First observations of seasonal bottom water deoxygenation off the Gironde estuary (Bay of Biscay, North East Atlantic)
 Nicolas Dubosq1*
Nicolas Dubosq1*  Sabine Schmidt2
Sabine Schmidt2  Joël Sudre3
Joël Sudre3  Sylvain Rigaud4
Sylvain Rigaud4  Bastien Lamarque1 Martin Danilo1
Bastien Lamarque1 Martin Danilo1  Antoine Grémare1
Antoine Grémare1  Bruno Deflandre1
Bruno Deflandre1- 1Univ. Bordeaux, CNRS, Bordeaux INP, EPOC, UMR 5805, Pessac, France
- 2CNRS, Univ. Bordeaux, Bordeaux INP, EPOC, UMR 5805, Pessac, France
- 3CNRS, UAR CPST, La Seyne sur mer, France
- 4Université de Nîmes, EA 7352 CHROME, Nîmes, France
Although not systematically considered as oxygen minimum zones, River-dominated Ocean Margins are sensitive to oxygen depletions. For example, the continental shelf off the Gironde, which flows into the Bay of Biscay, had not yet been studied from this perspective although recent simulations suggested that this area should already have experienced deoxygenations in recent decades. To fill this gap, profiles of temperature, salinity and dissolved oxygen were performed in the water column of the continental shelf off the Gironde during seven cruises distributed over the different seasons between 2016 and 2021. Turbidity, chlorophyll-a and pH were also measured during some of these cruises. In winter, the water column was slightly stratified due to high river flows. Then, a seasonal thermal stratification was present from spring to autumn. Similarly, dissolved oxygen showed a seasonal dynamic with: a well-oxygenated water column in winter, an oxygen oversaturation in the first 20 meters during the spring bloom, and then a progressive oxygen depletion in bottom waters until reaching an oxygen saturation minimum down to 45% in autumn. These deoxygenations are explained by the seasonal stratification that isolates the bottom waters from spring to autumn, and are likely enhanced by the advection of deoxygenated waters from the north of the Bay of Biscay and the settling of the organic matter produced in surface waters. A better understanding of these processes in the context of global warming undoubtedly requires better documentation of dissolved oxygen variability through the implementation of a long-term and continuous in situ monitoring.
1 Introduction
Worldwide the number of oceanic areas affected by hypoxia, as well as the frequency and the spatial and temporal extents of this phenomenon, are increasing (Breitburg et al., 2018; Oschlies et al., 2018; Wakelin et al., 2020). This trend is more pronounced in coastal regions (Gilbert et al., 2010; Breitburg et al., 2018) and constitutes a rising threat for ecosystems. Indeed, many impacts (e.g. migrations, local extinctions, population and individual size decrease) up to food web perturbations have been observed (Breitburg et al., 2018; Levin, 2018; Fennel and Testa, 2019; Roman et al., 2019). Despite these potential effects on ecosystems supporting fisheries, the consequences of climate changes on dissolved oxygen (DO) in the coastal ocean are still poorly documented (Wakelin et al., 2020).
Although not systematically considered as Oxygen Minimum Zones, River-dominated Ocean Margins (RiOMar) are vulnerable to DO depletions due to riverine nutrient inputs. Indeed, they can promote excess phytoplankton production inducing a DO consumption for their degradation (Diaz and Rosenberg, 2008; Cai et al., 2011; Testa and Kemp, 2011; Breitburg et al., 2018; Wang et al., 2021). A recent simulation suggested that several areas of the North-West European continental shelves, including the Bay of Biscay, a wide inlet of the North Atlantic Ocean, would be vulnerable to deoxygenation (Ciavatta et al., 2016). The shelf off the Loire and Gironde, the main rivers flowing into the Bay of Biscay (Figure 1A), is estimated to experience relatively frequent deoxygenations over recent decades. Moreover, an ecological model applied to the French Atlantic shelf also simulated hypoxia south from the Gironde but the authors themselves considered this result “too severe and unrealistic” (Ménesguen et al., 2019). In fact, the only reported anoxia episode along the French shelf of the Bay of Biscay has been registered in July 1982 in its northernmost sector, off the smaller and eutrophicated Vilaine river (Figure 1A, Rossignol-Strick, 1985). Following this event, a high frequency monitoring of the Vilaine Bay revealed recurrent deoxygenations and even hypoxia in bottom waters (Retho et al., 2020; Ratmaya et al., 2022). The lack of such in situ data in the southern sector of the Bay of Biscay precludes confirming the occurrence of deoxygenation suggested by the models.
