
Pax3 and Pax7 Exhibit Distinct and Overlapping Functions in Marking Muscle Satellite Cells and Muscle Repair in a Marine Teleost, Sebastes schlegelii

 ,
,
Abstract
:1. Introduction
2. Results
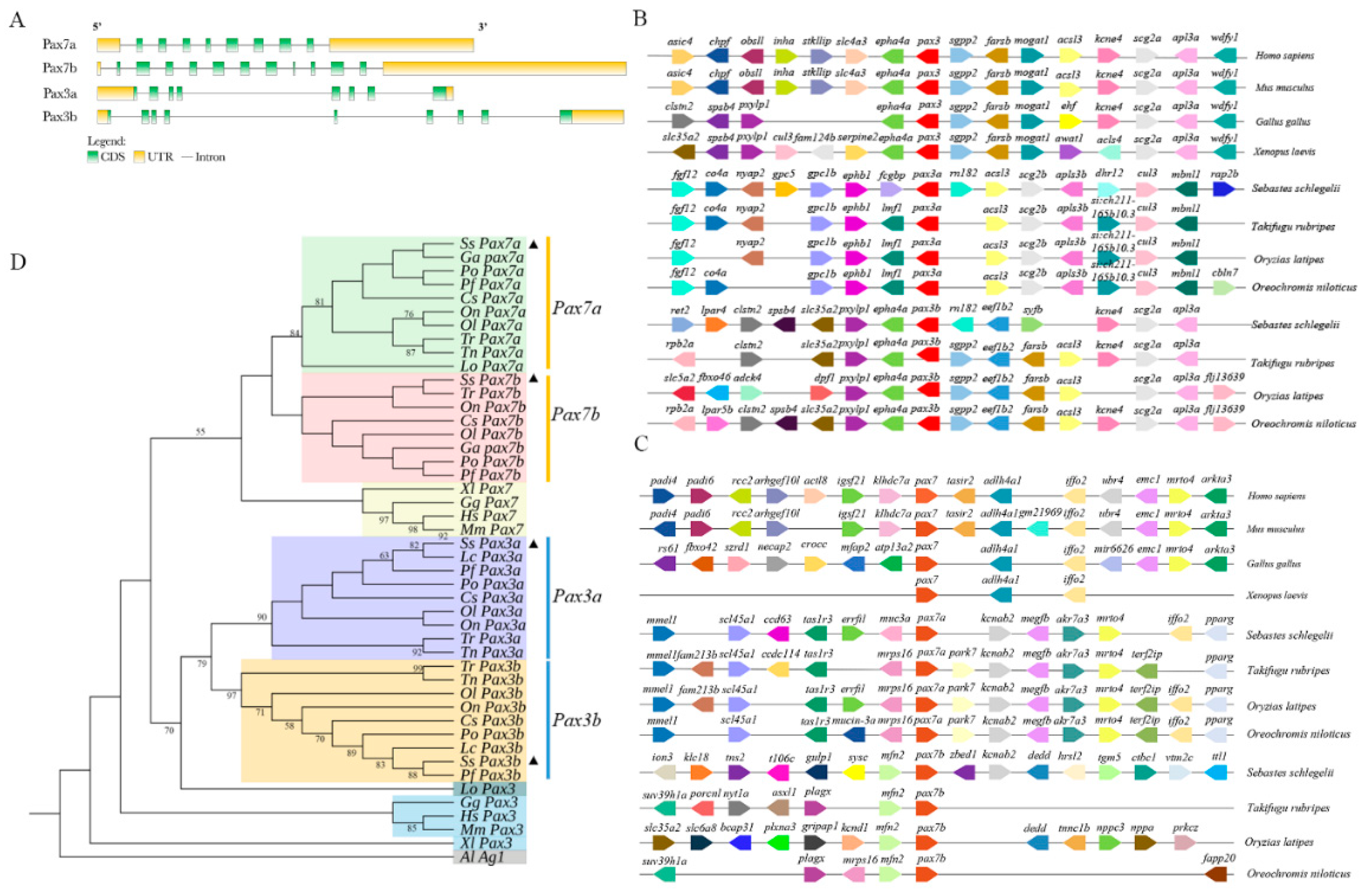
2.1. Gene Structure, Synteny Analyses, and Evolution of SsPax3 and SsPax7
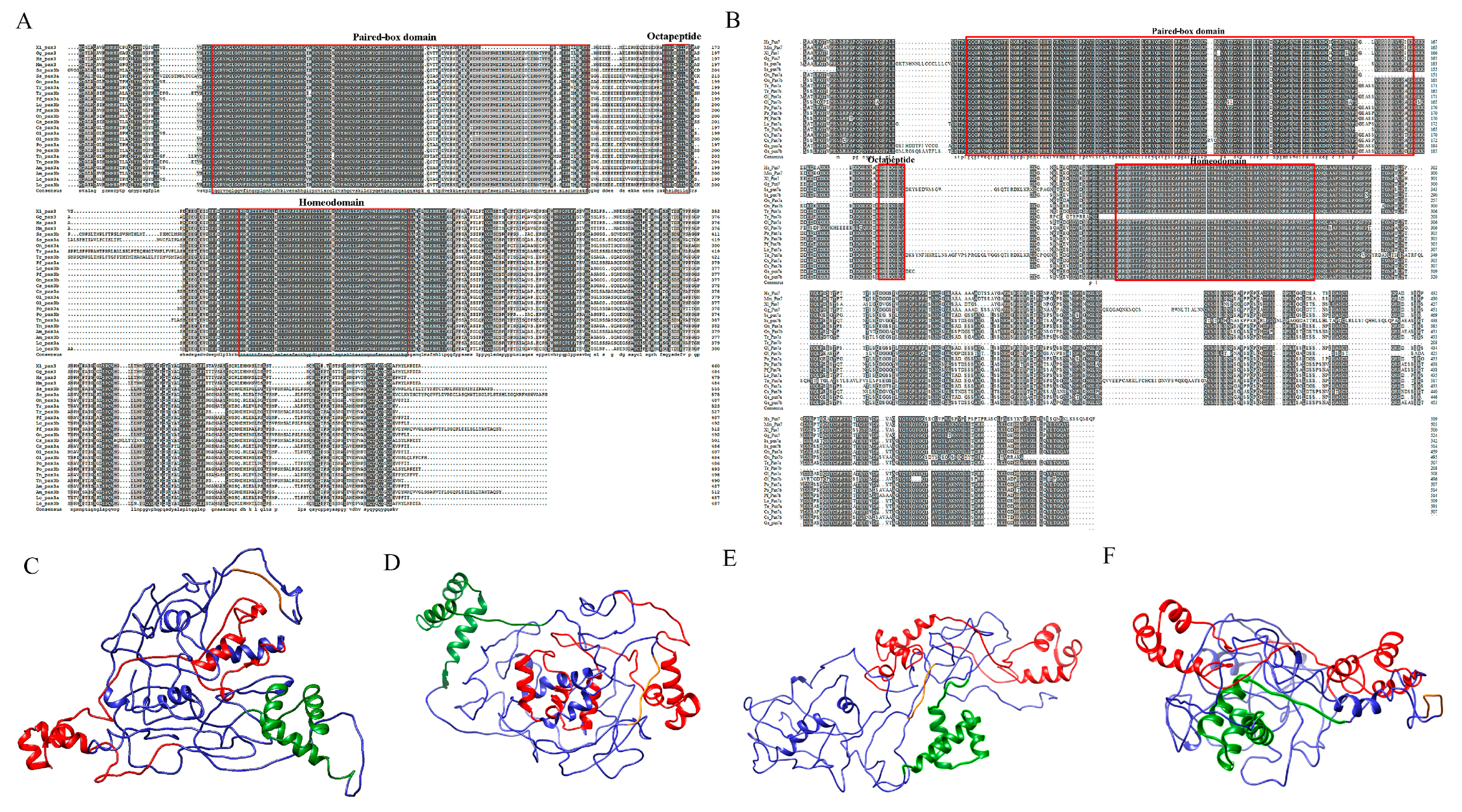
2.2. Sequence Analysis and 3D Protein Structures of SsPax3 and SsPax7
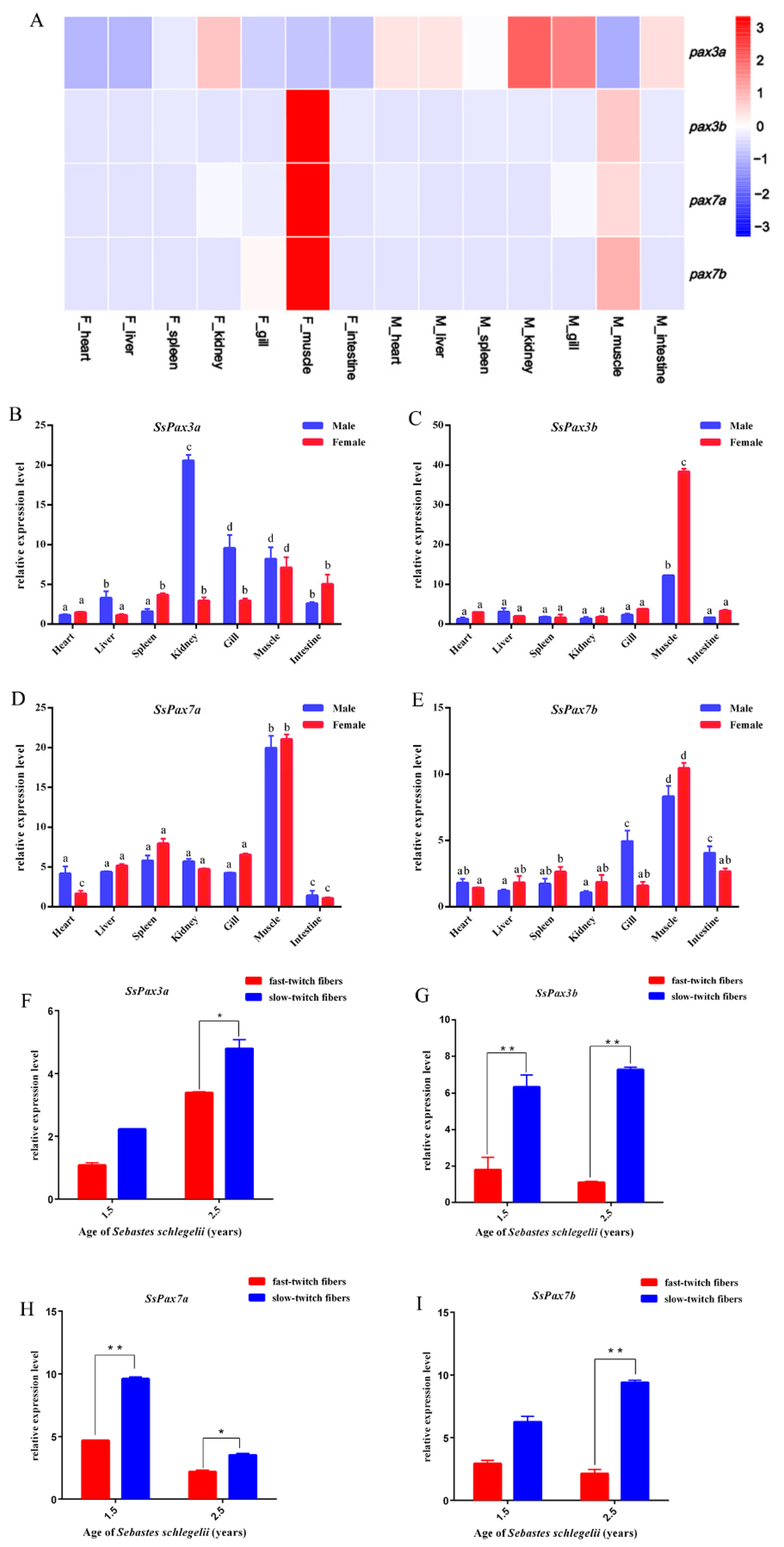
2.3. Expression of S. schlegelii Pax3 and Pax7 in Different Tissues
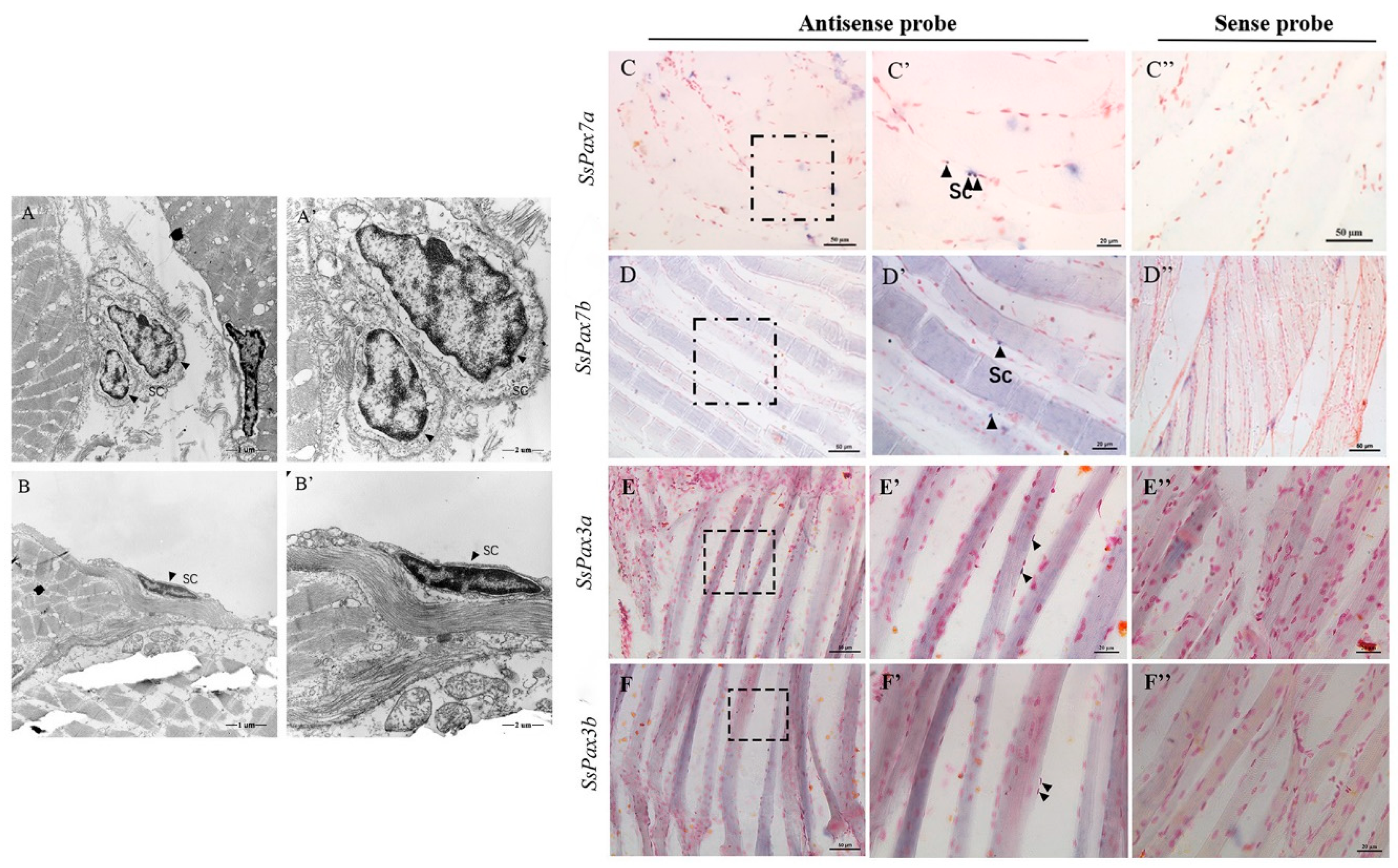
2.4. Muscle Satellite Cells Identification in S. schlegelii
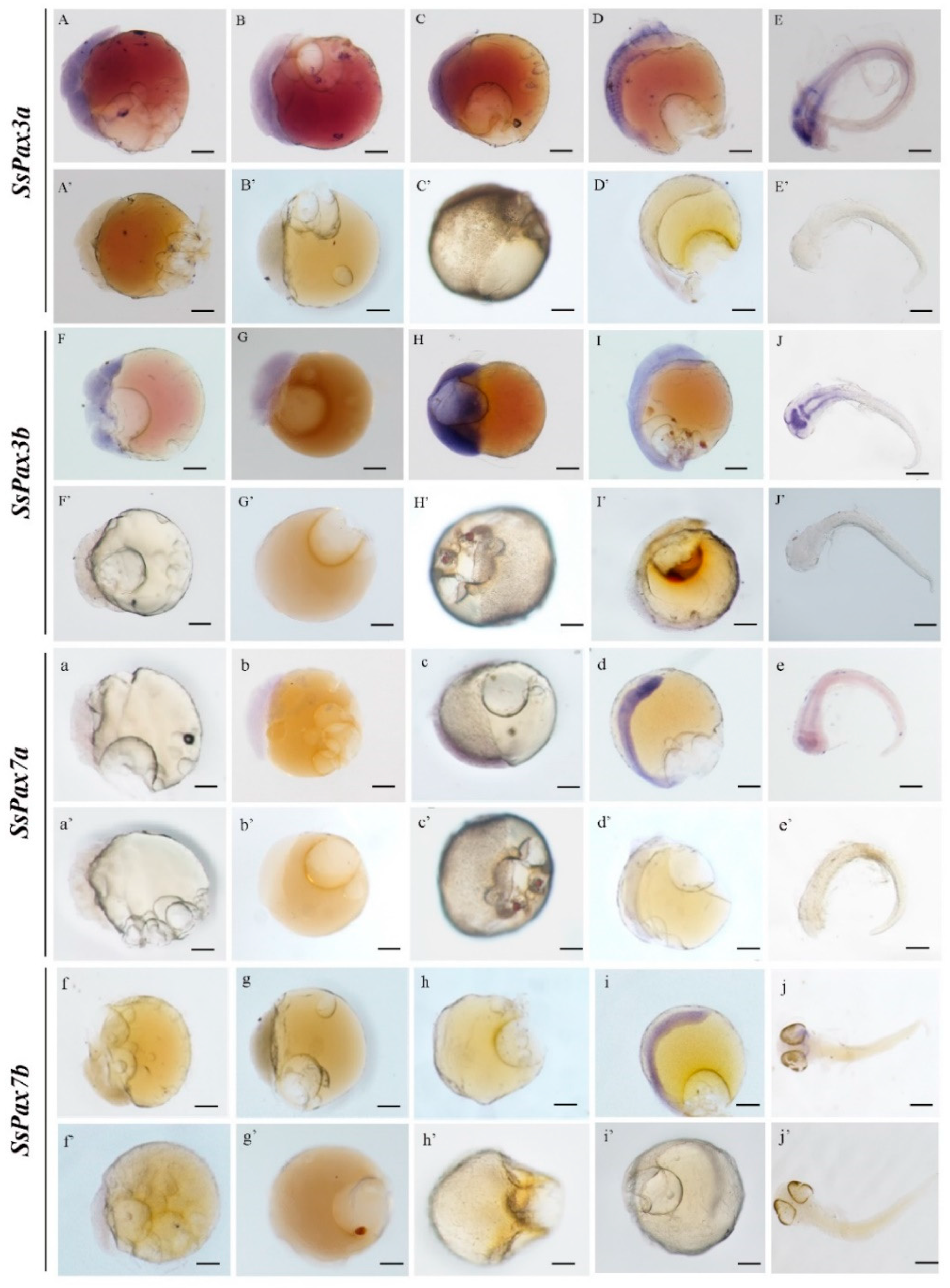
2.5. Tracking the Embryonic Origin of Muscle Satellite Cells in S. schlegelii
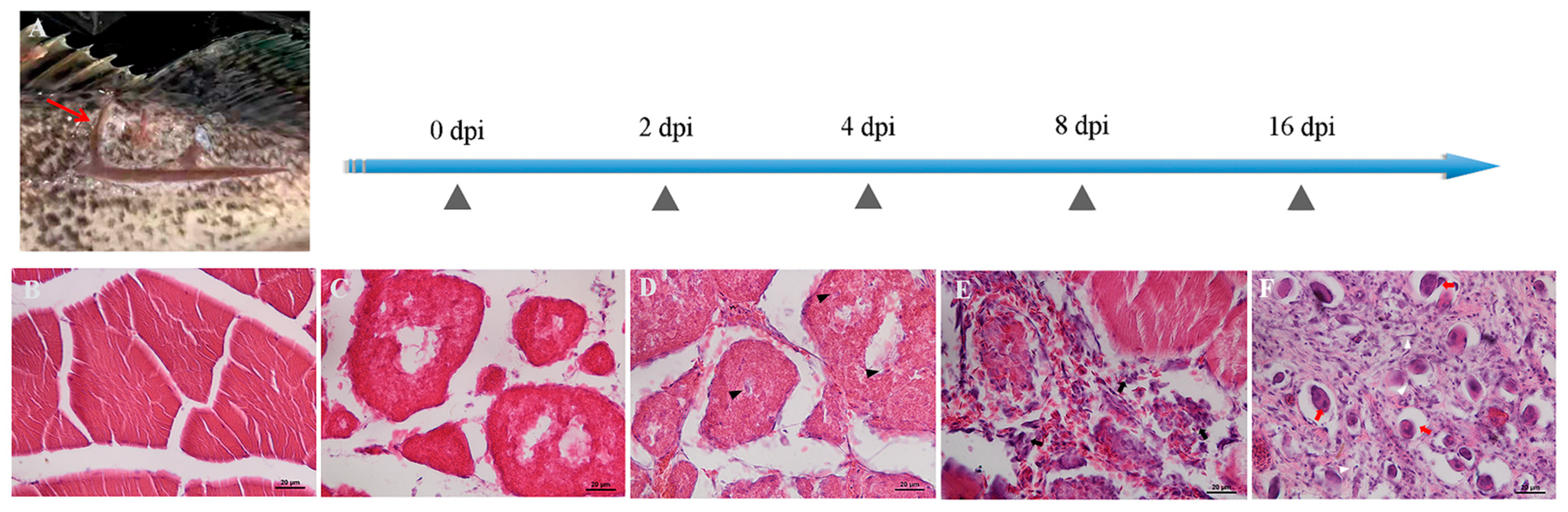
2.6. Histological Examination of S. schlegelii Muscle after Injury
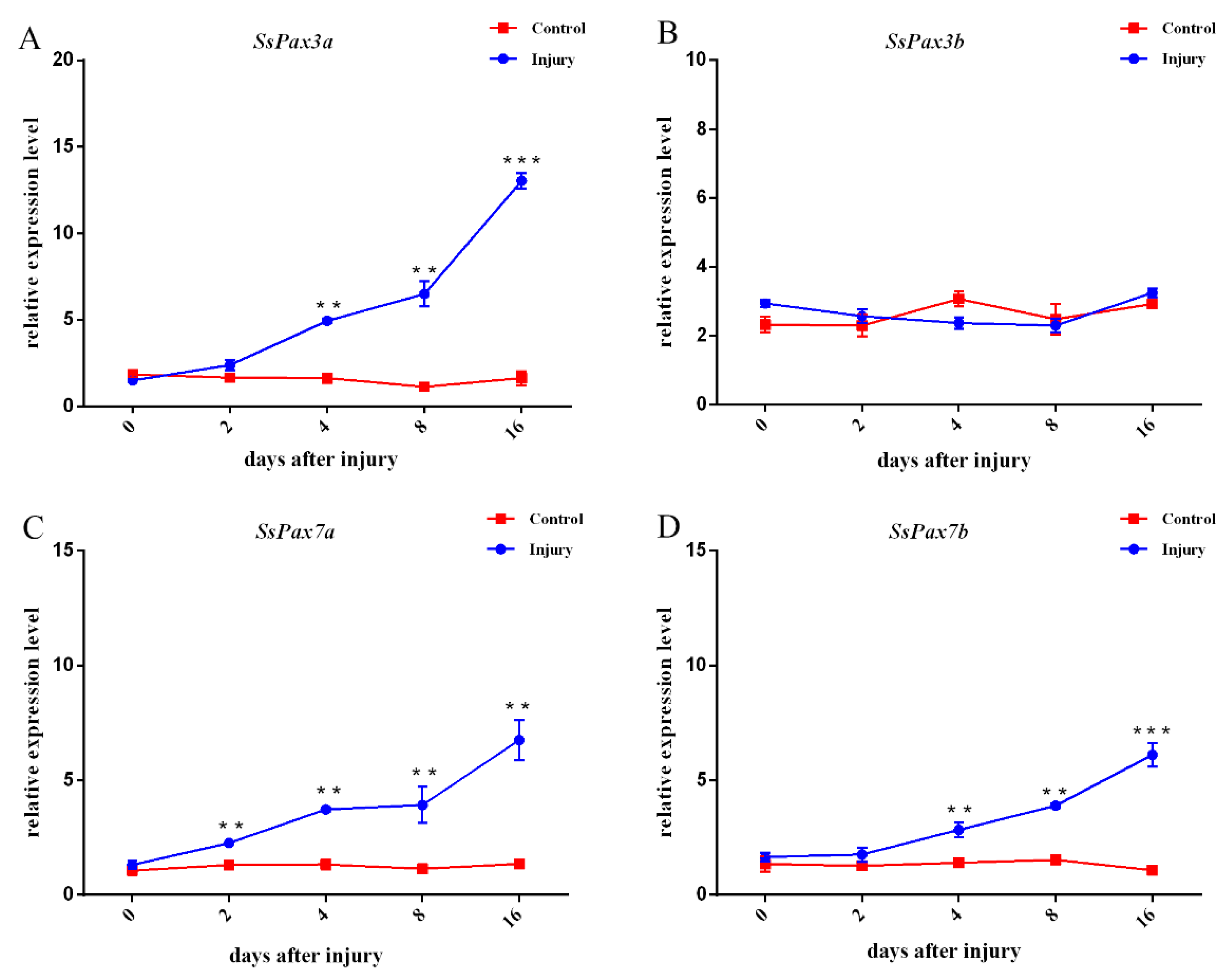
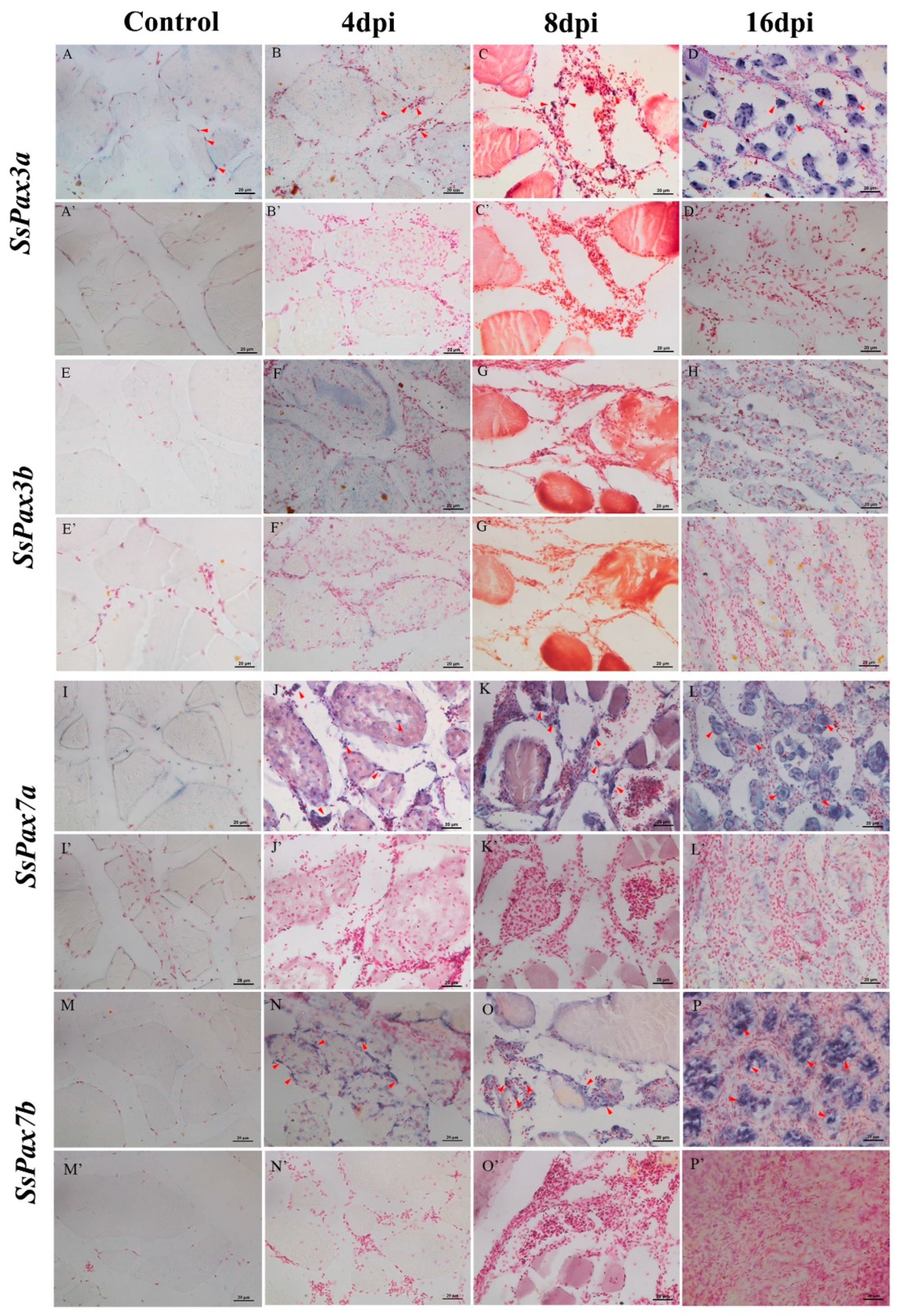
2.7. The Function of SsPax3 and SsPax7 in Injured Muscle during Repair
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fish and Sampling
4.3. Phylogenetic Analysis of Pax3 and Pax7 Genes
4.4. Protein 3D Structure Predictions of SsPax3 and SsPax7
4.5. Expression Analysis Using the Available RNA-seq Libraries and Quantitative Real-Time PCR (qRT-PCR)
4.6. Muscle Regeneration Experiment
4.7. In Situ Hybridization (ISH)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Pax3 | Paired box protein Pax-3 |
| Pax3a | Paired box protein Pax-3a |
| Pax3b | Paired box protein Pax-3b |
| Pax7 | Paired box protein Pax-7 |
| Pax7a | Paired box protein Pax-7a |
| Pax7b | Paired box protein Pax-7b |
| MyoD | Myoblast determination protein |
| Myf5 | Myogenic factor 5 |
References
- Sandow, A. Skeletal Muscle. Annu. Rev. Physiol. 1970, 32, 87–138. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Suh, Y.; Shin, S.; Lee, K. Skeletal Muscle Characterization of Japanese Quail Line Selectively Bred for Lower Body Weight as an Avian Model of Delayed Muscle Growth with Hypoplasia. PLoS ONE 2014, 9, e95932. [Google Scholar] [CrossRef] [PubMed]
