Colchicine Mutagenesis from Long-term Cultured Adventitious Roots Increases Biomass and Ginsenoside Production in Wild Ginseng (Panax ginseng Mayer)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Mutant Root Induction with Colchicine
2.3. DNA Content Analysis by Flow Cytometry
2.4. RAPD Analysis
2.4.1. DNA Isolation
2.4.2. PCR Amplification
2.5. RNA Isolation and Quantitative Real-Time PCR Analysis of Gene Expression
2.6. Preparation and Extraction of Mutant Lines
2.7. HPLC Analysis of Ginsenosides
2.8. Lipid Peroxidation
2.9. Catalase and Peroxidase Activity
2.10. Statistics
3. Results and Discussion
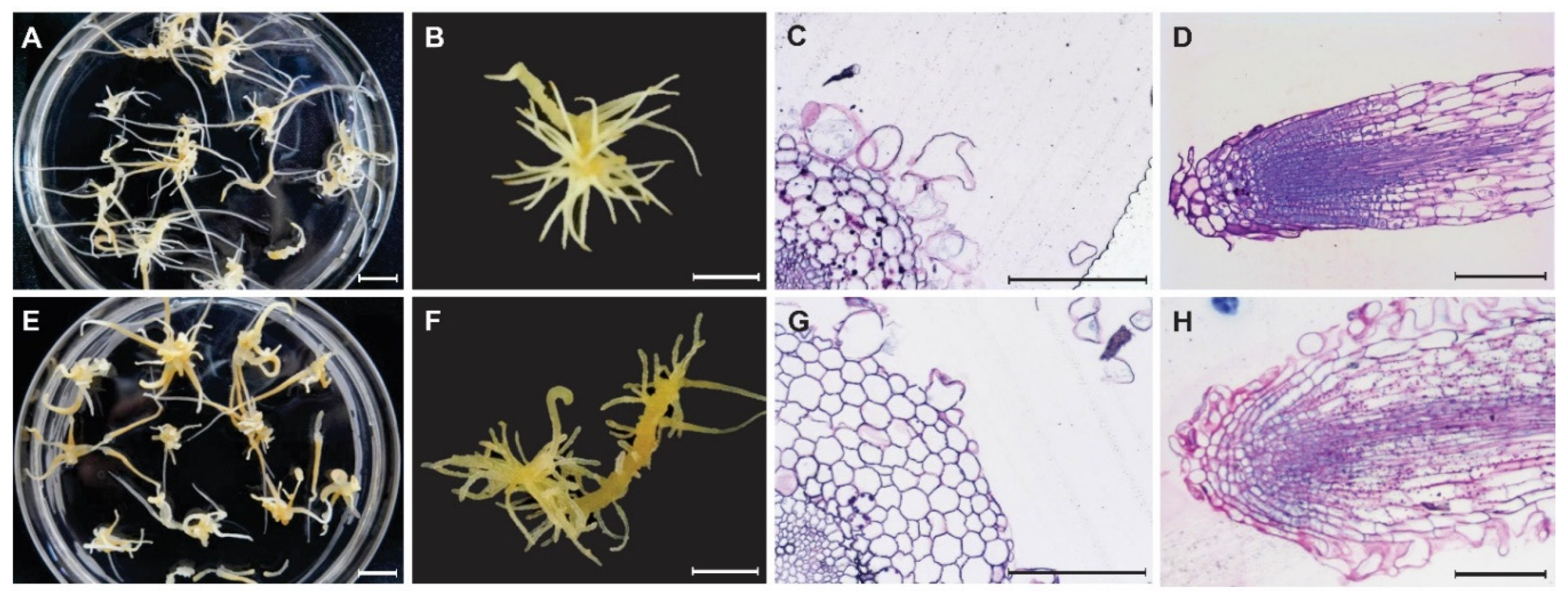
3.1. Effect of Colchicine Treatment on Lateral Root Induction and Growth
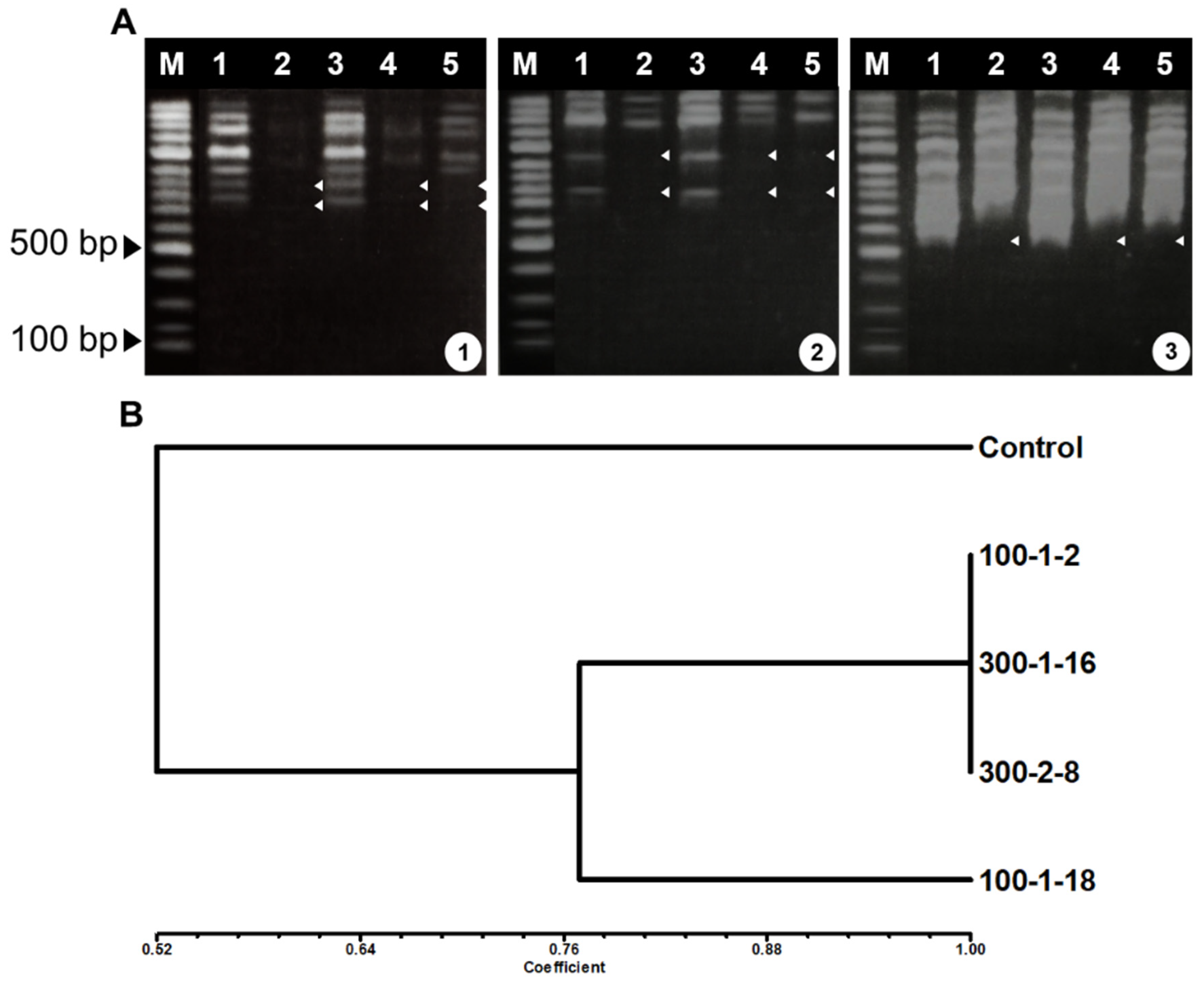
3.2. RAPD Analysis of Mutant Roots
3.3. Expression Analysis of Ginsenoside Biosynthetic Genes
3.4. Ginsenoside Content Analysis by HPLC
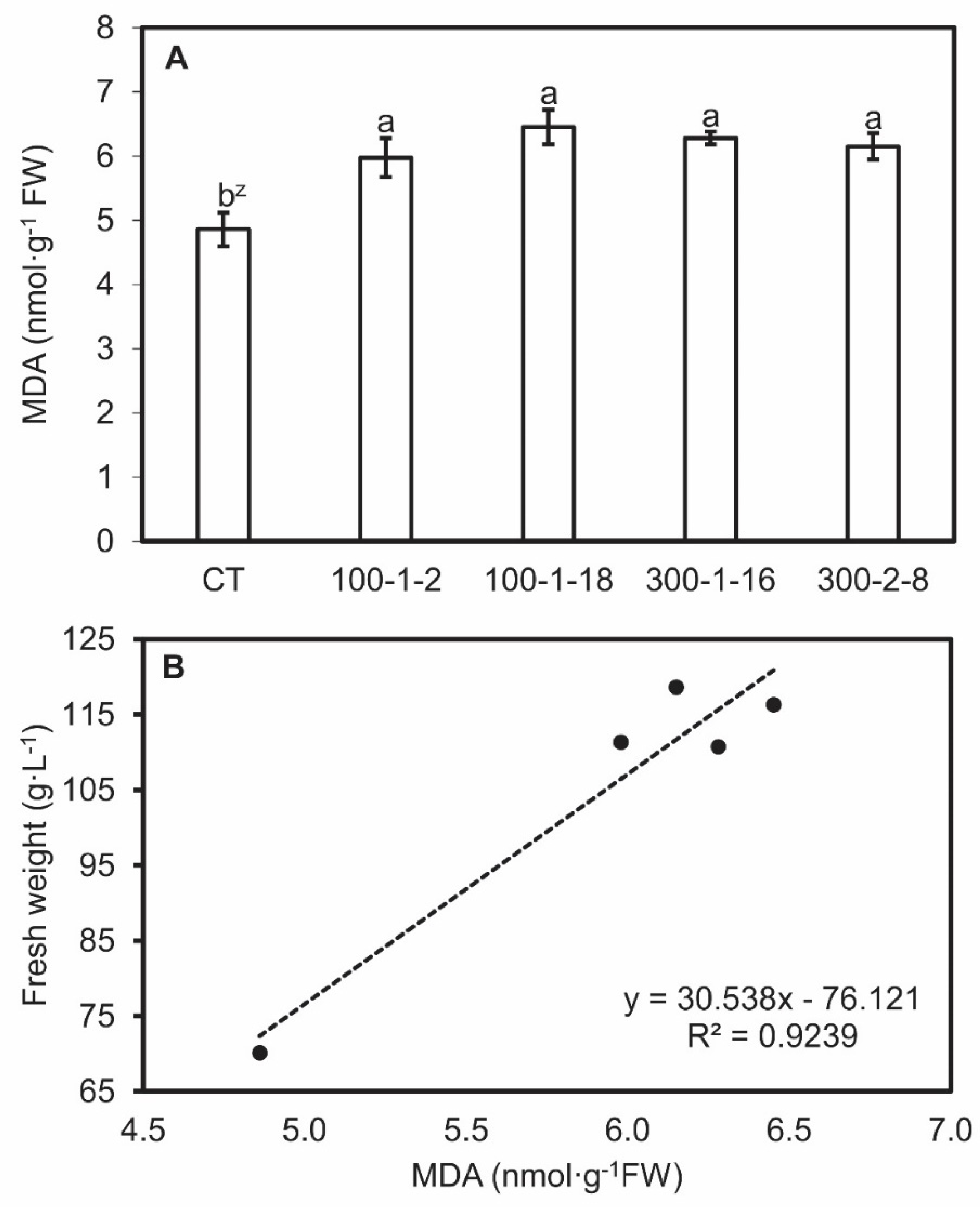
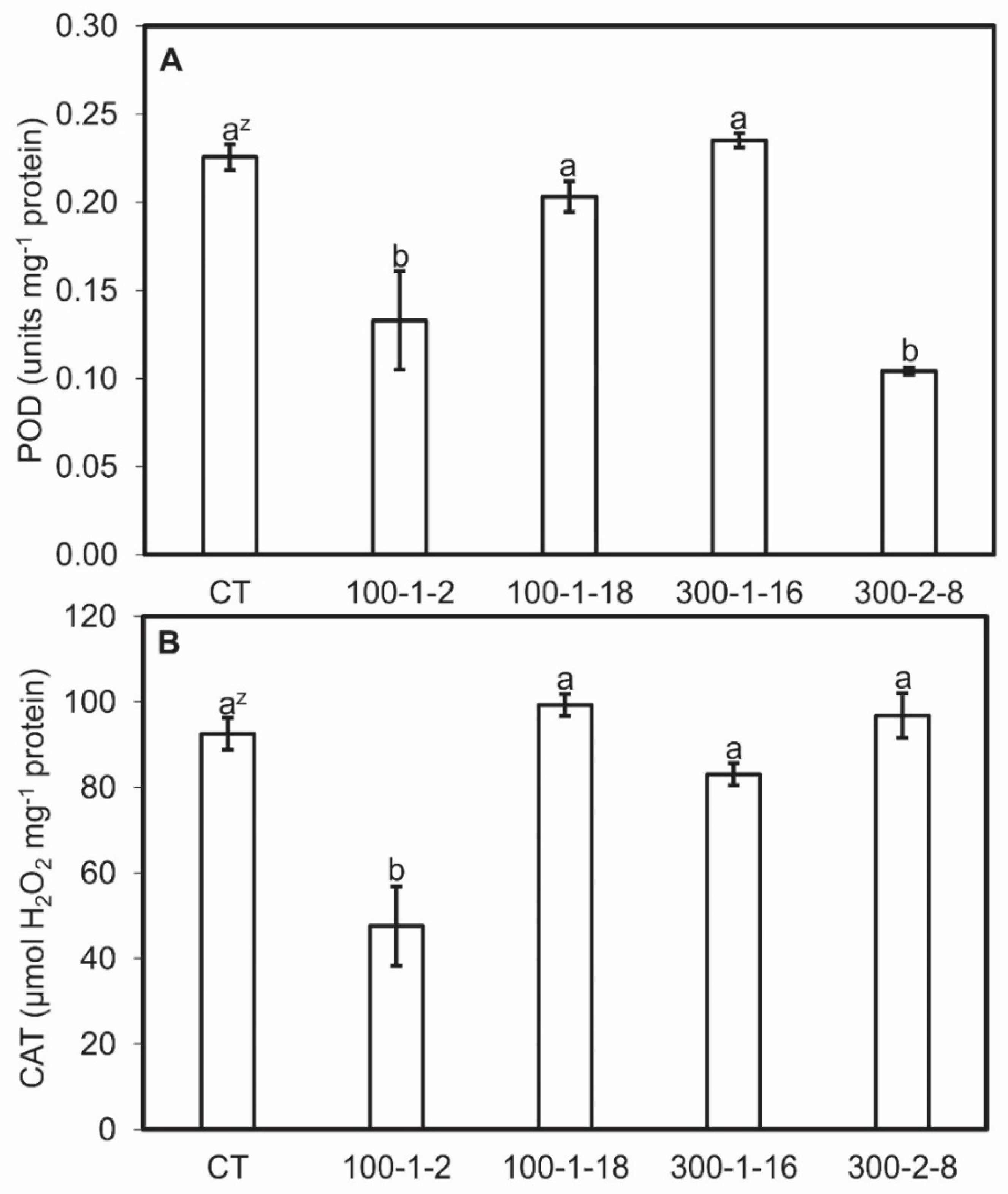
3.5. MDA, CAT, and POD Activity of Mutant Roots
3.6. Bioreactor Culture of Mutant Lines
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bairu, M.W.; Kane, M.E. Physiological and developmental problems encountered by in vitro cultured plants. Plant Growth Regul. 2011, 63, 101–103. [Google Scholar] [CrossRef] [Green Version]
- Bairu, M.W.; Aremu, A.O.; Van Staden, J. Somaclonal variation in plants: Causes and detection methods. Plant Growth Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, Y.E.; Yoon, Y.J.; Jeong, C.S.; Lian, M.L.; Paek, K.Y.; Park, S.Y. Highly endoreduplicated floral organs of somaclonal variants in clonally propagated Phalaenopsis “Spring Dancer”. Plant Cell Tissue Organ Cult. 2016, 126, 67–77. [Google Scholar] [CrossRef]
