 Miguel Balado1
Miguel Balado1 Marta A. Lages1Juan C. Fuentes-Monteverde2Diana Martínez-Matamoros2
Marta A. Lages1Juan C. Fuentes-Monteverde2Diana Martínez-Matamoros2 Jaime Rodríguez2
Jaime Rodríguez2 Carlos Jiménez2
Carlos Jiménez2 Manuel L. Lemos1*
Manuel L. Lemos1*- 1Department of Microbiology and Parasitology, Institute of Aquaculture, Universidade de Santiago de Compostela, Santiago de Compostela, Spain
- 2Department of Chemistry, Faculty of Sciences and Center for Advanced Scientific Research (CICA), Universidade da Coruña, A Coruña, Spain
Vibrio anguillarum causes vibriosis, a hemorrhagic septicaemia that affects many cultured marine fish species worldwide. Two catechol siderophores, vanchrobactin and anguibactin, were previously identified in this bacterium. While vanchrobactin is a chromosomally encoded system widespread in all pathogenic and environmental strains, anguibactin is a plasmid-encoded system restricted to serotype O1 strains. In this work, we have characterized, from a serotype O2 strain producing vanchrobactin, a novel genomic island containing a cluster of genes that would encode the synthesis of piscibactin, a siderophore firstly described in the fish pathogen Photobacterium damselae subsp. piscicida. The chemical characterization of this siderophore confirmed that some strains of V. anguillarum produce piscibactin. An in silico analysis of the available genomes showed that this genomic island is present in many of the highly pathogenic V. anguillarum strains lacking the anguibactin system. The construction of single and double biosynthetic mutants for vanchrobactin and piscibactin allowed us to study the contribution of each siderophore to iron uptake, cell fitness, and virulence. Although both siderophores are simultaneously produced, piscibactin constitute a key virulence factor to infect fish, while vanchrobactin seems to have a secondary role in virulence. In addition, a transcriptional analysis of the gene cluster encoding piscibactin in V. anguillarum showed that synthesis of this siderophore is favored at low temperatures, being the transcriptional activity of the biosynthetic genes three-times higher at 18°C than at 25°C. We also show that iron levels and temperature contribute to balance the synthesis of both siderophores.
Introduction
Vibrio anguillarum is a bacterium inhabitant of marine environments and also one of the most important pathogens in the aquaculture industry worldwide. It is the causative agent of vibriosis, a fatal hemorrhagic septicaemia affecting warm- and cold-water fish of highly economic importance (Toranzo et al., 2005). V. anguillarum isolates are classified into 23 different O-serogroups (O1 to O23) according to the European serotyping system (Pedersen et al., 1999), although most virulent strains belong to serotypes O1, O2 and, to a lesser extent, O3. The remaining V. anguillarum serotypes are mostly environmental strains isolated from seawater, marine animals or sediments (Frans et al., 2011; Toranzo et al., 2017).
It is well known that bacterial virulence is a multifactorial trait and that pathogenic bacteria must have a number of factors that enable them to colonize the host and establish an infection (Miller et al., 1989). The virulence-related factors identified in V. anguillarum so far include those related to chemotaxis and motility (O’Toole et al., 1996; Ormonde et al., 2000), adhesion (Wang et al., 1998), invasion (Croxatto et al., 2007), secretion of extracellular enzymes (Denkin and Nelson, 2004; Rodkhum et al., 2005; Li et al., 2008) and several iron uptake mechanisms (Li and Ma, 2017). Despite this knowledge, the pathogenesis of V. anguillarum is not completely understood (Hickey and Lee, 2017).
Iron restriction is an important host defense strategy, thus successful pathogens must possess mechanisms to acquire iron from host sources in order to cause disease. In V. anguillarum two catechol siderophores, vanchrobactin and anguibactin, have been described (Actis et al., 1986; Soengas et al., 2006; Lemos et al., 2010; Li and Ma, 2017). While anguibactin is encoded by pJM1-type plasmids and is restricted to some virulent strains of serotype O1, vanchrobactin is a chromosomally encoded siderophore that is widespread in all V. anguillarum isolates either environmental or pathogenic (Lemos et al., 2010; Li and Ma, 2017). It is noticeable that vanchrobactin is not produced by serotype O1 strains harboring a pJM1-type plasmid since the presence of this plasmid results in the inactivation of vanchrobactin synthesis by a transposition event from the plasmid to the vanchrobactin chromosomal gene cluster (Naka et al., 2008). Thus, to date the simultaneous production of more than one siderophore was not reported in V. anguillarum (Naka et al., 2008; Rønneseth et al., 2017). However, it was detected the simultaneous presence of anguibactin and vanchrobactin in a marine Vibrio sp. (Sandy et al., 2010). From previous reports, it is clear that anguibactin is a key virulence factor for V. anguillarum serotype O1 strains (Wolf and Crosa, 1986; Naka et al., 2011). However, the role of vanchrobactin in virulence remains unknown so far.
The in silico analysis of the V. anguillarum serotype O2 strain RV22 genome sequence (accession number AEZB00000000) showed the existence of a gene cluster whose closest homolog is the irp cluster encoding the siderophore piscibactin (Naka et al., 2011). This siderophore was firstly identified in the fish pathogen Photobacterium damselae subsp. piscicida (Souto et al., 2012) and it is one of the main virulence factors of this bacterium (Osorio et al., 2015). Piscibactin synthesis and transport are encoded by a pathogenicity island (PAI) located in the 69-kb plasmid pPHDP70 (Osorio et al., 2006, 2015). The presence of irp genes in V. anguillarum RV22 suggests that this strain, besides producing vanchrobactin, likely produces also piscibactin or a piscibactin-like siderophore, which would imply the existence of a third siderophore in V. anguillarum.
In this work, we describe a novel genomic island present in many virulent strains of V. anguillarum that encodes the piscibactin system. We demonstrate that some serotype O2 strains produce in fact piscibactin and that this siderophore plays a relevant role in the cell fitness and contribute to a greater extent than vanchrobactin to the virulence for fish. We also show that the synthesis of both siderophores is balanced in response to iron levels and to the growth temperature.
Materials and Methods
Bacterial Strains, Plasmids, and Media
The bacterial strains and plasmids used are listed in Table 1. V. anguillarum and P. damselae subsp. piscicida (Pdp) strains were routinely grown at 25°C on tryptic soy agar (TSA) or broth (TSB) (Cultimed) supplemented with 1% NaCl (TSA-1 or TSB-1). Escherichia coli strains were routinely grown at 37°C in Luria-Bertani (LB) broth and LB agar (Cultimed). When it was required, antibiotics were added at the following final concentrations: kanamycin (Km) at 50 μg mL-1, ampicillin (Ap) sodium salt at 50 μg mL-1, and gentamicin (Gm) at 12 μg mL-1.

TABLE 1. Strains and plasmids used in this study.
Construction of irp1 Defective Mutants by Allelic Exchange and Complementation
In-frame (non-polar) deletions of irp1 were constructed by allelic exchange in V. anguillarum RV22 wild type and ΔvabF (vanchrobactin synthesis deficient) backgrounds using the suicide vector pNidKan as previously described (Balado et al., 2006). The oligonucleotides used are shown in Table 2. The mutagenesis process led to the generation of V. anguillarum single mutant strains RV22Δirp1 (MB280) and the double mutant RV22ΔvabFΔirp1 (MB283). To ensure that all mutations were in frame, the deleted region was sequenced by the Sanger method using primers irp1_ang_1_XbaI and irp1_ang_4_EcoRI (Table 2). pPHDP70 plasmid (from Pdp DI21 and containing all necessary genes for piscibactin synthesis) marked with a Km cassette (pPHDP70::kan) (Osorio et al., 2015) was mobilized into the V. anguillarum RV22ΔvabFΔirp1 double mutant to restore piscibactin synthesis.

TABLE 2. Oligonucleotides used for construction of mutants by allele exchange, transcriptional fusions, and RT-PCR assays.
Growth Promotion and Siderophore Production Assays
Growth promotion assays were performed using 96-well microtiter plates. Each well contained 200 μL of CM9 medium (Lemos et al., 1988) supplemented with 10 μM FeCl3 to achieve iron excess conditions, or with the iron chelators EDDA [ethylenediamine-di(o-hydroxyphenyl-acetic acid)] 5 μM or 100 μM 2,2′-dipyridyl, to achieve iron restricted conditions. Each well was inoculated with a 1:50 dilution of a V. anguillarum overnight culture in TSB-1 adjusted to an OD600 = 0.5. The plates were incubated at 25°C or 18°C with shaking at 150 rpm. Growth (OD600) was recorded during 24 h in an iMACK Microplate reader (Bio-Rad). Bacterial cultures in CM9 with 50 μM 2,2′-dipyridyl and an OD600 ≈ 0.8 (after 6 h of incubation) were used to obtain supernatants and measure siderophore production using the chrome azurol-S (CAS) liquid assay (Schwyn and Neilands, 1987). For this purpose, equal volumes of cell free supernatants and CAS reagent were mixed and, after 15 min of incubation at room temperature, A630 was measured in a spectrophotometer (Hitachi).
Cross-Feeding Assays
To test whether V. anguillarum was able to use piscibactin as iron source, a cross-feeding assay was conducted using RV22 (wild type strain) and RV22ΔvabD mutant as indicator strains. These strains were inoculated into CM9 plates as follows: 0.5 mL of a TSB-1 culture at an OD600 = 0.5 were included into 20 mL of CM9 medium containing 0.8% agarose and 120 μM of the iron chelator 2,2′-dipyridyl, a concentration close to the MIC and at which growth halos are easily visualized (Balado et al., 2006). V. anguillarum strains to be tested for piscibactin production and P. damselae subsp. piscicida DI21 (wild type strain, piscibactin producer) or a cured strain (DI21 lacking pPHDP70, unable to produce piscibactin and used as negative control) were cultured in TSA-1 plates and the cells were harvested with a sterile loop and placed on the surface of the plates inoculated with the V. anguillarum strains. The presence of growth halos of the V. anguillarum indicator strains around cells of P. damselae subsp. piscicida DI21 or V. anguillarum after overnight incubation at 25°C was indicative of piscibactin utilization.
Piscibactin Detection by Mass Spectrometry
Siderophore piscibactin detection was carried out in cell free supernatants from cultures of V. anguillarum grown at 25°C in flasks containing 1 L of CM9 minimal medium with 40 μM 2,2′-dipyridyl with continuous shaking at 150 rpm for ca. 24 h (Souto et al., 2012). Flasks were inoculated with 20 mL of a fresh culture of V. anguillarum RV22wt, RV22ΔvabF or RV22ΔvabD mutants in TSB-1. When bacterial cultures achieved an OD600 = 1.0, 10 mg of GaBr3 were added to each 1.0-L batch. After 12 h of incubation at 4°C, bacterial cells were removed by centrifugation at 10,000 ×g for 10 min (Beckman J-21 High Speed Centrifuge) and supernatants were filtered through a 0.45-μm pore size membrane by a continuous filtration cartridge (Millipore). Subsequently, ca. 900 mL of each cell-free supernatant were concentrated under vacuum to 410 mL. Fifty milliliters of the solution were passed through an Oasis hydrophilic lipophilic (HLB) cartridge (Waters) (35 cm3, 6 g) (Espada et al., 2011), which was previously conditioned and equilibrated with 60 mL of acetonitrile (solvent B) and 60 mL of water (solvent A). They were eluted with 30 mL of the following mixtures of water and acetonitrile: 1:0, 7:3, 1:1, 3:7, and 0:1. The fraction eluted with a mixture of water and acetonitrile (1:1), named RV22WTF3, RV22ΔvabFF3, and RV22ΔvabDF3, were subjected to high-performance liquid chromatography (HPLC)-high-resolution electrospray ionization mass spectrometry (HRESIMS) analysis using an Atlantis dC18 column (100 mm × 4.6 mm, 5 μm) with a 35 min gradient from 10 to 100% CH3CN-H2O, then 5 min at 100% CH3CN, and finally a 10 min gradient from 100 to 10% H2O-CH3CN, at a flow rate of 1 mL min-1. Piscibactin-Ga(III) complex was detected with a tR of 9.56 min from RV22WTF3 and with a tR of 9.71 min from RV22ΔvabFF3 by comparison of the tR, HRESIMS and UV spectral data to those reported for that compound isolated from P. damselae subsp. piscicida DI21 (Souto et al., 2012). Vanchrobactin was detected with a tR of 2.74 min from RV22WTF3.
