 Sabine Prud'hon1,2
Sabine Prud'hon1,2 Samir Bekadar1,2
Samir Bekadar1,2 Agnès Rastetter1Justine Guégan1Florence Cormier-Dequaire1,2Lucette Lacomblez2,3Graziella Mangone1,2
Agnès Rastetter1Justine Guégan1Florence Cormier-Dequaire1,2Lucette Lacomblez2,3Graziella Mangone1,2 Hana You1,2Mailys Daniau1Yannick Marie1Hélène Bertrand1Suzanne Lesage1
Hana You1,2Mailys Daniau1Yannick Marie1Hélène Bertrand1Suzanne Lesage1 Sophie Tezenas Du Montcel4Mathieu Anheim5,6,7Alexis Brice1,8Fabrice Danjou1
Sophie Tezenas Du Montcel4Mathieu Anheim5,6,7Alexis Brice1,8Fabrice Danjou1 Jean-Christophe Corvol1,2* on behalf of the BADGE-PD study group1
Jean-Christophe Corvol1,2* on behalf of the BADGE-PD study group1- 1Sorbonne Université, INSERM UMRS 1127, CNRS UMR 7225, Institut du Cerveau et de la Moelle, ICM, Paris, France
- 2Assistance Publique Hôpitaux de Paris, Centre d'Investigation Clinique neurosciences, Department of Neurology, Hôpital Pitié-Salpêtrière, Paris, France
- 3Assistance Publique Hôpitaux de Paris, Department of Pharmacology, Hôpital Pitié-Salpêtrière, Paris, France
- 4Assistance Publique Hôpitaux de Paris, Sorbonne Université, INSERM, Institut Pierre Louis d'Epidémiologie et de Santé Publique, Hôpital Pitié-Salpêtrière, Paris, France
- 5Hôpitaux Universitaires de Strasbourg, Department of Neurology, Strasbourg, France
- 6Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC), UMR 7104 CNRS/Unistra, Inserm U1258, Illkirch, France
- 7Fédération de Médecine Translationnelle de Strasbourg, Université de Strasbourg, Strasbourg, France
- 8Assistance Publique Hôpitaux de Paris, Department of Genetics, Hôpital Pitié-Salpêtrière, Paris, France
Introduction: Impulse control disorders (ICDs) frequently complicate dopamine agonist (DA) therapy in Parkinson's disease (PD). There is growing evidence of a high heritability for ICDs in the general population and in PD. Variants on genes belonging to the reward pathway have been shown to account for part of this heritability. We aimed to identify new pathways associated with ICDs in PD.
Methods: Thirty-six Parkinsonian patients on DA therapy with (n = 18) and without ICDs (n = 18) matched on age at PD's onset, and gender was selected to represent the most extreme phenotypes of their category. Exome sequencing was performed, and variants with a strong functional impact in brain-expressed genes were selected. Allele frequencies and their distribution in genes and pathways were analyzed with single variant and SKAT-O tests. The 10 most associated variants, genes, and pathways were retained for replication in the Parkinson's progression markers initiative (PPMI) cohort.
Results: None of markers tested passed the significance threshold adjusted for multiple comparisons. However, the “Adenylate cyclase activating” pathway, one of the top associated pathways in the discovery data set (p = 1.6 × 10−3) was replicated in the PPMI cohort and was significantly associated with ICDs in a post hoc pooled analysis (combined p-value 3.3 × 10−5). Two of the 10 most associated variants belonged to genes implicated in cAMP and ERK signaling (rs34193571 in RasGRF2, p = 5 × 10−4; rs1877652 in PDE2A, p = 8 × 10−4) although non-significant after Bonferroni correction.
Conclusion: Our results suggest that genes implicated in the signaling pathways linked to G protein-coupled receptors participate to genetic susceptibility to ICDs in PD.
Introduction
Impulse control disorders (ICDs) and related behaviors are defined by the failure to resist an impulse to perform a self-rewarding act that will cause longer-term harm to oneself or others. Their prevalence is estimated between 1 and 3% in the general population while it raised up to 17% in treated Parkinsonian patients (1, 2). Common manifestations in Parkinson's disease (PD) are pathological gambling (PG), hypersexuality, and compulsive eating and buying. Clinical risk factors associated with ICDs in PD include younger age, younger age of PD onset, unmarried status, current smoking, novelty seeking traits, and family history of gambling (1, 3, 4). Above all, ICDs in PD are strongly associated with the dopamine replacement therapy as demonstrated by a similar prevalence of ICDs in untreated Parkinsonian patients as compared to healthy controls (5) and their independent association with dopaminergic drugs, particularly dopamine agonists (1).
A high heritability has been suggested for ICDs in the general population, estimated to 40–60% in twin studies for pathological gambling (6, 7) and in ICDs in PD patients (8). Genetic association studies have found associations with candidate variants in genes coding monoamine and glutamate receptors, transporters, and metabolism enzymes in both the general population (9, 10) and in PD (11–14). ICDs are also more frequent and more severe in PD patients with PRKN mutations (15). The reward pathway—involving ventral areas of basal ganglia and its related cortical structures—is thought to play a central role in the pathophysiology of ICDs as suggested in several studies (16, 17). However, these variants explain only a small part of the phenotype's variance, estimated to be 15–21% for pathological gambling in the general population (9).