- Biga, P.R.; Goetz, F.W. Zebrafish and giant danio as models for muscle growth: Determinate vs. indeterminate growth as determined by morphometric analysis. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R1327–R1337. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Rudnicki, M.A. Satellite cells, the engines of muscle repair. Nat. Rev. Mol. Cell Biol. 2011, 13, 127–133. [Google Scholar] [CrossRef]
- Ruparelia, A.A.; Ratnayake, D.; Currie, P.D. Stem cells in skeletal muscle growth and regeneration in amniotes and teleosts: Emerging themes. Wiley Interdiscip. Rev. Dev. Biol. 2019, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A. Satellite Cell of Skeletal Muscle Fibers. J. Cell Biol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Chargé, S.B.; Seale, P.; Huh, M.; Rudnicki, M.A. Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis. J. Cell Biol. 2006, 172, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, M. Skeletal muscle progenitor cells and the role of Pax genes. Comptes Rendus Biol. 2007, 330, 530–533. [Google Scholar] [CrossRef]
- Buckingham, M.; Relaix, F. The Role ofPaxGenes in the Development of Tissues and Organs: Pax3 and Pax7 Regulate Muscle Progenitor Cell Functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 645–673. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, K.; Jie, M.; Xiangbing, B.; Jun, H. PAX3+ skeletal muscle satellite cells retain long-term self-renewal and proliferation. Muscle Nerve 2016, 54, 943–951. [Google Scholar] [CrossRef]
- Kassar-Duchossoy, L.; Giacone, E.; Gayraud-Morel, B.; Jory, A.; Gomès, D.; Tajbakhsh, S. Pax3/Pax7 mark a novel population of primitive myogenic cells during development. Genes Dev. 2005, 19, 1426–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, D.; Powell, S.K.; Plummer, R.S.; Young, K.P.; Ruggeri, B.A. PAX genes: Roles in development, pathophysiology, and cancer. Biochem. Pharmacol. 2007, 73, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gruss, P.; Walther, C. Pax in development. Cell 1992, 69, 719–722. [Google Scholar] [CrossRef]