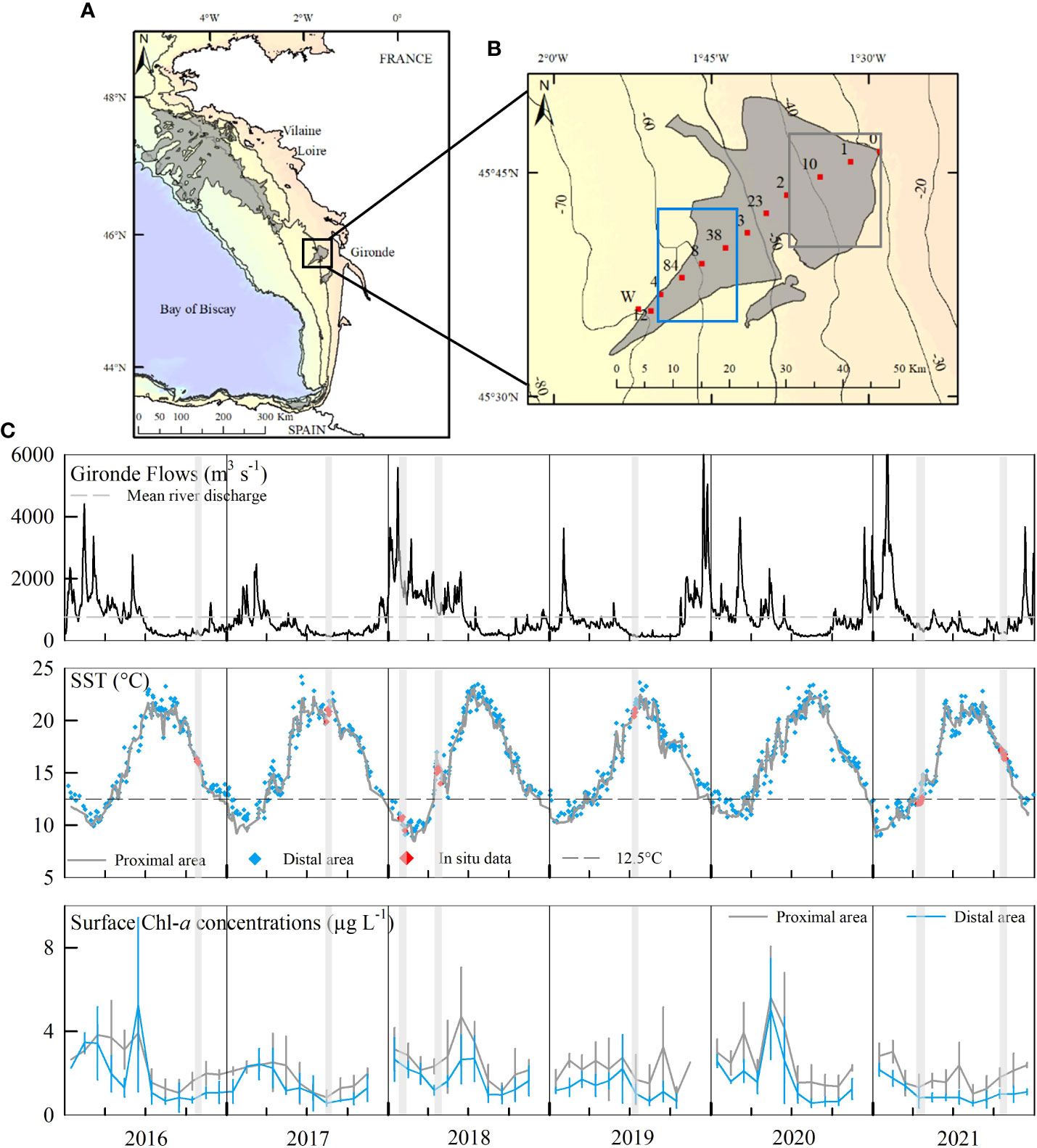
Figure 1 (A) The Bay of Biscay with the locations of mud patches (gray areas) and of the two main rivers, (B) map of the WGMP showing the locations of sampling stations (red squares) and the areas of satellite data extractions (rectangles). (C) Hydrological context: Gironde flows, Sea Surface Temperatures and monthly averages of surface Chl-a concentrations in the distal and proximal areas from the 01/01/2016 to the 12/31/2021. Gray lines indicate sampling periods.