Rare genetic variants or combinations of variants may explain this and new strategies that are being developed to explore genetic contribution to complex traits. For instance, significant genes in the course of diabetes mellitus or cystic fibrosis have been discovered by comparing exome data of extreme phenotype groups, reasonably supposed to be enriched in causal variants (18, 19). Moreover, when several genetic variants in a gene or group of genes may contribute to the trait, aggregation tests, by evaluating the cumulative effect of multiple variants, are particularly relevant. The power gained with this method, compared to a single test, allows considering a smaller sample (20).
In this proof-of-concept study, we propose an unbiased approach by comparing exome sequences of two extreme phenotypes of Parkinsonian patients with and without ICDs. Our main objective was to identify new genes and pathways associated to ICDs in PD.
Methods
Patients and Study Design
Patients were selected from a multicenter, national, case-control study named Behavioral ADdiction and GEnes in Parkinson's disease (BADGE-PD) conducted in France between 2012 and 2015 in 13 centers of the clinical research network for Parkinson's disease (NS-PARK/FCRIN) (21). Three hundred four patients were included with (n = 172) or without (n = 132) ICDs or related behaviors. Common inclusion criteria were diagnosis of PD according to the UK Parkinson's Disease Society brain bank (22) and age older than 30 years. The study consisted of one visit at the expert center for PD with a neurological and neuropsychological evaluation and a blood sample for DNA extraction. All patients were evaluated by a face-to-face interview with a neuropsychologist with the French PD behavioral scale, validated for the assessment of ICDs and related behaviors in PD (23). This scale explores and quantifies the different components of hyper- and hypo-dopaminergic behaviors in PD, each item being rated on a 5-point scale (severe disorder: 4, marked disorder: 3, moderate disorder: 2, mild disorder: 1, absence of disorder: 0). Patients with ICDs were defined as having two items with a score of ≥ 2 or at least one item with a score of ≥ 2 on the following items: eating behavior, creativity, hobbying, risk-taking behavior, compulsive shopping, punding, pathological gambling, and hypersexuality. Control PD patients were selected to have no more than one of the previous item with a score of 1. All patients had to be of European Caucasian ancestry for at least two generations, and patients with and without ICDs were matched for age and sex. In addition, patients in the control group had more than 5 years since PD diagnosis and were exposed to a dopamine agonist of at least 300 mg of levodopa equivalent daily dose (LEDD). LEDD were calculated as previously described (24). The study was conducted according to international good clinical practice guidelines and submitted to local regulatory and ethical committees. All patients signed an informed consent form prior any procedure. The study was sponsored by the Assistance Publique Hôpitaux de Paris.
For this ancillary study, we selected an extreme phenotype (EP) population corresponding to 36 cases: 18 PD patients with ICDs (cases) and 18 PD patients without ICDs (controls). Cases were selected in descending order of behavioral scale in PD's scores. The five following items were considered: eating behavior, creativity, compulsive shopping, pathological gambling, and hypersexuality. Controls were selected considering the same items if scoring 0 in all of them. Cases and controls were matched for gender and age at PD onset and had been treated with ropinirole or pramipexole, the two major dopamine agonists used in this cohort. When several controls were available for matching, the patient exposed to the highest dose of DA was selected. Finally, as mutations or risk genetic variants have been shown to be associated with neuropsychiatric symptoms in PD patients (15, 25), patients were screened for the G2019S variant on LRRK2 gene and for pathogenic mutations on PRKN gene.
Replication
For replication, we used the Parkinson's progression markers initiative (PPMI) data set (https://www.michaeljfox.org/ppmi-clinical-study, data downloaded on April 20, 2016), selecting cases and controls with similar criteria than for the hypothesis-generation cohort. In this cohort, ICD behaviors were screened by using the Questionnaire for Impulsive-Compulsive Disorders in Parkinson's disease (QUIP) (26) at each visit. Among the 431 de novo Parkinsonian patients included in the cohort, we selected as cases the patients who developed ICD (QUIP>0) during follow-up (n = 52) while treating with pramipexole or ropinirole (n = 45) with a QUIP = 0 at baseline (n = 30) and available exome sequencing data (n = 27). For controls, we selected patients with QUIP scores remaining null throughout the follow-up (n = 127) treated with ropinirole or pramipexole (n = 107) with a dose > 290 mg LEDD for more than 3 months (n = 29) and exome sequencing data available (n = 28). The 10 most associated genetic variants, genes, and pathways were selected for replication using the whole exome sequencing data.
Sequencing, Variant Calling, and Annotation
Exomes from the 36 selected patients were sequenced on an Illumina NextSeq 500 with the MedExome kit, and the resulting reads were aligned to the hg19 reference genome with BWA applying duplicate removal with Picard Tools MarkedDuplicates. We performed GATK base quality score recalibration, indel realignment, and SNP and INDEL discovery and genotyping across all samples simultaneously using HaplotypeCaller, followed by variant quality score recalibration according to the GATK Best Practices recommendation: GATK-HaplotypeCaller primary QC was applied at the variant level, excluding any variants that were not flagged as “PASS” by the VQSR algorithm, thus excluding variants outside the 95% sensitivity tranches (i.e., the lowest 5% of recalibrated quality scores). Finally, variants were annotated using the SnpEff tool and GENome MINing software (GEMINI, version 0.18.0) (27).
For all 36 samples, mean coverage on target was above 84, and 90% of targeted regions were covered at least at 30X.