- Paixão-Côrtes, V.R.; Salzano, F.M.; Bortolini, M.C. Origins and evolvability of the PAX family. Semin. Cell Dev. Biol. 2015, 44, 64–74. [Google Scholar] [CrossRef]
- Tremblay, P.; Dietrich, S.; Mericskay, M.; Schubert, F.R.; Li, Z.; Paulin, D. A Crucial Role for Pax3 in the Development of the Hypaxial Musculature and the Long-Range Migration of Muscle Precursors. Dev. Biol. 1998, 203, 49–61. [Google Scholar] [CrossRef]
- Schubert, F.R.; Tremblay, P.; Mansouri, A.; Faisst, A.M.; Kammandel, B.; Lumsden, A.; Gruss, P.; Dietrich, S. Early mesodermal phenotypes in splotch suggest a role for Pax3 in the formation of epithelial somites. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2001, 222, 506–521. [Google Scholar]
- Crist, C.G.; Montarras, D.; Pallafacchina, G.; Rocancourt, D.; Cumano, A.; Conway, S.J.; Buckingham, M. Muscle stem cell behavior is modified by microRNA-27 regulation of Pax3 expression. Proc. Natl. Acad. Sci. USA 2009, 106, 13383–13387. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, A.; Gruss, P. Pax3 and Pax7 are expressed in commissural neurons and restrict ventral neuronal identity in the spinal cord. Mech. Dev. 1998, 78, 171–178. [Google Scholar] [CrossRef]
- Le Grand, F.; A Rudnicki, M. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [Green Version]
- Maroto, M.; Reshef, R.; Münsterberg, A.E.; Koester, S.; Goulding, M.; Lassar, A.B. Ectopic Pax-3 activates MyoD and Myf-5 ex-pression in embryonic mesoderm and neural tissue. Cell 1997, 89, 139–148. [Google Scholar] [CrossRef]
- Bajard, L.; Relaix, F.; Lagha, M.; Rocancourt, D.; Daubas, P.; Buckingham, M.E. A novel genetic hierarchy functions during hypaxial myogenesis: Pax3 directly activates Myf5 in muscle progenitor cells in the limb. Genes Dev. 2006, 20, 2450–2464. [Google Scholar] [CrossRef] [Green Version]
