1. Introduction
Yield formation in rice mainly depends on the degree of coordination of carbon and nitrogen (C-N) metabolism during the filling stage [
1,
2]. It has been shown that 9–43% of grain yield comes from unstructured carbohydrates stored before flowering, while postanthesis photosynthetic products contribute 57–91% of yield [
3], and
13CO
2 tracing technology showed that the aboveground
13C assimilates fixed by photosynthesis accounted for 45.3–95% of such assimilates [
4]. N metabolism, accumulation, and redistribution in rice vegetative and regenerative organs are also important factors in determining yield [
5,
6,
7]. The previous work showed that a considerable portion of N is transported from vegetative organs to grains in rice, of which approximately 64% comes from leaves and 20% from stems [
5]. Moreover,
15N tracing showed that 39% and 46%, respectively, of the
15N absorbed at the rice tillering and panicle differentiation stages, are transported to grains at the maturity stage [
6].
The above studies focused on a single aspect of C or N metabolism. During the process of rice postanthesis yield formation, changes in C and N (C-N) metabolism in rice plants will directly affect the yield, transformation, and distribution of C and N substances [
3,
4,
7]. The mechanisms of C-N metabolism regulation are coupled to each other and restrict each other in that not only is C metabolism affected by the regulation of N levels, but N-metabolic-pathway–related enzymes and metabolites are also subject to feedback from C-metabolism–related products [
8]. Water and fertilizer are important parts of high-quality and high-yield rice cultivation techniques [
9,
10]. Previous studies by us and others showed that water and N have a coupling effect on rice yield formation and nitrogen use efficiency (NUE), and proper coordination of W-N regimes is conducive to improving grain yield and NUE [
9,
10,
11]. However, many research on rice postanthesis C-N metabolism has mainly focused on one factor of the effects of water and fertilizer [
12,
13,
14,
15]. It is reported that 79–85% of
14CO
2-labeled photosynthetic products were transported from flag leaves to grains under severe water stress, while only 55–66% were transported from flag leaves to grains under traditional basin irrigation; the proportion of
14CO
2-labeled photosynthetic products transported from stems to grains also increased with the degree of water-stress [
13]. Moreover, the proportions of
14CO
2-labeled photosynthetic products transported from stems to grains were, respectively, 19–22%, 45–46%, and 62–66% under flooded irrigation, mild water stress, and severe water stress at 33 d postanthesis [
14].
15N tracing showed that under different N regimes, the use efficiency of N applied at the booting and tillering stage reached 54–82% and 17–34%, respectively [
15]. Cassman et al. [
12] also showed that 53% of N was absorbed in rice within 10 d of N application during the booting stage, and the absorption rate was as high as 9–12 kg hm
−2 d
−1 within 4 days (d).
There are few reports on how W-N coupling regulates the synergistic process of postanthesis C-N metabolism in rice and on how this in turn improves yield and NUE. Specifically, it is rarely reported whether differences exist in the precise quantification of transport and distribution of postanthesis photoassimilates in rice between different irrigation regimes or different N regimes and whether the differences in postanthesis N transport from leaves and stems under W-N interaction have a synergistic effect on the transport and distribution efficiency of photoassimilates. To this end, based on the relevant research [
3,
4,
5,
12] and the results of our previous studies [
2,
7,
9,
10], using the
13C and
15N dual-isotope tracer and physiological/biochemical analysis methods, we studied the effects of different W-N regimes on postanthesis C-N metabolism, yield, and NUE of hybrid rice. We analyzed the common response mechanism of the postanthesis photoassimilate and N accumulation, transport, and distribution as well as their relationship with yield formation and with NUE in the context of W-N coupling. The findings of this study enrich our understanding of the W-N regulation mechanism in hybrid rice and provide a theoretical basis and practical guidance for the development of water-saving and high-yield rice production.
