 Gabriella Marincola1*
Gabriella Marincola1* Olivia Liong1
Olivia Liong1 Christoph Schoen2
Christoph Schoen2 Alaa Abouelfetouh3,4
Alaa Abouelfetouh3,4 Aisha Hamdy3
Aisha Hamdy3 Freya D. R. Wencker1
Freya D. R. Wencker1 Tessa Marciniak1,5
Tessa Marciniak1,5 Karsten Becker6
Karsten Becker6 Robin Köck7,8
Robin Köck7,8 Wilma Ziebuhr1*
Wilma Ziebuhr1*- 1Institute of Molecular Infection Biology, University of Würzburg, Würzburg, Germany
- 2Institute of Hygiene and Microbiology, University of Würzburg, Würzburg, Germany
- 3Department of Microbiology and Immunology, Faculty of Pharmacy, Alexandria University, Alexandria, Egypt
- 4Department of Microbiology and Immunology, Faculty of Pharmacy, AlAlamein International University, AlAlamein, Egypt
- 5Helmholtz Institute for RNA-based Infection Research (HIRI), Helmholtz Centre for Infection Research (HZI), Würzburg, Germany
- 6Friedrich Loeffler-Institute of Medical Microbiology, University Medicine Greifswald, Greifswald, Germany
- 7Deutsches Rotes Kreuz (DRK) Kliniken Berlin, Berlin, Germany
- 8Institute of Hygiene, University Hospital Münster, Münster, Germany
Coagulase-negative staphylococci (CoNS) are common opportunistic pathogens, but also ubiquitous human and animal commensals. Infection-associated CoNS from healthcare environments are typically characterized by pronounced antimicrobial resistance (AMR) including both methicillin- and multidrug-resistant isolates. Less is known about AMR patterns of CoNS colonizing the general population. Here we report on AMR in commensal CoNS recovered from 117 non-hospitalized volunteers in a region of Germany with a high livestock density. Among the 69 individuals colonized with CoNS, 29 had reported contacts to either companion or farm animals. CoNS were selectively cultivated from nasal swabs, followed by species definition by 16S rDNA sequencing and routine antibiotic susceptibility testing. Isolates displaying phenotypic AMR were further tested by PCR for presence of selected AMR genes. A total of 127 CoNS were isolated and Staphylococcus epidermidis (75%) was the most common CoNS species identified. Nine isolates (7%) were methicillin-resistant (MR) and carried the mecA gene, with seven individuals (10%) being colonized with at least one MR-CoNS isolate. While resistance against gentamicin, phenicols and spectinomycin was rare, high resistance rates were found against tetracycline (39%), erythromycin (33%) and fusidic acid (24%). In the majority of isolates, phenotypic resistance could be associated with corresponding AMR gene detection. Multidrug-resistance (MDR) was observed in 23% (29/127) of the isolates, with 33% (23/69) of the individuals being colonized with MDR-CoNS. The combined data suggest that MR- and MDR-CoNS are present in the community, with previous animal contact not significantly influencing the risk of becoming colonized with such isolates.
Introduction
Antimicrobial resistance (AMR) in bacteria is an increasing public health issue jeopardizing many achievements of modern medicine (1). Accordingly, monitoring the resistance situation in major human and veterinary pathogens such as enterobacteria, non-fermenters or Staphylococcus aureus is in the focus of surveillance programs. Far less attention however is paid to commensal, low pathogenic and environmental microorganisms, which may carry AMR genes as well. In this context, commensal and environmental bacteria are considered to play a role as putative AMR gene reservoirs that may fuel the resistance gene pool of more pathogenic bacteria through horizontal gene transfer (HGT) (2–4). Also, under certain conditions, these bacteria may be selected and emerge as opportunistic pathogens in their own right. A prime example for the dual role of commensals are coagulase-negative staphylococci (CoNS), which form a significant part of the skin and mucosa microbiota of warm blooded hosts, but also represent classical opportunistic pathogens that have been established as common causes of numerous healthcare-associated infections (5–7). Nosocomial CoNS are particularly notorious for readily acquiring numerous resistance traits, resulting in (multidrug-)resistance toward many commonly used antimicrobials. In addition, some species (e.g., Staphylococcus epidermidis) are capable of forming biofilms on indwelling medical devices, making CoNS infections sometimes extremely difficult to treat (8). While the detection and spread of multidrug-resistant (MDR) CoNS in hospital settings is well-documented (5, 9), we currently have less information on the resistance situation in CoNS outside of medical facilities. Previously, we performed a study on AMR in CoNS recovered from dust and manure samples in pig farms with a previous history of livestock-associated (LA) methicillin-resistant S. aureus (MRSA) detection (10). Unexpectedly, we found high AMR rates in CoNS from environmental samples, including resistance traits against last resort antibiotics such as oxazolidinones and lipopeptides. The reason(s) for the high multidrug-resistance rate in this distinct CoNS collection remained elusive. So, it was speculated that the selective pressure by antibiotics, commonly used in industrialized pig farming, might have favored AMR development. Also, contact of these dust- and manure-derived CoNS with soil microorganisms and their intrinsic resistance gene pool was hypothesized to have facilitated AMR acquisition (10). Finally, it is conceivable that, regardless of the ecological niche they are residing in, CoNS might be generally prone to increased AMR carriage. In order to shed more light on the presence and spread of AMR in CoNS, we currently aim at investigating CoNS from various ecological origins. In this report, we focus on the AMR profiles of human commensal CoNS isolates in non-hospitalized volunteers from the general population in Germany. The isolates were recovered from nasal swabs obtained in a previous cohort study on nasal colonization by important human bacterial pathogens (11, 12). Using standard microbiological methods, we assessed the species distribution and AMR profiles of 127 human commensal CoNS isolates. As the cohort study was performed in a geographic region of Germany with high livestock and industrialized farming intensity, we also asked the question whether or not contact with animals may represent a risk factor for individual AMR-CoNS carriage. Together, the analysis revealed that AMR is widespread among human commensal CoNS, many of which detected as MDR resistant isolates, with animal contact not significantly influencing individual AMR carriage.