- De Morree, A.; Klein, J.D.D.; Gan, Q.; Farup, J.; Urtasun, A.; Kanugovi, A.; Bilen, B.; Van Velthoven, C.T.J.; Quarta, M.; Rando, T.A. Alternative polyadenylation of Pax3 controls muscle stem cell fate and muscle function. Science 2019, 366, 734–738. [Google Scholar] [CrossRef]
- Der Vartanian, A.; Quétin, M.; Michineau, S.; Auradé, F.; Hayashi, S.; Dubois, C.; Rocancourt, D.; Drayton-Libotte, B.; Szegedi, A.; Buckingham, M.; et al. PAX3 Confers Functional Heterogeneity in Skeletal Muscle Stem Cell Responses to Environmental Stress. Cell Stem Cell 2019, 24, 958–973.e9. [Google Scholar] [CrossRef]
- Relaix, F.; Montarras, D.; Zaffran, S.; Gayraud-Morel, B.; Rocancourt, D.; Tajbakhsh, S.; Mansouri, A.; Cumano, A.; Buckingham, M. Pax3 and Pax7 have distinct and overlapping functions in adult muscle progenitor cells. J. Cell Biol. 2005, 172, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 Is Required for the Specification of Myogenic Satellite Cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Oustanina, S.; Hause, G.; Braun, T. Pax7 directs postnatal renewal and propagation of myogenic satellite cells but not their specification. EMBO J. 2004, 23, 3430–3439. [Google Scholar] [CrossRef] [Green Version]
- Berberoglu, M.A.; Gallagher, T.L.; Morrow, Z.T.; Talbot, J.C.; Hromowyk, K.J.; Tenente, I.M.; Langenau, D.M.; Amacher, S.L. Satellite-like cells contribute to pax7-dependent skeletal muscle repair in adult zebrafish. Dev. Biol. 2017, 424, 162–180. [Google Scholar] [CrossRef] [PubMed]
- Noden, D.M. Cell movements and control of patterned tissue assembly during craniofacial development. J. Craniofacial Genet. Dev. Boil. 1991, 11, 192–213. [Google Scholar]
- Trainor, A.P.; Tan, S.S.; Tam, P.P. Cranial paraxial mesoderm: Regionalisation of cell fate and impact on craniofacial development in mouse embryos. Development 1994, 120, 2397–2408. [Google Scholar]
