 Huhu Liu
Huhu Liu Yulan Song
Yulan Song Xiao Fan
Xiao Fan Yun Tian
Yun Tian- College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha, China
The microbial fermentation process has been used as an alternative pathway to the production of value-added natural products. Of the microorganisms, Yarrowia lipolytica, as an oleaginous platform, is able to produce fatty acid-derived biofuels and biochemicals. Nowadays, there are growing progresses on the production of value-added fatty acid-based bioproducts in Y. lipolytica. However, there are fewer reviews performing the metabolic engineering strategies and summarizing the current production of fatty acid-based bioproducts in Y. lipolytica. To this end, we briefly provide the fatty acid metabolism, including fatty acid biosynthesis, transportation, and degradation. Then, we introduce the various metabolic engineering strategies for increasing bioproduct accumulation in Y. lipolytica. Further, the advanced progress in the production of fatty acid-based bioproducts by Y. lipolytica, including nutraceuticals, biofuels, and biochemicals, is summarized. This review will provide attractive thoughts for researchers working in the field of Y. lipolytica.
Introduction
With the growing crisis of oil energy, microbial production of biochemicals, as one potential alternative route, has received increasing attention (Levering et al., 2015; Ji and Huang, 2019; Ji and Ledesma-Amaro, 2020). Among these microorganisms, the oleaginous yeasts, such as Rhodosporidium toruloides, Lipomyces starkeyi, and Yarrowia lipolytica, are able to produce oleochemicals (Probst et al., 2016; McNeil and Stuart, 2018; Park et al., 2018b; Miller and Alper, 2019). Y. lipolytica, as Food and Drug Administration (FDA)-regarded Generally Recognized as Safe (GRAS) yeast with lipids over 20% of its biomass, performs many attractive characteristics and applications, including having mature genetic tools, secreting functional enzymes, and producing organic acids, lipids, and non-native chemicals (Xie, 2017; Darvishi et al., 2018; Larroude et al., 2018; Madzak, 2018; Ma et al., 2019). Currently, many researchers focus on the biotechnological application of Y. lipolytica (Xie et al., 2015; Markham et al., 2018; Robles-Rodriguez et al., 2018; Li et al., 2019). In particular, the different metabolic engineering strategies are applied in the lipid production for Y. lipolytica (Abdel-Mawgoud et al., 2018; Wang J. et al., 2020). In fact, Y. lipolytica is able to produce fatty acids in the form of lipids, either grown on hydrophilic or hydrophobic materials (Spagnuolo et al., 2018; Ma et al., 2020). Generally, these fatty acid-based bioproducts from Y. lipolytica are divided into three different types, based on the chain length, the terminal reductive state, and the modifications to the main chain of target product (Yan and Pfleger, 2020). With the development of metabolic engineering and synthetic biology, there are growing progresses on the production of value-added fatty acid-based bioproducts in Y. lipolytica. In the past 5 years, researchers have reviewed the production of fatty acid-derived products by Y. lipolytica, including fatty alkanes, fatty alcohols, and polyunsaturated fatty acids (PUFAs) (Ledesma-Amaro and Nicaud, 2016b; Ma et al., 2020). However, there is less review performing the metabolic engineering strategies for improving the production of fatty acid-based products and summarizing the current biosynthesis of fatty acid-based bioproducts in Y. lipolytica.
Herein, in this review, we describe a brief overview of the biochemistry metabolism of fatty acid in Y. lipolytica. Then, we focus on introducing the various metabolic strategies for increasing bioproduct accumulation, including constructing and engineering metabolic pathways, optimizing fermentation conditions, and engineering compartmentalization system. Moreover, we summarize the recent progress in the production of fatty acid-based bioproducts in Y. lipolytica, including nutraceuticals, biofuels, and biochemicals (Table 1). This article will provide attractive thoughts for researchers working in the field of Y. lipolytica.
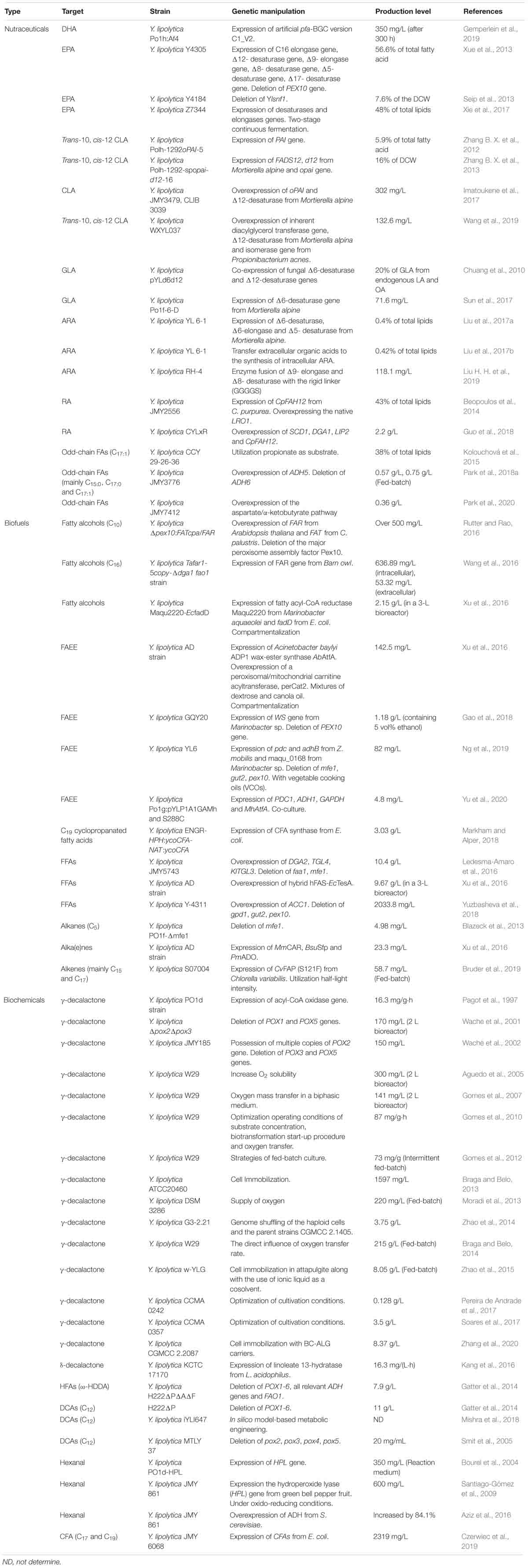
Table 1. Summary of the production of fatty acid-based bioproducts from the Y. lipolytica platform.
Biochemistry of Fatty Acid Metabolism
Currently, some articles have summarized the fatty acid metabolism of Y. lipolytica (Fickers et al., 2005; Abghari and Chen, 2014; Ledesma-Amaro and Nicaud, 2016a; Lazar et al., 2018). Previously, we reviewed in detail the characteristics of Y. lipolytica grown on various carbon substrates (Liu et al., 2015). Herein, the metabolism of fatty acid for producing its derived chemicals in Y. lipolytica is shown in Figure 1.
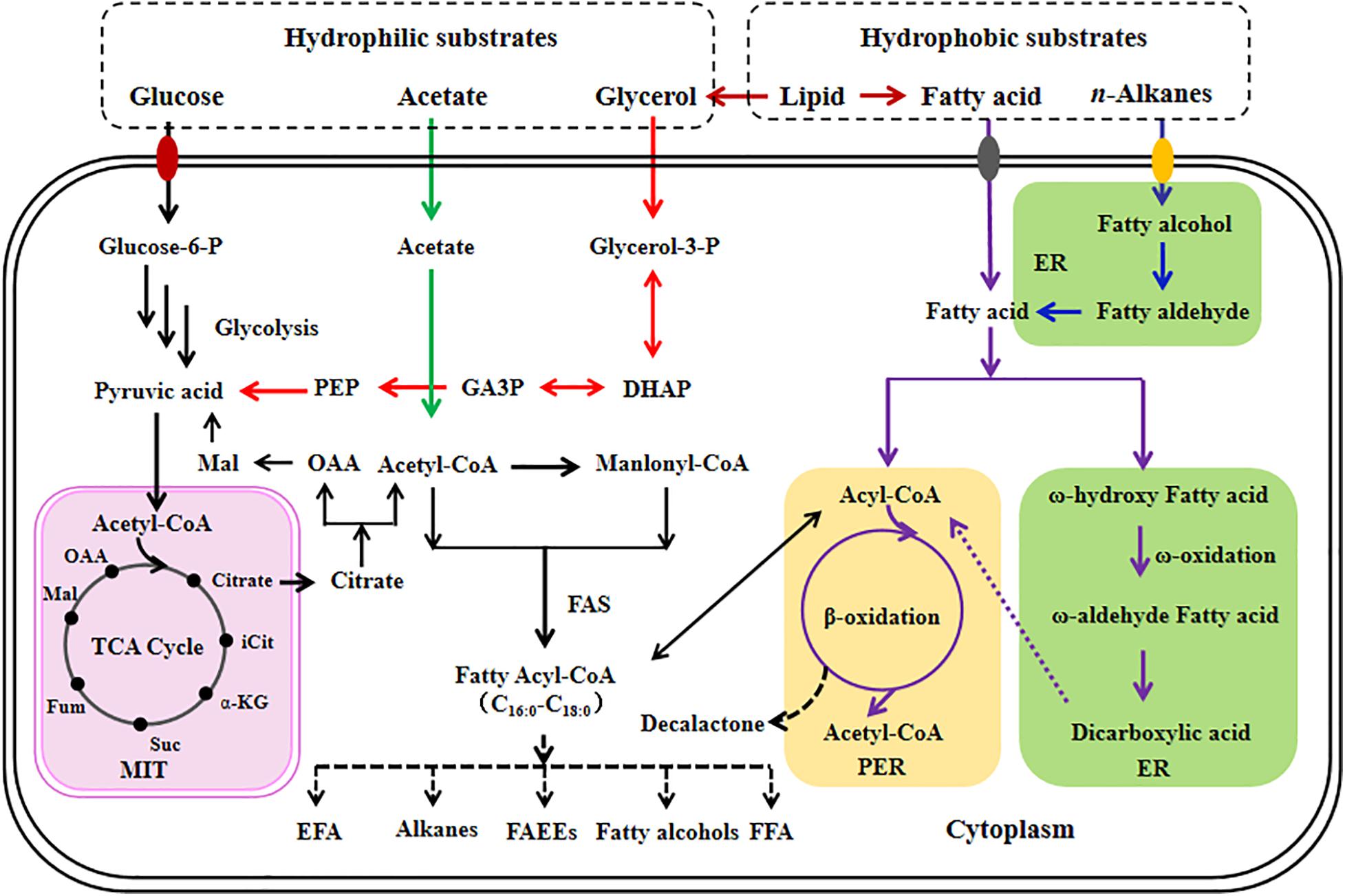
Figure 1. Overview of fatty acid metabolism for the production of its based chemicals in Y. lipolytica. Different colored arrows are used to represent different metabolic pathways; black, de novo fatty acid metabolic pathway; green, acetate metabolic pathway; red, glycerol metabolic pathway; dark red, heterologous lipid metabolic pathway; purple, ex novo fatty acid metabolic pathway; blue, heterologous alkane metabolic pathway. Pathway localization with respect to specific subcellular organelles are also depicted. ER, endoplasmic reticulum; PER, peroxisome; MIT, mitochondria; TCA cycle, tricarboxylic acid cycle; DHAP, dihydroxyacetone phosphate; GA3P, glycerol-3-phosphate; iCit, isocitrate; α-KG, α-ketoglutarate; Suc, succinate; Fum, fumarate; Mal, malate; OAA, oxaloacetate; FAS, fatty acid synthase; EFA, essential fatty acid; FFA, free fatty acid; FAEEs, fatty acid ethyl esters.
