) among any of the genotypes. Statistically significant difference (** p < 0.01 based on ANOVA in conjunction with a two-tailed Tukey’s HSD multiple comparison test) is indicated between genotypes. Error bars indicate 95% confidence intervals.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, T.G.; Hutton, S.F. Field Evaluation of CRISPR-Driven Jointless Pedicel Fresh-Market Tomatoes. Agronomy 2021, 11, 1957. https://doi.org/10.3390/agronomy11101957
AMA Style
Lee TG, Hutton SF. Field Evaluation of CRISPR-Driven Jointless Pedicel Fresh-Market Tomatoes. Agronomy. 2021; 11(10):1957. https://doi.org/10.3390/agronomy11101957
Chicago/Turabian StyleLee, Tong Geon, and Samuel F. Hutton. 2021. "Field Evaluation of CRISPR-Driven Jointless Pedicel Fresh-Market Tomatoes" Agronomy 11, no. 10: 1957. https://doi.org/10.3390/agronomy11101957
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.