- Whitmer, S.; Canel, C.; van der Heijden, R.; Verpoorte, R. Long-term instability of alkaloid production by stably transformed cell lines of Catharanthus roseus. Plant Cell Tissue Organ Cult. 2003, 74, 73–80. [Google Scholar] [CrossRef]
- Qu, J.; Zhang, W.; Yu, X.; Jin, M. Instability of anthocyanin accumulation in Vitis vinifera L. var. Gamay Fréaux suspension cultures. Biotechnol. Bioprocess. Eng. 2005, 10, 155. [Google Scholar] [CrossRef]
- Li, L.Q.; Fu, C.H.; Zhao, C.F.; Xia, J.; Wu, W.J.; Yu, L.J. Efficient extraction of RNA and analysis of gene expression in a long-term Taxus cell culture using real-time RT-PCR. Z. Naturforsch C 2009, 64, 125–130. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S.; Shumakova, O.A. DNA mutagenesis in 2-and 20-yr-old Panax ginseng cell cultures. In Vitro Cell Dev. Biol. 2013, 49, 175–182. [Google Scholar] [CrossRef]
- Yu, K.W.; Gao, W.Y.; Son, S.H.; Paek, K.Y. Improvement of ginsenoside production by jasmonic acid and some other elicitors in hairy root culture of ginseng (Panax ginseng CA Meyer). In Vitro Cell Dev. Biol. 2000, 36, 424–428. [Google Scholar] [CrossRef]
- Lian, M.L.; Chakrabarty, D.; Paek, K.Y. Effect of plant growth regulators and medium composition on cell growth and saponin production during cell-suspension culture of mountain ginseng (Panax ginseng C. A. mayer). J. Plant Biol. 2002, 45, 201–206. [Google Scholar] [CrossRef]
- Leitão, J.M. Chemical mutagenesis. In Plant Mutation Breeding and Biotechnology; Shu, Q., Forster, B., Nakagawa, H., Eds.; CABI: Wallingford, UK, 2012; pp. 135–158. [Google Scholar]