Fatty Acid Biosynthesis
With the development of metabolic engineering, it enables Y. lipolytica to utilize a wide range of carbon sources (Liu et al., 2015; Ledesma-Amaro and Nicaud, 2016b). Using hydrophilic substrates (such as glucose and glycerol) as carbon source, fatty acid is synthesized by de novo pathway in Y. lipolytica. With glucose as sole carbon source, it is converted into pyruvate via the glycolytic pathway in the cytosol. Then, pyruvate is transported to mitochondria and transformed into acetyl-CoA. Acetyl-CoA, a key precursor involved in fatty acid biosynthesis, can be produced by different metabolic routes, including citrate degradation catalyzed by ATP citrate lyase (ACL), fatty acid degradation from β-oxidation pathway, acetate transformation by acetyl-CoA synthetase (ACS, YALI0F05962p), and pyruvate transformation by pyruvate dehydrogenase complex. Under nitrogen-limited conditions, citrate is secreted into cytosol from mitochondria in Y. lipolytica and acetyl-CoA is produced by ACL catalysis. In Y. lipolytica, ACL is encoded by ACL1 (YALI0E34793p) and ACL2 (YALI0D24431p). Further, acetyl-CoA is transformed into malonyl-CoA by acetyl-CoA carboxylase (ACC, YALI0C11407p). Generally, acetyl-CoA and malonyl-CoA are used as substrates for fatty acid biosynthesis by fatty acid synthetases (FAS, YALI0B15059p, and YALI0B19382p) in Y. lipolytica. Naturally, Y. lipolytica can only produce C16 and C18 fatty acids (Beopoulos et al., 2009). Notably, the inherent long-chain PUFAs, including oleic acid (OA, C18:1) or linoleic acid (LA, C18:2), are synthesized by desaturase located in endoplasmic reticulum (ER).
Using hydrophobic materials (such as fats) as substrate, fatty acids are synthesized by ex novo pathway in Y. lipolytica. Generally, the extracellular fatty acids from the metabolism of hydrophobic materials are directly transported to cytosol in Y. lipolytica. Then, fatty acids are converted into derived chemicals by the corresponding oxidation process. Additionally, using alkane from oil refinery as carbon source, fatty acids are synthesized by the enzyme catalytic system located in ER, including cytochrome P450 reductase (EC 1.6.2.4), fatty alcohol oxidase (EC 1.1.3.20), and fatty aldehyde dehydrogenase (EC 1.2.1.3).
NADPH is an important reducing power involved in fatty acid biosynthesis in Y. lipolytica. Generally, there are two identified routes for providing NADPH pool in Y. lipolytica (Qiao et al., 2017). One route is from decarboxylation reaction catalyzed by malic enzyme (EC 1.1.1.40) that occurred in cytosol; the other metabolic route is from the pentose phosphate pathway in Y. lipolytica. Previously, it was reported that overexpression of malic enzyme has little impact on lipid accumulation in Y. lipolytica (Beopoulos et al., 2011; Zhang H. et al., 2013). Wasylenko et al. (2015) reported that the oxidative pentose phosphate pathway, harboring glucose 6-phosphate dehydrogenase (EC 1.1.1.49) and 6-phosphogluconolactonase (EC 3.1.1.31), is the primary source of lipogenic NADPH in Y. lipolytica.
Fatty Acid Transportation
To date, the mechanism of fatty acid transportation is unclear in Y. lipolytica. Generally, shorter carbon-chain fatty acids, such as C8:0 and C10:0, are toxic for Y. lipolytica. Using primrose oil containing C18 fatty acids as substrate, Y. lipolytica performs a higher assimilation rate for unsaturated fatty acids (C18:3, C18:2, and C18:1) than that for saturated fatty acid (C18:0) (Aggelis et al., 1997). In this research, it was deduced that the fatty acids with different saturated levels are assimilated and transported via a selective uptake mechanism in Y. lipolytica. Recently, Dulermo et al. (2015) proposed a model of fatty acid transportation with chain length preferences in Y. lipolytica. According to this model, the extracellular fatty acids are transported into Y. lipolytica via unidentified transporters. Then, the internal fatty acids are activated to acyl-CoA by YlFaa1p (YALI0D17864p) or transported into peroxisome by unknown transporters. Notably, the activated fatty acids can be sorted in the form of triacylglycerols or enter peroxisome via transporters YlPxa1p (YALI0A06655p) and YlPxa2p (YALI0D04246p). Importantly, fatty acids from lipid remobilization can enter the peroxisome via transporter YlFat1p (YALI0E16016p).
In particular, the intracellular medium-chain fatty acids (C12–C14) are converted into fatty acyl-CoAs by fatty acyl-CoA synthetase II in the peroxisome for further degradation, whereas long-chain fatty acids (C16–C18) are converted into fatty acyl-CoA by fatty acyl-CoA synthetase I in the cytosol (Dulermo et al., 2015). Then, long-chain fatty acyl-CoA is either transported into peroxisome from cytosol or used as substrate for triacylglyceride biosynthesis in Y. lipolytica.
Fatty Acid Degradation
Generally, fatty acids, either from intracellular triacylglyceride hydrolysis or from extracellular fatty acid transportation, can be transformed into fatty acid-based chemicals by oxidation in Y. lipolytica. Notably, the intracellular fatty acids are mainly degraded by peroxisomal β-oxidation or ω-oxidation pathway. In fact, the intracellular fatty acids from lipid remobilization are mainly converted into acetyl-CoA, via peroxisomal β-oxidation pathway. In particular, each cycle of β-oxidation consists of a four-step enzyme catalyzed reaction in Y. lipolytica. The first step is catalyzed by acyl-CoA oxidases (EC 1.3.3.6), the second step and third steps are catalyzed by multifunctional enzyme, and the last step is catalyzed by 3-ketoacyl-CoA thiolase (EC 2.3.1.16). In addition, the intracellular fatty acids can be degraded into derived chemicals by ω-oxidation pathway that occurred in ER. The fatty acids are firstly converted into ω-hydroxyl-fatty acids by cytochrome P450-containing fatty acid ω-hydroxylase. Then, ω-hydroxyl-fatty acids are converted into ω-aldo-fatty acids by fatty alcohol dehydrogenase or fatty alcohol oxidase, and ω-aldo-fatty acids are converted into long-chain diacids by fatty aldehyde dehydrogenase. In particular, the β-oxidation pathway can be engineered to synthesize β-hydroxy fatty acid (HFA) and lactones, whereas the ω-oxidation pathway can be engineered to produce ω-HFA and α, ω-dicarboxylic acids (DCAs) in Y. lipolytica.
Engineering Strategies to Increase Oleochemical Production
Nowadays, different metabolic strategies have been used to de novo produce the novel fatty acid-based bioproducts and accumulate the production of these derived biochemicals in Y. lipolytica (Table 2).
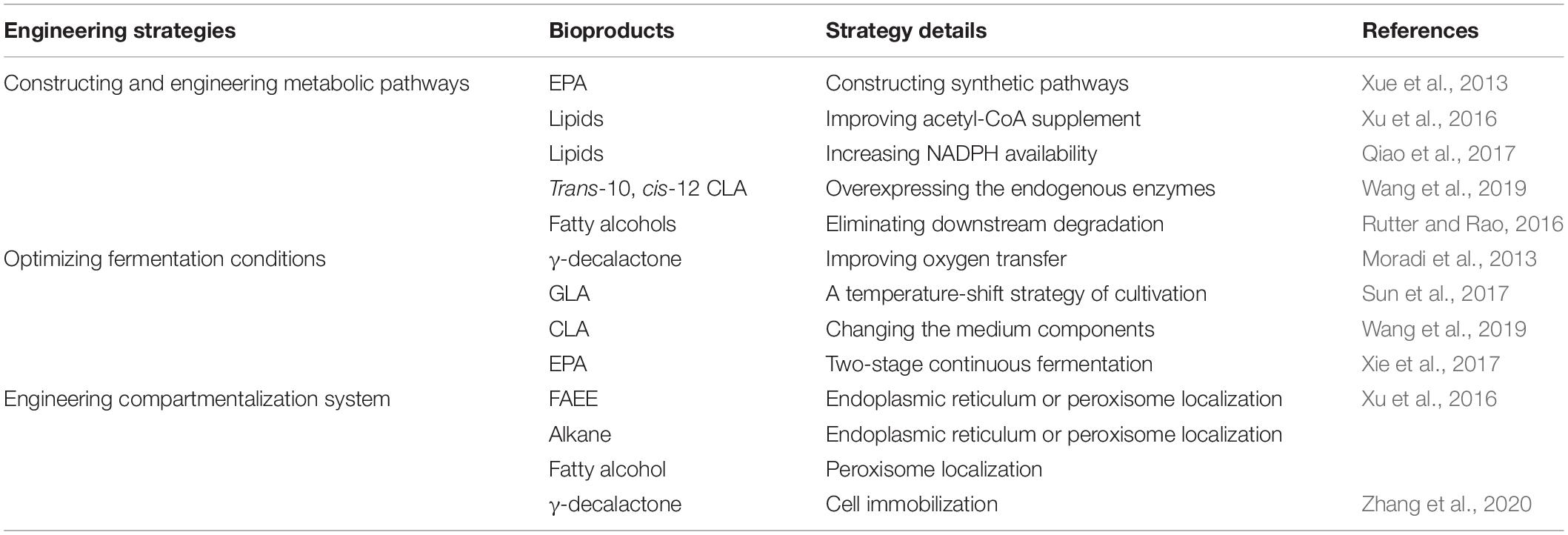
Table 2. Engineering strategies to improve fatty acid-based bioproducts accumulation in Y. lipolytica.