The continental shelf off the Gironde is subjected to strong hydrodynamics, and is thus mainly composed of sands and gravels but also of several mud patches (Lesueur et al., 2002). Among them, the West Gironde Mud Patch (WGMP) is a well-defined sedimentary area located about 25 km off the Gironde and comprised between 30 and 75 m depth (Figure 1B). The WGMP is of particular interest due to its economic importance for common sole (Solea solea) and Norway lobster (Nephrops norvegicus) fishing but also for its biological activity (Lamarque et al., 2021) and its capability to store carbon (Dubosq et al., 2021). Its sedimentary functioning is studied since the 1990’s and its benthic ecological structuration was recently defined (Lesueur et al., 1991; Lesueur et al., 2002; Dubosq et al., 2021; Lamarque et al., 2021; Fontanier et al., 2022). However, there was still no information on DO in the WGMP water column. The present study aims to fill this gap by presenting the first measurements of DO and ancillary data (temperature, salinity, chlorophyll-a, turbidity, pH) in the water column along a cross-shelf transect covering the WGMP. This gives a first insight of the DO dynamics in this RiOMar, from seasonal to inter-annual scales, and its controlling factors.
2 Materials and methods
2.1 Field measurements
Seven cruises were performed from 2016 to 2021 on the R/V “Côtes de la Manche” to investigate the biogeochemical and ecological functioning of the benthic ecosystem of the WGMP (Deflandre, 2016; Deflandre et al., 2021). Even if these cruises were mainly dedicated to sediments, depth profiles of DO, temperature and salinity were performed in the water column at each sampling station (Figure 1B; Supplementary Table 1). Following the observation of unexpected low bottom DO concentrations during the first cruise, the strategy was completed from 2018 by a day per cruise (except in July 2019) dedicated to the achievement of cross-shelf transects from station 0 to W or 12 (Figure 1B).
Additional parameters were measured more occasionally: pH (October 2021), chlorophyll-a (October 2016, August 2017, January and April 2018, July 2019) and turbidity (January and April 2018, July 2019, October 2021). Depth profiles were acquired using inter-calibrated sensors: a CTD SeaBird SBE-19plus, autonomous sensors (STPS, SDOT, STBD, NKE Instrumentation) and a WiMo Plus multiparameter probe (NKE Instrumentation, October 2021). Profile resolutions varied from two to five measurements by meter depending on the response time of each sensor. Turbidity sensors were inter-calibrated using formazin, allowing to provide turbidity data in Formazin Nephelometric Units (FNU). Water density was calculated from temperature, salinity and depth using the seawater state equation (Pond and Pickard, 2007). The significance of the correlations between salinity above the halocline at each site and daily river flows and between DO concentrations and pH was assessed using a Spearman’s rank correlation coefficient (software SigmaPlot 14).
2.2 Satellite and gironde flows data
The NASA Aqua/MODIS data (4 km resolution, NASA Ocean Color, https://oceancolor.gsfc.nasa.gov/) were used to replace in situ measurements in the annual cycles of sea surface temperatures (SST) and biological production. SST and surface chlorophyll-a (Chl-a) concentrations were extracted for the proximal (longitudinal and latitudinal limits: 1°28.80’W to 1°37.50’W; 45°47.50’N to 45°40.00’N) and distal (1°42.50’W to 1°50.00’W; 45°42.50’N to 45°35.00’N) areas of the WGMP from the 01/01/2016 to the 12/31/2021 (Figure 1B). Each area consisted in 25 pixels whose values were averaged. For the entire period, 2189 pictures were available for SST and 2192 for surface Chl-a concentrations. However, some of them contained no or very little data due to cloud cover. Only pictures with at least 11 pixels per area were retained to calculate mean values. Outliers, with no data in the three previous or following days to confirm the value, were excluded. Monthly-averages of surface Chl-a concentrations were calculated to reduce the day-to-day variability and better identify trends.