4. Discussion
C and N metabolism are the two most basic metabolic processes in plants [
8]. How to adjust the relationship between the two through water and fertilizer treatments such that assimilation is coordinately distributed between the two is key to the balance of C-N metabolism and the improvement in yield and NUE in rice. The previous work showed that water-saving irrigation can not only enhance the N metabolism and improve the photosynthetic productivity of leaves but also help increase the activities of ADP-glucose pyrophosphorylase, starch synthase, starch branching enzyme, and other key enzymes of C metabolism during filling stage [
23,
24,
25]. Honjyo et al. showed that increasing the application of N can increase the chlorophyll content and keep the activities of GS, SPS, and phosphoenolpyruvate carboxylase at high levels, which are conducive to promoting the formation of photosynthetic products and the accumulation of grain starch and protein, thereby achieving synergistic increases in yield and protein content [
26]. However, it has rarely been reported which factor between water and N treatment has a more significant effect on the regulation of C-N metabolism. It was also not known, under W-N coupling, what the postanthesis C-N metabolism characteristics are in rice, or what the accumulation and distribution characteristics of C and N substances are.
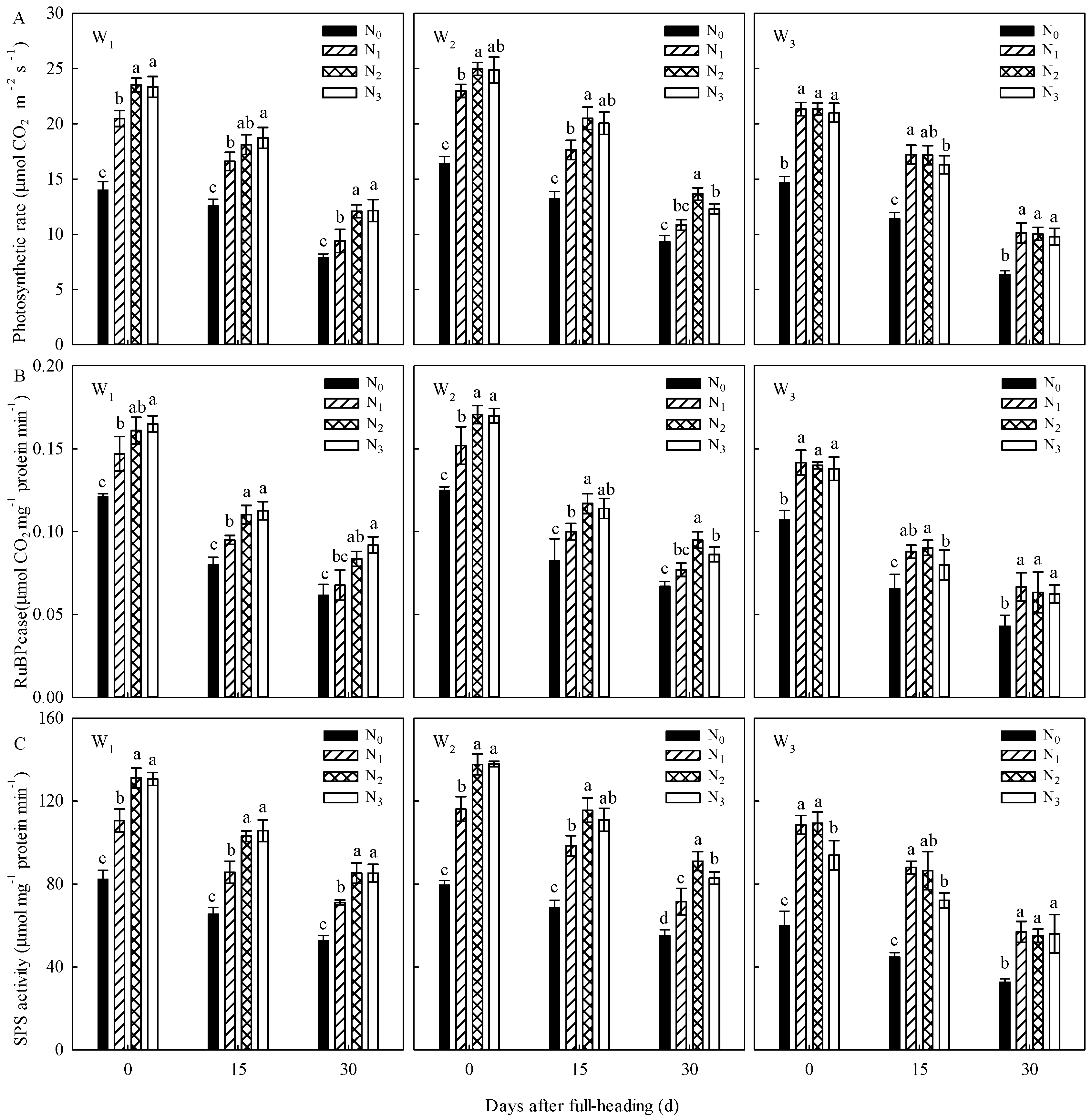
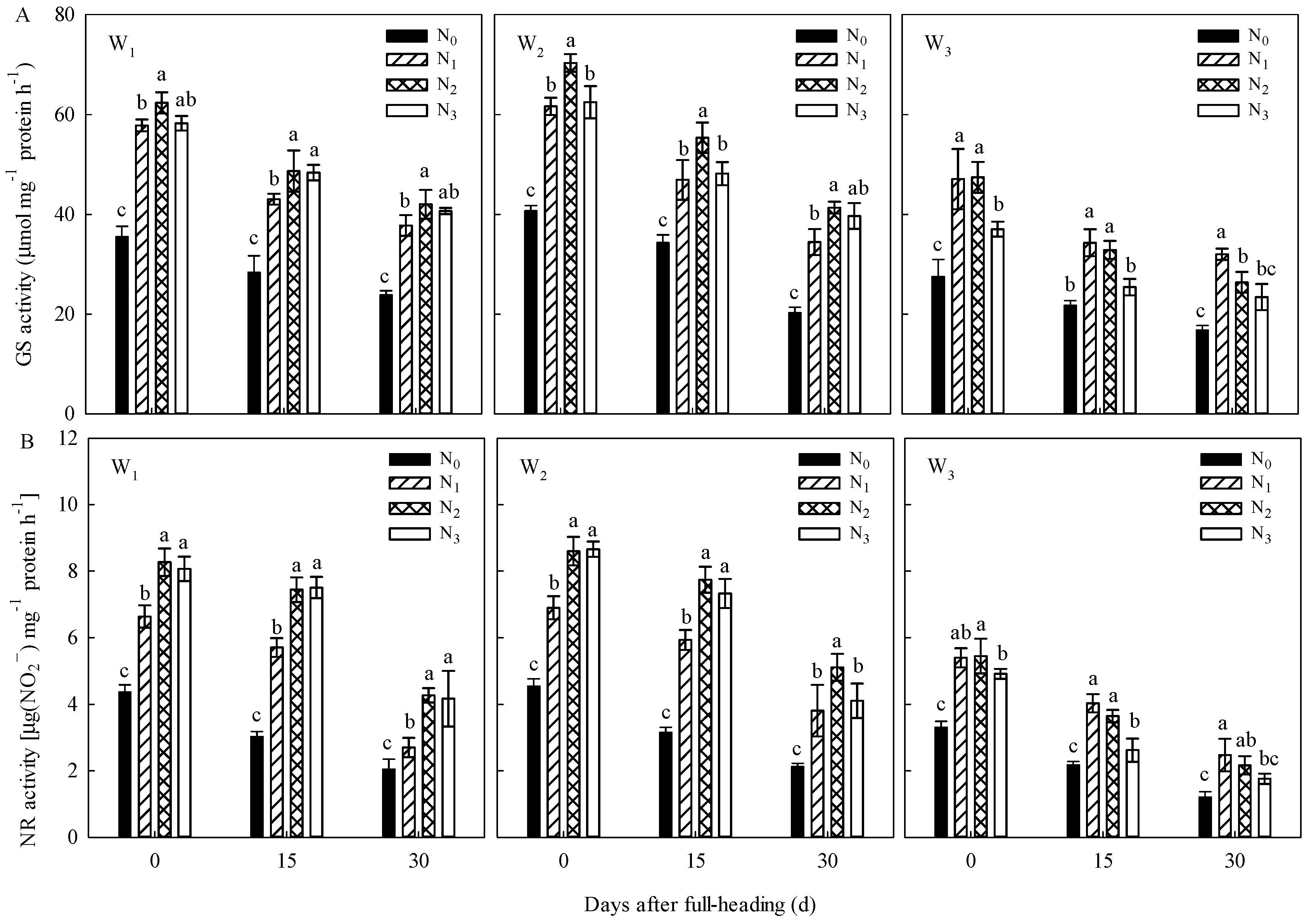
This study showed that the regulatory effects of irrigation regimes on WUE, NUE, and the accumulation of C-assimilates in panicles were significantly higher than the regulatory effects of N regimes; the regulatory effects of N regimes, on the other hand, had a significant regulatory effect on the N accumulation at the maturity stage, C/N, NGPE, NMPE, and yield. A proper water-saving regime and a proper proportion of N application shifted to postpone can coordinate the enzyme activities of C-N metabolism (
Figure 2 and
Figure 3) and promote the coordinated transport of postanthesis C assimilates and N (