Constructing and Engineering Metabolic Pathways
Researchers have focused on constructing and optimizing metabolic pathways to achieve efficient fatty acid and its derivatives biosynthesis in Y. lipolytica, using various metabolic engineering strategies, including constructing heterologous synthetic pathways, overexpressing endogenous enzymes. Naturally, Y. lipolytica can produce linoleic acid as the precursor of ω-3/6 fatty acids (Liu et al., 2017a). Generally, the novel linoleic acid-derived nutraceuticals, such as arachidonic acid (ARA, C20:4) and eicosapentaenoic acid (EPA, C20:5), can be de novo synthesized via constructing the synthetic pathway in Y. lipolytica. For example, to de novo produce EPA in Y. lipolytica, the selected and optimized multiple copies of different chimeric genes from different microorganisms were integrated into yeast genome (Δ9-elongase, Δ8-desaturase, and Δ5-desaturase from E. gracilis, C16/18-elongase from M. alpina, Δ12-desaturase gene from F. moniliforme, Δ17-desaturase from P. aphanidermatum, and CPT), which led to the first engineered commercial strain Y4305 under strong promoters, containing 30 copies of nine different genes, which can produce EPA at 56.6% of the total fatty acids (TFA), without γ-linolenic acid (GLA, C18:3) accumulation (Xue et al., 2013).
Through overexpressing and eliminating the endogenous enzymes involved in the lipid degradation, the accumulation of fatty acid and its derivatives has been greatly enhanced in Y. lipolytica (Dulermo and Nicaud, 2011). Generally, the availability of precursors, including acetyl-CoA and NADPH, limits the lipid biosynthesis. Previously, by harnessing the carnitine shuttle mechanism, the lipid titer was enhanced 1.75-fold via increasing acetyl-CoA supplement (Xu et al., 2016). Qiao et al. (2017) performed a specific strategy of converting NADH to NADPH in 13 engineered strains of Y. lipolytica for improving lipid synthesis. Recently, Wang et al. (2019) showed that the increased conjugated linoleic acid (CLA, C18:2) accumulation is reached by overexpressing the endogenous diacylglycerol transferase gene. Additionally, in order to block the lipid degradation in Y. lipolytica, Rutter and Rao (2016) showed that the peroxisome assembly factor Pex10 is the major enzyme involved in the peroxisomal β-oxidation or ω-oxidation pathway.
Optimizing Fermentation Conditions
The optimization of fermentation process, based on the microbial physiology, plays a key role in achieving the high titer, yield, and productivity of value-added products. Naturally, pH, temperature, and medium components are the common optimized approaches during the fermentation process of Y. lipolytica. Previously, the temperature-shift strategy of cultivation was successfully exhibited to increase GLA accumulation in Y. lipolytica (Sun et al., 2017). Recently, the production of CLA was increased by changing carbon and nitrogen source, carbon-t- nitrogen mass ratio, and CaCl2 concentrations (Wang et al., 2019). In addition, the fed-batch fermentation approach has been used to increase the production of drop-in biochemicals (Park et al., 2018a; Bruder et al., 2019). Compared with the continuous fermentation processes, the batch and fed-batch processes perform lower volumetric productivities (Li et al., 2011). In fact, the productivities utilizing continuous fermentation processes were improved, typically at the cost of product concentration, conversion yield, or both (Ethier et al., 2011). Previously, the novel two-stage continuous process for EPA accumulation in Y. lipolytica was developed (Xie et al., 2017). In this research, compared with the single-stage continuous and fed-batch fermentation, the novel continuous process, equipped with a small growth tank (Stage 1) and a large production tank (Stage 2), successfully improved the volumetric lipid productivities by 80%.
Generally, Y. lipolytica requires a high oxygen supply in the large-scale bioprocess. Previously, researchers have showed that the heterologous expression of gene encoding the bacterial hemoglobin from Vitreoscilla stercoraria (VHb) can improve the oxygen utilization efficiency and further increase the productivity (Suen et al., 2014; Zhang et al., 2017). Recently, Mirończuk et al. (2019) performed that the improved erythritol synthesis is obtained in Y. lipolytica, by overexpressing the codon-optimized bacterial hemoglobin (VHb). Through improving oxygen transfer rate using higher agitation rates or pure oxygen for aeration, the production of γ-decalactone was successfully enhanced (Moradi et al., 2013).
Engineering Compartmentalization System
Naturally, each subcellular compartment in Y. lipolytica provides a unique microenvironment, including enzyme, precursor, and cofactor composition. Due to the distinct organelle characteristics, the separation of organelles in the cytosol performs the potential to eliminate metabolic crosstalk and enhance compartmentalized pathway efficiency (Hammer and Avalos, 2017). Previously, Xu et al. (2016) reported that the titer of drop-in product performs a 10–15-fold improvement, by targeting the fatty acid ethyl ester (FAEE) pathway to either ER or peroxisome of Y. lipolytica. Compared to free cell systems, the immobilized cells could tolerate unsuitable conditions (Li et al., 2009; Macario et al., 2009). For example, using cell immobilization systems with bacterial cellulose-alginate (BC-ALG) carriers, γ-decalactone production was successfully reached with 8.37 g/L in the repeated experiments in Y. lipolytica, an approximately 3.7-fold improvement over with an ALG carrier alone (Zhang et al., 2020).
Modular co-culture metabolic engineering combines the strains carrying each pathway module in the engineered strains to form a synthetic complex, which can accommodate different modules expressing functional genes in different hosts to produce drop-in bioproducts (Jawed et al., 2019). Recently, by coculturing and engineered Y. lipolytica and S. cerevisiae strain, a synthetic microbial consortium was constructed to increase the titer of FAEE. In this research, the titer of FAEE biodiesel at 4.8 mg/L was reached by the synthetic microbial consortium under the optimum coculture conditions (Yu et al., 2020).
Production of Fatty Acid-Based Bioproducts
Nutraceuticals
Due to the potential applications of microbial lipids in the field of food supplements, the microbial production of PUFAs is becoming an industrial reality (Bellou et al., 2016). Of these oleaginous yeasts, Y. lipolytica can synthesize OA and LA.
Omega-3 PUFAs with special function, particularly α-linolenic acid (ALA, C18:3), EPA, and docosahexaenoic acid (DHA, C22:6), are gaining importance. Previously, using inherent LA as carbon substrate, Xue et al. (2013) constructed an engineered Y. lipolytica strain Y4305 capable of de novo producing EPA at 56.6% of TFA, by the combined metabolic engineering strategies. With Y. lipolytica as a host, the highest titer of ALA at 1.4 g/L was produced in the engineered strain containing a bifunctional Δ12–Δ15 desaturase from Rhodosporidium kratochvilovae, under the optimized fermentation conditions (Cordova and Alper, 2018). Recently, an artificial PUFA biosynthetic gene clusters, encoding DPA/DHA-type PUFA synthases, was expressed in Y. lipolytica. In this research, under the optimized fermentation process, the DHA level over 350 mg/L was reached (Gemperlein et al., 2019).
Omega-6 PUFAs, including conjugated CLA, GLA, and ARA, are a major family of PUFAs with diverse bioactivities (Xu and Qian, 2014). In 2017, the combined elimination of β-oxidation pathway and overexpression of Δ12-desaturase was conducted in Y. lipolytica, which leads to CLA production at 302 mg/L (Imatoukene et al., 2017). Recently, Wang et al. (2019) showed that the maximum content of trans-10, cis-12 CLA at 132.6 mg/L is reached by the engineered Y. lipolytica under the optimized fermentation conditions, by the overexpression of inherent diacylglycerol transferase from Y. lipolytica, Δ12 desaturase from Mortierella alpina, and Propionibacterium acnes isomerase. With LA as substrate, the GLA biosynthetic pathway was constructed in Y. lipolytica harboring Δ6-desaturase from M. alpina. Under the optimized fermentation process, the titer of GLA at 71.6 mg/L was achieved (Sun et al., 2017).
Arachidonic acid (ARA, C20:4) is also an essential ω-6 PUFA with special functions. Previously, we developed the in vivo one-step pathway assembly and integration method enabling Y. lipolytica to produce ARA (Liu et al., 2017a). Additionally, we showed that the ARA biosynthetic pathway is able to redirect the carbon flux toward intracellular fatty acid accumulation at the expense of extracellular organic acid secretion in the engineered Y. lipolytica strain (Liu et al., 2017b). Recently, using Δ9 elongase pathway engineering and fusion enzyme strategy, the ARA titer at 118.1 mg/L was achieved in the engineered Y. lipolytica (Liu H. H. et al., 2019).