RNA Purification and RT-PCR
The organization of piscibactin genes into operon(s) was tested by reverse transcription PCR. V. anguillarum RV22 was grown until exponential phase (ca. OD600 = 1) in 10 mL CM9 medium containing 5 μM EDDA or 10 μM Fe2(SO4)3. Cells were pelleted by centrifugation at 10,000 ×g for 10 min and total RNA was isolated with RNAwiz (Ambion) following the manufacturer’s recommendations. RT-PCR was performed with 1 μg RNA pre-treated with RQ1 RNase-Free DNase (Promega) by using the M-MLV reverse transcriptase (Invitrogen). An appropriate primer (Table 2), located at the 3′-end of irp5 gene, was used to obtain a cDNA that was then used as template for three PCR reactions targeted into araC1 (PCR1), frpA (PCR2) and between irp2 3′-end and irp1 5′-end (PCR3) (Figure 1). All primers used are listed in Table 2. A negative control reaction was performed with total RNA without M-MLV reverse transcriptase to confirm the lack of genomic DNA contamination in each reaction mixture. The PCR positive control reaction was done using 100 ng of genomic DNA as template.
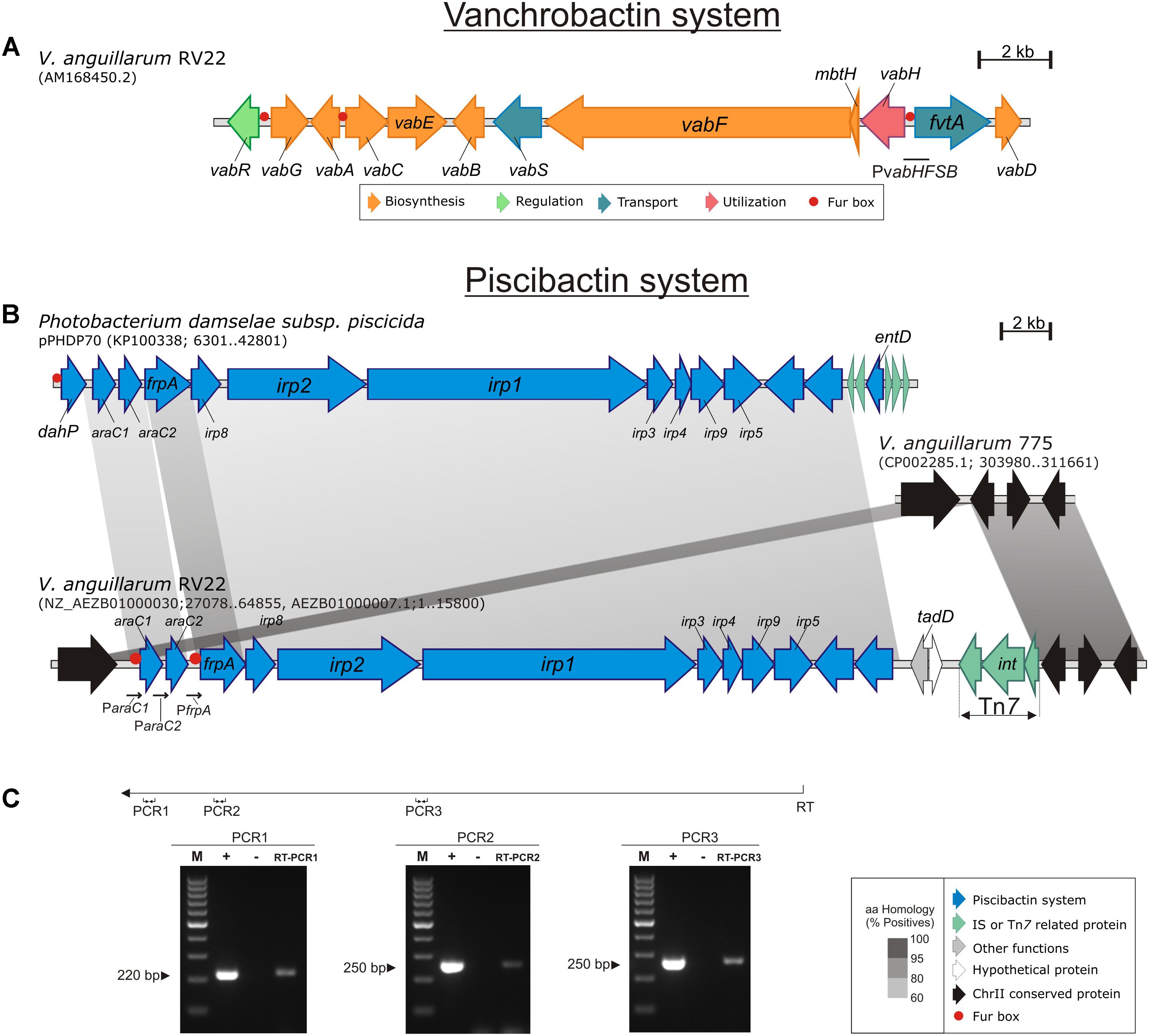
FIGURE 1. Genetic maps of the gene clusters encoding the vanchrobactin (A) and the piscibactin (B) systems in V. anguillarum RV22. In (B) the piscibactin genes, which are part of the plasmid pPHDP70 of P. damselae subsp. piscicida are included for comparative purposes. Gray blocks indicate percentages of similarity in the proteins sequence. (C) Displays the results of three RT-PCRs reactions designed to analyze the transcription of the piscibactin gene cluster in RV22. An appropriate primer (Primer RT, Table 2), located at the 3′-end of irp5 gene, was used to obtain a cDNA (denoted as RT in the figure) that was then used as template for three PCR reactions targeted into araC1 (PCR1), frpA (PCR2) and between irp2 3′-end and irp1 5′-end (PCR3). M, size marker from 100 to 1,000 bp; +, positive control PCR using genomic DNA as template; -, negative control PCR using total RNA without reverse transcriptase; RT-PCR1-3, results of the three different RT-PCRs.
lacZ Transcriptional Fusions and β-Galactosidase Assays
DNA fragments corresponding to V. anguillarum araC1, araC2 and frpA promoter regions were amplified by PCR. The PCR-amplified promoter regions were fragments of about 700 bp, including the region upstream of the start codon and the first nucleotides of the araC1, araC2 or frpA coding sequences (ca. 50 bp). These putative promoter regions were fused to a promoterless lacZ gene in the low-copy-number reporter plasmid pHRP309 (Parales and Harwood, 1993). The resulting transcriptional fusion constructs, araC1::lacZ (pMB34), araC2::lacZ (ML214) and frpA::lacZ (pMB33), were mobilized from E. coli S17-1 aaapir into V. anguillarum strains RV22 wild type or RV22ΔvabD mutant by conjugation. The construction vabH::lacZ (pMB12), corresponding to the promoter of the vanchrobactin operon (vabH) (Balado et al., 2008), was used as reference of a siderophore promoter. A housekeeping gene promoter (PproC) was used as control (Savioz et al., 1990). The V. anguillarum strains carrying the promoter–lacZ fusions or the plasmid pHRP309 alone (negative control) were grown in CM9 minimal medium under different iron-availability conditions. The β-galactosidase (LacZ) activities were measured by the method of Miller (1992). Results showed are means of three independent experiments, each one measured in triplicate.
In Silico Analysis of irpang Genomic Island Structure and Analysis of Its Distribution
The NCBI services were used to consult DNA and protein sequence databases using BLAST algorithms. All V. anguillarum genome sequences (43 genomes) deposited in GenBank were screened for the presence of the irpang genomic island. For this purpose, the whole nucleotide sequence of irpang genomic island of V. anguillarum RV22 and the 3′ and 5′-ends surrounding regions were used as query in a BLASTN search. Nucleotide sequences producing significant alignments were downloaded and a multiple alignment of the conserved genomic sequences was obtained by using MAUVE (Darling et al., 2010). The alignment was used to compare the genomic island structure and to identify the insertion point. The Virtual Footprint Promoter Matches from the online database PRODORIC Release 8.91 (Münch et al., 2003) was used to identify putative Fur-box sequences. BLASTP algorithm was also used to analyze homology of each protein encoded by the irpang gene cluster from V. anguillarum with its counterpart in P. damselae subsp. piscicida. All this information was used to create Figure 1 and Supplementary Figure S1.
Fish Virulence Assays
Virulence assays were carried out with turbot (Psetta maxima) fingerlings with an average weight of 5 g. Fish were divided in groups of 30 animals. All fish groups, one per strain tested and a control group, were maintained in 50-L seawater tanks at 18°C with continuous aeration and water recirculation. The bacterial inoculum was prepared as follows. Several colonies from a 24 h TSA-1 culture were suspended into saline solution (0.85% NaCl) to get a cell suspension with an OD600 = 0.5. The inoculum used was a 10-fold dilution of this suspension in saline solution. The actual number of injected bacterial cells was determined by plate count on TSA-1. Fish were inoculated intraperitoneally (ip) with 100 μL of bacterial suspensions, ranging the doses used between 2 and 4 × 104 CFU per fish. A control group was inoculated with 100 μL of saline solution. Mortalities were recorded daily during 10 days after injection and statistical significance of differences in percentage survival for V. anguillarum strains was determined using the Kaplan-Meier method with Mantel-Cox log-rank test using SPSS (version 20; IBM SPSS Inc., Chicago, IL, United States). P-values were considered significant when P was <0.05, <0.01, and <0.001. All protocols for animal experimentation used in this study have been reviewed and approved by the Animal Ethics Committee of the University of Santiago de Compostela.
Results
V. anguillarum Strain RV22 Harbors a Novel Genomic Island Likely Encoding Piscibactin Synthesis
In a previous work, we described and characterized the gene cluster involved in the synthesis and transport of vanchrobactin system in V. anguillarum RV22 (Figure 1A) (Balado et al., 2006). Inactivation of vanchrobactin synthesis resulted in a drastic decrease in siderophore production by this strain and in its growth ability under iron starvation conditions. However, when the complete genome sequence of RV22 was described (Naka et al., 2011), it was found that chromosome II contains a likely additional siderophore gene cluster with high similarity to the irp genes encoding piscibactin in P. damselae subsp. piscicida (Souto et al., 2012).
This irp gene cluster in V. anguillarum RV22 strain (that we named irpang) is part of a DNA region of ca. 40 kb located in chromosome II (between locus AEH34691 and AEH34692) (Figure 1B). This cluster shows identical gene structure and organization as the irp cluster described in P. damselae subsp. piscicida as part of pPHDP70 plasmid (Figure 1B) (Souto et al., 2012; Osorio et al., 2015). Each CDS present in the irpang cluster shows an amino acid similarity between 52 and 66% with the irp cluster ortholog from P. damselae subsp. piscicida (Supplementary Table S1). Although irpang genes include most of the functions for piscibactin synthesis and utilization (Supplementary Table S1), the cluster lacks an entD homolog, which encodes a 4′-phosphopantetheinyl transferase, that is required to activate the peptide synthesis domains of non-ribosomal peptide synthetases (NRPS) (Crosa and Walsh, 2002). However, the vab gene cluster (Figure 1A), encoding the vanchrobactin system, does include a 4′-phosphopantetheinyl transferase gene (vabD) (Balado et al., 2008) that could complement in trans this function (see below).
In addition, there are three orfs (loci: VAR_RS19255, VAR_RS0101310, and VAR_RS19260), adjacent to irpang, which encode a probable transposon belonging to the Tn7 superfamily (Figure 1B). The nucleotide sequence (accession No. AEZB01000007.1; region 10925–14484) including these three orfs shows a 86% nucleotide identity to the Tn7 transposon genes of the V. parahaemolyticus pathogenicity island (Vp-PAI) that encodes a type three secretion system (T3SSα) (Okada et al., 2009). Tn7 transposons are widespread in bacteria and are involved in the formation of some genomic islands that have the common feature of not being inserted near of tRNAs genes (Sugiyama et al., 2008). These findings suggest that irpang could be part of a novel genomic island. Interestingly, a BLASTN search revealed that four of highly virulent V. anguillarum strains (two belonging to serotype O1 and two from serotype O2) (Rønneseth et al., 2017) contain the irpang genes inserted in the same chromosomal location (Supplementary Figure S1). However, in two of these strains the irpang genes are not associated with transposon genes (Supplementary Figure S1). The genome sequences of many V. anguillarum strains were recently reported (Busschaert et al., 2015; Castillo et al., 2017; Holm et al., 2018). Thus, we performed an in silico search within the 43 genomes available in GenBank and we could find the presence of irpang gene cluster in several of these strains, some of which are highly virulent (Table 3). This fact suggests that the piscibactin cluster could have a relevant role in virulence.