Table 7 and
Table 8). Such optimization can also achieve simultaneous increases in rice yield and NUE and can further realize the advantages of W-N coupling (
Table 4,
Table 5 and
Table 6), which further confirms and supplements the results of previous studies, including ours [
7,
13,
14]. This study also showed that when the amount of N application shifted to postpone reached 60% (N
3 treatment), compared to W
1, the inadequate irrigation (W
2 and W
3 treatments) caused a decrease in the enzyme activities of C-N metabolism of postanthesis flag leaves, a significant decrease in N accumulation, an increased retention of C assimilates in leaves and stems, and a significant decrease in C assimilates in panicles, leading to a significant reduction in NAE and NPE. This may be because N cycling would deplete the metabolism/assimilation capacity of inorganic N in the organism and form organic C-N compounds [
8] and cause C-N metabolism to be dysregulated. The greater the application of base fertilizer and fertilizer at the tillering stage, the greater the loss, and the lower the overall NUE [
15]. This study further showed that when a large proportion of N application was shifted to postpone, the amount of N loss was decreased with regard to N accumulation, yet the accumulation and coordinated transport of N and photosynthetic C were low, which also led to a significant reduction in the overall NUE. This result further strengthens the conclusions of a previous study [
15,
24].
This study also found that the increase in water stress intensity under irrigation regimes increased the proportion of
13C photosynthetic products transported from leaves to grains, which is basically consistent with the findings of Yang et al. [
13]; however, in this study, it was shown that severe water stress was not conducive to the transport of
13C photosynthetic products from stem/sheath to grains, which is contrary to the result of Yang et al. [