Ricinoleic acid (RA, C18:1) and its derivatives perform oleochemical applications, due to the special characteristics. Meesapyodsuk and Qiu (2008) first identified an oleic acid-like hydroxylase (CpFAH12) from Claviceps purpurea. Previously, with LA as substrate, an engineered Schizosaccharomyces pombe strain capable of producing RA, harboring heterologous CpFAH12 from C. purpurea, was constructed (Holic et al., 2012). Using Y. lipolytica as a host, Beopoulos et al. (2014) reported that RA accumulation at 42% of total lipids is achieved, by overexpressing C. purpurea Δ12-hydroxylase and native Y. lipolytica Lro1p acyltransferase. Recently, by the combined overexpression of SCD1 gene encoding stearoyl-CoA desaturase, DGA1 gene encoding acyl-CoA:diacylglycerol acyltransferase, LIP2 gene encoding lipase, and CpFAH12 gene encoding hydroxylase, the production level of RA at 2.2 g/L was obtained by the engineered Y. lipolytica using cellulose as substrate (Gao et al., 2018).
Odd-chain fatty acids with special biochemical and biological activities are receiving growing attention on potential applications (Řezanka and Sigler, 2009). Previously, Kolouchová et al. (2015) performed that Y. lipolytica is capable of producing heptadecenoic acid (C17:1) using propionate as substrate. Recently, the deletion of the PHD1 gene and optimization of the fermentation process were applied to produce odd-chain fatty acids (mainly C15:0, C17:0, and C17:1) by Y. lipolytica grown on propionate (Park et al., 2018a). Additionally, Park et al. (2020) constructed an engineered Y. lipolytica capable of de novo producing odd-chain fatty acids, using glucose as sole substrate without any propionate supplementation.
Biofuels
The microbial production of fatty alcohols is becoming an alternative method to meet the increasing demand. Presently, various microorganisms, such as Escherichia coli and Saccharomyces cerevisiae, have been engineered for fatty alcohol production (Zhang et al., 2011; Zhou et al., 2016). Using Y. lipolytica as a host, Wang et al. (2016) constructed a novel fatty alcohol-producing workhorse, harboring Tafar1 gene coding fatty acyl-CoA reductase. Under the optimized tri-module condition, the intracellular hexadecanol at 636.89 mg/L and extracellular hexadecanol at 53.32 mg/L was produced, respectively. Meanwhile, through the overexpression of fatty acyl-ACP-thioesterases and fatty acyl-CoA reductase, and deletion of the major peroxisome assembly factor Pex10, the medium-chain alcohol, especially 1-decanol over 500 mg/L, was produced in the engineered Y. lipolytica (Rutter and Rao, 2016).
Researchers have performed that FAEEs or fatty acid methyl esters (FAMEs) can be produced via the microbial fermentation, using E. coli and S. cerevisiae (Steen et al., 2010; Nawabi et al., 2011; Yu et al., 2012). Fortunately, Xu et al. (2016) reported that the highest titer of FAEEs at 142.5 mg/L is produced in the engineered Y. lipolytica, using the compartmentalized metabolic engineering. Recently, an engineered Y. lipolytica strain, harboring the heterogenous pyruvate decarboxylase (pdc), alcohol dehydrogenase II (adhB) from Zymomonas mobilis, and wax ester synthases from Marinobacter sp., was constructed for producing FAEE. In this research, the titer of FAEE up to 82 mg/L was achieved by the supplementation of vegetable cooking oil (Ng et al., 2019). Meanwhile, Yu et al. (2020) developed the synthetic co-culture system comprising the engineered S. cerevisiae and Y. lipolytica strain, which was able to produce FAEE at 4.8 mg/L. To overcome the limitation of oxidative stability in the traditional FAMEs, Markham and Alper (2018) first performed the production of C19 cyclopropanated fatty acids in the engineered Y. lipolytica strain, harboring the heterologous cyclopropane fatty acid synthase from E. coli. In this research, the titer of C19 cyclopropanated fatty acids over 3.0 mg/L was produced under the bioreactor fermentation.
Free fatty acids (FFAs) are special oleochemicals with wide applications in the field of agricultural chemicals, soaps, and surfactants. Previously, Zhou et al. (2016) engineered S. cerevisiae capable of producing FFAs. Using Y. lipolytica as a workhorse, FFAs up to 9.67 g/L were produced by the engineered strain under the bioreactor scale with pH control (Xu et al., 2016). With the mixture of glucose and glycerol as carbon source, Yuzbasheva et al. (2018) showed that the engineered Y. lipolytica Y-4311 strain can produce FFAs (2033.8 mg/L) by the addition of dodecane.
Alka(e)nes are the major components of gasoline, diesel, and jet fuel. Presently, many studies have explored that the microbial production of alkanes is a conceivable method (Choi and Lee, 2013; Zhou et al., 2016). Using Y. lipolytica as a host expressing soybean lipoxygenase enzyme, Blazeck et al. (2013) first developed a microbial platform capable of producing pentane. In particular, in this research, using LA as substrate, the high titer of pentane at 4.98 g/L was produced. Recently, Bruder et al. (2019) revealed that the engineered Y. lipolytica is able to produce odd-numbered alkanes and alkenes (mainly C15 and C17), by the expression of light-driven oxidase. Interestingly, using the lighting bioreactors, the titer of alkenes at 58.7 mg/L was first reached in this research.
Biochemicals
γ-decalactone, a well-known aroma compound, is mainly synthesized via β-oxidation. Previously, we have summarized in detail the γ-decalactone production by Y. lipolytica (Liu et al., 2015). Recently, using the immobilized culture technology, the maximum production of γ-decalactone reached 8.37 g/L by Y. lipolytica strain on bacterial cellulose-alginate carriers (Zhang et al., 2020). Additionally, using a one-pot biotransformation process containing whole Y. lipolytica cells, the highest production of δ-decalactone at 58.7 mg/L was first performed (Kang et al., 2016).
HFAs, as valuable building blocks, can be synthesized by the biotransformation of fatty acids via the terminal carbon oxygenation (Seo et al., 2015). To date, the microbial production of ω-HFAs by the engineered E. coli has received specific progress (Kim and Park, 2019). Using Y. lipolytica as a promising workhorse, an engineered strain capable of synthesizing ω-hydroxy dodecanoic acid was constructed, through the deletion of acyl-CoA oxidase-coding genes (POX 1–6), fatty alcohol oxidase gene (FAO1), and alcohol dehydrogenase genes (ADH 1–8) (Gatter et al., 2014). Recently, Rigouin et al. (2019) showed that the engineered Y. lipolytica is able to produce polyhydroxyalkanoates composed of 3-HFAs, using methyl myristate as precursor.
DCAs are also important intermediates in the industrial field. At present, the microbial production of DCAs, as an alternative method, are gaining interests (Huf et al., 2011; Ledesma-Amaro and Nicaud, 2016b; Werner and Zibek, 2017). Y. lipolytica can produce DCAs via alkane degradation (Nicaud et al., 2006). Previously, researchers have shown that the engineered Y. lipolytica can produce dioic acids (Smit et al., 2005; Nicaud et al., 2006). In particular, Gatter et al. (2014) showed that the overexpression of FAO1 leads to an improved production of dodecane dioic acid at 11 g/L. Recently, using the in silico model-based metabolic engineering strategies, the metabolic flux toward DCAs production was obviously increased in Y. lipolytica (Mishra et al., 2018).
Hexanal, one of C-6 aldehydes with green odor, can be synthesized via the degradation from LA using lipoxygenase and hydroperoxide lyase. Previously, using Y. lipolytica as a host, Bourel et al. (2004) showed that hexanal is produced by expressing of fatty acid hydroperoxide lyase. Further, Santiago-Gómez et al. (2009) reported the effect of oxido-reduction environment on hexanal production. Interestingly, in this research, under the optimized conditions, the highest titer of hexanal at 600 mg/L was produced by the engineered Y. lipolytica.
In addition, cyclopropane fatty acids (CFAs), as good unusual fatty acid candidates, were produced by the engineered Y. lipolytica (Czerwiec et al., 2019). In this research, by expressing genes from various organisms and optimizing the expression level of CFAs synthase and fed-batch fermentation, it was shown that CFAs at 2319 mg/L (mainly C17:0 and C19:0 cyclopropanated form) are finally synthesized in the strain JMY 6068. Compared with E. coli and S. cerevisiae, the fatty acid derivatives produced by Y. lipolytica are more abundant (Table 3).
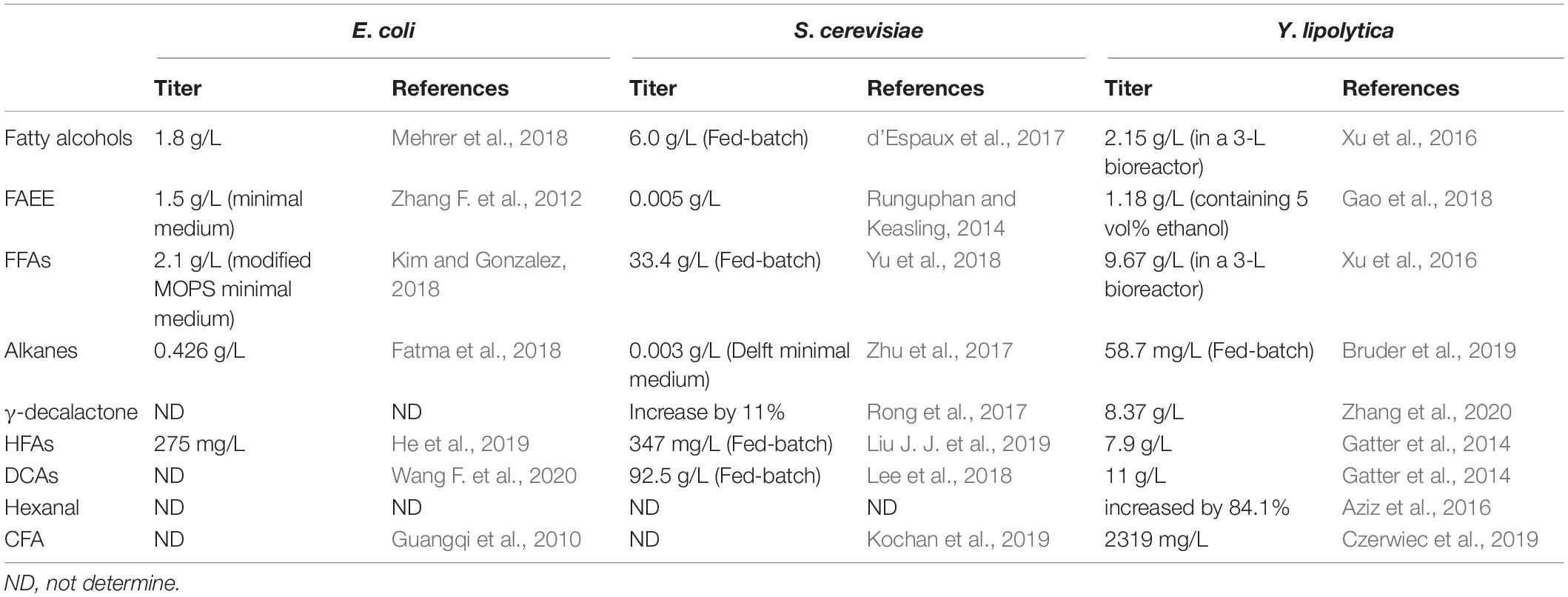
Table 3. Comparison of the productivity of fatty acid-derived biofuels between E. coli, S. cerevisiae, and Y. lipolytica.