Methods
Sample Isolation, Isolate Recovery, and Species Identification
For the analysis, we referred to a previous cohort study in which 1,878 nasal swabs were obtained from non-hospitalized volunteers from the German general population (11, 12). Recruitment was done by asking persons for their agreement to voluntarily participate in the study; written informed consent was obtained prior to enrolment and ethical clearance was granted by the institutional review board of the Westphalian Wilhelms-University Münster (no. 2006-268-f-S) (11). For CoNS recovery, 65 nasal swabs from persons without animal contact and 52 with animal contact (five veterinarians, 11 farmers and 36 pet owners) were randomly selected for the analysis. Samples were recovered through enrichment in LB broth for 6 h at 37°C. Dilutions of the cultures were plated onto Columbia colistin-aztreonam blood agar (CAP, Oxoid, Germany) and incubated for 24 h at 37°C to select for Gram-positive bacteria and to obtain single colonies. The next day, six randomly selected colonies from each nasal swab were picked and patched onto chromogenic medium to differentiate between methicillin-resistant (MR) and methicillin-susceptible (MS) CoNS (CAMSA/MPK, Medco Diagnostika, Germany). Species identification of the isolates was done by 16S rDNA locus sequencing after PCR amplification using the primers listed in Supplementary Table 1. By this approach, a total of 176 CoNS arising from 69 nasal swabs were obtained for further analysis.
Antimicrobial Susceptibility Testing
MICs for oxacillin (OXA), gentamicin (GEN), levofloxacin (LEV), erythromycin (ERY), clindamycin (CLI), linezolid (LNZ), daptomycin (DAP), teicoplanin (TEC), vancomycin (VAN), tetracycline (TET), tigecycline (TIG), fosfomycin (FOS), fusidic acid (FUS), rifampicin (RIF), trimethoprim/sulfamethoxazole (TMP-SMX) were determined using the VITEK2 system (bioMérieux Deutschland GmbH, Nürtingen) according to standard procedures provided by the manufacturer (Vitek Card AST-P654). MIC results were evaluated through the Advanced Expert System (AESTM) according to EUCAST guidelines1 and clinical breakpoints for CoNS. Isolates arising from the same nasal swab, which were the same species and had the same VITEK antibiogram were considered as duplicates and only one isolate was included into further analyses. Antibiotic susceptibilities for apramycin (APR), spectinomycin (SPC), florfenicol (FFC), chloramphenicol (CM) and quinupristin-dalfopristin (QD) were performed by agar disk diffusion assays using disks with 15, 100, 30, 30, and 15 μg of the respective antimicrobial agent according to EUCAST guidelines. As no interpretive criteria applicable to staphylococci are available for APR, SPC, and FFC, inhibition zone distributions were determined. Isolates displaying reduced zone diameters were further tested by molecular analysis for the presence of the respective resistance genes (see below). Isolates which harbored a respective resistance gene were considered as resistant, even in absence of available interpretive criteria for the antibiotic.
Molecular Analysis of Resistance
CoNS displaying a resistant phenotype based on the VITEK or disk diffusion analyses were tested by PCR for presence of the respective resistance genes [oxacillin: mecA, mecB, mecC; chloramphenicol: cat194, cat221, cat223; florfenicol: fexA and fexB; spectinomycin: spc, spd and spw; apramycin: apmA; fusidic acid: fusB, fusD, fusC; gentamicin; aac(6')/aph(2”) and aadD; erythromycin: ermA, ermB and ermC; tetracycline: tetK/L and tetM], using the primers and conditions listed in Supplementary Table 1. Thus, isolates were cultured on CAP sheep blood agar and DNA was extracted using the NucleoSpin Tissue Kit (Macherey-Nagel, #740952) according to the manufacturer's protocol with the addition of 15 μl of lysostaphin (2 mg/ml) to the lysis buffer. Primers used for 16S rDNA amplification were included in each PCR reaction as a control for gDNA template integrity.
Statistical Analysis
Whenever appropriate, contingency analyses were performed using Fisher's exact test by employing the GraphPad Prism software package. Differences with p < 0.01 were considered statistically significant.
Results
CoNS Recovery and Species Determination
For CoNS recovery, 65 nasal swabs from persons without animal contact and 52 with animal contact (five veterinarians, 11 farmers and 36 pet owners) were randomly selected for the analysis. 176 CoNS (arising from 69 nasal swabs) were initially picked from selective media as described in Methods. Next, species and antibiograms of the 176 CoNS isolates were assessed. Isolates arising from the same nasal swab and displaying identical species and resistance profiles were considered as duplicates, with only one isolate being subjected to further analysis. This led to a final pool of 127 CoNS isolates obtained from 69 nasal swabs. In 34 swabs growth of only one CoNS isolate was detectable, while 35 samples exhibited simultaneous growth of more than one CoNS isolate (18 swabs displayed two CoNS isolates, 12 swabs showed three CoNS isolates, four swabs had four CoNS isolates and in one swab we found five different CoNS isolates). Species determination by 16S rDNA locus sequencing identified 75% of the isolates as S. epidermidis (95/127). The remaining 32 isolates represented five additional species (Figure 1). S. epidermidis was detected in the majority of swabs (61/69; 88%). In the 35 nasal swabs in which more than one CoNS isolate was recovered, at least one S. epidermidis isolate was present in the majority of samples (32/35; 91%), while the species was absent in only three swabs (3/35; 9%).
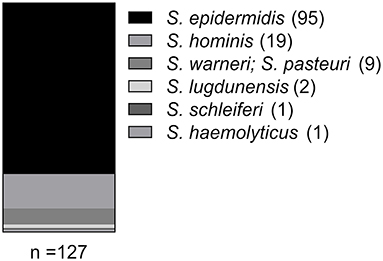
Figure 1. Species distribution among the CoNS isolates as identified by 16S rDNA locus sequence analysis.