- Gros, J.; Manceau, M.; Thomé, V.; Marcelle, C. A common somitic origin for embryonic muscle progenitors and satellite cells. Nature 2005, 435, 954–958. [Google Scholar] [CrossRef]
- Ordahl, C.P.; Williams, B.A.; Denetclaw, W. Determination and morphogenesis in myogenic progenitor cells: An experimental embryological approach. Curr. Top. Dev. Biol. 1999, 48, 319–367. [Google Scholar] [CrossRef]
- Hollway, G.E.; Bryson-Richardson, R.J.; Berger, S.; Cole, N.J.; Hall, T.E.; Currie, P.D. Whole-Somite Rotation Generates Muscle Progenitor Cell Compartments in the Developing Zebrafish Embryo. Dev. Cell 2007, 12, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Lepper, C.; Conway, S.J.; Fan, C.-M. Adult satellite cells and embryonic muscle progenitors have distinct genetic requirements. Nat. Cell Biol. 2009, 460, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Chang, Y.; Bao, L.; Yu, M.; Li, R.; Niu, J.; Fan, G.; Song, W.; Seim, I.; Qin, Y.; et al. A chromosome-level genome of black rockfish, Sebastes schlegelii, provides insights into the evolution of live birth. Mol. Ecol. Resour. 2019, 19, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.C.; Schultz, E. The distribution of satellite cells and their relationship to specific fiber types in soleus and extensor digitorum longus muscles. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1982, 202, 329–337. [Google Scholar] [CrossRef]
- Holland, L.Z.; Schubert, M.; Kozmik, Z.; Holland, N.D. AmphiPax3/7, an amphioxus paired box gene: Insights into chordate myogenesis, neurogenesis, and the possible evolutionary precursor of definitive vertebrate neural crest. Evol. Dev. 1999, 1, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Tan, X.; Wang, Q.; Li, M.; Du, S.J. The olive flounder (Paralichthys olivaceus) Pax3 homologues are highly conserved, encode multiple isoforms and show unique expression patterns. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 180, 7–15. [Google Scholar] [CrossRef]