The Gironde fluvial discharge corresponds to the sum of those of its two main tributaries, the Garonne and Dordogne rivers, whose measuring gauging stations are at Tonneins and Lamonzie Saint Martin, respectively. Data were extracted from the data portal hydro.eaufrance.fr.
3 Results
3.1 Gironde flow and satellite data
For the period 2016 to 2021, the daily Gironde flow ranged from 117 (August 2019) to 7900 m3 s-1 (February 2021) with a mean of 762 m3 s-1 (Figure 1C). It is slightly higher than the average over the period 2008 – 2021 (745 m3 s-1, hydro.eaufrance.fr). The lowest flows were recorded in summer and the highest in winter and spring. The intensity and duration of the high flow period presented a large inter-annual variability. For example, daily flows were above mean from January to June 2018, whereas it only happened for a few weeks in 2019. SST also showed a well-defined seasonal cycle (Figure 1C) with warmer waters in summer (>20°C) and colder in winter (<11°C). The inter-annual variability of SST was mainly reflected in a variation of the period during which extreme values were reached. For example, the lowest temperature was measured at the end of February in 2018 but in mid-January in 2021.
Surface Chl-a concentrations are an indicator of the net primary production. Monthly averages varied from 0.6 to 5.6 µg L-1 with higher values in the proximal than in the distal area (Figure 1C; Supplementary Table 2). They were at their highest in late winter and spring, and then decreased to a minimum in summer. A slight increase occurred in autumn. Interestingly, longer and more intense blooms periods were observed during the years with longer high flow periods (i.e., 2016, 2018 and 2020, Figure 1C).
Satellite data allowed defining the hydrological context of each cruise. The winter cruise took place during a high flow and cold period. Not surprisingly, the summer cruises correspond to the warmest and driest (the lowest river flows) period. River flows and temperatures showed the highest variability in spring and autumn. However, the two autumnal cruises presented rather similar characteristics: a surface temperature around 16-17°C, a slight increase in flows rate with autumnal rains, and a moderate net primary production in surface waters. On the contrary, the two spring cruises had contrasting hydrological contexts: Gironde flows were still high in April 2018, combined with relatively high Chl-a levels, whereas they were below the mean in April 2021.
3.2 Temperature and salinity