Data Analysis
Genetic variants were filtered to keep variants likely to have a functional impact based on the Combined Annotation Dependent Depletion (CADD) score (28) as made available in GEMINI (27). Expression values of genes were obtained from the Brain eQTL Almanac (http://www.braineac.org). Variants with a CADD score ≥ 12.37 were kept for further analysis, according to published recommendations (29), if also lying in genes expressed in at least one region among putamen, substantia nigra, thalamus, frontal cortex, and temporal cortex. For a pathway to be considered expressed in these brain regions and kept for analysis, 95% or more of its genes should be expressed in at least one of the latter brain areas. Furthermore, pathways were kept for analysis if at least 80% of their genes were represented by at least one variant in the experiment while a minimum of two variants per gene was required for gene-wise test and a minimum allele count of 4 for single-variant testing. For population description, paired sample t-test, McNemar, and χ2 or Fisher's exact tests were used for group comparisons of quantitative and qualitative variables, using Statistica (version 9.1) software.
Association of single variants with the case/control status was tested using the likelihood ratio test as made available in the Efficient and Parallelizable Association Container Toolbox, version 3.2.6 (EPACTS). Group-wise tests were conducted using SKAT-O tests as made available in EPACTS on both genes and Reactome pathways (30, 31). The significance threshold was adjusted with Bonferroni's method regarding the number of tests ran: 6,953 for variants, 4,769 for genes, and 123 for pathways leading to a threshold for significance of 7.2 × 10−6, 1 × 10−5, and 4.1 × 10−4, respectively.
For the replication stage, combined p-values were obtained using the Fisher test when SKAT-O p-values were found to be nominally significant in both EP and PPMI cohorts. P-values were considered as significant when passing the Bonferroni-corrected significance threshold considering the number of tests run during the discovery and replication phases (6,963 for variants, 4,779 for genes, and 133 for pathways, resulting in a threshold of significance of 7.2 × 10−6, 1 × 10−5, and 3.8 × 10−4). Tests were adjusted on age at PD diagnosis and gender.
Results
Population
The demographic and clinical characteristics of the 36 selected patients with extreme phenotypes (cases, n = 18; controls, n = 18) are described in Table 1. Cases and controls were similar in terms of PD age onset, disease duration, motor severity (MDS-UPDRS 3 and Hoehn & Yahr stage), and cognitive assessment (MMSE). Cases had significantly higher scores than controls at the MDS-UPDRS part I, which was related to a higher score at item I.6 assessing addictive behaviors (respectively, 2.4 vs. 0.8, p = 5 × 10−3). DA doses were significantly higher in control subjects (370.3 vs. 258.7 mg/day, p = 3 × 10−4) due to the study design (controls were recruited based on the absence of ICD behaviors despite high doses of DA). Among the cases, all had at least two different ICDs, five patients had a maximum score of 4 for at least one item, and eight patients accumulated at least two types of ICDs with a score equal to 3. There was no difference in ropinirole/pramipexole or gender distributions regarding to the type of ICDs except for hypersexuality, which was exclusively present in males (see Supplementary Data 1).
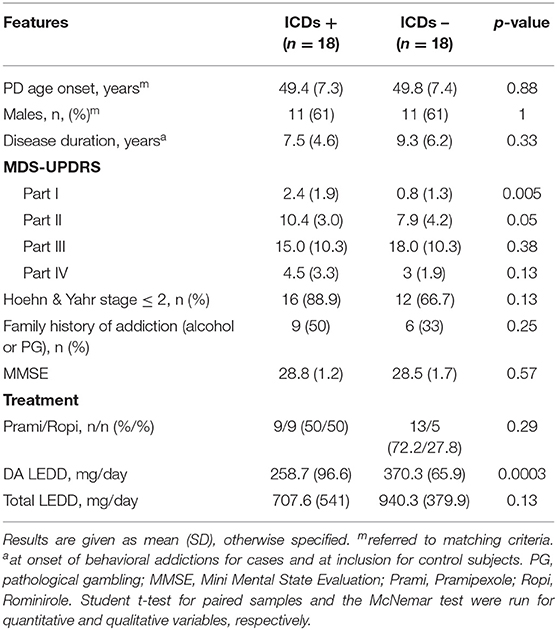
Table 1. Clinical features of the study population.
Single-Variant Analysis
After quality controls, 354,174 variants were available for further analyses. Among the 34,981 with a CADD-score ≥ 12.37, 26,418 satisfied the condition of a minimal occurrence of 4 in the cohort for the single test. After selection upon brain expression of genes, 6,953 variants remained for further analyses. The 10 most associated variants are shown in Table 2. Five variants were exonic missense—in PDCD6IP, RasGRF2, ACAN, COL12A1, and OMA1—and 4 variants were intronic—in GDA, PGK1, ITGA6, and PDE2A. The last variant was nonsense in the IL17RB gene. Two variants (rs34193571 RasGRF2, rs1877652 PDE2A) were involved in intracellular signal transduction, related to ERK, and cAMP signaling pathways. Three variants were laid on genes belonging to the pathway of focal adhesion (rs2293647, rs3743398, and rs970547 on ITGA6, ACAN, and COL12A1). No mutation was found in the LRRK2 or PRKN genes.