14] that severe water stress promoted the proportion of photosynthetic products transported from stems (up to 62–66%). This may be because although severe water deficit could increase the proportion of
13C photosynthetic products transported from leaves, the significant decrease in the amount of stem/sheath transport from leaves is the main stage for the decrease of
13C assimilates in grains. Under each irrigation regime, with the proportion of
15N-labeled N application shifted to postpone, the difference in
15N accumulation between various W and N regimes at the full heading stage was significant. This is not quite consistent with the results in Ye et al. [
27], in which the absorption of total
15N in rice was similar between various treatments until the full heading stage, when
15N-labeled N fertilizer was applied twice, as the base fertilizer and before the full heading stage. In this study, the accumulation of
15N in different organs of rice plants at the full heading stage decreased as the proportion of N application shifted to postpone increased (
Table 8), and it was especially significantly reduced under the N
3 treatment. From the full heading to maturity stage, the absorption of
15N was further improved under the N
3 treatment over the N
2 treatment. These results show that shifting N application to postpone could promote the absorption of N in postanthesis rice, but the total
15N absorption under the N
3 treatment was still significantly lower than that under N
2. This result may indicate that under the N
3 treatment, a gap of denitrification existed between the application of base fertilizer (40% N) and topdressing (60%), and postanthesis N retention occurred (
Table 8)—rice plants absorbed N in the soil before topdressing, while the later-dressed
15N-labeled fertilizer may cause the N absorbed at the postanthesis stage to not be transported to grains in a timely manner but rather be retained in leaves and roots. This study only focused on the whole plant level in full-heading and maturity stages, and the dynamic accumulation of
13C and
15N during grain filling, and their relationship needs to be further studied.