Based on temperature and salinity profiles, the water column can be divided into two layers (Figure 2; Supplementary Figure 1). Whatever the season, bottom water salinities were quite constant with values between 34.8 and 35.4. The same pattern was observed for temperatures in the central and distal areas with values from 11.5 to 12.5°C, excepted for the cruise of August 2017 during which bottom waters were slightly warmer (between 12.8 and 13.5°C). In contrast, surface temperatures and salinities varied seasonally, partly due to freshwater inputs from the Gironde as previously described by Puillat et al. (2004).
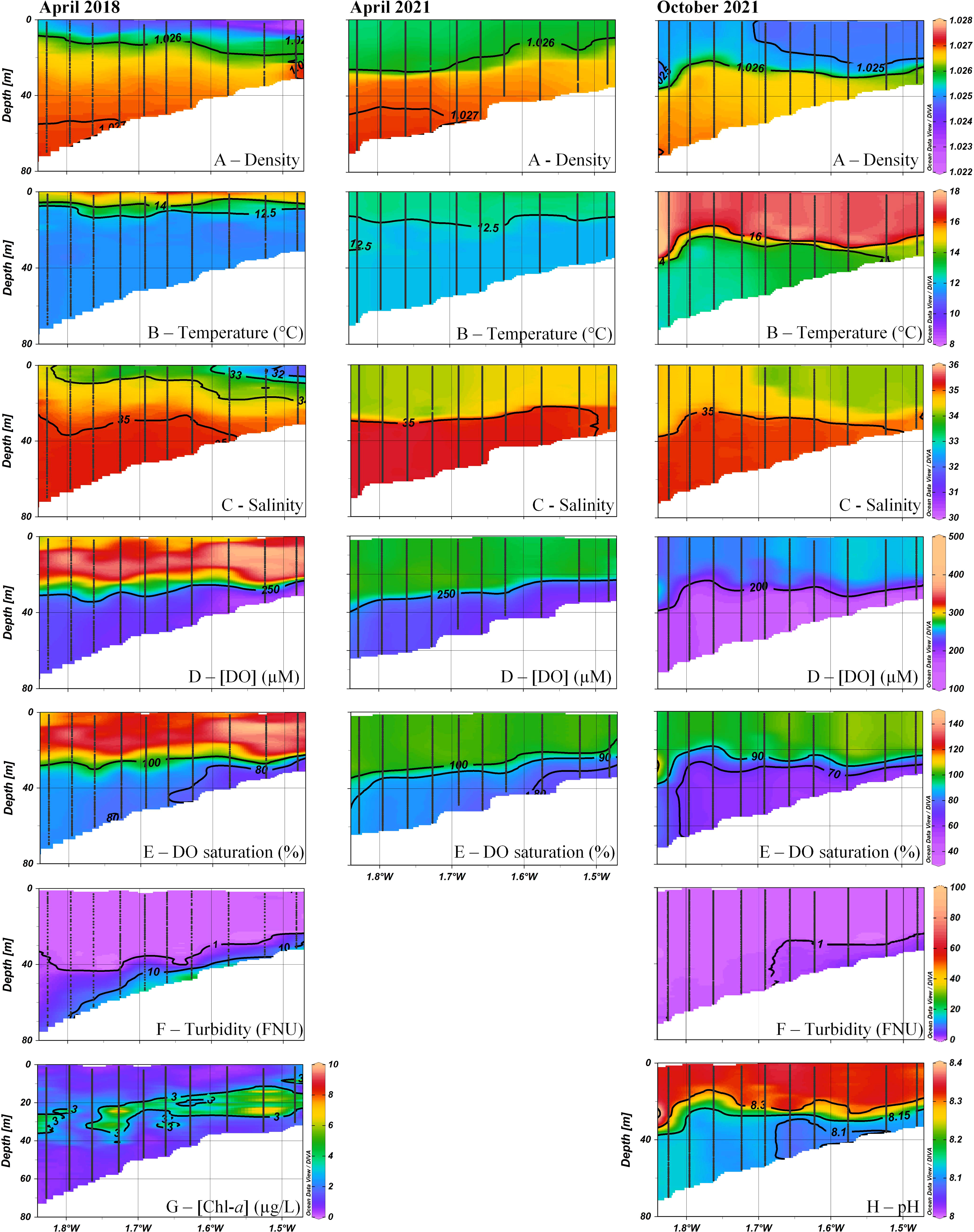
Figure 2 Transects (10 sites, from station 0 to 4) of density, temperature, salinity, dissolved oxygen (DO) concentrations and saturations, turbidity, Chl-a (April 2018) and pH (October 2021) in the water column of the West Gironde Mud Patch against longitude at three different periods (i.e., April 2018 and 2021 and October 2021). Profiles were interpolated with the software Ocean Data View. Gray dots represent the measurements points.
In winter 2018, surface waters were colder and fresher (mean salinity between 30 and 32) than bottom waters (Supplementary Figure 1) due to the Gironde plume spread, inducing a haline stratification. After the period of the highest flows, usually observed in winter (Figure 1C), flows decreased and the associated haline stratification weakened until to become negligible, as observed on salinity profiles in August 2017 and October 2016 where the salinity difference between surface and bottom waters was less than 0.5 (Supplementary Figure 1). The intensity of the spring haline stratification varied depending on years due to the variability of river flows. Indeed, the difference in flows rates resulted in lower surface salinities and a more pronounced haline stratification in April 2018 compared to April 2021 (Figure 2).
Along with the decrease in river flows and associated haline stratification, the warming of surface waters in spring induced the establishment of a thermocline, the timing of which is variable. For example, temperatures decreased with depth from 15°C to 11.6°C in April 2018 while they were rather constant along the depth profile in April 2021 (about 12.5°C, Figure 2). The thermal stratification was particularly marked in August 2017 with a temperature decrease from 21°C to 13.5°C (Supplementary Figure 1). During the two autumnal cruises, a thermocline was still present but shallower in 2016 than in 2021. The thermocline depth was highly variable from cruise to cruise, from about 15 m (April 2018) to 30 m (October 2021) in the central WGMP (i.e., station 3, Figure 2). Temperature and salinity as well as the seasonal variability of the thermal stratification are consistent with previous hydrological studies at the scale of the Bay of Biscay continental shelf (Vincent and Kurc, 1969; Koutsikopoulos and Le Cann, 1996; Puillat et al., 2004).