- Akolkar, D.B.; Asaduzzaman, M.; Kinoshita, S.; Asakawa, S.; Watabe, S. Characterization of Pax3 and Pax7 genes and their expression patterns during different development and growth stages of Japanese pufferfish Takifugu rubripes. Gene 2016, 575, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Tan, X.; Jun, S.Y.; Feng, M.; Jiao, S. The duplicated paired box protein 7 (pax7) genes differentially transcribed during Japanese flounder (Paralichthys olivaceus) embryogenesis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 189, 62–68. [Google Scholar]
- Treisman, J.; Harris, E.; Desplan, C. The paired box encodes a second DNA-binding domain in the paired homeo domain protein. Genes Dev. 1991, 5, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulding, M.D.; Chalepakis, G.; Deutsch, U.; Erselius, J.R.; Gruss, P. Pax-3, a novel murine DNA binding protein expressed during early neurogenesis. EMBO J. 1991, 10, 1135–1147. [Google Scholar] [CrossRef]
- Goulding, M.; Lumsden, A.; Paquette, A.J. Regulation of Pax-3 expression in the dermomyotome and its role in muscle devel-opment. Development 1994, 120, 957–971. [Google Scholar]
- Jostes, B.; Walther, C.; Gruss, P. The murine paired box gene, Pax7, is expressed specifically during the development of the nervous and muscular system. Mech. Dev. 1990, 33, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-C.; Sætre, B.O.; Håvik, B.; Ellingsen, S.; Fjose, A. The zebrafish Pax3 and Pax7 homologues are highly conserved, encode multiple isoforms and show dynamic segment-like expression in the developing brain. Mech. Dev. 1998, 70, 49–63. [Google Scholar] [CrossRef]
- Neal, A.; Boldrin, L.; Morgan, J.E. The Satellite Cell in Male and Female, Developing and Adult Mouse Muscle: Distinct Stem Cells for Growth and Regeneration. PLoS ONE 2012, 7, e37950. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.A.; Zembrzycki, A.; Mansouri, A.; Ziman, M. Pax7 is requisite for maintenance of a subpopulation of superior col-licular neurons and shows a diverging expression pattern to Pax3 during superior collicular development. BMC Dev. Biol. 2008, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Maczkowiak, F.; Mateos, S.; Wang, E.; Roche, D.; Harland, R.; Monsoro-Burq, A.H. The Pax3 and Pax7 paralogs cooperate in neural and neural crest patterning using distinct molecular mechanisms, in Xenopus laevis embryos. Dev. Biol. 2010, 340, 381–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christ, B.; Ordahl, C.P. Early stages of chick somite development. Beiträge Ref. Anat. Entwickelungsgeschichte 1995, 191, 381–396. [Google Scholar] [CrossRef]
- Prisk, V.; Huard, J. Muscle injuries and repair: The role of prostaglandins and inflammation. Histol. Histopathol. 2003, 18, 14. [Google Scholar]
- Tierney, M.T.; Stec, M.J.; Rulands, S.; Simons, B.D.; Sacco, A. Muscle Stem Cells Exhibit Distinct Clonal Dynamics in Response to Tissue Repair and Homeostatic Aging. Cell Stem Cell 2018, 22, 119–127.e3. [Google Scholar] [CrossRef] [Green Version]
- Chargé, S.B.P.; Rudnicki, M.A. Cellular and Molecular Regulation of Muscle Regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef]
- Tian, Z.-L.; Jiang, S.-K.; Zhang, M.; Wang, M.; Li, J.-Y.; Zhao, R.; Wang, L.-L.; Li, S.-S.; Liu, M.; Zhang, M.-Z.; et al. Detection of satellite cells during skeletal muscle wound healing in rats: Time-dependent expressions of Pax7 and MyoD in relation to wound age. Int. J. Leg. Med. 2015, 130, 163–172. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2001, 30, 2275–2729. [Google Scholar] [CrossRef] [Green Version]
- Lalitha, S. Primer premier 5. Biotech. Softw. Internet Rep. Comput. Softw. J. Sci. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Du, X.; Wang, H.; Jin, C.; Gao, C.; Liu, J.; Zhang, Q. Comparative studies on duplicated tdrd7 paralogs in teleosts: Mo-lecular evolution caused neo-functionalization. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 347–357. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence |
|---|---|