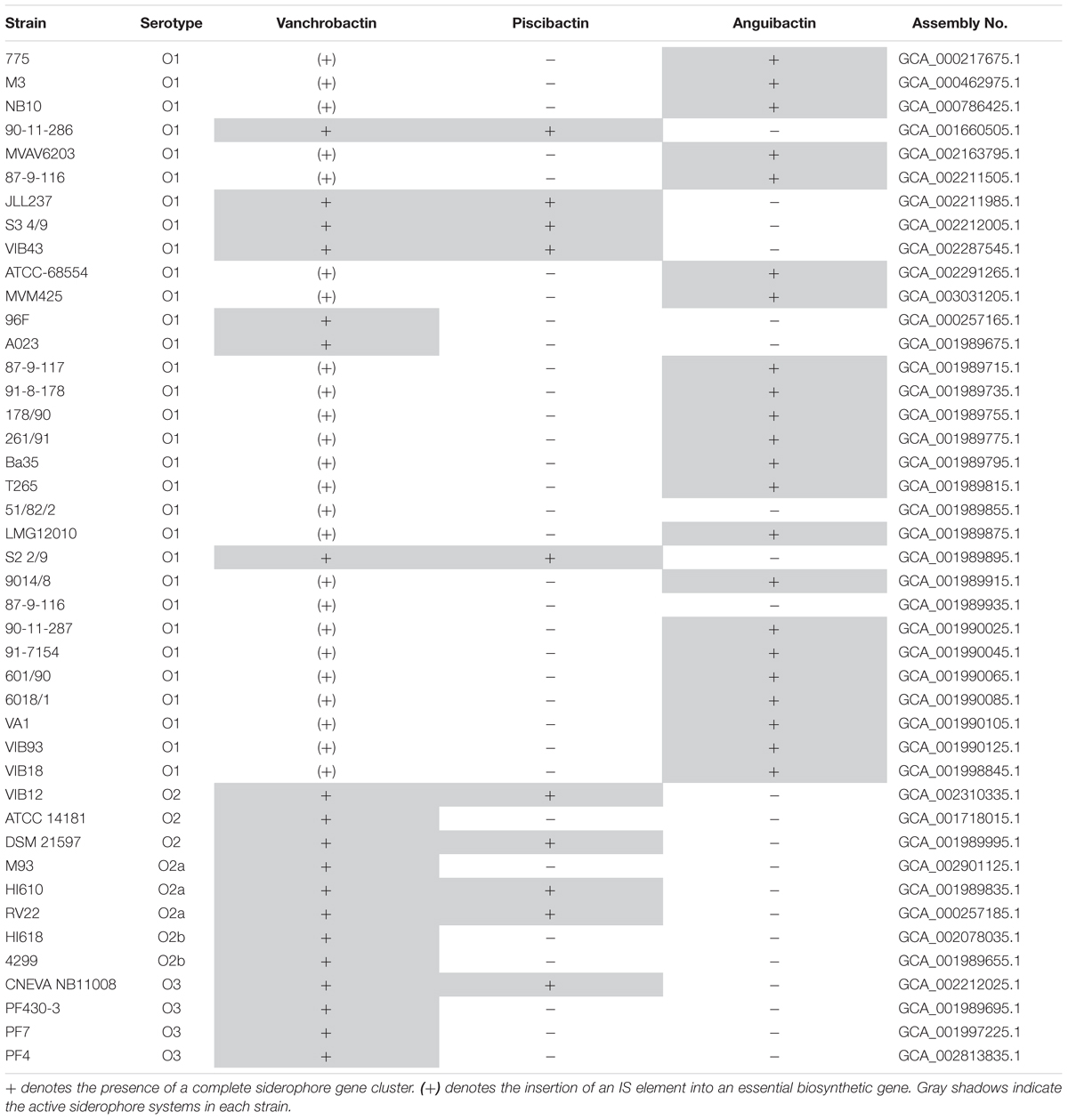
TABLE 3. Distribution of vanchrobactin, piscibactin and anguibactin gene clusters among the V. anguillarum genome projects available in GenBank.
To test if irpang genes are actually expressed, a series of reverse-transcriptase PCR reactions were conducted. The results showed that this gene cluster is transcribed in a polycistronic mRNA comprising araC1, araC2, frpA, irp1-5, irp8 and irp9 genes (Figures 1B,C). Therefore, the 10 genes putatively encoding piscibactin biosynthetic, regulatory, and uptake functions could be co-transcribed from the promoter upstream of araC1. A similar result was found for the piscibactin gene cluster harbored by P. damselae subsp. piscicida plasmid pPHDP70, which is transcribed in a single polycistronic mRNA from the promoter upstream of dahP (Osorio et al., 2006). Interestingly, the irpang cluster does not include a dahP homolog and many other differences were found at the nucleotide level in intergenic regions. Thereby, even though irpang genes can be transcribed from a promoter upstream of araC1, the existence of additional promoters, particularly upstream of frpA, cannot be ruled out (see below).
V. anguillarum RV22 Produces Vanchrobactin and Piscibactin Simultaneously
To demonstrate that RV22 synthesizes piscibactin in addition to vanchrobactin (Soengas et al., 2006, 2007), we used the previously described methodology, based on the use of HLB cartridges and liquid chromatography-mass spectrometry (LC-MS), that we developed for isolation of piscibactin (Souto et al., 2012). Vanchrobactin was first detected with a retention time (tR) of 2.74 min (Figures 2A,B), which showed the [M+H]+ and [M+Na]+ ions at m/z 398.1667 and 420.1487, respectively (Figure 2C). Furthermore, from the cell-free supernatants of RV22 cultures under iron restriction and after addition of GaBr3, in order to stabilize the siderophore as its corresponding complex, it was detected the characteristic piscibactin-Ga(III) complex with (tR) of 9.56 min (Figure 2D). The chemical identification was done on the basis of its tR value, UV spectral data and HRESIMS analysis. Thus, piscibactin-Ga(III) complexes obtained from V. anguillarum RV22 and from P. damselae subsp. piscicida (Souto et al., 2012) showed similar retention times and absorbance maxima (223, 267, and 354 nm) in its UV spectrum (Figure 2F) in HPLC analysis. Moreover, HRESIMS analyses showed the [M+H]+ ions at m/z 519.9949/521.9938 (calculated for C19H21N3O4S3Ga, 519.9950/521.9941), with the distinctive isotopic ratio of gallium (Mr = 69 and 71, ratio 3:2) (Figure 2E), indicating the same molecular mass to that reported for piscibactin-Ga (III) complex isolated from P. damselae subsp. piscicida (Souto et al., 2012).
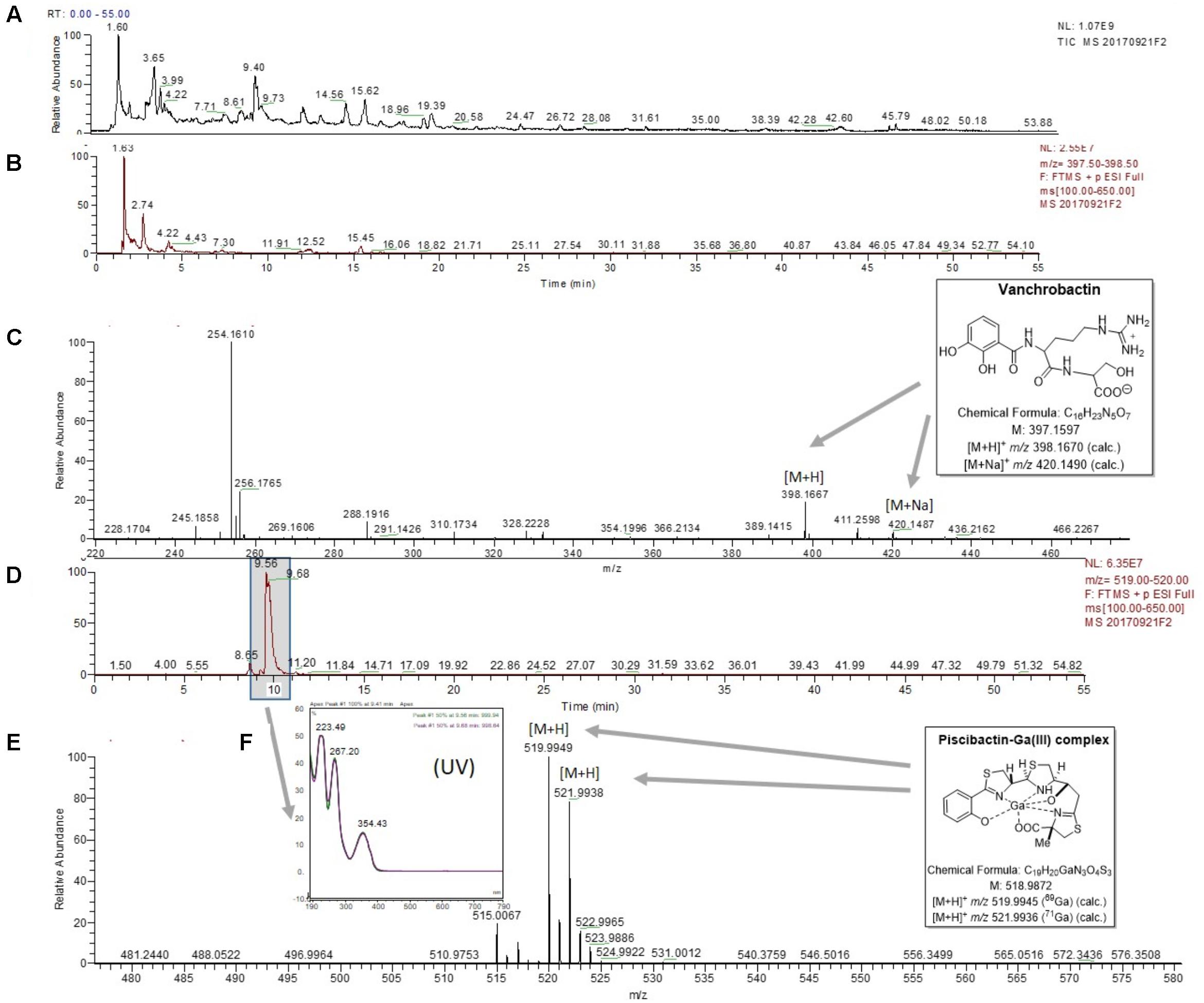
FIGURE 2. LC-MS experiments for the detection of vanchrobactin and piscibactin-Ga(III) complex in the RV22wt strain. (A) Total ion chromatogram (TIC) of the fraction RV22WTF3 eluted with H2O-CH3CN (1:1) from the Oasis HLB cartridge. (B) Extracted mass chromatogram (m/z 397.50 to 398.50) showing the peak with retention time of 2.74 min. (C) (+)-HRESIMS of the peak at tR of 2.74 min identified as vanchrobactin: m/z 398.1667 ([M+H]+; calc. for C16H24N5O7, 398.1670) and m/z 420.1487 ([M+Na]+; calc. for C16H23N5O7Na, 420.1490). (D) Extracted mass chromatogram (m/z 519.00 to 520.00) showing the peak with retention time of 9.56 min. (E) (+)-HRESIMS of the peak at tR of 9.56 min identified as piscibactin-Ga(III) complex: m/z 519.9946/521.9933 ([M+H]+, calc. for C19H21N3O4S3Ga, 519.9950/521.9941). (F) UV spectrum of the peak at tR of 9.56 min.