3.3 Dissolved oxygen, pH, Chl-a and turbidity
Dissolved oxygen concentrations in the water column were highly contrasted between cruises and seasons with values ranging from 121 to 414 µM. In January 2018, the whole water column was well-oxygenated (Figure 3; Supplementary Figure 1) due to water mixing related to winter storminess. However, in spite of this apparent homogeneity, a minimum of 90% DO saturation was observed in proximal bottom waters. From spring to autumn, the first 20 to 30 meters were also well-oxygenated (Figure 2), and even oversaturated in April 2018 up to 147%. This was related to a spring bloom as highlighted by satellite-derived data and depth profiles of Chl-a concentrations (Figures 1C, 2). On the contrary, bottom waters were DO under-saturated over the whole WGMP. These depletions were relatively weak in April 2018 and 2021 with DO saturations always above 70% but more intense in October 2016, July 2019, and October 2021 with minimum saturations of 45, 53 and 59%, respectively (Figure 3). These depletions were more pronounced in the proximal area.
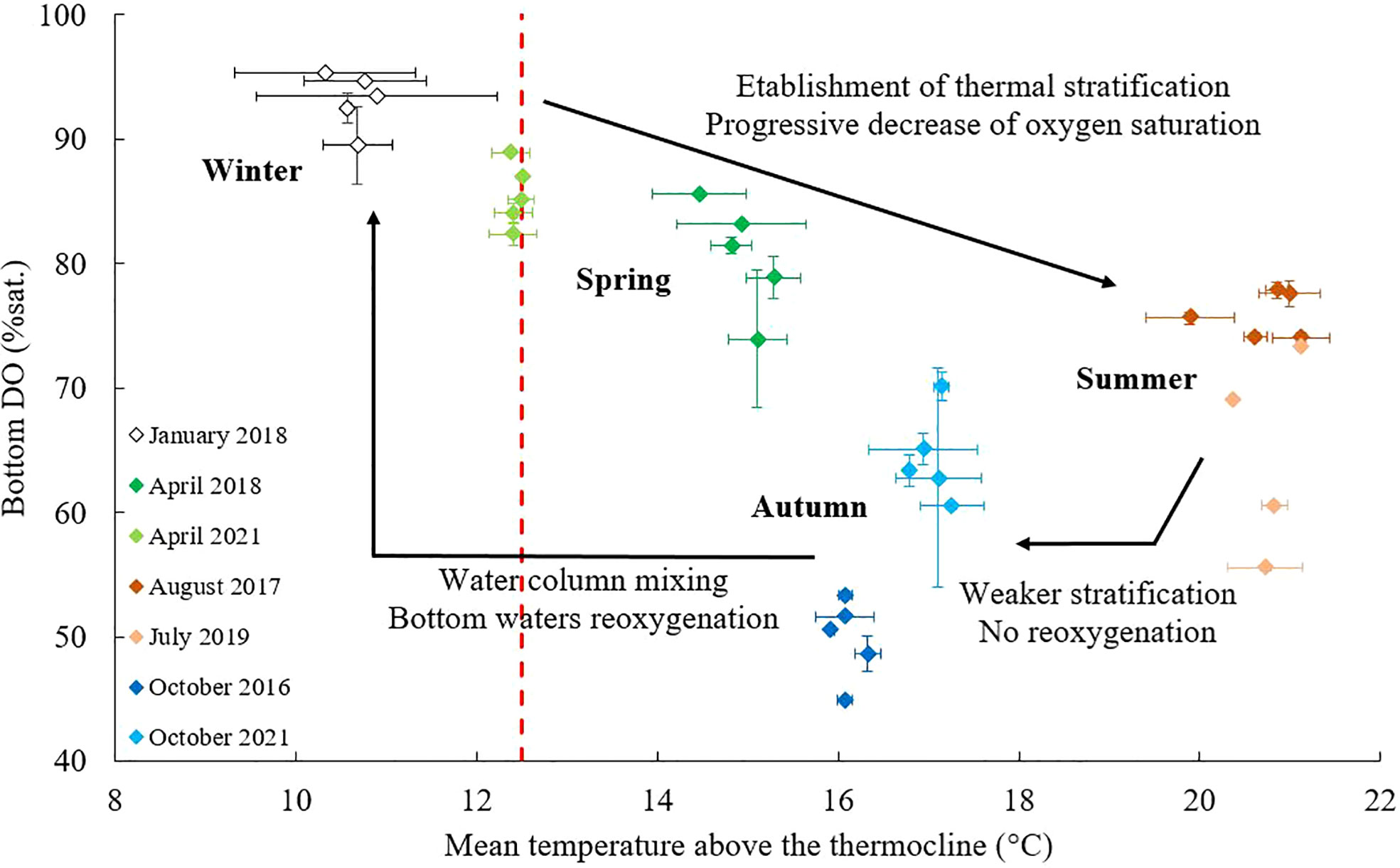
Figure 3 Mean dissolved oxygen (DO) saturation in bottom waters (based on the 5 meters above the bottom) against the mean temperature above the thermocline for each cruise and stations 1, 2, 3, 8 and 4. When several profiles were available, DO saturations and temperatures of the several profiles were averaged. The red dash line represents the threshold of 12.5°C above which the thermal stratification occurs.