- Davidson, D.; Pertens, E.; Zhao, J.P. Chromosome distribution between two restitution nuclei in a cell following colchicine treatment. Can. J. Genet. Cytol. 1983, 25, 437–445. [Google Scholar] [CrossRef]
- Ade, R.; Rai, M.K. Colchicine, current advances and future prospects. Nusant. Biosci. 2010, 2, 90–96. [Google Scholar]
- Naranjo, T. Contribution of structural chromosome mutants to the study of meiosis in plants. Cytogenet Genome Res. 2015, 147, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Bragdo, M. Production of polyploids by colchicine. Euphytica 1955, 4, 76–82. [Google Scholar]
- Balkanjieva, J. Influence of genotype on mutagenic variability in barley (H. vulgare L.) following colchicine treatment. Barley Genet. Newsl. 1980, 10, 7–10. [Google Scholar]
- Downes, R.W.; Marshall, D.R. Colchicine-induced variants in sunflower. Euphytica 1983, 32, 757–766. [Google Scholar] [CrossRef]
- Mensah, J.K.; Obadoni, B.O.; Akomeah, P.A.; Ikhajiagbe, B.; Ajibolu, J. The effects of sodium azide and colchicine treatments on morphological and yield traits of sesame seed (Sesame indicum L.). Afr. J. Biotechnol. 2007, 6, 534–538. [Google Scholar]
- El-Nashar, Y.I.; Ammar, M.H. Mutagenic influences of colchicine on phenological and molecular diversity of Calendula officinalis L. Genet. Mol. Res. GMR 2016, 15. [Google Scholar] [CrossRef]
- Kamwean, P.; Chaisan, T.; Thobunluep, P.; Phumichai, C.; Bredemeier, M. Changing of morphological characteristic and biomass properties in Pennisetum purpureum by colchicine treatment. J. Agron. 2016, 16, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Kharde, A.V.; Chavan, N.S.; Chandre, M.A.; Autade, R.H.; Khetmalas, M.B. In vitro enhancement of bacoside in brahmi (Bacopa monnieri) using colchicine. J. Plant Biochem. Physiol. 2017, 5, 1–6. [Google Scholar]