As expected, chemical analysis of cell-free supernatants of the V. anguillarum RV22ΔvabF mutant (unable to produce vanchrobactin), following the same methodology, showed the absence of vanchrobactin (Figures 3A–C) and the presence of a peak with a tR of 9.71 min (Figure 3D), displaying a m/z value of 519.9945/521.9933 (Figure 3E) and a UV spectrum (Figure 3F) which correspond to piscibactin-Ga(III) complex. The same methodology was used to test piscibactin synthesis by V. anguillarum RV22ΔvabD mutant (Balado et al., 2008), which lacks an active 4′-phosphopantetheinyl transferase. In this mutant, neither vanchrobactin or piscibactin were detected (Supplementary Figure S2), confirming the vabD gene is essential for the synthesis of both siderophores.
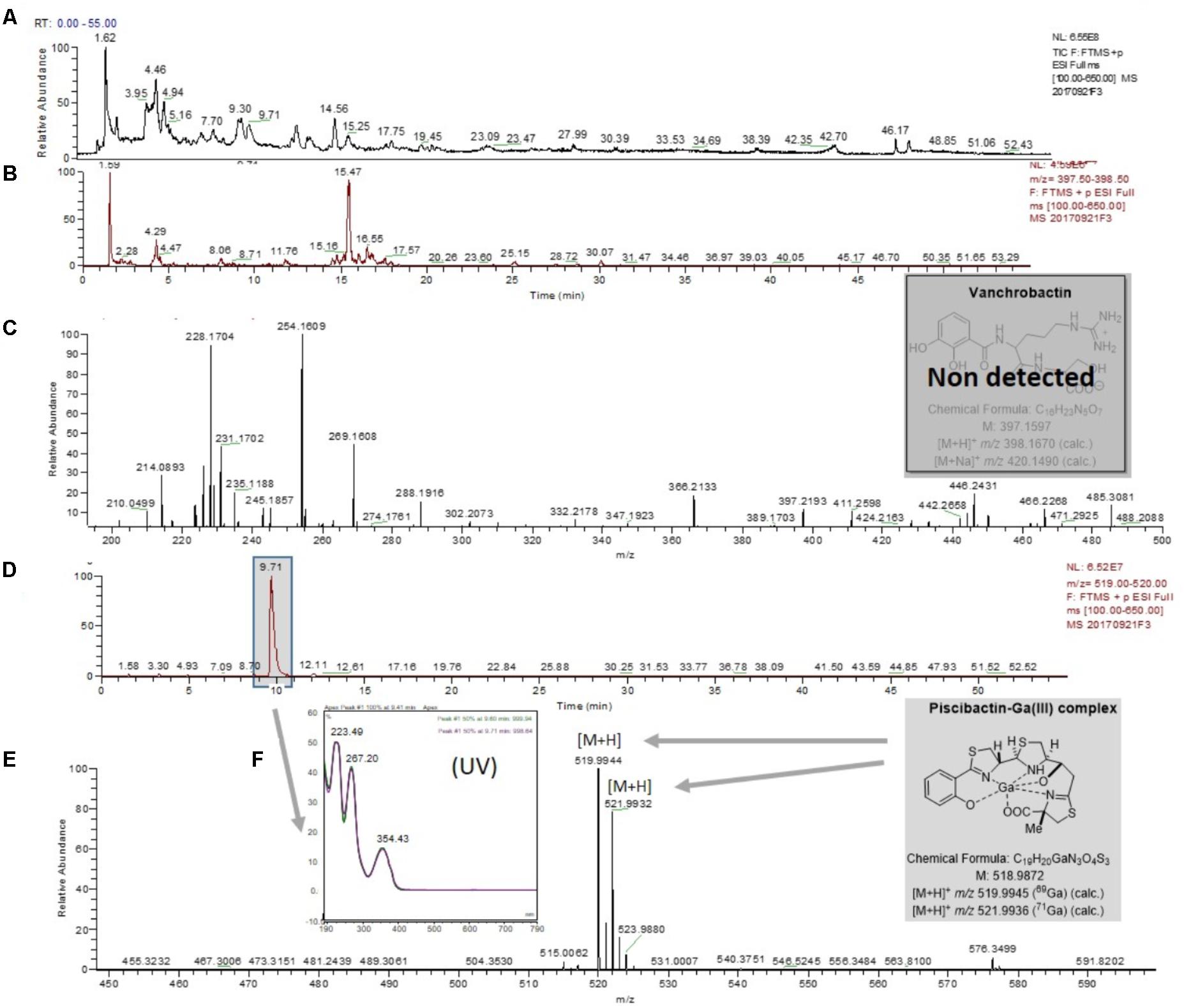
FIGURE 3. LC-MS experiments for the detection of piscibactin-Ga(III) complex in the RV22ΔvabF mutant strain. (A) Total ion chromatogram (TIC) of the fraction RV22ΔvabFF3 eluted with H2O-CH3CN (1:1) from the Oasis HLB cartridge. (B) Extracted mass chromatogram (m/z 397.50 to 398.50). (C) (+)-HRESIMS corresponding to retention time window from tR of 0.19–3.00 min which shows the lack of vanchrobactin. (D) Extracted mass chromatogram (m/z 519.00 to 520.00) showing the peak with retention time of 9.71 min. (E) (+)-HRESIMS of the peak at tR of 9.71 min identified as piscibactin-Ga(III) complex: m/z 519.9944/521.9922 ([M+H]+, calc. for C19H21N3O4S3Ga, 519.9950/521.9941). (F) UV spectrum of the peak at tR of 9.71 min.
In addition, cross-feeding assays were used to ascertain piscibactin production and utilization by V. anguillarum RV22 (Figure 4). RV22 wild type and RV22ΔvabD mutant could be cross-fed by a piscibactin-producing strain (P. damselae subsp. piscicida DI21). The V. anguillarum mutant RV22ΔvabF (impaired for vanchrobactin production) still could cross-fed both indicator strains, indicating the production of a second siderophore. However, no growth halo was obtained when using RV22ΔvabD mutant as tested strain (Figure 4). All these results prove that V. anguillarum RV22 is able to produce and use piscibactin, which denotes that irpang genes are functional. Moreover, the results clearly suggest that irpang gene cluster needs a functional vabD gene (encoding the 4′-phosphopantetheinyl transferase) to complete the biosynthesis of piscibactin.
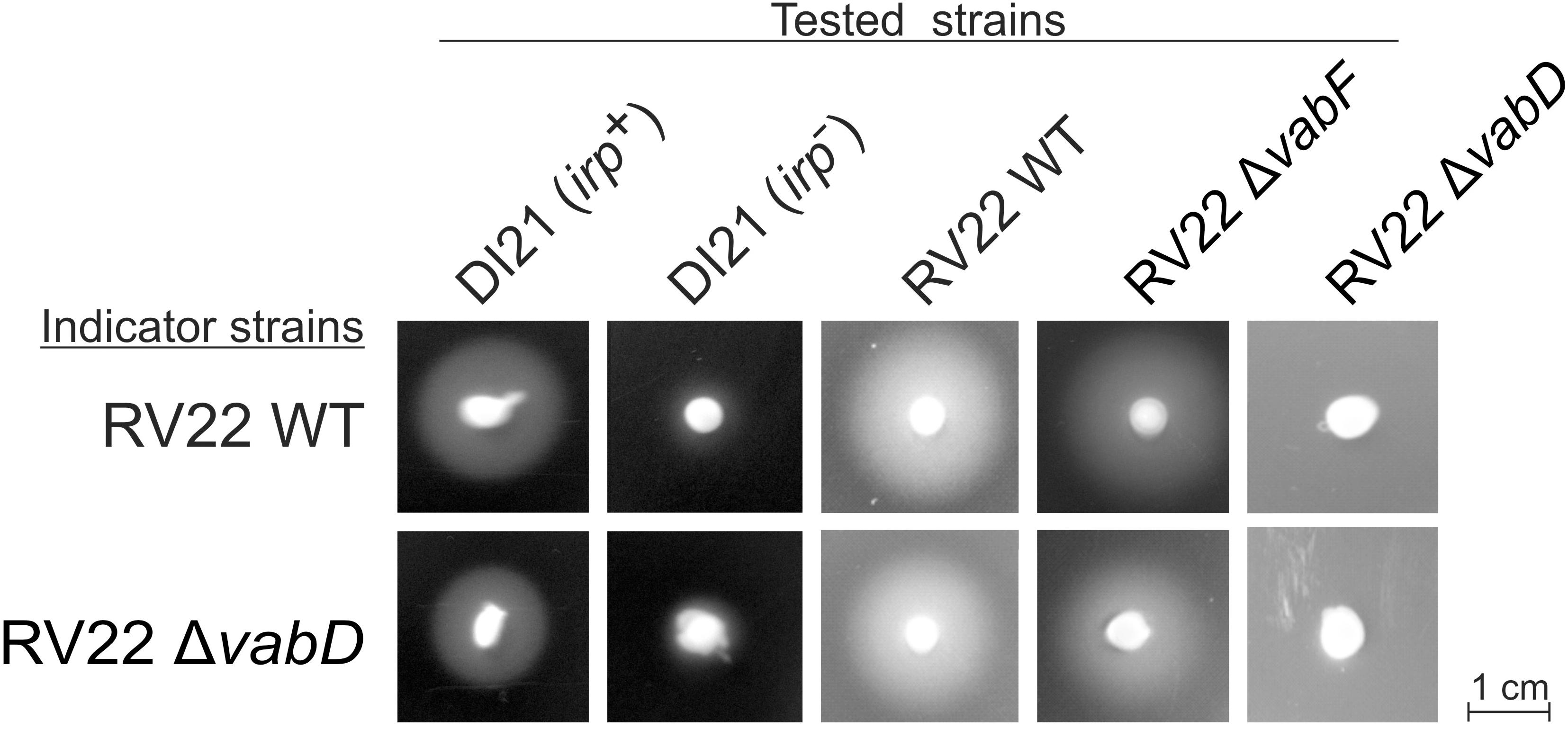
FIGURE 4. Cross feeding assay to determine the production and use of piscibactin by V. anguillarum RV22 wild type and RV22ΔvabD mutant. Indicator strains are those inoculated within the CM9 plates containing 120 μM 2,2′-dipyridyl. Tested strains are those tested by placing a loopful of bacterial biomass on the agar surface. A growth halo indicates that indicator strains can use the siderophores produced by tested strains to overcome the iron limitation conditions. Photobacterium damselae subsp. piscicida DI21 wild type strain (irp+, piscibactin producer) and DI21 cured from plasmid pPHDP70 (irp-, unable to produce piscibactin) were used as controls. RV22ΔvabF does not produce vanchrobactin; RV22ΔvabD does not produce vanchrobactin nor piscibactin.
Piscibactin Synthesis Contributes to the Growth Ability Under Iron Restriction Depending of Environmental Conditions
To test whether irp1 is required for piscibactin production and to study the influence of piscibactin on V. anguillarum cell fitness, in-frame deletion mutants of irp1 were obtained in RV22wt and RV22ΔvabF (deficient in vanchrobactin synthesis) backgrounds. The growth ability of resultant RV22Δirp1 and RV22ΔvabFΔirp1 mutant strains was measured at 25°C under iron excess and under iron-deprivation conditions using the strong iron-chelating agent ethylenediamine-di-(o-hydroxyphenyl-acetic acid) (EDDA), or a weaker one, 2,2′-dipyridyl. RV22 wild type strain (that produces both vanchrobactin and piscibactin) was used as positive growth control showing a minimal inhibitory concentration (MIC) of EDDA and 2,2′-dipyridyl of 10 μM and 150 μM, respectively. Conversely, RV22ΔvabD strain (Balado et al., 2008), impaired to produce any of these siderophores, showed MICs of 5 μM for EDDA and 100 μM for 2,2′-dipyridyl. When the mutants were cultured under iron-excess conditions (CM9 plus 10 μM ferric chloride) no significant differences in growth levels were observed with respect to the wild type strain (Figure 5A). However, significant differences were observed between RV22wt and some of the siderophore mutants when cultivated under iron deficiency. When EDDA (CM9 plus 5 μM EDDA) was used as chelating agent, the RV22Δirp1 mutant (producing only vanchrobactin) and the parental strain showed identical growth levels. However, the mutant producing only piscibactin (RV22ΔvabF) was unable to grow (Figure 5A). These results corroborate those previously published (Balado et al., 2006) and confirm that inactivation of vanchrobactin synthesis strongly reduce the growth ability of V. anguillarum under in vitro iron starvation. Interestingly, when growth was measured in CM9 containing 100 μM of the chelator 2,2′-dipyridyl the mutant RV22ΔvabF reached around 50% of the growth levels showed by the parental strain (Figure 5A). Siderophore-like activity present in cell free supernatants, measured by the CAS assay after growing each V. anguillarum strain at 50 μM 2,2′-dipyridyl, correlated with the growth levels observed.