Antibiotic Susceptibility and Resistance Genes Detection
The resistance phenotypes and AMR genes detected among the 127 CoNS isolates are summarized in Figure 2 and Table 1. Methicillin (oxacillin) resistance occurred in 7% (9/127) of the isolates (seven S. epidermidis and two S. hominis), with all isolates carrying the methicillin resistance conferring mecA gene (Table 1). None of the MR-CoNS harbored mecB or mecC. The most abundant resistance phenotypes in the collection were found toward tetracycline (39%; 49/127), fosfomycin (35%; 45/127), erythromycin (33%; 42/127) and fusidic acid (17%; 22/127) (Figure 2A). Gentamicin resistance occurred in five isolates (4%) all harboring the aminoglycoside resistance-mediating aac-aph gene. Except for one isolate, tetracycline resistance was conferred either by tetK/L (44/49) or tetM (4/49) (Table 1). With respect to erythromycin, all 42 resistant isolates harbored at least one of the macrolide resistance genes tested. The most abundant gene was ermA (24/42), followed by ermC (17/42) and ermB (11/42) (Table 1). Interestingly, eight isolates carried more than one of the macrolide resistance genes. Among the 22 fusidic acid resistant isolates, 19 tested positive for fusB and three for fusC (Table 1). For the antibiotics listed in Figure 2B, agar disk diffusion tests were performed. Based on zone diameter breakpoint definitions for quinupristin-dalfopristin (i.e., R <18 mm), all isolates were found to be susceptible to the streptogramin combination (Figure 2B). For spectinomycin, florfenicol, and apramycin no clinical interpretive criteria are currently defined for CoNS. Therefore, inhibition zone diameter distributions were determined and isolates displaying reduced inhibition zones were tested for resistance gene presence (Figures 2C–E). For spectinomycin, one isolate displayed no inhibition zone and carried the spectinomycin resistance-mediating spc gene (Table 1). Of the two isolates with reduced zone diameters toward florfenicol (<20 mm) one was found to harbor fexB (Table 1). Four isolates with reduced zone diameters (<19 mm) toward apramycin, were analyzed for the presence of apmA, but all lacked the resistance gene (Figure 2E). Finally, seven isolates displaying reduced chloramphenicol inhibition zone diameters (≤18 mm) were tested for the cat genes, revealing the detection of cat194 in all isolates (Table 1). Between individuals with and without animal contact, no statistically significant differences were recorded regarding AMR-CoNS carriage, although a slightly increased tendency for the detection of fosfomycin resistant isolates was observed in persons with reported animal contacts (Figure 3A).
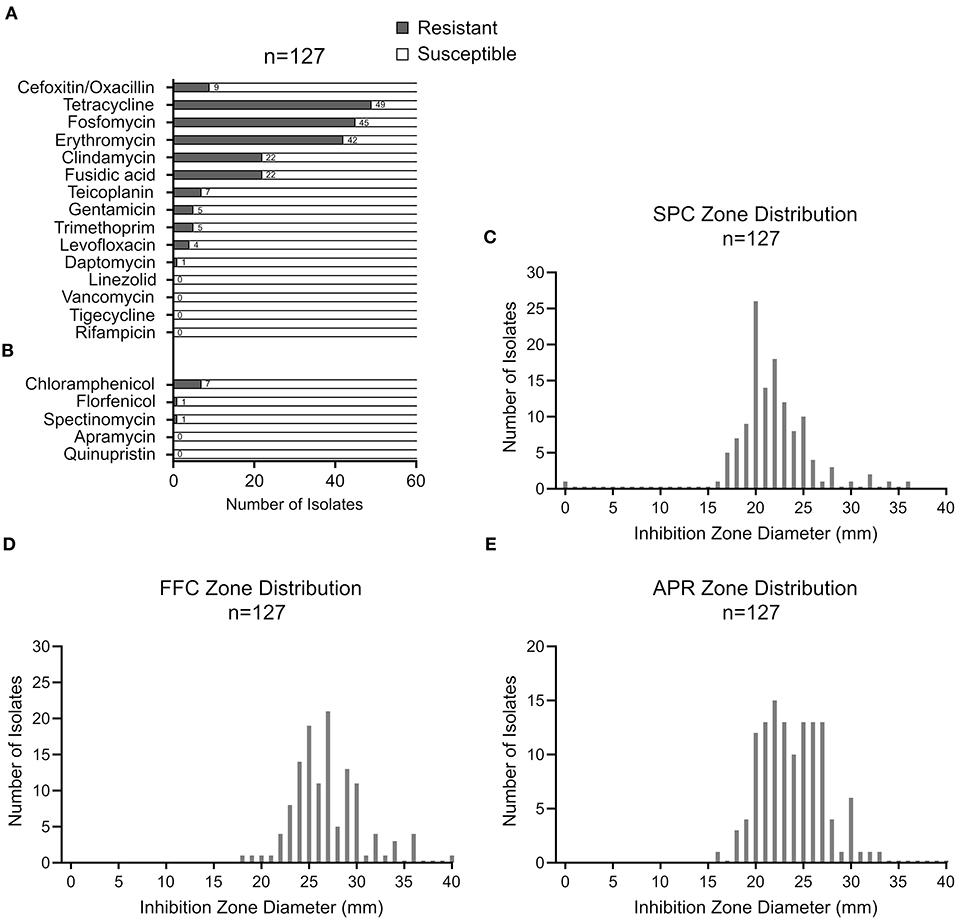
Figure 2. CoNS resistance profiles assessed by VITEK2 (A) or by agar disk diffusion (B). Inhibition zone diameter distributions for spectinomycin (C), florfenicol (D) and apramycin (E) with 100, 30, and 15 μg of the respective antimicrobial agent, respectively, according to EUCAST guidelines.