- Nura, S.; Adamu, A.K.; Mu’Azu, S.; Dangora, D.B.; Fagwalawa, L.D. Morphological characterization of colchicine-induced mutants in sesame (Sesamum indicum L.). J. Biol. Sci. 2013, 13, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Ahloowalia, B.S.; Maluszynski, M. Induced mutations–A new paradigm in plant breeding. Euphytica 2001, 118, 167–173. [Google Scholar] [CrossRef]
- Obute, G.C.; Ndukwu, B.C.; Chukwu, O.F. Targeted mutagenesis in Vigna unguiculata (L.) Walp. and Cucumeropsis mannii (NAUD) in Nigeria. Afr. J. Biotechnol. 2007, 6, 2467–2472. [Google Scholar] [CrossRef] [Green Version]
- Shehu, A.S.; Adelanwa, M.A.; Alonge, S.O. Effects of gamma radiation and colchicine on crude protein content of soy-bean [Glycine max (L.) Merrill] varieties. Int. J. Res. 2016, 3, 1186–1193. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Doyle, J.J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Am. 1987, 19, 11–15. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bisht, S.S.; Sharma, A.; Chaturvedi, K. Certain metabolic lesions of chromium toxicity in radish. Indian J. Agric. Biochem. 1989, 2, 109–115. [Google Scholar]
- Roy, A.; Leggett, G.; Koutoulis, A. In vitro tetraploid induction and generation of tetraploids from mixoploids in hop (Humulus lupulus L.). Plant Cell Rep. 2001, 20, 489–495. [Google Scholar] [CrossRef]
- Dhooghe, E.; Van Laere, K.; Eeckhaut, T.; Leus, L.; Van Huylenbroeck, J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tissue Organ Cult. 2011, 104, 359–373. [Google Scholar] [CrossRef]
- Hewawasam, W.; Bandara, D.C.; Aberathne, W.M. New phenotypes of Crossandra infundibuliformis var. Danica through in vitro culture and induced mutations. Trop. Agric. Res. 2004, 16, 253–270. [Google Scholar]