Conclusion and Future Perspectives
Y. lipolytica is a promising workhorse gaining great attention. Currently, the advance of metabolic engineering and synthetic biology enables Y. lipolytica to produce various value-added chemicals with different substrates and metabolic engineering strategies, including the design and construction of synthetic pathways, regulation of endogenous genes, and optimization of the fermentation process. However, several challenges remain in limiting the wide applications of Y. lipolytica.
When developing and optimizing Y. lipolytica for improving the production of value-added chemicals, the whole bioprocess, including the upstream of strain development and bioproducts production, the midstream of scale-up fermentation, and the downstream of recovery and purification, is needed to be considered first. Ko et al. (2020) showed that systems metabolic engineering, integrating systems biology, synthetic biology, and evolutionary engineering can enable microbial strains to efficiently produce chemicals. Therefore, systems metabolic engineering can be further applied to better manipulate the engineered Y. lipolytica to synthesize the desired bioproducts. Meanwhile, to optimize cell metabolism, such as reducing the negative effects of intermediate accumulation and metabolic perturbations, the dynamic metabolic engineering capable of tuning the cell growth and bioproducts formation is becoming a promising approach to better engineer the host strain (Xu, 2018). Moreover, due to the limits of dimorphic nature, cellular engineering and bioprocess engineering can be used to improve the yield of products at the industrial scale (Soong et al., 2019). Additionally, to reduce the cost of bioprocess, other low-value carbon sources, especially single-carbon substrates, will be utilized and converted to valuable fatty acid-based bioproducts by metabolic engineering Y. lipolytica. Conclusively, the application of Y. lipolytica for fatty acid-based chemicals production shows a great promise for researchers working in this field.
Author Contributions
HL conceived the outline and revised the manuscript. YT finalized the topic of this review, and all authors wrote the manuscript. All authors read and approved the final manuscript for publication.
Funding
This work was supported by the National Key R&D Program of China (No. 2019YFC1604903), the National Natural Science Foundation of China (No. 21808052), the China Postdoctoral Science Foundation (No. 2019TQ0088), the Scientific Research Foundation of Hunan Provincial Education Department (Nos. 18B090 and 18K039), and the Double First-Class Construction Project of Hunan Agricultural University (No. SYL201802002).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abdel-Mawgoud, A. M., Markham, K. A., Palmer, C. M., Liu, N., Stephanopoulos, G., and Alper, H. S. (2018). Metabolic engineering in the host Yarrowia lipolytica. Metab. Eng. 50, 192–208. doi: 10.1016/j.ymben.2018.07.016
Abghari, A., and Chen, S. L. (2014). Yarrowia lipolytica as an oleaginous cell factory platform for production of fatty acid-based biofuel and bioproducts. Front. Energy Res. 2:21. doi: 10.3389/fenrg.2014.00021
Aggelis, G., Papadiotis, G., and Komaitis, M. (1997). Microbial fatty acid specificity. Folia Microbiol. 42, 117–120. doi: 10.1007/BF02898718
Aguedo, M., Gomes, N., Garcia, E. E., Wache, Y., Mota, M., Teixeira, J. A., et al. (2005). Decalactone production by Yarrowia lipolytica under increased O2 transfer rates. Biotechnol. Lett. 27, 1617–1621. doi: 10.1007/s10529-005-2517-z
Aziz, M., St-Louis, R., Husson, F., and Kermasha, S. (2016). Selected dehydrogenases in Yarrowia lipolytica JMY 861: their role in the synthesis of flavor compounds. Biosci. Biotechnol. Biochem. 80, 2184–2191. doi: 10.1080/09168451.2016.1214531
Bellou, S., Triantaphyllidou, I. E., Aggeli, D., Elazzazy, A. M., Baeshen, M. N., and Aggelis, G. (2016). Microbial oils as food additives: recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 37, 24–35. doi: 10.1016/j.copbio.2015.09.005
Beopoulos, A., Cescut, J., Haddouche, R., Uribelarrea, J.-L., Molina-Jouve, C., and Nicaud, J.-M. (2009). Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 48, 375–387. doi: 10.1016/j.plipres.2009.08.005
Beopoulos, A., Nicaud, J. M., and Gaillardin, C. (2011). An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbiol. Biotechnol. 90, 1193–1206. doi: 10.1007/s00253-011-3212-8
Beopoulos, A., Verbeke, J., Bordes, F., Guicherd, M., Bressy, M., Marty, A., et al. (2014). Metabolic engineering for ricinoleic acid production in the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 98, 251–262. doi: 10.1007/s00253-013-5295-x
Blazeck, J., Liu, L., Knight, R., and Alper, H. S. (2013). Heterologous production of pentane in the oleaginous yeast Yarrowia lipolytica. J. Biotechnol. 165, 184–194. doi: 10.1016/j.jbiotec.2013.04.003
Bourel, G., Nicaud, J. M., Nthangeni, B., Santiago-Gomez, P., Belin, J. M., and Husson, F. (2004). Fatty acid hydroperoxide lyase of green bell pepper: cloning in Yarrowia lipolytica and biogenesis of volatile aldehydes. Enzyme Microb. Technol. 35, 293–299. doi: 10.1016/j.enzmictec.2003.12.014
Braga, A., and Belo, I. (2013). Immobilization of Yarrowia lipolytica for aroma production from castor oil. Appl. Biochem. Biotechnol. 169, 2202–2211. doi: 10.1007/s12010-013-0131-4
Braga, A., and Belo, I. (2014). Production of γ-decalactone by Yarrowia lipolytica: insights into experimental conditions and operating mode optimization. J. Chem. Technol. Biotechnol. 90, 559–565. doi: 10.1002/jctb.4349
Bruder, S., Moldenhauer, E. J., Lemke, R. D., Ledesma-Amaro, R., and Kabisch, J. (2019). Drop-in biofuel production using fatty acid photodecarboxylase from Chlorella variabilis in the oleaginous yeast Yarrowia lipolytica. Biotechnol. Biofuels 12:202. doi: 10.1186/s13068-019-1542-4
Choi, Y. J., and Lee, S. Y. (2013). Microbial production of short-chain alkanes. Nature 502, 571–574. doi: 10.1038/nature12536
Chuang, L. T., Chen, D. C., Nicaud, J. M., Madzak, C., Chen, Y. H., and Huang, Y. S. (2010). Co-expression of heterologous desaturase genes in Yarrowia lipolytica. N. Biotechnol. 27, 277–282. doi: 10.1016/j.nbt.2010.02.006
Cordova, L. T., and Alper, H. S. (2018). Production of α-linolenic acid in Yarrowia lipolytica using low-temperature fermentation. Appl. Microbiol. Biotechnol. 102, 8809–8816. doi: 10.1007/s00253-018-9349-y
Czerwiec, Q., Idrissitaghki, A., Imatoukene, N., Nonus, M., Thomasset, B., Nicaud, J. M., et al. (2019). Optimization of cyclopropane fatty acids production in Yarrowia lipolytica. Yeast 36, 143–151. doi: 10.1002/yea.3379
Darvishi, F., Ariana, M., Marella, E. R., and Borodina, I. (2018). Advances in synthetic biology of oleaginous yeast Yarrowia lipolytica for producing non-native chemicals. Appl. Microbiol. Biotechnol. 102, 5925–5938. doi: 10.1007/s00253-018-9099-x
d’Espaux, L., Ghosh, A., Runguphan, W., Wehrs, M., Xu, F., Konzock, O., et al. (2017). Engineering high-level production of fatty alcohols by Saccharomyces cerevisiae from lignocellulosic feedstocks. Metab. Eng. 42, 115–125. doi: 10.1016/j.ymben.2017.06.004
Dulermo, R., Gamboa-Melendez, H., Ledesma-Amaro, R., Thevenieau, F., and Nicaud, J. M. (2015). Unraveling fatty acid transport and activation mechanisms in Yarrowia lipolytica. Biochim. Biophys. Acta 1851, 1202–1217. doi: 10.1016/j.bbalip.2015.04.004
Dulermo, T., and Nicaud, J. M. (2011). Involvement of the G3P shuttle and β-oxidation pathway in the control of TAG synthesis and lipid accumulation in Yarrowia lipolytica. Metab. Eng. 13, 482–491. doi: 10.1016/j.ymben.2011.05.002
Ethier, S., Woisard, K., Vaughan, D., and Wen, Z. (2011). Continuous culture of the microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for producing docosahexaenoic acid. Bioresour. Technol. 102, 88–93. doi: 10.1016/j.biortech.2010.05.021
Fatma, Z., Hartman, H., Poolman, M. G., Fell, D. A., Srivastava, S., Shakeel, T., et al. (2018). Model-assisted metabolic engineering of Escherichia coli for long chain alkane and alcohol production. Metab. Eng. 46, 1–12. doi: 10.1016/j.ymben.2018.01.002
Fickers, P., Benetti, P. H., Wache, Y., Marty, A., Mauersberger, S., Smit, M. S., et al. (2005). Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 5, 527–543. doi: 10.1016/j.femsyr.2004.09.004
Gao, Q., Cao, X., Huang, Y. Y., Yang, J. L., Chen, J., Wei, L. J., et al. (2018). Overproduction of fatty acid ethyl esters by the oleaginous yeast Yarrowia lipolytica through metabolic engineering and process optimization. ACS. Synth. Biol. 7, 1371–1380. doi: 10.1021/acssynbio.7b00453
Gatter, M., Forster, A., Bar, K., Winter, M., Otto, C., Petzsch, P., et al. (2014). A newly identified fatty alcohol oxidase gene is mainly responsible for the oxidation of long-chain omega-hydroxy fatty acids in Yarrowia lipolytica. FEMS Yeast Res. 14, 858–872. doi: 10.1111/1567-1364.12176