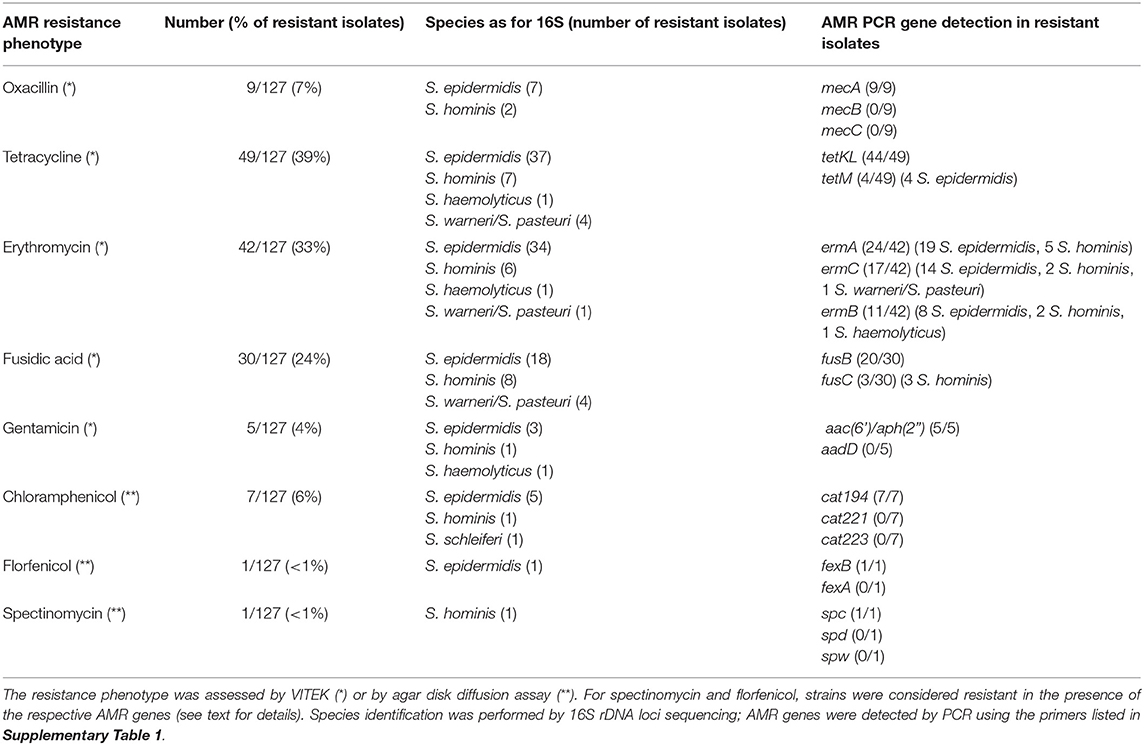
Table 1. Antimicrobial resistance gene detection among phenotypically resistant CoNS isolates.
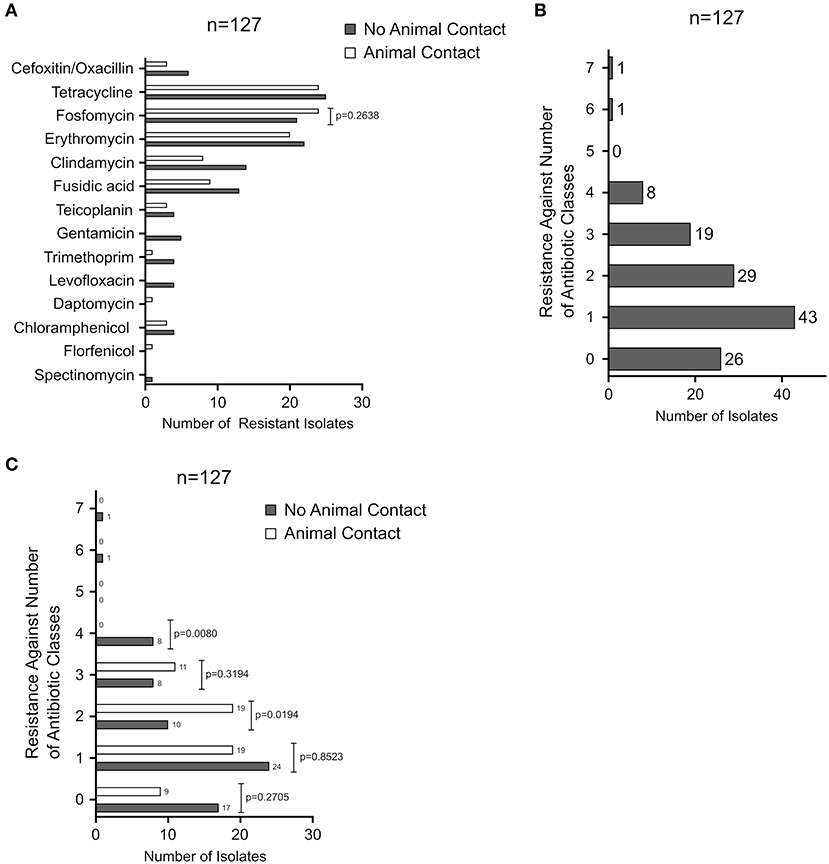
Figure 3. (A) Comparison of the number of resistant isolates arising from nasal swabs of individuals with animal contact (white bars) or without animal contact (gray bars). (B) Analysis of multidrug-resistant isolates. The number of resistant isolates is plotted against the simultaneous resistance toward 0–7 antibiotic classes. (C) Comparison of the number of multidrug-resistant isolates arising from nasal swabs of individuals with animal contact (white bars) or without animal contact (gray bars). Contingency analysis in (A,B) was performed using Fisher's Exact Test by employing the GraphPad Prism software package.
CoNS Multidrug-Resistance
Multidrug-resistance (defined as insusceptibility toward at least three antimicrobial classes) is a known hallmark and major issue particularly in nosocomial and infection-associated CoNS (5, 9). Analysis of our community-obtained commensal CoNS collection revealed that 20% (26/127) of the isolates were fully susceptible to all antimicrobials tested, while 57% (72/127) exhibited resistance toward one (43/127) or two (29/127) of the antibiotics tested (Figure 3B). Multidrug-resistance was recorded in 23% (29/127) of all isolates, from which 15% (19/127) displayed resistance against three and 6% (8/127) against four antibiotics. Two isolates were simultaneously resistant to six and seven of the antibiotics, respectively (Figure 3B). As shown in Figure 3C, previous animal contact of the volunteers did not significantly influence individual multidrug-resistant CoNS carriage (Figure 3C).