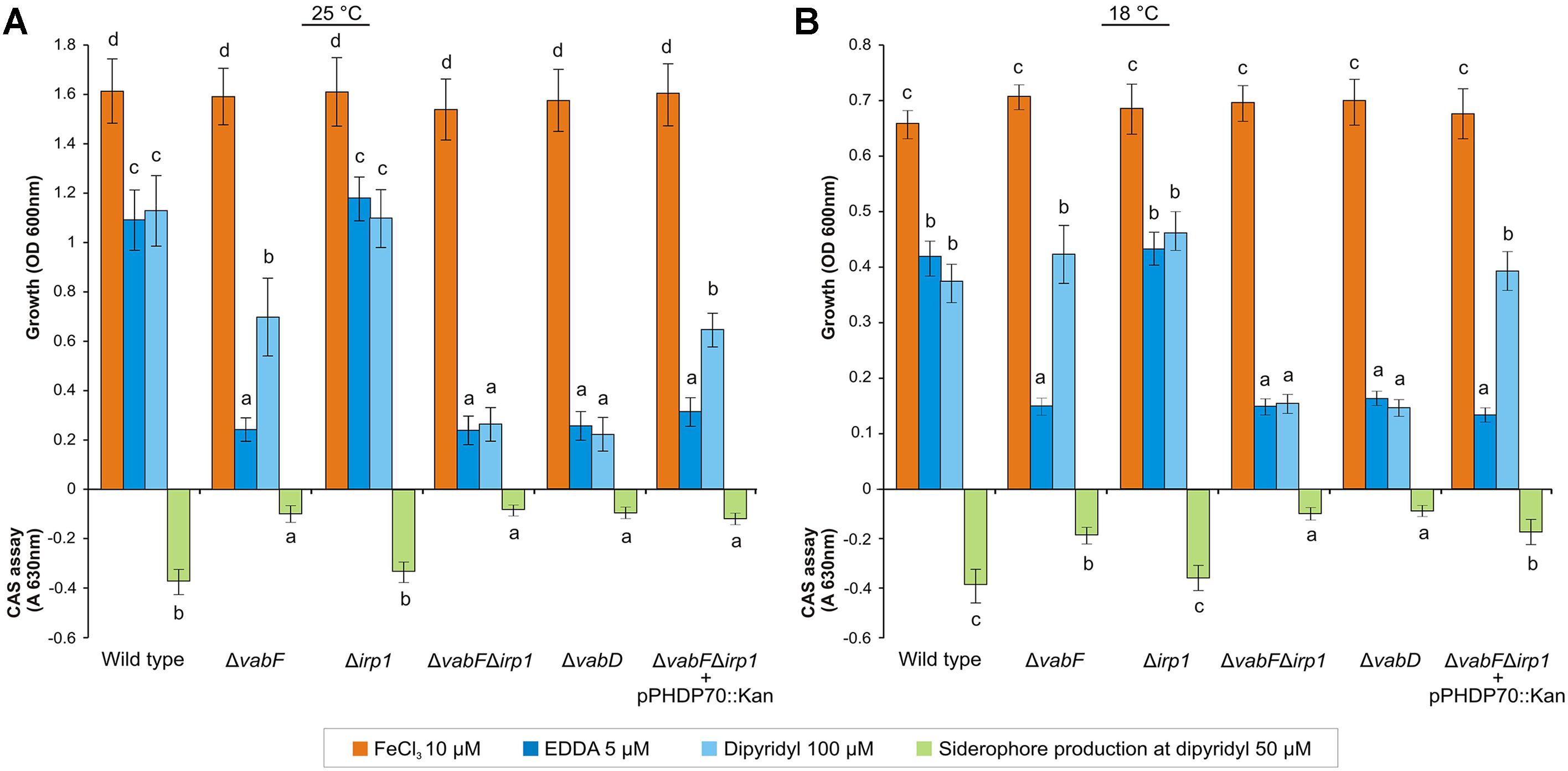
FIGURE 5. Growth of V. anguillarum strains cultivated for 24 h under iron excess or iron starvation in CM9 medium and incubated at 25°C (A) or 18°C (B). Siderophore production (indicated as A630 values), determined by the CAS test performed on culture supernatants, is also displayed for all strains. In each panel, bars labeled with the same letter (a, b, c, d) are not significantly different from each other (t-test). Error bars denote standard deviations.
The same growth experiments were conducted also at 18°C since it is a temperature closer to the conditions encountered in vivo during infection. At this temperature of incubation, and in presence of 100 μM 2,2′-dipyridyl, RV22ΔvabF (producing only piscibactin) behaved similarly to the wild type strain (Figure 5B), which suggests that piscibactin production by V. anguillarum could be favored at low temperatures (see below). Reactivation of piscibactin synthesis in RV22ΔvabFΔirp1 double mutant by introduction of pPHDP70 plasmid (Osorio et al., 2015), resulting in strain RV22ΔvabFΔirp1(pPHDP70), showed a phenotype almost identical to RV22ΔvabF strain in both temperatures assayed (Figures 5A,B).
Overall, these results suggest that (i) V. anguillarum may use vanchrobactin as primary siderophore and that (ii) piscibactin efficacy to provide iron to support growth depends on the environmental conditions, in particular on the iron source and the temperature. Additionally, the results also suggest that the role of vanchrobactin in the biology of V. anguillarum could be previously overestimated (Balado et al., 2006) probably owing to the fact that vanchrobactin defective mutants have been studied only under stringent conditions of iron limitation by the use of EDDA as iron chelator.
Piscibactin Contributes to a Greater Extent Than Vanchrobactin to Virulence for Turbot
In order to study the contribution of each siderophore (vanchrobactin and piscibactin) to V. anguillarum pathogenesis, the virulence of the RV22 wild type strain, as well as of the mutants deficient in one of the two siderophores or in both of them, was evaluated by experimental infections in turbot. The fish were kept at 18–20°C and were inoculated intraperitoneally with 2–4 × 104 CFU of the wild type or with one of the single or double siderophore mutants. The wild type strain RV22 killed 80% of the fish after 5 days of challenge (Figure 6). When the challenge was done with the mutant that produces only vanchrobactin (RV22Δirp1) the mortality reached 56% after 6 days of infection. However, when the fish were inoculated with the mutant that produces only piscibactin (RV22ΔvabF) the mortality reached 88% after 5 days, at the same level of the wild type strain. The double mutant RV22ΔvabFΔirp1 and the RV22ΔvabD (both unable to produce any of the two siderophores) give mortalities of 12% and 8%, respectively, after 7 days. Interestingly, when piscibactin production was reactivated by the introduction of pPHDP70 plasmid (that contains the piscibactin synthesis gene cluster) (Osorio et al., 2015) into RV22ΔvabFΔirp1 double mutant, this complemented strain caused a mortality of 78% after 6 days (Figure 6). These results clearly demonstrate that the ability to synthesize a siderophore has a great impact in the virulence of V. anguillarum. However, each siderophore system contributes in different grade to virulence being piscibactin production, more than vanchrobactin, the main virulence factor in the tested conditions.
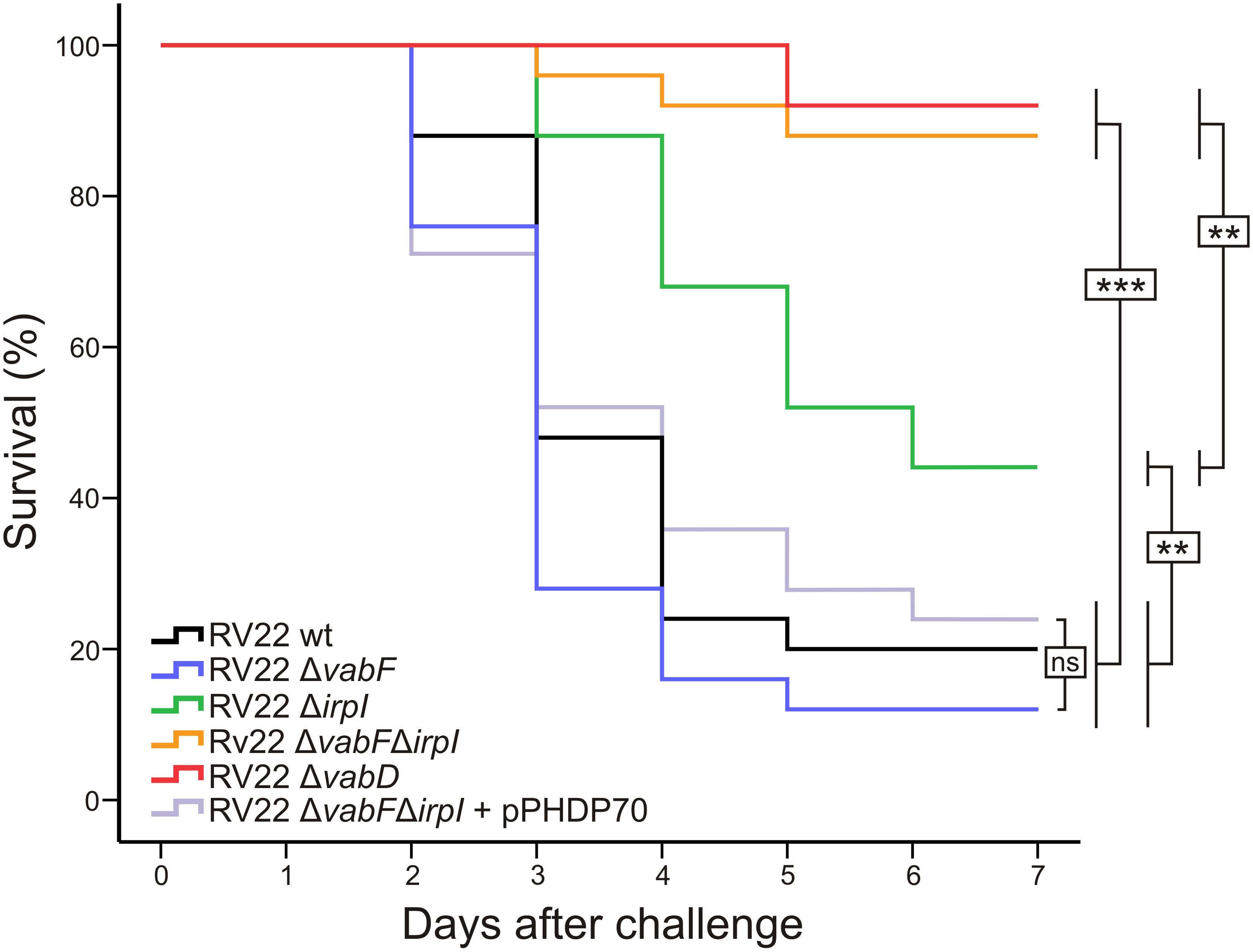
FIGURE 6. Percentage survival after 7 days of turbot fingerlings challenged with 2–4 × 104 CFU/fish of V. anguillarum RV22 wild type strain or with the siderophore mutants analyzed. Asterisks denote statistically significant differences between strains: ∗P < 0.05; ∗∗P < 0.01; ∗∗∗P < 0.001; ns, no statistically significant differences.
Transcriptional Analysis of the irpang Gene Cluster: The Expression of Piscibactin Genes Are Favored at Low Temperatures
The synthesis of siderophores are tightly regulated at transcriptional level which ensures that siderophore production occurs only when it is necessary (Cornelis et al., 2011). In order to analyze the expression levels of the irpang putative promoters, DNA fragments of ca. 700 nucleotides upstream frpAang, araC1ang and araC2ang genes, that were named PfrpA ParaC1 and ParaC2, respectively (Figure 1B), were cloned into the plasmid pHRP309 upstream of a promoterless lacZ gene. frpA encodes the presumptive ferri-piscibactin outer membrane receptor while araC1 and araC2 encode two putative AraC-type transcriptional regulators (Supplementary Table S1). Resulting plasmids were mobilized into V. anguillarum RV22wt and the transcription levels were measured by determining β-galactosidase activities under different iron availability conditions at 25°C (Figure 7A). The expression levels were compared with the promoter of the vanchrobactin system located upstream of vabH (Balado et al., 2008). Interestingly, the use of the PfrpA, ParaC1, and ParaC2 presumptive promoters produced significant β-galactosidase activity (Figure 7) which demonstrates that the three sequences serve as transcriptional starts. In addition, under the iron-restricted conditions tested, the expression from promoter PfrpA was three-times higher than from ParaC1 or ParaC2 (Figure 7A). Thus, although the transcription of piscibactin genes could start from any of the three promoters, it seems to occur to a greater extent from the promoter immediately upstream of frpA (PfrpA), which encodes the probable piscibactin receptor.