- Atak, Ç.; Celik, O.; Acik, L. Genetic analysis of Rhododendron mutants using random amplified polymorphic DNA (RAPD). Pak. J. Bot. 2011, 43, 1173–1182. [Google Scholar]
- Zainudin, A.; Fitriani, H. Analysis of genetic diversity on mutants Jatropha curcas using RAPD. Energy Procedia 2014, 47, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, H.N.; Georgiev, M.I.; Kim, Y.S.; Jeong, C.S.; Kim, S.J.; Park, S.Y.; Paek, K.Y. Ginsenosides: Prospective for sustainable biotechnological production. Appl. Microbiol. Biotechnol. 2014, 98, 6243–6254. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zhang, D.; Yang, D.C. Biosynthesis and biotechnological production of ginsenosides. Biotechnol. Adv. 2015, 33, 717–735. [Google Scholar] [CrossRef]
- Abe, I.; Rohmer, M.; Prestwich, G.D. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpenes. Chem. Rev. 1993, 93, 2189–2206. [Google Scholar] [CrossRef]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef]
- Mehreen, K. Assessment of biochemical attributes of Praecitrullus fistulosus treated with mutagens. J. Plant Breed. Genet. 2015, 3, 39–47. [Google Scholar]
- Caruso, I.; Lepore, L.; De Tommasi, N.; Dal Piaz, F.; Frusciante, L.; Aversano, R.; Garramone, R.; Carputo, D. Secondary metabolite profile in induced tetraploids of wild Solanum commersonii Dun. Chem. Biodivers. 2011, 8, 2226–2237. [Google Scholar] [CrossRef]
- Xu, C.; Tang, T.; Chen, R.; Liang, C.; Liu, X.; Wu, C.; Yang, Y.S.; Yang, D.P.; Wu, H. A comparative study of bioactive secondary metabolite production in diploid and tetraploid Echinacea purpurea (L.) Moench. Plant Cell Tissue Organ Cult. 2014, 116, 323–332. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence |
|---|---|
| A-05 | 5ʹ-AGGGGTCTTG-3ʹ |
| A-08 | 5ʹ-GTGACGTAGG-3ʹ |
| A-09 | 5ʹ-GGGTAACGCC-3ʹ |
| A-10 | 5ʹ-GTGATCGCAG-3ʹ |
| A-13 | 5ʹ-CAGCACCCAC-3ʹ |
| A-15 | 5ʹ-TTCCGAACCC-3ʹ |
| Colchicine Concentration (mg·L−1) | Exposure Time (day) | Lateral Root Formation (%) | No. of Lateral Roots/Explant | Lateral Root Length (cm) |
|---|---|---|---|---|
| 0 | 0 | 100.0a z | 27.75a | 2.51a |
| 100 | 1 | 100.0a | 25.75b | 2.24c |
| 2 | 100.0a | 20.13c | 1.79e | |
| 3 | 100.0a | 12.25g | 1.50g | |
| 200 | 1 | 100.0a | 18.25d | 2.31b |
| 2 | 100.0a | 15.00e | 2.01d | |
| 3 | 97.5ab | 11.13h | 1.60f | |
| 300 | 1 | 100.0a | 17.50d | 2.34b |
| 2 | 97.5ab | 13.38f | 2.05d | |
| 3 | 95.0b | 10.75h | 2.01d |
| Culture Scale | Lines | DNA Index z | No. of Lateral Roots/Explant | Fresh Mass (g·L−1) | Dry Mass (g·L−1) | Growth Ratio y |
|---|---|---|---|---|---|---|
| Flask (250 mL) | Control | 1.00a | 24.00c x | 70.08b | 4.94b | 6.01b |
| 100–1-2 | 1.08a | 32.00bc | 111.32a | 8.69a | 10.14a | |
| 100–1-18 | 0.99a | 38.25ab | 116.27a | 9.95a | 10.63a | |
| 300–1-16 | 1.00a | 28.50bc | 118.60a | 8.14ab | 10.86a | |
| 300–2-8 | 0.96a | 48.00a | 110.71a | 8.45a | 10.07a | |
| Bioreactor (3 L) | Control | 1.00a | 25.83c | 73.36d | 5.39c | 13.91c |
| 100–1-2 | 1.08a | 39.83b | 84.31c | 6.25bc | 16.10b | |
| 100–1-18 | 0.99a | 50.50a | 94.87b | 7.19ab | 17.98a | |
| 300–1-16 | 1.00a | 39.50b | 93.28b | 7.60ab | 17.66a | |
| 300–2-8 | 0.96a | 38.50b | 99.84a | 6.87ab | 18.98a |
| Line | PPD (mg·g−1 DW) | PPT (mg·g−1 DW) | PPD (mg·g−1 DW) | PPT (mg·g−1 DW) | PPD/PPT | Total Ginsenoside (mg·g−1 DW) z | Ginsenoside Productivity (mg·L−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rb1 | Rb2 | Rb3 | Rc | Rd | Rg3 | Rh2 | Re | Rf | Rg1 | Rg2 | ||||||