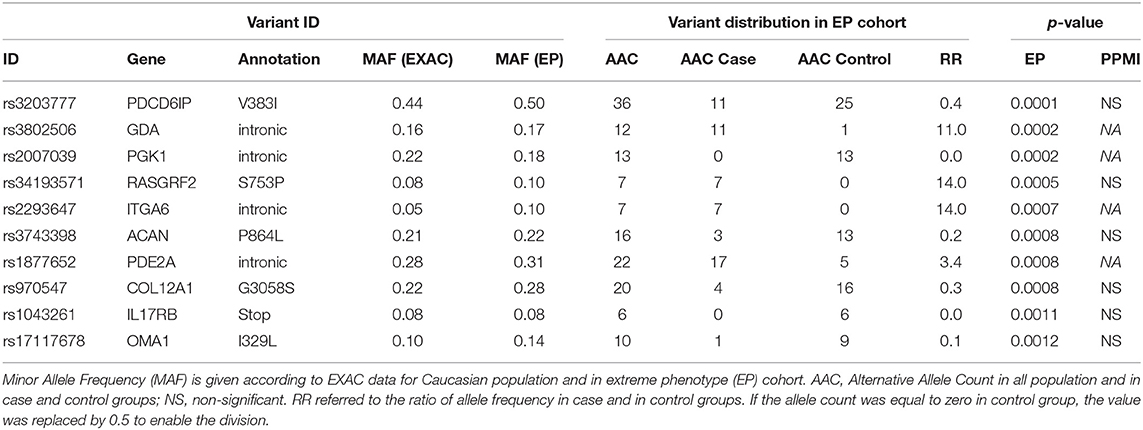
Table 2. Single test association study results (top 10 variants).
Genes Analysis
Of the 19,962 genes covered by the MedExome kit, 6,250 were represented by at least two variants with a CADD score ≥ 12.37. After selection upon brain expression, 4,769 genes remained for association testing. The 10 most associated genes upon SKAT-O test are shown in Table 3. Four of them, DOCK4, RasGRF2, ITGA6, and ITGA11, were linked to the ERK pathway. PDE2A belongs to cAMP signaling. For some genes, such as PDE2A and RasGRF2, association was led by only one frequent variant, which was strongly associated in the single test. For others, as for DOCK4 and MYH14, association resulted from different rare variants laying in this gene in different individuals of the cohort (see Supplementary Data 2).
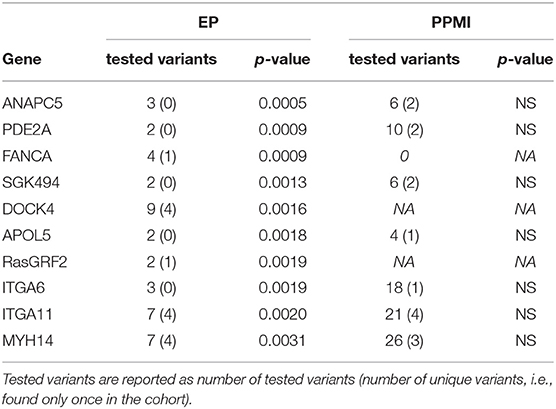
Table 3. SKAT-O on genes results (top 10 genes).
Pathways Analysis
Of the 1,816 pathways extracted from the Reactome database, 1,130 were represented by at least two variants with a CADD score ≥ 12.37. Among them, only 312 had more than 95% of their genes expressed in the brain's regions of interest, and the 123 that had more than 80% of their genes represented by at least one variant were considered for further analysis. The five most associated pathways using SKAT-O are shown in Table 4. The most associated one (R-HSA-446343, “Localization of the PINCH-ILK-PARVIN complex to focal adhesions” from Reactome database, p-value 1.52 × 10−3) was related to focal adhesions with a total of eight variants (two of which were unique, i.e., found only once in the cohort) tested. The second most associated pathway was the adenylate activating pathway (R-HSA-170660, “Adenylate cyclase activating pathway”, p = 1.56 × 10−3) with a total of 29 (10 of which were unique) variants tested distributed on eight genes.

Table 4. SKAT-O on pathways results (top 5 pathways).
None of the variants, genes, or pathways tested passed the significance threshold adjusted with Bonferroni's method (significance threshold of 7.2 × 10−6, 1 × 10−5, and 4.1 × 10−4 for a single test, SKAT-O on genes and pathways, respectively), in the discovery phase only.
Replication
The replication data set consisted of 59 patients (27 cases and 28 controls). Mean age at PD diagnosis was 58 years old in both groups. There were seven women in the cases and seven in the controls. Pramipexole was the most frequent DA in each group: 60 and 76% in cases and controls, respectively. Doses of DA were significantly higher in controls than in cases (372 LEDD vs. 193 mg/day LEDD, p < 0.001).
We applied the same analysis for the 10 most associated variants, genes, and pathways. Only the “adenylate cyclase activating pathway” replicated in the PPMI cohort (Table 4, p = 2 × 10−2 in the PPMI cohort and p = 1.56 × 10−3 in the EP cohort, resulting in a Fisher combined p-value of 3.3 × 10−5). Twenty-nine variants were tested in the EP and PPMI cohorts (see Supplementary Data 3 and 4). In the EP cohort, variants laid on eight different genes: ADCY 1, 3, 4, 5, 6, 8, 9, and GNAL. In the PPMI cohort, the nine genes encoding the adenylate cyclase from 1 to 9 were represented by at least one variant. Six variants of the 29 tested in each cohort were common to both cohorts, resulting in a total of 52 variants tested for this pathway. For five of them (rs3181385 on ADCY4, rs3730071, rs115315671, rs55770045 on ADCY6, and rs2228949 on ADCY8), the effect was observed in the same direction in both cohorts.
Discussion
In this study comparing whole exome sequences from two extreme phenotypes, we found enrichment of functional variants laying on brain-expressed genes of the “adenylate cyclase activating pathway” in PD patients with ICDs. This result is in accordance with a recent genome-wide association study founding an association between pathological gambling and the cAMP protein kinase signaling (32).