Some studies argue that the level of rice yield is closely related to the absorption and utilization of N. C-N metabolism, as the most basic metabolic pathway in crops, provides a material basis for crop growth and development, which largely determines grain yield [
28,
29]. Matt et al. showed that under sufficient supply of N, the decrease in Rubisco activity can lead to decreases in NR activity, nitrate accumulation, and amino acid level [
30]. Therefore, the inhibition of leaf photosynthesis would reduce leaf sugar levels and thus lead to the inhibition of N metabolism. Moreover, the accumulation of photosynthetic products under low-light or basin irrigation conditions decreased the leaf photosynthesis rate and affected N metabolism. Under too much N accumulation, N assimilation would compete with and consume too much C, so the unstructured carbohydrates required for the development of young panicles would then be reduced, thereby affecting C metabolism [
31,
32,
33]. However, all the above studies studied a single vegetative organ, such as leaves or grains, and few studies have reported the relationship between C metabolism and N metabolism in various vegetative organs or whether N metabolism or C metabolism has a more significant effect on nutrient accumulation in grains. This study showed that during the filling stage, under appropriate W-N treatments, the activities of enzymes of C-N metabolism in leaves can also directly regulate C and N accumulation in grains. Specifically, the activities of key enzymes of N metabolism in leaves had a more obvious effect in promoting the accumulation of
13C in panicles, as evidenced by their high correlations (
Table 10). Thus, C metabolism in leaves plays a positive role in regulating N accumulation in grains to achieve the synergy of C-N metabolism between various vegetative organs. This also further showed that under the W-N interaction, the stimulatory effect of the enzyme activities of postanthesis N metabolism on C accumulation in panicles and the regulatory effect of the enzyme activities of C metabolism on N accumulation in panicles are important reasons that C-N metabolism coupling can help achieve high yield and high NUE in rice.
Selecting indicators that can evaluate NUE and yield is key to the evaluation of crop NUE and high yield [
34]. The endopeptidase activity in rice leaves during filling stage can be used as an indicator of rice filling characteristics, yield composition, and quality, and GS activity and soluble protein content also have important reference value for evaluating the NUE in rice [
35,
36]. Our previous research results also showed that the GS activity in functional leaves can be used as an indicator to accurately quantify the N accumulation at various growth stages of rice plants, and the enzyme activities of NR and GS in the flag leaf at the heading stage can be used as a comprehensive indicator of the simultaneous increase in rice yield and NUE [
9]. According to this study, under different irrigation and N regimes, the photosynthesis rate of the flag leaf and the enzyme activities of C-N metabolism, such as RuBPCase and GS, at various postanthesis growth stages of rice plants can accurately reflect the NUE and yield levels of rice (
Table 11), which further confirms the results of previous studies [
9,
36].
At the same time, this study also showed by
13C labeling that the postanthesis accumulation and transport of photosynthetic C assimilates had significant consistency and synergy with the photosynthesis rate of the flag leaf and the activities of key enzymes of C-N metabolism. It further clarified that C-N metabolism can succinctly and clearly reflect rice yield and NUE as well as the accumulation and transport of photoassimilates. It showed that the improvement in N metabolism capacity can simultaneously improve the level of C metabolism and that the activity of key enzymes of C metabolism can be used as physiological and biochemical indicators for screening and identifying high-NUE rice genotypes. Weigelt et al. showed that the normal growth of crops does not require high N and C contents but can be accomplished when the C/N ratio reaches a certain balance [
37]. However, few studies have reported how the balance of photosynthetic C assimilate accumulation in rice plants changes with water and N absorption dynamics in roots, or how the C/N ratio changes under W-N coupling. This study found that under high-yield conditions, from the full heading to maturity stage, the C/N ratio was approximately twice that under other conditions in leaves and in panicles and approximately half in stems and in roots (
Table 9). Hence, the C/N change value of postanthesis vegetative organs in rice can be used as an indicator and a theoretical basis for the simultaneous improvement in yield and NUE. The optimization of the C/N ratios of various rice vegetative organs from the full heading to maturity stage serves as another important approach to achieve high yield, high WUE, and high NUE; this approach is more accurate than the determination of rice yield and NUE through measuring the activity of a specific enzyme of N or C metabolism at a specific growth stage.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}