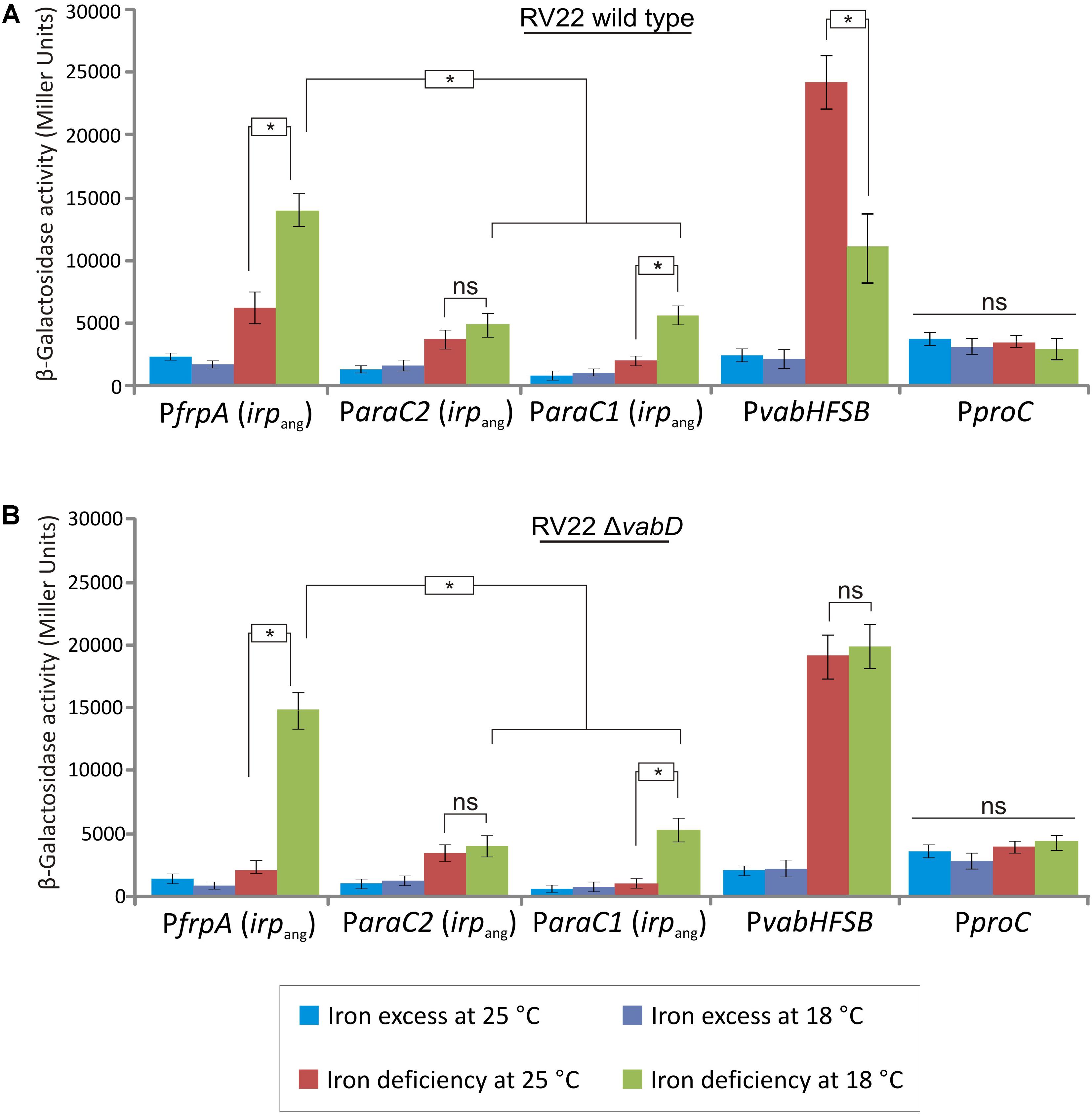
FIGURE 7. Transcriptional activity (β-galactosidase units) of the lacZ fusions to the P1, P2 and vabH promoters in V. anguillarum RV22wt (A) or RV22ΔvabD mutant impaired to produce siderophores (B) grown at 25°C or 18°C in CM9 medium under different iron availability conditions: iron excess and CM9 containing 50 μM 2,2′-dipyridyl. lacZ fusion with the constitutive promoter of proC (PproC) was used as control. Asterisks indicate statistically significant differences between bars (P < 0.05); ns, no statistically significant differences. Error bars denote standard deviations.
Usually, expression of siderophore systems promoters are down-regulated under iron availability (Cornelis et al., 2011). As expected, the addition of an iron chelator to the medium produced a significant increase of PfrpA, ParaC1, and ParaC2 activities (Figure 7A). However, the maximal expression levels achieved by piscibactin promoters under high iron deprivation (by addition of 2,2′-dipyridyl 100 μM) at 25°C were fourfold lower with respect to the vanchrobactin promoter (Figure 7A). These findings strongly contrast with the high relevance of piscibactin in virulence, as demonstrated above (Figure 6). This low expression at 25°C could be due to: (1) irpang genes could remain down-regulated because enough iron is entering the cell through the vanchrobactin system; or (2) piscibactin promoters need additional environmental signals, not only low iron levels, to achieve their maximal activity. These two hypothesis need to be further studied.
Since, as shown above, temperature could have a relevant role in piscibactin production, we wanted to ascertain if the expression levels of piscibactin genes could be favored at low temperatures. For this purpose, transcriptional levels of the piscibactin promoters were measured in RV22wt strain incubated at 18°C and compared to those observed at 25°C. While ParaC2 showed almost the same activity at both temperatures, we found an increment of ca. twofold in the PfrpA and ParaC1 activity at 18°C with respect to that measured at 25°C (Figure 7A). In contrast, the activity of the vanchrobactin promoter decreased twofold. When constitutive proC-promoter was tested, no significant differences in lacZ activity were observed between conditions assayed either temperature or iron availability (Figures 7A,B). These findings show that expression of piscibactin genes are favored when V. anguillarum grows at low temperatures. In addition, the increase of piscibactin promoters expression, together with the decrease of vanchrobactin promoters at low temperatures, strongly suggests that V. anguillarum is able to balance the synthesis of each siderophore responding to environmental signals like iron levels and temperature (Figure 7). To ascertain if the decrease in vancrobactin promoter activity at 18°C was due to the temperature or if it was caused by the intake of iron mediated by piscibactin, the expression levels of the three promoters were measured in the RV22ΔvabD strain, which is unable to produce any siderophore. While PfrpA and ParaC1 piscibactin promoters showed a strong temperature-dependent expression pattern again, the vanchrobactin promoter (PvabH) showed almost identical β-galactosidase activity at 25 or 18°C (Figure 7B). These results suggest that the lower activity of vanchrobactin promoter at 18°C in the RV22wt strain may be due to the increased intake of iron through the piscibactin system more than to a temperature-dependent regulation. Overall, these results strongly suggest that the expression of irpang genes in V. anguillarum are favored at low growth temperatures.
Discussion
In the present work, we could identify in V. anguillarum a siderophore gene cluster, named irpang, that is contained within a pathogenicity island and that shows a strong homology to its counterpart described in the pPHDP70 plasmid from P. damselae subsp. piscicida (Osorio et al., 2015). This PAI is located in the second chromosome of some highly pathogenic strains of V. anguillarum, including strain RV22, from which vanchrobactin genes were previously identified (Balado et al., 2006, 2008). The irpang cluster shows the same genetic organization than the irp genes of P. damselae subsp. piscicida and both confer the ability to produce the siderophore piscibactin. The PAI island harboring the piscibactin genes in P. damselae subsp. piscicida encodes all functions required to produce and use piscibactin (Osorio et al., 2015). However, piscibactin synthesis in V. anguillarum depends on the presence in the recipient genome of a functional 4′-phosphopantetheinyl transferase gene (entD). A similar case was reported for Aeromonas salmonicida, where acinetobactin and amonabactin biosynthetic pathways share the same set of genes encoding the biosynthesis of 2,3-DHBA, a common moiety of both siderophores, and also share an EntD homolog (Balado et al., 2015).
Chemical analysis demonstrated that piscibactin is indeed being synthesized by the vanchrobactin-producing V. anguillarum strains. Although the production of more than one siderophore by a bacterial cell could be deleterious (Cordero et al., 2012), in some cases the ability to produce two siderophores can enhance niche flexibility (Sandy et al., 2010; Dumas et al., 2013) and pathogenesis (Garcia et al., 2011). Some pathogens carry multiple iron acquisition systems that seem redundant in laboratory culture conditions. For instance, the fish pathogen Aeromonas salmonicida subsp. salmonicida produces acinetobactin and amonabactin, two catechol siderophores and, although there are strains that produce both siderophores simultaneously, most strains have the amonabactin cluster inactivated (Balado et al., 2015). By contrast, the co-expression of siderophores with different chemical properties can play specialized roles at the host-pathogen interface (Fetherston et al., 2010; Koh and Henderson, 2015). Piscibactin, along with yersiniabactin and pyochelin, constitute a group of siderophores that share similar structural characteristics (Crosa and Walsh, 2002; Souto et al., 2012). It has been shown that the co-expression of yersiniabactin with other catecholate siderophores enhances the virulence properties of uropathogenic Escherichia coli (Garcia et al., 2011). Yersiniabactin and pyochelin seem to play several roles, other than iron uptake, during infection since they can efficiently bind other metals besides iron. For instance, yersiniabactin participates in zinc (Zn+2) acquisition and also confers protection against copper toxicity, which promotes bacterial colonization, dissemination and resistance against phagocytosis (Fetherston et al., 2010; Chaturvedi et al., 2012; Bobrov et al., 2014; Koh and Henderson, 2015; Holden et al., 2016). Pyochelin has been also associated with some traits such as oxidative or inflammatory activities in Pseudomonas aeruginosa infections (Britigan et al., 1997), decomposition of organotin compounds (like TPT, triphenyltin chloride or DPT, diphenyltin dichloride) (Sun et al., 2006) or plants defense against fungal plagues (Audenaert et al., 2002). Some of these or other related roles could also be attributed to piscibactin in V. anguillarum.
Interestingly, the inactivation of piscibactin synthesis in V. anguillarum RV22 produced just a small decrease in the growth ability under iron restriction. This suggests that these mutants would use vanchrobactin to internalize iron. Thus, vanchrobactin would serve in vitro as an efficient iron carrier (Iglesias et al., 2011) for V. anguillarum while the piscibactin efficacy to supply iron to the cell would depend on which is the iron source available (Figure 5). However, this observation highly contrasts with our results of the virulence assays where piscibactin production alone was sufficient to obtain the maximal virulence. The inactivation of piscibactin system resulted in a severe loss of the virulence degree even if vanchrobactin system was functional (Figure 6). Hence, although the ability to synthesize one siderophore, either vanchrobactin or piscibactin, has a great impact in V. anguillarum virulence, the ability to produce piscibactin seems to be sufficient to confer maximal virulence to V. anguillarum, at least in the tested conditions.