The dopamine replacement therapy that has been strongly associated with ICDs in PD patients activates dopamine receptors, 7-transmembrane domain G protein-coupled receptors (GPCR), regulating adenylate cyclase activity. D2-like receptors inhibit adenylyl cyclase, whereas stimulation of D1-like receptors leads to its activation through Galpha(olf) in the striatum (33). Cyclic-AMP then acts as a second messenger by activating the protein kinase A (PKA), which, in turn, phosphorylates several substrates and activates transcription factors. In the case of concomitant glutamatergic activation, the extracellular regulated kinase (ERK) MAPK pathway is also activated in medium spiny neurons in the striatum (34). These different routes converge to alteration in gene expression (35), which is supposed to underlie long-term neuronal plasticity induced by dopamine in the striatum (36). ERK activation in the ventral striatum has been shown to play a central role in the reward-associated pathway and in addictive behaviors related to alcohol (37) and psychostimulant consumption (38). In mice, activation of the ERK pathway in the dorsal striatum has been shown to underlie abnormal involuntary movements, dyskinesia, developed in response to chronic L-DOPA (39). In our study, the pathway directly related to cAMP signaling the “Adenylate cyclase activating pathway” was found to be associated with ICDs in both EP and PPMI cohorts.
Interestingly, although below the significance threshold, many of the most associated variants or genes tested were linked to the cAMP and ERK-dependent pathways.
The most associated variant in our study was a missense variant (rs3203777) in the PDCD6IP gene, also named DRIP4 for dopamine receptor interacting protein 4. This gene encodes a protein that has been shown to upregulate D1 and D3 receptor expression (40).
In addition, the rs1877652 variant, enriched in patients with ICDs, is an intronic variant of the PDE2A gene encoding the phosphodiesterase 2A controlling cAMP rates and is shown to play a key role in the modulation of the signal induced by dopamine in rodents (41). Seven patients with ICDs and none of the controls had a coding variant on RasGRF2 (rs34193571), previously associated with alcohol addiction (42), and coding a protein that mediates calcium-dependent activation of the ERK pathway, modulating the presynaptic effect on the activation of the basal ganglia mesolimbic pathway (43).
Several top associated variants, genes, and pathways were related to integrin systems, which also act on ERK signaling via downstream transducers, including the focal adhesion kinase, FAK. Integrin and its downstream effectors have been shown to be involved in structural changes that occurred in the dendritic tree within the addiction to alcohol (44) or cocaine process (45). A rare variant, rs2293647, lying in the integrin subunit α 6 gene ITGA6, was found only among cases (p = 7 × 10−4) while the rs3743398, a variant lying on the ACAN gene belonging to the aggrecan family, was found more frequently in controls, suggesting a protective role. The most associated pathway in the EP cohort was related to the integrin system although it was not replicated in the PPMI cohort.
None of these variants was previously found to be associated with ICDs in PD or in the general population. However, most previous studies focused on candidate gene coding receptors, transporters, or metabolism enzymes of the reward pathway but did not investigate their downstream signaling effectors (9–14).
The main limitation of our study is the small sample sizes of both our hypothesis generation and the replication data sets, which partly resulted from our stringent inclusion criteria (high doses of DA, severe ICDs, or no ICDs at all), and our results deserve further replication in larger cohorts. The underpowered analysis may explain that only one pathway was replicated and that most of the p-values obtained were far from reaching the adjustment for the multiple comparison threshold of significance. The lack of replicated results might also be due to differences between the two cohorts in terms of exome sequencing coverage or phenotypic definition of ICDs in the two cohorts. The selection of extreme cases strongly associated with DA-induced ICDs might have biased our results toward genes directly interacting with these drugs, which was the purpose of the design of the study. Our results also suggest that genetic susceptibility to ICDs in PD would rather result from the combination of multigenic variations than unique rare variants with strong effects, which was already suggested for ICDs in the general population (9).
This study highlights the potential important role of enrichment of functional variants in genes coding proteins of intracellular signaling cascades beyond neurotransmitter receptors in the genetic susceptibility of ICDs. Subjects with stronger signal transduction of GPCRs might be at higher risk to develop ICDs when exposed to dopaminergic therapy in PD. How these pathways interfere with drugs or increase the individual susceptibility to ICDs remain to be explored. The combination of clinical, neuroimaging, and exome sequencing data focused on the study of the polymorphism of the different pathways could help in providing some predictability, in the future, for ICDs in PD patients.
Data Availability Statement
Data are available upon request to any interested researcher or for the purpose of replicating our results (direct requests to jean-christophe.corvol@aphp.fr).
Ethics Statement
The studies involving human participants were reviewed and approved by CPP Paris VI. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
J-CC, FD, and SP contributed to the design of the work, data acquisition, interpretation of the data, and draft the manuscript. AR, HB, JG, and YM participated to data analysis. All authors contributed to the interpretation of the data, revised the manuscript for important intellectual content, provide approval for the publication, and agree to be accountable for all aspects of the work.
Funding
SB was funded by a grant from the program Investissements d'Avenir ANR-10-IAIHU-06. BADGE-PD study was sponsored by the Assistance Publique Hôpitaux de Paris, and funded by a grant of the French Ministry of Health (PHRC-2010_AOM10078). We thank the DNA and Cell bank of the ICM, the genetic platform of the ICM, and the NS-PARK/FCRIN network for their support in the conduct of this study. PPMI—a public-private partnership—is funded by the Michael J. Fox Foundation for Parkinson's Research and funding partners, including AbbVie, Avid Radiopharmaceuticals, Biogen, Bristol-Myers Squibb, Covance, GE Healthcare, Genentech, GlaxoSmithKline, Lilly, Lundbeck, Merck, Meso Scale Discovery, Pfizer, Piramal, Roche, Sanofi Genzyme, Servier, Takeda, Teva, UCB. This work received support from an European ERA-net grant (EraCoSysMed, PD-Strat).