Individual Colonization by Resistant CoNS
Among the 69 CoNS-carrying individuals (40 with no and 29 with animal contact), seven (10%) were colonized by at least one mecA-positive MR-CoNS. The carriage rates were 5/40 and 2/29 for individuals with no or with animal contact, respectively (Fisher's Exact Test, p = 0.69, ns). Carriage of fully susceptible and of strains resistant to 1–2 antibiotic classes occurred in 12 (17%) and 34 (49%) of the 69 persons, respectively, from which 4 (6%) and 16 (23%) had previous animal contact (Fisher's Exact Test, p = 0.51, ns). Colonization by MDR-CoNS was overall detected in 23 individuals (33%). Among these, there were 9/29 individuals with animal contact vs. 14/40 without animal contact (Fisher Exact Test, p = 0.80, ns). The combined data suggest that MR- and MDR-CoNS are widely disseminated in the community with animal contact not significantly influencing the risk of becoming colonized by such isolates.
Discussion
AMR in bacterial pathogens continues to represent a major challenge for infection control. In order to tackle the problem holistically, the One Health concept, which takes humans, domestic and wild animals and the environment equally into account, is currently being pursued. Across these sectors, the approach also includes commensal and environmental bacteria to assess the risk factors for AMR development in pathogens (13). CoNS are typical skin and mucosa commensals which share the same ecological niche in the human anterior nares with S. aureus and many other bacteria (5, 7, 11), providing CoNS ample opportunity for horizontal gene transfer and the exchange of resistance genes (14). Indeed, CoNS have been identified as reservoirs and source of resistance traits that are transferred across the Staphylococcaceae family (15), including resistance genes against last resort antibiotics such as linezolid or daptomycin (10, 16). Across the geographic regions and infection sites, high resistance rates are common and typical among CoNS from health care settings (9, 17–20). In this report, we show that methicillin- and multidrug- resistant CoNS are also present in the community in healthy non-hospitalized volunteers. In staphylococci, methicillin/oxacillin resistance is of particular interest. It is mainly mediated by mecA (encoding an alternative penicillin-binding protein) located on transferrable SCCmec genomic elements whose genetic origins have been associated with Staphylococcus sciuri (now Mammaliicoccus sciuri) (21–24) and macrococcal species (25). SCCmec elements readily integrate other mobile genetic elements, and in addition to beta-lactam insusceptibility, they may therefore confer resistance to unrelated antibiotic classes too (15), making mecA-carriage a marker for multidrug- resistant isolates as well. Moreover, co-selection processes may favor the manifestation of multidrug resistance. In our study, mecA detection among the CoNS isolates was low (i.e., 7%) and only seven of the 69 individuals tested (10%) were colonized by at least one MR-CoNS isolate. These low mecA detection rates are in good agreement with previous reports on community-acquired commensal CoNS in Europe (26–28); but numbers may vary considerably in studies conducted in other geographic regions, with MR-CoNS rates ranging between 16 and 50% (29–33). We also noticed relatively high resistance rates toward tetracycline (39% 49/127) and erythromycin (33%; 42/127) in the sample (Figure 2). Macrolides are among the antibiotics most frequently prescribed on an outpatient basis and tetracyclines are commonly used in veterinary medicine. It is therefore well conceivable that the frequent detection of these resistances might be associated with a high selective pressure imposed by these antibiotics. However, as we lack concrete data on antibiotic consumption, this is currently mere speculation. We further found that the majority of isolates (i.e., 77%) are either completely susceptible to the tested antibiotics (20%, 26/127) or show resistance to a maximum of two antibiotic classes (57%, 72/127) (Figure 3B). Of note, however, in about a quarter of the strains (i.e., 23%), we detected multidrug-resistance toward three or more antibiotics, with two isolates even displaying simultaneous resistance against six and seven antibiotics, respectively (Figure 3B). Fortunately, we did not find any resistance to newer antibiotics such as linezolid or daptomycin in our sample, as described earlier in CoNS from animal and human sources (10, 16). Our results are supported by other studies reporting co- and multidrug-resistance not only in CoNS from humans but also among isolates from animals and the environment (34–41). We interpret these findings as an alarming signal for the continued introduction of (multidrug) resistant CoNS isolates into habitats outside of hospitals. Since the selective pressure by antibiotics is the main driving force behind the emergence and maintenance of resistant bacteria, it is tempting to speculate that the ubiquitous use of antimicrobial agents in human and veterinary medicine as well as in agriculture is the key factor behind this worrying development. In addition to their accepted role as resistance gene reservoirs for the more pathogenic S. aureus (15, 42, 43), MDR-CoNS selected in the community may pose a risk as opportunistic pathogens in immunocompromised patients when such strains are transferred into hospitals, highlighting the need for effective AMR surveillance also outside of the medical sector. In this respect, both farm and wild animals as well as pets are increasingly recognized as so far neglected source of MDR bacteria, including CoNS (40, 41, 44–50). Another question that we therefore wanted to answer with the study addressed the possible influence of animal contact on colonization rates with MDR-CoNS isolates. Interestingly, the data suggest that animal contact (both to farm and companion animals) did not increase the risk of becoming colonized by such isolates. However, our study was carried out in a region in northwest Germany with high livestock density and a previously proven LA-MRSA prevalence (11). It is well conceivable that the general population in this area is already increasingly exposed to resistant CoNS from animal husbandry, which would make it more difficult to reveal putative animal contact effects. A comparison of the CoNS resistance situation in a comparable region without extensive livestock husbandry would be helpful in order to finally answer this question. Together, the study shows that CoNS, including MDR strains, are present in the community, irrespective of a history of animal contact.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The studies involving human participants were reviewed and approved by Westphalian Wilhelms-University Münster (nr. 2006-268-f-S). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
GM, CS, AA, KB, RK, and WZ conceived and designed the experiments. GM and OL performed the experiments. GM, OL, AH, AA, FW, TM, and WZ analyzed the data. GM and WZ wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The study was supported by the German Research Council (DFG) through grant ZI665/3-1 to WZ as well as by the German Federal Ministry of Education and Research (BMBF), grant numbers 01KI1727A to KB and RK and 01KI1727E to WZ. This publication was supported by the Open Access Publication Fund of the University of Würzburg.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the Prevalence of Multiresistant Microorganisms (PMM) Study Group (O. Bindewald, T. T. Bui, C. Eckhoff, R. Epping, C. Fegeler, L. Kähmann, M. Meurer, M. Pobiruchin, J. Steger, L. von Auenmüller, and P. Werner) for sampling and data collection software development. We are grateful to students and technicians Antje Enders, Anja Hassing, Greta Jaschkowitz, Sabrina Kömm, and Martina Schulte for excellent technical assistance and to Beate Stirzel for supporting the VITEK analyses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpubh.2021.684456/full#supplementary-material
Footnotes
References
1. O'Neill J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. The Review on Antimicrobial Resistance. (2016). Available online at: https://wellcomecollection.org/works/thvwsuba (accessed March 17, 2021).