A total of 43 V. anguillarum genomes are currently available in the genomic databases and the virulence properties of 32 of these strains were recently reported (Busschaert et al., 2015; Rønneseth et al., 2017). An in silico analysis showed that many of these virulent V. anguillarum strains harbor both, vanchrobactin and piscibactin, gene clusters (Table 3). Thus, piscibactin genes are widespread in V. anguillarum strains and they are associated with a highly virulent phenotype for several fish species, such as turbot (Psetta maxima), cod (Gadus morhua), halibut (Hippoglossus hippoglossus), rainbow trout (Oncorhynchus mykiss) or sea bass (Dicentrarchus labrax) (Busschaert et al., 2015; Rønneseth et al., 2017). These results support the previously suggested hypothesis that vanchrobactin is the ancient siderophore of V. anguillarum (Lemos et al., 2010). However, according to our results and to previously published works (Sandy et al., 2010), vanchrobactin production could be more related to the persistence of V. anguillarum in the marine environment than to pathogenesis. Hence, piscibactin would be the most important siderophore during the vibriosis outbreaks in fish. Interestingly, it was recently found that piscibactin is one of the most extended siderophore systems in Vibrionaceae family. Phylogenetic analysis of three siderophore systems (piscibactin, vibrioferrin, and aerobactin) suggested that their current distribution could be explained by an old insertion that was followed by the action of diverse evolutionary forces (Thode et al., 2018). An interesting observation is that the piscibactin gene cluster is never present in anguibactin-producing strains (Table 3). It has been previously reported that the presence of pJM1-like plasmids encoding the anguibactin system inactivates the vanchrobactin cluster through a transposition event (Naka et al., 2008). A similar genetic incompatibility could explain the mutual exclusion between piscibactin and anguibactin gene clusters.
Many of the genes encoding virulence factors, including iron uptake systems, are preferentially expressed within the host in response to particular environmental signals (Lee and Camilli, 2000). The analysis of promoters activity showed that irpang genes in V. anguillarum are not only strongly iron-regulated, but also, that piscibactin production has a requirement for low temperatures. These findings suggest that V. anguillarum is able to modulate the synthesis of vanchrobactin and piscibactin according to the surrounding temperature, which adjust the energy costs of siderophore production to the environmental circumstances. Some bacterial species that are able to produce more than one siderophore can modulate the expression as a response to environmental factors, e.g., pH, carbon source, amino sugars or temperature (Valdebenito et al., 2006; Craig et al., 2012; Cornelis and Dingemans, 2013; Dumas et al., 2013; Tanabe et al., 2014). The temperature-dependent regulation is characteristic of pathogenic bacteria affecting ectothermic hosts such as fish, where the infection processes generally occur at temperatures lower than those required for bacterial optimal growth (Guijarro et al., 2015). However, these regulation mechanisms are yet poorly understood and few examples were reported of siderophore synthesis regulated by temperature. One of these is ruckerbactin, a catecholate siderophore produced by Yersinia ruckeri (Fernández et al., 2004); other one is the hydroxamate bisucaberin produced by Aliivibrio salmonicida, the causative agent of cold-water vibriosis (Colquhoun and Sørum, 2001; Winkelmann et al., 2002). In both bacteria, the maximal siderophore production take place around the temperatures at which the diseases outbreaks occur. Our results also suggest that piscibactin synthesis in V. anguillarum is higher at temperatures (16–19°C) at which the vibriosis outbreaks usually occur in turbot (Toranzo et al., 2005, 2017).
Conclusion
Pathogenic V. anguillarum strains lacking the anguibactin system produce two chromosomally encoded siderophores, vanchrobactin and piscibactin. While vanchrobactin synthesis seems to be regulated only by the iron availability, piscibactin is preferentially produced at low temperatures at which fish vibriosis outbreaks usually occur in natural conditions. Piscibactin seems to contribute to a greater extent than vanchrobactin to the virulence for turbot. Further studies are currently under way to decipher the regulation pathways that control piscibactin synthesis in V. anguillarum.
Author Contributions
MB, JR, CJ, and MLL contributed to the conception and design of the study. MB, MAL, JF-M, and DM-M performed the lab experiments. MB, CJ, and MLL analyzed the data. MB wrote the first draft of the manuscript. CJ, JR, and MLL corrected the draft and build the final version of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
This work was supported by grants AGL2015-63740-C2-1-R and AGL2015-63740-C2-2-R (AEI/FEDER, EU) from the State Agency for Research (AEI) of Spain, and co-funded by the FEDER Programme from the European Union. The support of Xunta de Galicia (Spain) with grant GRC-2014/007 is also acknowledged.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01766/full#supplementary-material
Footnotes
References
Actis, L. A., Fish, W., Crosa, J. H., Kellerman, K., Ellenberger, S. R., Hauser, F. M., et al. (1986). Characterization of anguibactin, a novel siderophore from Vibrio anguillarum 775(pJM1). J. Bacteriol. 167, 57–65. doi: 10.1128/jb.167.1.57-65.1986
Audenaert, K., Pattery, T., Cornelis, P., and Höfte, M. (2002). Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant Microbe Interact. 15, 1147–1156. doi: 10.1094/MPMI.2002.15.11.1147
Balado, M., Osorio, C. R., and Lemos, M. L. (2006). A gene cluster involved in the biosynthesis of vanchrobactin, a chromosome-encoded siderophore produced by Vibrio anguillarum. Microbiology 152, 3517–3528. doi: 10.1099/mic.0.29298-0
Balado, M., Osorio, C. R., and Lemos, M. L. (2008). Biosynthetic and regulatory elements involved in the production of the siderophore vanchrobactin in Vibrio anguillarum. Microbiology 154, 1400–1413. doi: 10.1099/mic.0.2008/016618-0
Balado, M., Souto, A., Vences, A., Careaga, V. P., Valderrama, K., Segade, Y., et al. (2015). Two catechol siderophores, acinetobactin and amonabactin, are simultaneously produced by Aeromonas salmonicida subsp. salmonicida sharing part of the biosynthetic pathway. ACS Chem. Biol. 10, 2850–2860. doi: 10.1021/acschembio.5b00624
Bobrov, A. G., Kirillina, O., Fetherston, J. D., Miller, M. C., Burlison, J. A., and Perry, R. D. (2014). The Yersinia pestis siderophore, yersiniabactin, and the ZnuABC system both contribute to zinc acquisition and the development of lethal septicaemic plague in mice. Mol. Microbiol. 93, 759–775. doi: 10.1111/mmi.12693
Britigan, B. E., Rasmussen, G. T., and Cox, C. D. (1997). Augmentation of oxidant injury to human pulmonary epithelial cells by the Pseudomonas aeruginosa siderophore pyochelin. Infect. Immun. 65, 1071–1076.
Busschaert, P., Frans, I., Crauwels, S., Zhu, B., Willems, K., Bossier, P., et al. (2015). Comparative genome sequencing to assess the genetic diversity and virulence attributes of 15 Vibrio anguillarum isolates. J. Fish Dis. 38, 795–807. doi: 10.1111/jfd.12290
Castillo, D., Alvise, P. D., Xu, R., Zhang, F., Middelboe, M., and Gram, L. (2017). Comparative genome analyses of Vibrio anguillarum strains reveal a link with pathogenicity traits. mSystems 2:e00001-17. doi: 10.1128/mSystems.00001-17
Chaturvedi, K. S., Hung, C. S., Crowley, J. R., Stapleton, A. E., and Henderson, J. P. (2012). The siderophore yersiniabactin binds copper to protect pathogens during infection. Nat. Chem. Biol. 8, 731–736. doi: 10.1038/nchembio.1020
Colquhoun, D. J., and Sørum, H. (2001). Temperature dependent siderophore production in Vibrio salmonicida. Microb. Pathog. 31, 213–219. doi: 10.1006/mpat.2001.0464
Cordero, O. X., Ventouras, L.-A., DeLong, E. F., and Polz, M. F. (2012). Public good dynamics drive evolution of iron acquisition strategies in natural bacterioplankton populations. Proc. Natl. Acad. Sci. U.S.A. 109, 20059–20064. doi: 10.1073/pnas.1213344109
Cornelis, P., and Dingemans, J. (2013). Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front. Cell. Infect. Microbiol. 3:75. doi: 10.3389/fcimb.2013.00075
Cornelis, P., Wei, Q., Andrews, S. C., and Vinckx, T. (2011). Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 3, 540–549. doi: 10.1039/c1mt00022e
Craig, M., Lambert, S., Jourdan, S., Tenconi, E., Colson, S., Maciejewska, M., et al. (2012). Unsuspected control of siderophore production by N-acetylglucosamine in streptomycetes. Environ. Microbiol. Rep. 4, 512–521. doi: 10.1111/j.1758-2229.2012.00354.x
Crosa, J. H., and Walsh, C. T. (2002). Genetics and assembly line enzymology of siderophore biosynthesis in bacteria. Microbiol. Mol. Biol. Rev. 66, 223–249. doi: 10.1128/MMBR.66.2.223-249.2002
Croxatto, A., Lauritz, J., Chen, C., and Milton, D. L. (2007). Vibrio anguillarum colonization of rainbow trout integument requires a DNA locus involved in exopolysaccharide transport and biosynthesis. Environ. Microbiol. 9, 370–382. doi: 10.1111/j.1462-2920.2006.01147.x
Darling, A. E., Mau, B., and Perna, N. T. (2010). progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One 5:e11147. doi: 10.1371/journal.pone.0011147
Denkin, S. M., and Nelson, D. R. (2004). Regulation of Vibrio anguillarum empA metalloprotease expression and its role in virulence. Appl. Environ. Microbiol. 70, 4193–4204. doi: 10.1128/AEM.70.7.4193-4204.2004
Dumas, Z., Ross-Gillespie, A., and Kümmerli, R. (2013). Switching between apparently redundant iron-uptake mechanisms benefits bacteria in changeable environments. Proc. Biol. Sci. 280:20131055. doi: 10.1098/rspb.2013.1055
Espada, A., Anta, C., Bragado, A., Rodríguez, J., and Jiménez, C. (2011). An approach to speed up the isolation of hydrophilic metabolites from natural sources at semipreparative level by using a hydrophilic-lipophilic balance/mixed-mode strong cation exchange-high-performance liquid chromatography/mass spectrometry system. J. Chromatogr. A 1218, 1790–1794. doi: 10.1016/j.chroma.2011.01.072
Fernández, L., Márquez, I., and Guijarro, J. A. (2004). Identification of specific in vivo-induced (ivi) genes in Yersinia ruckeri and analysis of ruckerbactin, a catecholate siderophore iron acquisition system. Appl. Environ. Microbiol. 70, 5199–5207. doi: 10.1128/AEM.70.9.5199-5207.2004
Fetherston, J. D., Kirillina, O., Bobrov, A. G., Paulley, J. T., and Perry, R. D. (2010). The yersiniabactin transport system is critical for the pathogenesis of bubonic and pneumonic plague. Infect. Immun. 78, 2045–2052. doi: 10.1128/IAI.01236-09
Frans, I., Michiels, C. W., Bossier, P., Willems, K. A., Lievens, B., and Rediers, H. (2011). Vibrio anguillarum as a fish pathogen: virulence factors, diagnosis and prevention. J. Fish Dis. 34, 643–661. doi: 10.1111/j.1365-2761.2011.01279.x
Garcia, E. C., Brumbaugh, A. R., and Mobley, H. L. T. (2011). Redundancy and specificity of Escherichia coli iron acquisition systems during urinary tract infection. Infect. Immun. 79, 1225–1235. doi: 10.1128/IAI.01222-10
Guijarro, J. A., Cascales, D., García-Torrico, A. I., García-Domínguez, M., and Méndez, J. (2015). Temperature-dependent expression of virulence genes in fish-pathogenic bacteria. Front. Microbiol. 6:700. doi: 10.3389/fmicb.2015.00700
Herrero, M., de Lorenzo, V., and Timmis, K. N. (1990). Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram-negative bacteria. J. Bacteriol. 172, 6557–6567. doi: 10.1128/jb.172.11.6557-6567.1990
Hickey, M. E., and Lee, J. L. (2017). A comprehensive review of Vibrio (Listonella) anguillarum: ecology, pathology and prevention. Rev. Aquac. 1893, 1–26. doi: 10.1111/raq.12188
Holden, V. I., Breen, P., Houle, S., Dozois, C. M., and Bachman, M. A. (2016). Klebsiella pneumoniae siderophores induce inflammation, bacterial dissemination, and HIF-1α stabilization during pneumonia. mBio 7:e01397-16. doi: 10.1128/mBio.01397-16
Holm, K. O., Bækkedal, C., Söderberg, J. J., and Haugen, P. (2018). Complete genome sequences of seven Vibrio anguillarum strains as derived from PacBio sequencing. Genome Biol. Evol. 10, 1127–1131. doi: 10.1093/gbe/evy074
Iglesias, E., Brandariz, I., Jiménez, C., and Soengas, R. G. (2011). Iron(III) complexation by Vanchrobactin, a siderophore of the bacterial fish pathogen Vibrio anguillarum. Metallomics 3, 521–528. doi: 10.1039/c0mt00071j
Koh, E.-I., and Henderson, J. P. (2015). Microbial copper-binding siderophores at the host-pathogen interface. J. Biol. Chem. 290, 18967–18974. doi: 10.1074/jbc.R115.644328
Lee, S. H., and Camilli, A. (2000). Novel approaches to monitor bacterial gene expression in infected tissue and host. Curr. Opin. Microbiol. 3, 97–101. doi: 10.1016/S1369-5274(99)00058-2
Lemos, M. L., Balado, M., and Osorio, C. R. (2010). Anguibactin- versus vanchrobactin-mediated iron uptake in Vibrio anguillarum: evolution and ecology of a fish pathogen. Environ. Microbiol. Rep. 2, 19–26. doi: 10.1111/j.1758-2229.2009.00103.x
Lemos, M. L., Salinas, P., Toranzo, A. E., Barja, J. L., and Crosa, J. H. (1988). Chromosome-mediated iron uptake system in pathogenic strains of Vibrio anguillarum. J. Bacteriol. 170, 1920–1925. doi: 10.1128/jb.170.4.1920-1925.1988
Li, L., Rock, J. L., and Nelson, D. R. (2008). Identification and characterization of a repeat-in-toxin gene cluster in Vibrio anguillarum. Infect. Immun. 76, 2620–2632. doi: 10.1128/IAI.01308-07
Li, Y., and Ma, Q. (2017). Iron acquisition strategies of Vibrio anguillarum. Front. Cell. Infect. Microbiol. 7:342. doi: 10.3389/fcimb.2017.00342
Miller, J. F., Mekalanos, J. J., and Falkow, S. (1989). Coordinate regulation transduction in the sensory of control and bacterial virulence. Scienc 243, 916–922. doi: 10.1126/science.2537530
Miller, J. H. (1992). A Short Course in Bacterial Genetics. Plainview, NY: Cold Spring Harbor Laboratory Press.