Gemperlein, K., Dietrich, D., Kohlstedt, M., Zipf, G., Bernauer, H. S., Wittmann, C., et al. (2019). Polyunsaturated fatty acid production by Yarrowia lipolytica employing designed myxobacterial PUFA synthases. Nat. Commun. 10:4055. doi: 10.1038/s41467-019-12025-8
Gomes, N., Aguedo, M., Teixeira, J., and Belo, I. (2007). Oxygen mass transfer in a biphasic medium: influence on the biotransformation of methyl ricinoleate into γ-decalactone by the yeast Yarrowia lipolytica. Biochem. Eng. J. 35, 380–386. doi: 10.1016/j.bej.2007.02.002
Gomes, N., Teixeira, J. A., and Belo, I. (2010). The use of methyl ricinoleate in lactone production by Yarrowia lipolytica: aspects of bioprocess operation that influence the overall performance. Biocatal. Biotransformation 28, 227–234. doi: 10.3109/10242422.2010.493208
Gomes, N., Teixeira, J. A., and Belo, I. (2012). Fed-batch versus batch cultures of Yarrowia lipolytica for gamma-decalactone production from methyl ricinoleate. Biotechnol. Lett. 34, 649–654. doi: 10.1007/s10529-011-0824-0
Guangqi, E., Lesage, D., and Ploux, O. (2010). Insight into the reaction mechanism of the Escherichia coli cyclopropane fatty acid synthase: isotope exchange and kinetic isotope effects. Biochimie 92, 1454–1457. doi: 10.1016/j.biochi.2010.05.019
Guo, Z. P., Robin, J., Duquesne, S., O’Donohue, M. J., Marty, A., and Bordes, F. (2018). Developing cellulolytic Yarrowia lipolytica as a platform for the production of valuable products in consolidated bioprocessing of cellulose. Biotechnol. Biofuels 11:141. doi: 10.1186/s13068-018-1144-6
Hammer, S. K., and Avalos, J. L. (2017). Harnessing yeast organelles for metabolic engineering. Nat. Chem. Biol. 13, 823–832. doi: 10.1038/nchembio.2429
He, Q., Bennett, G. N., San, K. Y., and Wu, H. (2019). Biosynthesis of medium-chain omega-hydroxy fatty acids by AlkBGT of Pseudomonas putida GPo1 with native FadL in engineered Escherichia coli. Front. Bioeng. Biotechnol. 7:273. doi: 10.3389/fbioe.2019.00273
Holic, R., Yazawa, H., Kumagai, H., and Uemura, H. (2012). Engineered high content of ricinoleic acid in fission yeast Schizosaccharomyces pombe. Appl. Microbiol. Biotechnol. 95, 179–187. doi: 10.1007/s00253-012-3959-6
Huf, S., Krügener, S., Hirth, T., Rupp, S., and Zibek, S. (2011). Biotechnological synthesis of long-chain dicarboxylic acids as building blocks for polymers. Eur. J. Lipid Sci. Technol. 113, 548–561. doi: 10.1002/ejlt.201000112
Imatoukene, N., Verbeke, J., Beopoulos, A., Idrissi Taghki, A., Thomasset, B., Sarde, C. O., et al. (2017). A metabolic engineering strategy for producing conjugated linoleic acids using the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 101, 4605–4616. doi: 10.1007/s00253-017-8240-6
Jawed, K., Yazdani, S. S., and Koffas, M. A. G. (2019). Advances in the development and application of microbial consortia for metabolic engineering. Metab. Eng. Commun. 9:e00095. doi: 10.1016/j.mec.2019.e00095
Ji, X. J., and Huang, H. (2019). Engineering microbes to produce polyunsaturated fatty acids. Trends Biotechnol. 37, 344–346. doi: 10.1016/j.tibtech.2018.10.002
Ji, X.-J., and Ledesma-Amaro, R. (2020). Microbial lipid biotechnology to produce polyunsaturated fatty acids. Trends Biotechnol. 38, 832–834. doi: 10.1016/j.tibtech.2020.02.003
Kang, W. R., Seo, M. J., An, J. U., Shin, K. C., and Oh, D. K. (2016). Production of delta-decalactone from linoleic acid via 13-hydroxy-9(Z)-octadecenoic acid intermediate by one-pot reaction using linoleate 13-hydratase and whole Yarrowia lipolytica cells. Biotechnol. Lett. 38, 817–823. doi: 10.1007/s10529-016-2041-3
Kim, S., and Gonzalez, R. (2018). Selective production of decanoic acid from iterative reversal of β-oxidation pathway. Biotechnol. Bioeng. 115, 1311–1320. doi: 10.1002/bit.26540
Kim, S. K., and Park, Y. C. (2019). Biosynthesis of ω-hydroxy fatty acids and related chemicals from natural fatty acids by recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 103, 191–199. doi: 10.1007/s00253-018-9503-6
Ko, Y. S., Kim, J. W., Lee, J. A., Han, T., Kim, G. B., Park, J. E., et al. (2020). Tools and strategies of systems metabolic engineering for the development of microbial cell factories for chemical production. Chem. Soc. Rev. 49, 4615–4636. doi: 10.1039/d0cs00155d
Kochan, K., Peng, H., Gwee, E. S. H., Izgorodina, E., Haritos, V., and Wood, B. R. (2019). Raman spectroscopy as a tool for tracking cyclopropane fatty acids in genetically engineered Saccharomyces cerevisiae. Anal. 144, 901–912. doi: 10.1039/c8an01477a
Kolouchová, I., Schreiberová, O., Sigler, K., Masák, J., and Řezanka, T. (2015). Biotransformation of volatile fatty acids by oleaginous and non-oleaginous yeast species. FEMS Yeast Res. 15:fov076. doi: 10.1093/femsyr/fov076
Larroude, M., Rossignol, T., Nicaud, J. M., and Ledesma-Amaro, R. (2018). Synthetic biology tools for engineering Yarrowia lipolytica. Biotechnol. Adv. 36, 2150–2164. doi: 10.1016/j.biotechadv.2018.10.004
Lazar, Z., Liu, N., and Stephanopoulos, G. (2018). Holistic approaches in lipid production by Yarrowia lipolytica. Trends Biotechnol. 36, 1157–1170. doi: 10.1016/j.tibtech.2018.06.007
Ledesma-Amaro, R., Dulermo, R., Niehus, X., and Nicaud, J. M. (2016). Combining metabolic engineering and process optimization to improve production and secretion of fatty acids. Metab. Eng. 38, 38–46. doi: 10.1016/j.ymben.2016.06.004
Ledesma-Amaro, R., and Nicaud, J. M. (2016a). Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol. 34, 798–809. doi: 10.1016/j.tibtech.2016.04.010
Ledesma-Amaro, R., and Nicaud, J. M. (2016b). Yarrowia lipolytica as a biotechnological chassis to produce usual and unusual fatty acids. Prog. Lipid Res. 61, 40–50. doi: 10.1016/j.plipres.2015.12.001
Lee, H., Han, C., Lee, H.-W., Park, G., Jeon, W., Ahn, J., et al. (2018). Development of a promising microbial platform for the production of dicarboxylic acids from biorenewable resources. Biotechnol. Biofuels 11:310. doi: 10.1186/s13068-018-1310-x
Levering, J., Broddrick, J., and Zengler, K. (2015). Engineering of oleaginous organisms for lipid production. Curr. Opin. Biotechnol. 36, 32–39. doi: 10.1016/j.copbio.2015.08.001
Li, J., Jiang, Z., Wu, H., Long, L., Jiang, Y., and Zhang, L. (2009). Improving the recycling and storage stability of enzyme by encapsulation in mesoporous CaCO3-alginate composite gel. Composites Sci. Technol. 69, 539–544. doi: 10.1016/j.compscitech.2008.11.017
Li, S.-Y., Srivastava, R., Suib, S. L., Li, Y., and Parnas, R. S. (2011). Performance of batch, fed-batch, and continuous A-B-E fermentation with pH-control. Bioresour. Technol. 102, 4241–4250. doi: 10.1016/j.biortech.2010.12.078
Li, X., Hu, B., Li, H., and You, B. (2019). Application of artificial intelligence in the diagnosis of multiple primary lung cancer. Thorac. Cancer 10, 2168–2174. doi: 10.1111/1759-7714.13185
Liu, H. H., Ji, X. J., and Huang, H. (2015). Biotechnological applications of Yarrowia lipolytica: past, present and future. Biotechnol. Adv. 33, 1522–1546. doi: 10.1016/j.biotechadv.2015.07.010
Liu, H. H., Madzak, C., Sun, M. L., Ren, L. J., Song, P., Huang, H., et al. (2017a). Engineering Yarrowia lipolytica for arachidonic acid production through rapid assembly of metabolic pathway. Biochem. Eng. J. 119, 52–58. doi: 10.1016/j.bej.2016.12.004
Liu, H. H., Zeng, S. Y., Shi, T. Q., Ding, Y., Ren, L. J., Song, P., et al. (2017b). A Yarrowia lipolytica strain engineered for arachidonic acid production counteracts metabolic burden by redirecting carbon flux towards intracellular fatty acid accumulation at the expense of organic acids secretion. Biochem. Eng. J. 128, 201–209. doi: 10.1016/j.bej.2017.10.007
Liu, H. H., Wang, C., Lu, X. Y., Huang, H., Tian, Y., and Ji, X. J. (2019). Improved production of arachidonic acid by combined pathway engineering and synthetic enzyme fusion in Yarrowia lipolytica. J. Agric. Food Chem. 67, 9851–9857. doi: 10.1021/acs.jafc.9b03727
Liu, J. J., Zhang, C. B., and Lu, W. Y. (2019). Biosynthesis of long-chain omega-hydroxy fatty acids by engineered Saccharomyces cerevisiae. J. Agric. Food Chem 67, 4545–4552. doi: 10.1021/acs.jafc.9b00109
Ma, J., Gu, Y., Marsafari, M., and Xu, P. (2020). Synthetic biology, systems biology, and metabolic engineering of Yarrowia lipolytica toward a sustainable biorefinery platform. J. Ind. Microbiol. Biotechnol. 47, 845–862. doi: 10.1007/s10295-020-02290-8
Ma, Y. R., Wang, K. F., Wang, W. J., Ding, Y., Shi, T. Q., Huang, H., et al. (2019). Advances in the metabolic engineering of Yarrowia lipolytica for the production of terpenoids. Bioresour. Technol. 281, 449–456. doi: 10.1016/j.biortech.2019.02.116