2. Allen HK, Donato J, Wang HH, Cloud-Hansen KA, Davies J, Handelsman J. Call of the wild: antibiotic resistance genes in natural environments. Nat Rev Microbiol. (2010) 8:251–9. doi: 10.1038/nrmicro2312
3. Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev. (2010) 74:417–33. doi: 10.1128/MMBR.00016-10
4. Nemeghaire S, Argudín MA, Feßler AT, Hauschild T, Schwarz S, Butaye P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet Microbiol. (2014) 171:342–56. doi: 10.1016/j.vetmic.2014.02.005
5. Becker K, Heilmann C, Peters G. Coagulase-negative staphylococci. Clin Microbiol Rev. (2014) 27:870–926. doi: 10.1128/CMR.00109-13
6. Morad Asaad A, Ansar Qureshi M, Mujeeb Hasan S. Clinical significance of coagulase-negative staphylococci isolates from nosocomial bloodstream infections. Infect Dis. (2016) 48:356–60. doi: 10.3109/23744235.2015.1122833
7. Heilmann C, Ziebuhr W, Becker K. Are coagulase-negative staphylococci virulent? Clin Microbiol Infect. (2019) 25:1071–80. doi: 10.1016/j.cmi.2018.11.012
8. Schilcher K, Horswill AR. Staphylococcal biofilm development: structure, regulation, and treatment strategies. Microbiol Mol Biol Rev. (2020) 84:e00026–19. doi: 10.1128/MMBR.00026-19
9. Widerström M. Significance of Staphylococcus epidermidis in health care-associated infections, from contaminant to clinically relevant pathogen: this is a wake-up call! J Clin Microbiol. (2016) 54:1679–81. doi: 10.1128/JCM.00743-16
10. Schoenfelder SMK, Dong Y, Feßler AT, Schwarz S, Schoen C, Köck R, et al. Antibiotic resistance profiles of coagulase-negative staphylococci in livestock environments. Vet Microbiol. (2017) 200:79–87. doi: 10.1016/j.vetmic.2016.04.019
11. Köck R, Werner P, Friedrich AW, Fegeler C, Becker K, Prevalence of Multiresistant Microorganisms (PMM) Study Group. Persistence of nasal colonization with human pathogenic bacteria and associated antimicrobial resistance in the German general population. New Microbes New Infect. (2016) 9:24–34. doi: 10.1016/j.nmni.2015.11.004
12. Becker K, Schaumburg F, Fegeler C, Friedrich AW, Köck R, Prevalence of Multiresistant Microorganisms PMM Study. Staphylococcus aureus from the German general population is highly diverse. Int J Med Microbiol. (2017) 307:21–27. doi: 10.1016/j.ijmm.2016.11.007
13. Dalton KR, Rock C, Carroll KC, Davis MF. One health in hospitals: how understanding the dynamics of people, animals, and the hospital built-environment can be used to better inform interventions for antimicrobial-resistant gram-positive infections. Antimicrob Resist Infect Control. (2020) 9:78. doi: 10.1186/s13756-020-00737-2
14. Otto M. Coagulase-negative staphylococci as reservoirs of genes facilitating MRSA infection: Staphylococcal commensal species such as Staphylococcus epidermidis are being recognized as important sources of genes promoting MRSA colonization and virulence. Bioessays. (2013) 35:4–11. doi: 10.1002/bies.201200112
15. John J, George S, Nori SRC, Nelson-Sathi S. Phylogenomic analysis reveals the evolutionary route of resistant genes in Staphylococcus aureus. Genome Biol Evol. (2019) 11:2917–26. doi: 10.1093/gbe/evz213
16. Cuny C, Arnold P, Hermes J, Eckmanns T, Mehraj J, Schoenfelder S, et al. Occurrence of cfr-mediated multiresistance in staphylococci from veal calves and pigs, from humans at the corresponding farms, and from veterinarians and their family members. Vet Microbiol. (2017) 200:88–94. doi: 10.1016/j.vetmic.2016.04.002
17. May L, Klein EY, Rothman RE, Laxminarayan R. Trends in antibiotic resistance in coagulase-negative staphylococci in the United States, 1999 to 2012. Antimicrob Agents Chemother. (2014) 58:1404–9. doi: 10.1128/AAC.01908-13
18. Protic D, Savic D, Andjelkovic D, Djukanovic N, Zdravkovic M, Djurasevic SF, et al. Nosocomial coagulase-negative staphylococci in Belgrade: between Scylla and Charybdis. J Infect Dev Ctries. (2016) 10:907–12. doi: 10.3855/jidc.7939
19. Asante J, Amoako DG, Abia ALK, Somboro AM, Govinden U, Bester LA, et al. Review of clinically and epidemiologically relevant coagulase-negative Staphylococci in Africa. Microb Drug Resist. (2020) 26:951–70. doi: 10.1089/mdr.2019.0381
20. Noshak MA, Rezaee MA, Hasani A, Mirzaii M. The role of the coagulase-negative Staphylococci (CoNS) in infective endocarditis; a narrative review from 2000 to 2020. Curr Pharm Biotechnol. (2020) 21:1140–53. doi: 10.2174/1389201021666200423110359
21. Madhaiyan M, Wirth JS, Saravanan VS. Phylogenomic analyses of the Staphylococcaceae family suggest the reclassification of five species within the genus Staphylococcus as heterotypic synonyms, the promotion of five subspecies to novel species, the taxonomic reassignment of five Staphylococcus species to Mammaliicoccus gen. nov., and the formal assignment of Nosocomiicoccus to the family Staphylococcaceae. Int J Syst Evol Microbiol. (2020) 70:5926–36. doi: 10.1099/ijsem.0.004498