Conflict of Interest
J-CC has served on advisory boards for Air Liquide, Biogen, Denali, Ever Pharma, Idorsia, Prevail Therapeutic, Theranexus; and received grants from Ipsen, the Michael J Fox Foundation, and Sanofi. MA received honoraria and travel grants from Johnson and Johnson, Actelion Pharmaceuticals, Teva, LVL, Aguettant, Orkyn, AbbVie.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
BADGE-PD study group: Steering committee: Jean-Christophe Corvol, Mathieu Anheim, Pierre-Michel Llorca, Lucette Lacomblez, Antoine Pelissolo, Jean-Luc Houeto, Bénédicte Lebrun-Vignes, Sophie Tezenas du Montcel; CHU Pitié-Salpêtrière (Paris): Jean-Christophe Corvol, Marie Vidailhet, Bertrand Degos, Florence Cormier-Dequaire, Lucette Lacomblez, Andreas Hartman, Anne-Marie Bonnet, David Grabli, Bertrand Degos, Emmanuel Roze, Graziella Mangone, Eve Benchetrit, Julie Socha, Said Lebbah; CHU Grenoble: Paul Krack, Eugénie Lhommée, Anna Castroto; CHU Clermont-Ferrand: Franck Durif; CHU Marseille: Jean-Philippe Azulay, Alexandre Eusebio, Tatiana Witjas; CHU Nantes: Pascal Derkinderen; CHU Bordeaux: François Tison, Wassilios Meissner; CHU Poitiers: Jean-Luc Houeto, Isabelle Benatru, Olivier, Colin, Anne Fradet, Anziza Manssouri, Emilie Rabois; CHU Toulouse: Olivier Rascol, Christine Brefel-Courbon, Fabienne Ory-Magne, Claire Thalamas, Monique Galistky, Fabienne Calvas, Estelle Harroche, Antoine Faurie-Grepon. CHU Lille: Luc Lefebvre, Alexandre Kreisler, Alain Destée, David Devos; CHU Rouen: David Maltête; CHU Amiens: Pierre Krystkowiak; CHU Strasbourg: Christine Tranchant, Mathieu Anheim, Clélie Phillipps, Nadine Longato, Thomas Wirth, Ouhaid Lagha-Boukbiza; Genetic committee: Suzanne Lesage, Alexis Brice, Fanny Charbonnier-Beaupel, Yannick Marie, Khadija Tahiri.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fneur.2020.00641/full#supplementary-material
References
1. Weintraub D, Koester J, Potenza MN, Siderowf AD, Stacy M, Voon V, et al. Impulse control disorders in Parkinson disease: a cross-sectional study of 3090 patients. Arch Neurol. (2010) 67:589–95. doi: 10.1001/archneurol.2010.65
2. Dell'Osso B, Altamura AC, Allen A, Marazziti D, Hollander E. Epidemiologic and clinical updates on impulse control disorders: a critical review. Eur Arch Psychiatry Clin Neurosci. (2006) 256:464–75. doi: 10.1007/s00406-006-0668-0
3. Voon V, Sohr M, Lang AE, Potenza MN, Siderowf AD, Whetteckey J, et al. Impulse control disorders in Parkinson disease: a multicenter case–control study. Ann Neurol. (2011) 69:986–96. doi: 10.1002/ana.22356
4. Pirritano D, Plastino M, Bosco D, Gallelli L, Siniscalchi A, De Sarro G. Gambling disorder during dopamine replacement treatment in Parkinson's disease: a comprehensive review. BioMed Res Int. (2014) 2014:728038. doi: 10.1155/2014/728038
5. de la Riva P, Smith K, Xie SX, Weintraub D. Course of psychiatric symptoms and global cognition in early Parkinson disease. Neurology. (2014) 83:1096–103. doi: 10.1212/WNL.0000000000000801
6. Slutske WS, Zhu G, Meier MH, Martin NG. Genetic and environmental influences on disordered gambling in men and women. Arch Gen Psychiatry. (2010) 67:624–30. doi: 10.1001/archgenpsychiatry.2010.51
7. Eisen SA, Lin N, Lyons MJ, Scherrer JF, Griffith K, True WR, et al. Familial influences on gambling behavior: an analysis of 3359 twin pairs. Addict Abingdon Engl. (1998) 93:1375–84. doi: 10.1046/j.1360-0443.1998.93913758.x
8. Kraemmer J, Smith K, Weintraub D, Guillemot V, Nalls MA, Cormier-Dequaire F, et al. Clinical-genetic model predicts incident impulse control disorders in Parkinson's disease. J Neurol Neurosurg Psychiatry. (2016) 87:1106–11. doi: 10.1136/jnnp-2015-312848
9. Comings DE, Gade-Andavolu R, Gonzalez N, Wu S, Muhleman D, Chen C, et al. The additive effect of neurotransmitter genes in pathological gambling. Clin Genet. (2001) 60:107–16. doi: 10.1034/j.1399-0004.2001.600204.x
10. Guo G, Tong Y, Xie CW, Lange LA. Dopamine transporter, gender, and number of sexual partners among young adults. Eur J Hum Genet. (2007) 15:279–87. doi: 10.1038/sj.ejhg.5201763
11. Lee JY, Lee EK, Park SS, Lim JY, Kim HJ, Kim JS, et al. Association of DRD3 and GRIN2B with impulse control and related behaviors in Parkinson's disease. Mov Disord. (2009) 24:1803–10. doi: 10.1002/mds.22678