| Control | 1.08d y | 0.29c | 1.23a | 0.39b | 0.24a | 0.06a | 0.52a | 1.64c | 0.07b | 0.62b | 0.22b | 3.71b | 2.54c | 1.46b | 6.25b | 38.45c |
| 100–1-2 | 2.98b | 0.51b | 0.15b | 0.59b | 0.46a | 0.06ab | 0.53a | 1.13c | 0.09b | 0.53b | 0.18b | 5.08b | 1.93c | 2.63a | 7.01b | 77.21b |
| 100–1-18 | 4.01a | 1.19a | 0.28b | 0.88a | 0.50a | 0.05a | 0.48a | 4.72a | 0.33a | 1.06a | 0.71a | 7.85a | 6.81a | 1.15c | 14.66a | 186.60a |
| 300–1-16 | 2.18bc | 0.52b | 0.14b | 0.35b | 0.30a | 0.05ab | 0.44a | 1.60c | 0.05b | 0.64b | 0.25b | 4.22b | 2.54c | 1.66b | 6.76b | 72.60b |
| 300–2-8 | 1.74cd | 0.45bc | 0.10b | 0.29b | 0.31a | 0.10b | 0.44a | 2.29b | 0.12b | 1.16a | 0.30b | 3.56b | 3.87b | 0.92d | 7.43b | 71.83b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, K.-C.; Ho, T.-T.; Lee, J.-D.; Paek, K.-Y.; Park, S.-Y. Colchicine Mutagenesis from Long-term Cultured Adventitious Roots Increases Biomass and Ginsenoside Production in Wild Ginseng (Panax ginseng Mayer). Agronomy 2020, 10, 785. https://doi.org/10.3390/agronomy10060785
Le K-C, Ho T-T, Lee J-D, Paek K-Y, Park S-Y. Colchicine Mutagenesis from Long-term Cultured Adventitious Roots Increases Biomass and Ginsenoside Production in Wild Ginseng (Panax ginseng Mayer). Agronomy. 2020; 10(6):785. https://doi.org/10.3390/agronomy10060785
Chicago/Turabian StyleLe, Kim-Cuong, Thanh-Tam Ho, Jong-Du Lee, Kee-Yoeup Paek, and So-Young Park. 2020. "Colchicine Mutagenesis from Long-term Cultured Adventitious Roots Increases Biomass and Ginsenoside Production in Wild Ginseng (Panax ginseng Mayer)" Agronomy 10, no. 6: 785. https://doi.org/10.3390/agronomy10060785