22. Rolo J, Worning P, Nielsen JB, Sobral R, Bowden R, Bouchami O, et al. Evidence for the evolutionary steps leading to mecA -mediated β -lactam resistance in staphylococci. PLoS Genet. (2017) 13:e1006674. doi: 10.1371/journal.pgen.1006674
23. Rolo J, Worning P, Nielsen JB, Bowden R, Bouchami O, Damborg P, et al. Evolutionary origin of the staphylococcal cassette chromosome mec (SCCmec). Antimicrob Agents Chemother. (2017) 61:e02302–16. doi: 10.1128/AAC.02302-16
24. Miragaia M. Factors contributing to the evolution of mecA-Mediated β-lactam resistance in staphylococci: update and new insights from whole genome sequencing (WGS). Front Microbiol. (2018) 9:2723. doi: 10.3389/fmicb.2018.02723
25. Baba T, Kuwahara-Arai K, Uchiyama I, Takeuchi F, Ito T, Hiramatsu K. Complete genome sequence of Macrococcus caseolyticus strain JCSCS5402, [corrected] reflecting the ancestral genome of the human-pathogenic staphylococci. J Bacteriol. (2009) 191:1180–90. doi: 10.1128/JB.01058-08
26. Widerström M, Wiström J, Ek E, Edebro H, Monsen T. Near absence of methicillin-resistance and pronounced genetic diversity among Staphylococcus epidermidis isolated from healthy persons in northern Sweden. APMIS. (2011) 119:505–12. doi: 10.1111/j.1600-0463.2011.02757.x
27. Cavanagh JP, Wolden R, Heise P, Esaiassen E, Klingenberg C, Aarag Fredheim EG. Antimicrobial susceptibility and body site distribution of community isolates of coagulase-negative staphylococci. APMIS. (2016) 124:973–8. doi: 10.1111/apm.12591
28. Budri PE, Shore AC, Coleman DC, Kinnevey PM, Humpreys H, Fitzgerald-Hughes D. Observational cross-sectional study of nasal staphylococcal species of medical students of diverse geographical origin, prior to healthcare exposure: prevalence of SCCmec, fusC, fusB and the arginine catabolite mobile element (ACME) in the absence of sele. BMJ Open. (2018) 8:e020391. doi: 10.1136/bmjopen-2017-020391
29. Ruppé E, Barbier F, Mesli Y, Maiga A, Cojocaru R, Benkhalfat M, et al. Diversity of staphylococcal cassette chromosome mec structures in methicillin-resistant Staphylococcus epidermidis and Staphylococcus haemolyticus strains among outpatients from four countries. Antimicrob Agents Chemother. (2009) 53:442–9. doi: 10.1128/AAC.00724-08
30. Barbier F, Ruppé E, Hernandez D, Lebeaux D, Francois P, Felix B, et al. Methicillin-resistant coagulase-negative staphylococci in the community: high homology of SCCmec IVa between Staphylococcus epidermidis and major clones of methicillin-resistant Staphylococcus aureus. J. Infect. Dis. (2010) 202:270–81. doi: 10.1086/653483
31. Lebeaux D, Barbier F, Angebault C, Benmahdi L, Ruppé E, Felix B, et al. Evolution of nasal carriage of methicillin-resistant coagulase-negative staphylococci in a remote population. Antimicrob Agents Chemother. (2012) 56:315–23. doi: 10.1128/AAC.00547-11
32. He S, Lin J, Li Y, Zhang W, Zhou J, Han Z, et al. Insights into the epidemiology of methicillin-resistant coagulase-negative Staphylococci carriage in community-based drug users. J Infect Public Health. (2020) 13:1742–8. doi: 10.1016/j.jiph.2020.09.011
33. Kateete DP, Asiimwe BB, Mayanja R, Najjuka CF, Rutebemberwa E. Species and drug susceptibility profiles of staphylococci isolated from healthy children in Eastern Uganda. PLoS ONE. (2020) 15:e0229026. doi: 10.1371/journal.pone.0229026
34. Murugesan S, Perumal N, Mahalingam SP, Dilliappan SK, Krishnan P. Analysis of antibiotic resistance genes and its associated SCCmec types among nasal carriage of methicillin resistant coagulase negative staphylococci from community settings, Chennai, Southern India. J Clin Diagn Res. (2015) 9:DC01-5. doi: 10.7860/JCDR/2015/11733.6307
35. Xu Z, Mkrtchyan HV, Cutler RR. Antibiotic resistance and mecA characterization of coagulase-negative staphylococci isolated from three hotels in London, UK. Front Microbiol. (2015) 6:947. doi: 10.3389/fmicb.2015.00947
36. Xu Z, Shah HN, Misra R, Chen J, Zhang W, Liu Y, et al. The prevalence, antibiotic resistance and mecA characterization of coagulase negative staphylococci recovered from non-healthcare settings in London, UK. Antimicrob Resist Infect Control. (2018) 7:1–10. doi: 10.1186/s13756-018-0367-4
37. Xu Z, Liu S, Chen L, Liu Y, Tan L, Shen J, et al. Journal of global antimicrobial resistance antimicrobial resistance and molecular characterization of methicillin-resistant coagulase-negative staphylococci from public shared bicycles in Tianjin, China. Integr Med Res. (2019) 19:231–5. doi: 10.1016/j.jgar.2019.03.008