| SsPax3a-qRT-PCR-Fw | GGGTCGTATTCTCATGGTTATC |
| SsPax3a-qRT-PCR-Rv | GGTTGCAGTGGTTTCACTA |
| SsPax3b-qRT-PCR-Fw | CCATTCACCAGGACACATT |
| SsPax3b-qRT-PCR-Rv | GCTGCACCAACCTCATTA |
| SsPax7a-qRT-PCR-Fw | TGTACTTTGACCTTGCTGTT |
| SsPax7a-qRT-PCR-Rv | CTCATCATCCAGTCGTGTTT |
| SsPax7b-qRT-PCR-Fw | TCAGCTGTACTGGGACTG |
| SsPax7b-qRT-PCR-Rv | ACACACACACACACACAC |
| SsEif5a1-qRT-PCR-Fw | CTTTGCTCTGGTTCCTGAGTGG |
| SsEif5a1-qRT-PCR-Rv | AGCTTTGACATGCTGGGGTG |
| SsPax3a-ISH-Fw | ATTTAGGTGACACTATAGAAGAGGACAGAAAGACAAGAGGTACAG |
| SsPax3a-ISH-Rv | TAATACGACTCACTATAGGGAGACCATTAAGCGGCTATTGTAAAC |
| SsPax3b-ISH-Fw | ATTTAGGTGACACTATACGCGGATCCAGGCATCACTTCAAACATGG |
| SsPax3b-ISH-Rv | TAATACGACTCACTATAGCGAGCTCCCTCATTTGTCTCCACCTT |
| SsPax7a-ISH-Fw | ATTTAGGTGACACTATAGAAGAGGGACAGGCCTACTAAAG |
| SsPax7a-ISH-Rv | TAATACGACTCACTATAGGGAGACCATGACAAACAGGAAC |
| SsPax7b-ISH-Fw | ATTTAGGTGACACTATAGAAGAGACATACAGCACAACCAG |
| SsPax7b-ISH-Rv | TAATACGACTCACTATAGGGAGACAGCTGCATCATTCAATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Song, W.; Jin, C.; Huang, K.; Yu, Q.; Qi, J.; Zhang, Q.; He, Y. Pax3 and Pax7 Exhibit Distinct and Overlapping Functions in Marking Muscle Satellite Cells and Muscle Repair in a Marine Teleost, Sebastes schlegelii. Int. J. Mol. Sci. 2021, 22, 3769. https://doi.org/10.3390/ijms22073769
Wang M, Song W, Jin C, Huang K, Yu Q, Qi J, Zhang Q, He Y. Pax3 and Pax7 Exhibit Distinct and Overlapping Functions in Marking Muscle Satellite Cells and Muscle Repair in a Marine Teleost, Sebastes schlegelii. International Journal of Molecular Sciences. 2021; 22(7):3769. https://doi.org/10.3390/ijms22073769
Chicago/Turabian StyleWang, Mengya, Weihao Song, Chaofan Jin, Kejia Huang, Qianwen Yu, Jie Qi, Quanqi Zhang, and Yan He. 2021. "Pax3 and Pax7 Exhibit Distinct and Overlapping Functions in Marking Muscle Satellite Cells and Muscle Repair in a Marine Teleost, Sebastes schlegelii" International Journal of Molecular Sciences 22, no. 7: 3769. https://doi.org/10.3390/ijms22073769