Mourino, S., Osorio, C. R., and Lemos, M. L. (2004). Characterization of heme uptake cluster genes in the fish pathogen Vibrio anguillarum. J. Bacteriol. 186, 6159–6167. doi: 10.1128/JB.186.18.6159-6167.2004
Münch, R., Hiller, K., Barg, H., Heldt, D., Linz, S., Wingender, E., et al. (2003). PRODORIC: prokaryotic database of gene regulation. Nucleic Acids Res. 31, 266–269. doi: 10.1093/nar/gkg037
Naka, H., Dias, G. M., Thompson, C. C., Dubay, C., Thompson, F. L., and Crosa, J. H. (2011). Complete genome sequence of the marine fish pathogen Vibrio anguillarum harboring the pJM1 virulence plasmid and genomic comparison with other virulent strains of V. anguillarum and V. ordalii. Infect. Immun. 79, 2889–2900. doi: 10.1128/IAI.05138-11
Naka, H., Lopez, C. S., and Crosa, J. H. (2008). Reactivation of the vanchrobactin siderophore system of Vibrio anguillarum by removal of a chromosomal insertion sequence originated in plasmid pJM1 encoding the anguibactin siderophore system. Environ. Microbiol. 10, 265–277.
Okada, N., Iida, T., Park, K.-S., Goto, N., Yasunaga, T., Hiyoshi, H., et al. (2009). Identification and characterization of a novel type III secretion system in trh-positive Vibrio parahaemolyticus strain TH3996 reveal genetic lineage and diversity of pathogenic machinery beyond the species level. Infect. Immun. 77, 904–913. doi: 10.1128/IAI.01184-08
Ormonde, P., Horstedt, P., O’Toole, R., and Milton, D. L. (2000). Role of motility in adherence to and invasion of a fish cell line by Vibrio anguillarum. J. Bacteriol. 182, 2326–2328. doi: 10.1128/JB.182.8.2326-2328.2000
Osorio, C. R., Juiz-Rio, S., and Lemos, M. L. (2006). A siderophore biosynthesis gene cluster from the fish pathogen Photobacterium damselae subsp. piscicida is structurally and functionally related to the Yersinia high-pathogenicity island. Microbiology 152, 3327–3341. doi: 10.1099/mic.0.29190-0
Osorio, C. R., Rivas, A. J., Balado, M., Fuentes-Monteverde, J. C., Rodríguez, J., Jiménez, C., et al. (2015). A transmissible plasmid-borne pathogenicity island confers piscibactin biosynthesis in the fish pathogen Photobacterium damselae subsp. piscicida. Appl. Environ. Microbiol. 81, 5867–5879. doi: 10.1128/AEM.01580-15
O’Toole, R., Milton, D. L., and Wolf-Watz, H. (1996). Chemotactic motility is required for invasion of the host by the fish pathogen Vibrio anguillarum. Mol. Microbiol. 19, 625–637. doi: 10.1046/j.1365-2958.1996.412927.x
Parales, R. E., and Harwood, C. S. (1993). Construction and use of a new broad-host-range lacZ transcriptional fusion vector, pHRP309, for Gram- bacteria. Gene 133, 23–30. doi: 10.1016/0378-1119(93)90220-W
Pedersen, K., Grisez, L., van Houdt, R., Tiainen, T., Ollevier, F., and Larsen, J. L. (1999). Extended serotyping scheme for Vibrio anguillarum with the definition and characterization of seven provisional O-serogroups. Curr. Microbiol. 38, 183–189. doi: 10.1007/PL00006784
Rodkhum, C., Hirono, I., Crosa, J. H., and Aoki, T. (2005). Four novel hemolysin genes of Vibrio anguillarum and their virulence to rainbow trout. Microb. Pathog. 39, 109–119. doi: 10.1016/j.micpath.2005.06.004
Rønneseth, A., Castillo, D., D’Alvise, P., Tønnesen,Ø., Haugland, G., Grotkjaer, T., et al. (2017). Comparative assessment of Vibrio virulence in marine fish larvae. J. Fish Dis. 40, 1373–1385. doi: 10.1111/jfd.12612
Sandy, M., Han, A., Blunt, J., Munro, M., Haygood, M., and Butler, A. (2010). Vanchrobactin and anguibactin siderophores produced by Vibrio sp. DS40M4. J. Nat. Prod. 73, 1038–1043. doi: 10.1021/np900750g
Savioz, A., Jeenes, D., Kocher, H., and Haas, D. (1990). Comparison of proC and other housekeeping genes of Pseudomonas aeruginosa with their counterparts in Escherichia coli. Gene 86, 107–111. doi: 10.1016/0378-1119(90)90121-7
Schwyn, B., and Neilands, J. B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. doi: 10.1016/0003-2697(87)90612-9
Soengas, R. G., Anta, C., Espada, A., Nieto, R. M., Larrosa, M., Rodríguez, J., et al. (2007). Vanchrobactin: absolute configuration and total synthesis. Tetrahedron Lett. 48, 3021–3024. doi: 10.1016/j.tetlet.2007.02.125
Soengas, R. G., Anta, C., Espada, A., Paz, V., Ares, I. R., Balado, M., et al. (2006). Structural characterization of vanchrobactin, a new catechol siderophore produced by the fish pathogen Vibrio anguillarum serotype O2. Tetrahedron Lett. 47, 7113–7116. doi: 10.1016/j.tetlet.2006.07.104
Souto, A., Montaos, M. A., Rivas, A. J., Balado, M., Osorio, C. R., Rodríguez, J., et al. (2012). Structure and biosynthetic assembly of piscibactin, a siderophore from Photobacterium damselae subsp. piscicida, predicted from genome analysis. Eur. J. Org. Chem. 2012, 5693–5700. doi: 10.1002/ejoc.201200818
Sugiyama, T., Iida, T., Izutsu, K., Park, K.-S., and Honda, T. (2008). Precise region and the character of the pathogenicity island in clinical Vibrio parahaemolyticus strains. J. Bacteriol. 190, 1835–1837. doi: 10.1128/JB.01293-07
Sun, G. X., Zhou, W. Q., and Zhong, J. J. (2006). Organotin decomposition by pyochelin, secreted by Pseudomonas aeruginosa even in an iron-sufficient environment. Appl. Environ. Microbiol. 72, 6411–6413. doi: 10.1128/AEM.00957-06
Tanabe, T., Kato, A., Shiuchi, K., Miyamoto, K., Tsujibo, H., Maki, J., et al. (2014). Regulation of the expression of the Vibrio parahaemolyticus peuA gene encoding an alternative ferric enterobactin receptor. PLoS One 9:e105749. doi: 10.1371/journal.pone.0105749
Thode, S. K., Rojek, E., Kozlowski, M., Ahmad, R., and Haugen, P. (2018). Distribution of siderophore gene systems on a Vibrionaceae phylogeny: database searches, phylogenetic analyses and evolutionary perspectives. PLoS One 13:e0191860. doi: 10.1371/journal.pone.0191860
Toranzo, A. E., Barreiro, S., Casal, J. F., Figueras, A., Magariños, B., and Barja, J. L. (1991). Pasteurellosis in cultured gilthead seabream (Sparus aurata): first report in Spain. Aquaculture 99, 1–15. doi: 10.1016/0044-8486(91)90284-E
Toranzo, A. E., Magariños, B., and Avendaño-Herrera, R. (2017). “Vibriosis: Vibrio anguillarum, V. ordalii and Aliivibrio salmonicida,” in Fish Viruses and Bacteria: Pathobiology and Protection, eds P. T. K. Woo and R. C. Cipriano (Wallingford: CABI), 314–333. doi: 10.1079/9781780647784.0314
Toranzo, A. E., Magariños, B., and Romalde, J. L. (2005). A review of the main bacterial fish diseases in mariculture systems. Aquaculture 246, 37–61. doi: 10.1016/j.aquaculture.2005.01.002
Valdebenito, M., Crumbliss, A. L., Winkelmann, G., and Hantke, K. (2006). Environmental factors influence the production of enterobactin, salmochelin, aerobactin, and yersiniabactin in Escherichia coli strain Nissle 1917. Int. J. Med. Microbiol. 296, 513–520. doi: 10.1016/j.ijmm.2006.06.003
Wang, R. F., and Kushner, S. R. (1991). Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichia coli. Gene 100, 195–199. doi: 10.1016/0378-1119(91)90366-J
Wang, X. H., Oon, H. L., Ho, G. W., Wong, W. S., Lim, T. M., and Leung, K. Y. (1998). Internalization and cytotoxicity are important virulence mechanisms in Vibrio-fish epithelial cell interactions. Microbiology 144, 2987–3002. doi: 10.1099/00221287-144-11-2987
Winkelmann, G., Schmid, D. G., Nicholson, G., Jung, G., and Colquhoun, D. J. (2002). Bisucaberin–a dihydroxamate siderophore isolated from Vibrio salmonicida, an important pathogen of farmed Atlantic salmon (Salmo salar). Biometals 15, 153–160. doi: 10.1023/A:1015206419613
Keywords: Vibrio anguillarum, siderophores, vanchrobactin, piscibactin, bacterial virulence, fish pathogens
Citation: Balado M, Lages MA, Fuentes-Monteverde JC, Martínez-Matamoros D, Rodríguez J, Jiménez C and Lemos ML (2018) The Siderophore Piscibactin Is a Relevant Virulence Factor for Vibrio anguillarum Favored at Low Temperatures. Front. Microbiol. 9:1766. doi: 10.3389/fmicb.2018.01766
Received: 21 May 2018; Accepted: 16 July 2018;
Published: 02 August 2018.
Edited by:
Marie-Joelle Virolle, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Julian Ferreras, Instituto de Biología Subtropical (IBS), ArgentinaSébastien Rigali, University of Liège, Belgium
Rafi Ahmad, Inland Norway University of Applied Sciences, Norway
Hans Rediers, KU Leuven, Belgium
Alexandra Rebecca Mey, University of Texas at Austin, United States
Copyright © 2018 Balado, Lages, Fuentes-Monteverde, Martínez-Matamoros, Rodríguez, Jiménez and Lemos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel L. Lemos, manuel.lemos@usc.es