12. Cilia R, Benfante R, Asselta R, Marabini L, Cereda E, Siri C, et al. Tryptophan hydroxylase type 2 variants modulate severity and outcome of addictive behaviors in Parkinson's disease. Parkinsonism Relat Disord. (2016) 29:96–103. doi: 10.1016/j.parkreldis.2016.05.017
13. Zainal Abidin S, Tan EL, Chan SC, Jaafar A, Lee AX, Abd Hamid MHN, et al. DRD and GRIN2B polymorphisms and their association with the development of impulse control behaviour among Malaysian Parkinson's disease patients. BMC Neurol. (2015) 15:59. doi: 10.1186/s12883-015-0316-2
14. Krishnamoorthy S, Rajan R, Banerjee M, Kumar H, Sarma G, Krishnan S, et al. Dopamine D3 receptor Ser9Gly variant is associated with impulse control disorders in Parkinson's disease patients. Parkinsonism Relat Disord. (2016) 30:13–7. doi: 10.1016/j.parkreldis.2016.06.005
15. Morgante F, Fasano A, Ginevrino M, Petrucci S, Ricciardi L, Bove F, et al. Impulsive-compulsive behaviors in parkin-associated Parkinson disease. Neurology. (2016) 87:1436–41. doi: 10.1212/WNL.0000000000003177
16. Strafella AP. Mesolimbic dopamine and anterior cingulate cortex connectivity changes lead to impulsive behaviour in Parkinson's disease. Brain. (2019) 142:496–8. doi: 10.1093/brain/awz010
17. van Eimeren T, Pellecchia G, Cilia R, Ballanger B, Steeves TDL, Houle S, et al. Drug-induced deactivation of inhibitory networks predicts pathological gambling in PD. Neurology. (2010) 75:1711–6. doi: 10.1212/WNL.0b013e3181fc27fa
18. Emond MJ, Louie T, Emerson J, Zhao W, Mathias RA, Knowles MR, et al. Exome sequencing of extreme phenotypes identifies DCTN4 as a modifier of chronic Pseudomonas aeruginosa infection in cystic fibrosis. Nat Genet. (2012) 44:886–9. doi: 10.1038/ng.2344
19. Shtir C, Aldahmesh MA, Al-Dahmash S, Abboud E, Alkuraya H, Abouammoh MA, et al. Exome-based case-control association study using extreme phenotype design reveals novel candidates with protective effect in diabetic retinopathy. Hum Genet. (2016) 135:193–200. doi: 10.1007/s00439-015-1624-8
20. Lee S, Abecasis GR, Boehnke M, Lin X. Rare-variant association analysis: study designs and statistical tests. Am J Hum Genet. (2014) 95:5–23. doi: 10.1016/j.ajhg.2014.06.009
21. Cormier-Dequaire F, Bekadar S, Anheim M, Lebbah S, Pelissolo A, Krack P, et al. Suggestive association between OPRM1 and impulse control disorders in Parkinson's disease. Mov Disord. (2018) 33:1878–86. doi: 10.1002/mds.27519
22. Hughes AJ, Daniel SE, Kilford L, Lees AJ. Accuracy of clinical diagnosis of idiopathic Parkinson's disease: a clinico-pathological study of 100 cases. J Neurol Neurosurg Psychiatry. (1992) 55:181–4. doi: 10.1136/jnnp.55.3.181
23. Rieu I, Martinez-Martin P, Pereira B, De Chazeron I, Verhagen Metman L, Jahanshahi M, et al. International validation of a behavioral scale in Parkinson's disease without dementia. Mov Disord. (2015) 30:705–13. doi: 10.1002/mds.26223
24. Tomlinson CL, Stowe R, Patel S, Rick C, Gray R, Clarke CE. Systematic review of levodopa dose equivalency reporting in Parkinson's disease. Mov Disord. (2010) 25:2649–3. doi: 10.1002/mds.23429
25. Thaler A, Gonen T, Mirelman A, Helmich RC, Gurevich T, Orr-Urtreger A, et al. LRRK2 Ashkenazi Jewish consortium. altered reward-related neural responses in non-manifesting carriers of the Parkinson disease related LRRK2 mutation. Brain Imaging Behav. (2019) 13:1009–20. doi: 10.1007/s11682-018-9920-2
26. Weintraub D, Hoops S, Shea JA, Lyons KE, Pahwa R, Driver-Dunckley ED, et al. Validation of the questionnaire for impulsive-compulsive disorders in Parkinson's disease. Mov Disord. (2009) 24:1461–7. doi: 10.1002/mds.22571
27. Paila U, Chapman BA, Kirchner R, Quinlan AR. GEMINI: integrative exploration of genetic variation and genome annotations. PLoS Comput Biol. (2013) 9:e1003153. doi: 10.1371/journal.pcbi.1003153
28. Kircher M, Witten DM, Jain P, O'Roak BJ, Cooper GM, Shendure J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. (2014) 46:310–5. doi: 10.1038/ng.2892
29. Amendola LM, Dorschner MO, Robertson PD, Salama JS, Hart R, Shirts BH, et al. Actionable exomic incidental findings in 6503 participants: challenges of variant classification. Genome Res. (2015) 25:305–15. doi: 10.1101/gr.183483.114
30. Fabregat A, Sidiropoulos K, Garapati P, Gillespie M, Hausmann K, Haw R, et al. The Reactome pathway Knowledgebase. Nucleic Acids Res. (2016) 44:D481–7. doi: 10.1093/nar/gkv1351