Macario, A., Moliner, M., Corma, A., and Giordano, G. (2009). Increasing stability and productivity of lipase enzyme by encapsulation in a porous organic-inorganic system. Microporous Mesoporous Mater. 118, 334–340. doi: 10.1016/j.micromeso.2008.09.003
Madzak, C. (2018). Engineering Yarrowia lipolytica for use in biotechnological applications: a review of major achievements and recent innovations. Mol. Biotechnol. 60, 621–635. doi: 10.1007/s12033-018-0093-4
Markham, K. A., and Alper, H. S. (2018). Engineering Yarrowia lipolytica for the production of cyclopropanated fatty acids. J. Ind. Microbiol. Biotechnol. 45, 881–888. doi: 10.1007/s10295-018-2067-8
Markham, K. A., Palmer, C. M., Chwatko, M., Wagner, J. M., Murray, C., Vazquez, S., et al. (2018). Rewiring Yarrowia lipolytica toward triacetic acid lactone for materials generation. Proc. Natl. Acad. Sci. U.S.A. 115, 2096–2101. doi: 10.1073/pnas.1721203115
McNeil, B. A., and Stuart, D. T. (2018). Lipomyces starkeyi: an emerging cell factory for production of lipids, oleochemicals and biotechnology applications. World J. Microbiol. Biotechnol. 34:147. doi: 10.1007/s11274-018-2532-6
Meesapyodsuk, D., and Qiu, X. (2008). An oleate hydroxylase from the fungus Claviceps purpurea: cloning, functional analysis, and expression in Arabidopsis. Plant Physiol. 147, 1325–1333. doi: 10.1104/pp.108.117168
Mehrer, C. R., Incha, M. R., Politz, M. C., and Pfleger, B. F. (2018). Anaerobic production of medium-chain fatty alcohols via a β-reduction pathway. Metab. Eng. 48, 63–71. doi: 10.1016/j.ymben.2018.05.011
Miller, K. K., and Alper, H. S. (2019). Yarrowia lipolytica: more than an oleaginous workhorse. Appl. Microbiol. Biotechnol. 103, 9251–9262. doi: 10.1007/s00253-019-10200-x
Mirończuk, A. M., Kosiorowska, K. E., Biegalska, A., Rakicka-Pustułka, M., Szczepañczyk, M., and Dobrowolski, A. (2019). Heterologous overexpression of bacterial hemoglobin VHb improves erythritol biosynthesis by yeast Yarrowia lipolytica. Microb. Cell Fact. 18:176. doi: 10.1186/s12934-019-1231-9
Mishra, P., Lee, N. R., Lakshmanan, M., Kim, M., Kim, B. G., and Lee, D. Y. (2018). Genome-scale model-driven strain design for dicarboxylic acid production in Yarrowia lipolytica. BMC Syst. Biol. 12:12. doi: 10.1186/s12918-018-0542-5
Moradi, H., Asadollahi, M. A., and Nahvi, I. (2013). Improved γ-decalactone production from castor oil by fed-batch cultivation of Yarrowia lipolytica. Biocatal. Agric. Biotechnol. 2, 64–68. doi: 10.1016/j.bcab.2012.11.001
Nawabi, P., Bauer, S., Kyrpides, N., and Lykidis, A. (2011). Engineering Escherichia coli for biodiesel production utilizing a bacterial fatty acid methyltransferase. Appl. Environ. Microbiol. 77, 8052–8061. doi: 10.1128/AEM.05046-11
Ng, T.-K., Yu, A.-Q., Ling, H., Pratomo Juwono, N. K., Choi, W. J., Leong, S. S. J., et al. (2019). Engineering Yarrowia lipolytica towards food waste bioremediation: production of fatty acid ethyl esters from vegetable cooking oil. J. Biosci. Bioeng. 129, 31–40. doi: 10.1016/j.jbiosc.2019.06.009
Nicaud, J. M., Thevenieau, F., Dall, M. T. L., and Marchal, R. (2006). Production of Dicarboxylic Acids by Improved Mutant Strains of Yarrowia lipolytica. European Patent No 1828392B1. Paris: European patent office.
Pagot, Y., Endrizzi, A., Nicaud, J. M., and Belin, J. M. (1997). Utilization of an auxotrophic strain of the yeast Yarrowia lipolytica. Lett. Appl. Microbiol. 25, 113–116.
Park, Y. K., Dulermo, T., Ledesma-Amaro, R., and Nicaud, J. M. (2018a). Optimization of odd chain fatty acid production by Yarrowia lipolytica. Biotechnol. Bioeng. 11:158. doi: 10.1186/s13068-018-1154-4
Park, Y. K., Nicaud, J. M., and Ledesma-Amaro, R. (2018b). The engineering potential of Rhodosporidium toruloides as a workhorse for biotechnological applications. Trends Biotechnol. 36, 304–317. doi: 10.1016/j.tibtech.2017.10.013
Park, Y. K., Ledesma-Amaro, R., and Nicaud, J. M. (2020). De novo biosynthesis of odd-chain fatty acids in Yarrowia lipolytica enabled by modular pathway engineering. Front. Bioeng. Biotechnol. 7:484. doi: 10.3389/fbioe.2019.00484
Pereira de Andrade, D., Carvalho, B. F., Schwan, R. F., and Dias, D. R. (2017). Production of gamma-decalactone by yeast strains under different conditions. Food Technol. Biotechnol. 55, 225–230. doi: 10.17113/ftb.55.02.17.5009
Probst, K. V., Schulte, L. R., Durrett, T. P., Rezac, M. E., and Vadlani, P. V. (2016). Oleaginous yeast: a value-added platform for renewable oils. Crit. Rev. Biotechnol. 36, 942–955. doi: 10.3109/07388551.2015.1064855
Qiao, K., Wasylenko, T. M., Zhou, K., Xu, P., and Stephanopoulos, G. (2017). Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nat. Biotechnol. 35, 173–177. doi: 10.1038/nbt.3763
Řezanka, T., and Sigler, K. (2009). Odd-numbered very-long-chain fatty acids from the microbial, animal and plant kingdoms. Prog. Lipid Res. 48, 206–238. doi: 10.1016/j.plipres.2009.03.003
Rigouin, C., Lajus, S., Ocando, C., Borsenberger, V., Nicaud, J. M., Marty, A., et al. (2019). Production and characterization of two medium-chain-length polydroxyalkanoates by engineered strains of Yarrowia lipolytica. Microb. Cell Fact. 18:99. doi: 10.1186/s12934-019-1140-y
Robles-Rodriguez, C. E., Munoz-Tamayo, R., Bideaux, C., Gorret, N., Guillouet, S. E., Molina-Jouve, C., et al. (2018). Modeling and optimization of lipid accumulation by Yarrowia lipolytica from glucose under nitrogen depletion conditions. Biotechnol. Bioeng. 115, 1137–1151. doi: 10.1002/bit.26537
Rong, S., Yang, S., Li, Q., Cai, B., Guan, S., Wang, J., et al. (2017). Improvement of γ-decalactone production by stimulating the import of ricinoleic acid and suppressing the degradation of γ-decalactone in Saccharomyces cerevisiae. Biocatal. Biotransformation 35, 96–102. doi: 10.1080/10242422.2017.1289182
Runguphan, W., and Keasling, J. D. (2014). Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid-derived biofuels and chemicals. Metab. Eng. 21, 103–113. doi: 10.1016/j.ymben.2013.07.003
Rutter, C. D., and Rao, C. V. (2016). Production of 1-decanol by metabolically engineered Yarrowia lipolytica. Metab. Eng. 38, 139–147. doi: 10.1016/j.ymben.2016.07.011
Santiago-Gómez, M. P., Thanh, H. T., De Coninck, J., Cachon, R., Kermasha, S., Belin, J.-M., et al. (2009). Modeling hexanal production in oxido-reducing conditions by the yeast Yarrowia lipolytica. Process Biochem. 44, 1013–1018. doi: 10.1016/j.procbio.2009.04.028
Seip, J., Jackson, R., He, H., Zhu, Q., and Hong, S. P. (2013). Snf1 is a regulator of lipid accumulation in Yarrowia lipolytica. Appl. Environ. Microbiol. 79, 7360–7370. doi: 10.1128/AEM.02079-13
Seo, J. H., Lee, S. M., Lee, J., and Park, J. B. (2015). Adding value to plant oils and fatty acids: biological transformation of fatty acids into ω-hydroxycarboxylic, α,ω-dicarboxylic, and ω-aminocarboxylic acids. J. Biotechnol. 216, 158–166. doi: 10.1016/j.jbiotec.2015.10.024
Smit, M. S., Mokgoro, M. M., Setati, E., and Nicaud, J. M. (2005). α,ω-Dicarboxylic acid accumulation by acyl-CoA oxidase deficient mutants of Yarrowia lipolytica. Biotechnol. Lett. 27, 859–864. doi: 10.1007/s10529-005-6719-1
Soares, G. P. A., Souza, K. S. T., Vilela, L. F., Schwan, R. F., and Dias, D. R. (2017). γ-decalactone production by Yarrowia lipolytica and Lindnera saturnus in crude glycerol. Prep. Biochem. Biotechnol. 47, 633–637. doi: 10.1080/10826068.2017.1286601
Soong, Y. H. V., Liu, N., Yoon, S., Lawton, C., and Xie, D. (2019). Cellular and metabolic engineering of oleaginous yeast Yarrowia lipolytica for bioconversion of hydrophobic substrates into high-value products. Eng. Life Sci. 19, 423–443. doi: 10.1002/elsc.201800147
Spagnuolo, M., Shabbir Hussain, M., Gambill, L., and Blenner, M. (2018). Alternative substrate metabolism in Yarrowia lipolytica. Front. Microbiol. 9:1077. doi: 10.3389/fmicb.2018.01077
Steen, E. J., Kang, Y., Bokinsky, G., Hu, Z., Schirmer, A., McClure, A., et al. (2010). Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature 463, 559–562. doi: 10.1038/nature08721
Suen, Y. L., Tang, H., Huang, J., and Chen, F. (2014). Enhanced production of fatty acids and astaxanthin in Aurantiochytrium sp by the expression of Vitreoscilla Hemoglobin. J. Agric. Food Chem. 62, 12392–12398. doi: 10.1021/jf5048578
Sun, M. L., Madzak, C., Liu, H. H., Song, P., Ren, L. J., Huang, H., et al. (2017). Engineering Yarrowia lipolytica for efficient γ-linolenic acid production. Biochem. Eng. J. 117, 172–180. doi: 10.1016/j.bej.2016.10.014
Wache, Y., Aguedo, M., Choquet, A., Gatfield, I. L., Nicaud, J. M., and Belin, J. M. (2001). Role of β-oxidation enzymes in γ-decalactone production by the yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 67, 5700–5704. doi: 10.1128/AEM.67.12.5700-5704.2001
Waché, Y., Aguedoa, M., LeDall, M. T., Nicaud, J. M., and Belin, J. M. (2002). Optimization of Yarrowia lipolytica’s β-oxidation pathway for γ-decalactone production. J. Mol. Catal. 19–20, 347–351.