38. Seng R, Leungtongkam U, Thummeepak R, Chatdumrong W, Sitthisak S. High prevalence of methicillin-resistant coagulase-negative staphylococci isolated from a University environment in Thailand. Int Microbiol. (2017) 20:65–73. doi: 10.2436/20.1501.01.286
39. Seng R, Kitti T, Thummeepak R, Kongthai P, Leungtongkam U, Wannalerdsakun S, et al. Biofilm formation of methicillin-resistant coagulase negative staphylococci (MR-CoNS) isolated from community and hospital environments. PLoS ONE. (2017) 12:e0184172. doi: 10.1371/journal.pone.0184172
40. Gómez-Sanz E, Ceballos S, Ruiz-Ripa L, Zarazaga M, Torres C. Clonally diverse methicillin and multidrug resistant coagulase negative staphylococci are ubiquitous and pose transfer ability between pets and their owners. Front Microbiol. (2019) 10:485. doi: 10.3389/fmicb.2019.00485
41. Elnageh HR, Hiblu MA, Abbassi MS, Abouzeed YM, Ahmed MO. Prevalence and antimicrobial resistance of Staphylococcus species isolated from cats and dogs. Open Vet J. (2021) 10:452–6. doi: 10.4314/ovj.v10i4.13
42. Rossi CC, Souza-silva T, Araújo-alves AV, Giambiagi-demarval M. CRISPR-Cas systems features and the gene-reservoir role of coagulase-negative staphylococci. Front Microbiol. (2017) 8:1–9. doi: 10.3389/fmicb.2017.01545
43. Fišarová L, Pantuček R, Botka T, Doškar J. Variability of resistance plasmids in coagulase-negative staphylococci and their importance as a reservoir of antimicrobial resistance. Res Microbiol. (2019) 170:105–11. doi: 10.1016/j.resmic.2018.11.004
44. Wang Y, He T, Schwarz S, Zhao Q, Shen Z, Wu C, et al. International journal of medical microbiology multidrug resistance gene cfr in methicillin-resistant coagulase-negative staphylococci from chickens, ducks, and pigs in China. Int J Med Microbiol. (2013) 303:84–7. doi: 10.1016/j.ijmm.2012.12.004
45. He T, Wang Y, Schwarz S, Zhao Q, Shen J, Wu C. Genetic environment of the multi-resistance gene cfr in methicillin-resistant coagulase-negative staphylococci from chickens, ducks, and pigs in China. Int J Med Microbiol. (2014) 304:257–61. doi: 10.1016/j.ijmm.2013.10.005
46. Wendlandt S, Kadlec K, Feßler AT, Schwarz S. Identification of ABC transporter genes conferring combined pleuromutilin-lincosamide-streptogramin A resistance in bovine methicillin-resistant Staphylococcus aureus and coagulase-negative staphylococci. Vet Microbiol. (2015) 177:353–8. doi: 10.1016/j.vetmic.2015.03.027
47. Loncaric I, Kübber-Heiss A, Posautz A, Ruppitsch W, Lepuschitz S, Schauer B, et al. Characterization of mecC gene-carrying coagulase-negative Staphylococcus spp. isolated from various animals. Vet Microbiol. (2019) 230:138–44. doi: 10.1016/j.vetmic.2019.02.014
48. Mama OM, Ruiz-Ripa L, Lozano C, González-Barrio D, Ruiz-Fons JF, Torres C. High diversity of coagulase negative staphylococci species in wild boars, with low antimicrobial resistance rates but detection of relevant resistance genes. Comp Immunol Microbiol Infect Dis. (2019) 64:125–9. doi: 10.1016/j.cimid.2019.03.006
49. Ruiz-Ripa L, Feßler AT, Hanke D, Sanz S, Olarte C, Mistourath O, et al. Coagulase-negative staphylococci carrying cfr and PVL genes, and MRSA/MSSA-CC398 in the swine farm environment. Vet Microbiol. (2020) 243:108631. doi: 10.1016/j.vetmic.2020.108631
50. Teixeira IM, de Oliveira Ferreira E, de Araújo Penna B, Mello I, De Oliveira E, De Araújo B. Microbial pathogenesis dogs as reservoir of methicillin resistant coagulase negative staphylococci strains – A possible neglected risk. Microb Pthogenes. (2019) 135:103616. doi: 10.1016/j.micpath.2019.103616
Keywords: coagulase-negative staphylococci, antimicrobial resistance, One Health, community settings, Germany
Citation: Marincola G, Liong O, Schoen C, Abouelfetouh A, Hamdy A, Wencker FDR, Marciniak T, Becker K, Köck R and Ziebuhr W (2021) Antimicrobial Resistance Profiles of Coagulase-Negative Staphylococci in Community-Based Healthy Individuals in Germany. Front. Public Health 9:684456. doi: 10.3389/fpubh.2021.684456
Received: 23 March 2021; Accepted: 24 May 2021;
Published: 17 June 2021.
Edited by:
Ilana L. B. C. Camargo, University of São Paulo, BrazilReviewed by:
Hui-Min Neoh, National University of Malaysia, MalaysiaMarthie Magdaleen Ehlers, University of Pretoria, South Africa
Cemil Kurekci, Mustafa Kemal University, Turkey
Copyright © 2021 Marincola, Liong, Schoen, Abouelfetouh, Hamdy, Wencker, Marciniak, Becker, Köck and Ziebuhr. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriella Marincola, gabriella.marincola@uni-wuerzburg.de; gabriella@marincola.eu; Wilma Ziebuhr, w.ziebuhr@mail.uni-wuerzburg.de