31. Croft D, Mundo AF, Haw R, Milacic M, Weiser J, Wu G, et al. The Reactome pathway knowledgebase. Nucleic Acids Res. (2014) 42:D472–7. doi: 10.1093/nar/gkt1102
32. Lang M, Leménager T, Streit F, Fauth-Bühler M, Frank J, Juraeva D, et al. Genome-wide association study of pathological gambling. Eur Psychiatry. (2016) 36:38–46. doi: 10.1016/j.eurpsy.2016.04.001
33. Corvol JC, Studler JM, Schonn JS, Girault JA, Hervé D. Galpha(olf) is necessary for coupling D1 and A2a receptors to adenylyl cyclase in the striatum. J Neurochem. (2001) 76:1585–8. doi: 10.1046/j.1471-4159.2001.00201.x
34. Valjent E, Pascoli V, Svenningsson P, Paul S, Enslen H, Corvol JC, et al. Regulation of a protein phosphatase cascade allows convergent dopamine and glutamate signals to activate ERK in the striatum. Proc Natl Acad Sci USA. (2005) 102:491–6. doi: 10.1073/pnas.0408305102
35. Stipanovich A, Valjent E, Matamales M, Nishi A, Ahn JH, Maroteaux M, et al. A phosphatase cascade by which rewarding stimuli control nucleosomal response. Nature. (2008) 453:879–84. doi: 10.1038/nature06994
36. Calabresi P, Picconi B, Tozzi A, Di Filippo M. Dopamine-mediated regulation of corticostriatal synaptic plasticity. Trends Neurosci. (2007) 30:211–9. doi: 10.1016/j.tins.2007.03.001
37. Faccidomo S, Besheer J, Stanford PC, Hodge CW. Increased operant responding for ethanol in male C57BL/6J mice: specific regulation by the ERK1/2, but not JNK, MAP kinase pathway. Psychopharmacology. (2009) 204:135–47. doi: 10.1007/s00213-008-1444-9
38. Valjent E, Corvol JC, Pages C, Besson MJ, Maldonado R, Caboche J. Involvement of the extracellular signal-regulated kinase cascade for cocaine-rewarding properties. J Neurosci. (2000) 20:8701–9. doi: 10.1523/JNEUROSCI.20-23-08701.2000
39. Santini E, Valjent E, Usiello A, Carta M, Borgkvist A, Girault JA, et al. Critical involvement of cAMP/DARPP-32 and extracellular signal-regulated protein kinase signaling in L-DOPA-induced dyskinesia. J Neurosci. (2007) 27:6995–7005. doi: 10.1523/JNEUROSCI.0852-07.2007
40. Zhan L, Liu B, Jose-Lafuente M, Chibalina MV, Grierson A, Maclean A, et al. ALG-2 interacting protein AIP1: a novel link between D1 and D3 signalling. Eur J Neurosci. (2008) 27:1626–33. doi: 10.1111/j.1460-9568.2008.06135.x
41. Lin DTS, Fretier P, Jiang C, Vincent SR. Nitric oxide signaling via cGMP-stimulated phosphodiesterase in striatal neurons. Synap N Y N. (2010) 64:460–6. doi: 10.1002/syn.20750
42. Schumann G, Coin LJ, Lourdusamy A, Charoen P, Berger KH, Stacey D, et al. Genome-wide association and genetic functional studies identify autism susceptibility candidate 2 gene (AUTS2) in the regulation of alcohol consumption. Proc Natl Acad Sci USA. (2011) 108:7119–24. doi: 10.1073/pnas.1106917108
43. Stacey D, Bilbao A, Maroteaux M, Jia T, Easton AC, Longueville S, et al. RASGRF2 regulates alcohol-induced reinforcement by influencing mesolimbic dopamine neuron activity and dopamine release. Proc Natl Acad Sci USA. (2012) 109:21128–33. doi: 10.1073/pnas.1211844110
44. Neasta J, Ben Hamida S, Yowell QV, Carnicella S, Ron D. AKT signaling pathway in the nucleus accumbens mediates excessive alcohol drinking behaviors. Biol Psychiatry. (2011) 70:575–82. doi: 10.1016/j.biopsych.2011.03.019
Keywords: Parkinson's disease (PD), dopamine agonists (DA), impulse control disorders (ICD), pharmacogenetics, exome sequencing
Citation: Prud'hon S, Bekadar S, Rastetter A, Guégan J, Cormier-Dequaire F, Lacomblez L, Mangone G, You H, Daniau M, Marie Y, Bertrand H, Lesage S, Tezenas Du Montcel S, Anheim M, Brice A, Danjou F and Corvol J (2020) Exome Sequencing Reveals Signal Transduction Genes Involved in Impulse Control Disorders in Parkinson's Disease. Front. Neurol. 11:641. doi: 10.3389/fneur.2020.00641
Received: 17 January 2020; Accepted: 29 May 2020;
Published: 21 July 2020.
Edited by:
Chin-Hsien Lin, National Taiwan University Hospital, TaiwanReviewed by:
Luca Marsili, University of Cincinnati, United StatesKuo-Hsuan Chang, Linkou Chang Gung Memorial Hospital, Taiwan
Copyright © 2020 Prud'hon, Bekadar, Rastetter, Guégan, Cormier-Dequaire, Lacomblez, Mangone, You, Daniau, Marie, Bertrand, Lesage, Tezenas Du Montcel, Anheim, Brice, Danjou and Corvol. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Christophe Corvol, jean-christophe.corvol@aphp.fr