Wang, F., Zhao, J., Li, Q., Yang, J., Li, R., Min, J., et al. (2020). One-pot biocatalytic route from cycloalkanes to α,ω-dicarboxylic acids by designed Escherichia coli consortia. Nat. Commun. 11:5035. doi: 10.1038/s41467-020-18833-7
Wang, G., Xiong, X., Ghogare, R., Wang, P., Meng, Y., and Chen, S. (2016). Exploring fatty alcohol-producing capability of Yarrowia lipolytica. Biotechnol. Biofuels 9:107. doi: 10.1186/s13068-016-0512-3
Wang, J., Ledesma-Amaro, R., Wei, Y., Ji, B., and Ji, X. J. (2020). Metabolic engineering for increased lipid accumulation in Yarrowia lipolytica-a review. Bioresour. Technol. 313:123707. doi: 10.1016/j.biortech.2020.123707
Wang, X., Xia, Q., Wang, F., Zhang, Y., and Li, X. (2019). Modulating heterologous pathways and optimizing culture conditions for biosynthesis of trans-10, cis-12 conjugated linoleic acid in Yarrowia lipolytica. Molecules 24:1753. doi: 10.3390/molecules24091753
Wasylenko, T. M., Ahn, W. S., and Stephanopoulos, G. (2015). The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab. Eng. 30, 27–39. doi: 10.1016/j.ymben.2015.02.007
Werner, N., and Zibek, S. (2017). Biotechnological production of bio-based long-chain dicarboxylic acids with oleogenious yeasts. World J. Microbiol. Biotechnol. 33:194. doi: 10.1007/s11274-017-2360-0
Xie, D. (2017). Integrating cellular and bioprocess engineering in the non-conventional yeast Yarrowia lipolytica for biodiesel production: a review. Front. Bioeng. Biotechnol. 5:65. doi: 10.3389/fbioe.2017.00065
Xie, D., Jackson, E. N., and Zhu, Q. (2015). Sustainable source of omega-3 eicosapentaenoic acid from metabolically engineered Yarrowia lipolytica: from fundamental research to commercial production. Appl. Microbiol. Biotechnol. 99, 1599–1610. doi: 10.1007/s00253-014-6318-y
Xie, D., Miller, E., Sharpe, P., Jackson, E., and Zhu, Q. (2017). Omega-3 production by fermentation of Yarrowia lipolytica: from fed-batch to continuous. Biotechnol. Bioeng. 114, 798–812. doi: 10.1002/bit.26216
Xu, P. (2018). Production of chemicals using dynamic control of metabolic fluxes. Curr. Opin. Biotechnol. 53, 12–19. doi: 10.1016/j.copbio.2017.10.009
Xu, P., Qiao, K., Ahn, W. S., and Stephanopoulos, G. (2016). Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. U.S.A. 113, 10848–10853. doi: 10.1073/pnas.1607295113
Xu, Y., and Qian, S. Y. (2014). Anti-cancer activities of omega-6 polyunsaturated fatty acids. Biomed. J. 37, 112–119. doi: 10.4103/2319-4170.131378
Xue, Z., Sharpe, P. L., Hong, S.-P., Yadav, N. S., Xie, D., Short, D. R., et al. (2013). Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 31, 734–740. doi: 10.1038/nbt.2622
Yan, Q., and Pfleger, B. F. (2020). Revisiting metabolic engineering strategies for microbial synthesis of oleochemicals. Metab. Eng. 58, 35–46. doi: 10.1016/j.ymben.2019.04.009
Yu, A., Zhao, Y., Li, J., Li, S., Pang, Y., Zhao, Y., et al. (2020). Sustainable production of FAEE biodiesel using the oleaginous yeast Yarrowia lipolytica. Microbiologyopen 9:e1051. doi: 10.1002/mbo3.1051
Yu, K. O., Jung, J., Kim, S. W., Park, C. H., and Han, S. O. (2012). Synthesis of FAEEs from glycerol in engineered Saccharomyces cerevisiae using endogenously produced ethanol by heterologous expression of an unspecific bacterial acyltransferase. Biotechnol. Bioeng. 109, 110–115. doi: 10.1002/bit.23311
Yu, T., Zhou, Y. J., Huang, M., Liu, Q., Pereira, R., David, F., et al. (2018). Reprogramming yeast metabolism from alcoholic fermentation to lipogenesis. Cell 174, 1549.e14–1558.e14. doi: 10.1016/j.cell.2018.07.013
Yuzbasheva, E. Y., Mostova, E. B., Andreeva, N. I., Yuzbashev, T. V., Fedorov, A. S., Konova, I. A., et al. (2018). A metabolic engineering strategy for producing free fatty acids by the Yarrowia lipolytica yeast based on impairment of glycerol metabolism. Biotechnol. Bioeng. 115, 433–443. doi: 10.1002/bit.26402
Zhang, B. X., Chen, H. Q., Li, M., Gu, Z. N., Song, Y. D., Colin, R., et al. (2013). Genetic engineering of Yarrowia lipolytica for enhanced production of trans-10, cis-12 conjugated linoleic acid. Microb. Cell Fact. 12:70. doi: 10.1186/1475-2859-12-70
Zhang, B. X., Rong, C. C., Chen, H. Q., Song, Y. D., Zhang, H., and Chen, W. (2012). De novo synthesis of trans-10, cis-12 conjugated linoleic acid in oleaginous yeast Yarrowia lipolytica. Microb. Cell Fact. 11:51. doi: 10.1186/1475-2859-11-51
Zhang, F., Ouellet, M., Batth, T. S., Adams, P. D., Petzold, C. J., Mukhopadhyay, A., et al. (2012). Enhancing fatty acid production by the expression of the regulatory transcription factor FadR. Metab. Eng. 14, 653–660. doi: 10.1016/j.ymben.2012.08.009
Zhang, F., Rodriguez, S., and Keasling, J. D. (2011). Metabolic engineering of microbial pathways for advanced biofuels production. Curr. Opin. Biotechnol. 22, 775–783. doi: 10.1016/j.copbio.2011.04.024
Zhang, H., Feng, Y., Cui, Q., and Song, X. (2017). Expression of Vitreoscilla hemoglobin enhances production of arachidonic acid and lipids in Mortierella alpina. BMC Biotechnol. 17:68. doi: 10.1186/s12896-017-0388-8
Zhang, H., Zhang, L., Chen, H., Chen, Y. Q., Ratledge, C., Song, Y., et al. (2013). Regulatory properties of malic enzyme in the oleaginous yeast, Yarrowia lipolytica, and its non-involvement in lipid accumulation. Biotechnol. Lett. 35, 2091–2098. doi: 10.1007/s10529-013-1302-7
Zhang, S., He, H., Guan, S., Cai, B., Li, Q., and Rong, S. (2020). Bacterial cellulose-alginate composite beads as Yarrowia lipolytica cell carriers for lactone production. Molecules 25:928. doi: 10.3390/molecules25040928
Zhao, Y., Xu, Y., and Jiang, C. (2015). Efficient biosynthesis of γ-decalactone in ionic liquids by immobilized whole cells of Yarrowia lipolytica G3-3.21 on attapulgite. Bioprocess Biosyst. Eng. 38, 2045–2052. doi: 10.1007/s00449-015-1431-6
Zhao, Y. P., Mu, X. Q., and Xu, Y. (2014). Improvement in γ-decalactone production by Yarrowia sp. after genome shuffling. Chem. Pap. 68, 1030–1040. doi: 10.2478/s11696-014-0551-9
Zhou, Y. J., Buijs, N. A., Zhu, Z., Qin, J., Siewers, V., and Nielsen, J. (2016). Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 7:11709. doi: 10.1038/ncomms11709
Keywords: Yarrowia lipolytica, cell factory, fatty acid metabolism, bioproducts, metabolic engineering
Citation: Liu H, Song Y, Fan X, Wang C, Lu X and Tian Y (2021) Yarrowia lipolytica as an Oleaginous Platform for the Production of Value-Added Fatty Acid-Based Bioproducts. Front. Microbiol. 11:608662. doi: 10.3389/fmicb.2020.608662
Received: 21 September 2020; Accepted: 26 November 2020;
Published: 05 January 2021.
Edited by:
Junbiao Dai, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Paul Christakopoulos, Luleå University of Technology, SwedenDebarati Paul, Amity University, India
Copyright © 2021 Liu, Song, Fan, Wang, Lu and Tian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huhu Liu, liuhuhu@hunau.edu.cn; Yun Tian, tianyun@hunau.edu.cn