Turbidity varied from negligible values to 90 FNU, highlighting the presence of a benthic nepheloid layer (Figure 2; Supplementary Figure 1). This layer was more marked in the proximal area and varied seasonally with higher turbidity in January and April 2018 (values up to 90 and 26 FNU, respectively) than in July 2019 (< 2 FNU) and October 2021 (< 11 FNU). These results are consistent with a previous study of the distribution of suspended sediments on the Bay of Biscay continental shelf (Castaing et al., 1999).
The pH profiles measured in October 2021 showed rather constant values around 8.3 in the first 20 meters, followed by a decrease in depth below 8.15, slightly more pronounced in the proximal area (Figure 2).
4 Discussion
4.1 Structuring of the water column from seasonal to inter-annual scales
Both salinity and temperature control the water column stratification. However, they are considered separately because their respective contributions to stratification are difficult to define with density alone. As bottom water temperatures and salinities were quite constant regardless of season, surface values were used to infer the intensities of thermal and haline stratifications. The steady level of bottom temperatures is consistent with previous studies that identified a water mass characterized by a constant temperature of around 12°C, “the Cold Pool”, extending from Brittany to the mouth of the Gironde (Vincent and Kurc, 1969; Koutsikopoulos and Le Cann, 1996; Puillat et al., 2004). In fact, this value is of the order of magnitude of the bottom temperatures measured in the central and distal WGMP. Considering that the bottom temperatures of the central area (i.e., station 3) varied between 11.6 and 12.5°C (except in August 2017), we defined 12.5°C as the surface temperature threshold above which thermal stratification occurs. SST records and temperature profiles support a classical annual pattern with the gradual establishment of a thermocline, the precise timing of which is variable (Figures 1C, 2, 3). After its establishment, the thermal stratification intensifies until reaching a maximum in summer, and then progressively decreases in autumn although it is still present (Figures 1C, 3). Finally, the thermocline inversion in winter indicates its complete disappearance. A significant correlation (p<0.01) between the mean salinity above the halocline and Gironde daily flows indicates that the flows control the establishment of the haline stratification. An annual pattern of the water column structuration over the shelf off the Gironde can therefore be inferred. First, a haline stratification is well established in winter. It then decreases due to decreasing flows while a thermal stratification is gradually established in spring. It intensifies until summer, and decreases in autumn until its complete removal. During the period of the thermocline disappearance, flows rates increase and a haline stratification is established again. However, this stratification varies due to contrasted flows: it was almost absent in October 2016 and slightly higher in 2021 (Figure 1C; Supplementary Figure 1).
4.2 Probable causes of deoxygenations
Dissolved oxygen measurements highlighted substantial deoxygenation of the bottom waters of the WGMP, down to a DO saturation of 45%, in July 2019, October 2016 and 2021. This was very surprising for such an environment subjected to strong hydrodynamics that control its sedimentological and ecological structuration (Lesueur et al., 2002; Dubosq et al., 2021; Lamarque et al., 2021). These DO depletions are related to the establishment of a thermal stratification from spring to autumn (Figure 3). The isolation of bottom waters from the atmosphere limits DO renewal by preventing mixing with well-oxygenated surface waters. In the isolated waters, DO is consumed by several processes, i.e., bacterial degradation of organic matter, respiration of organisms, consumption by sediment, that have gradually led to the observed depletions. It is interesting to note that the maximum DO depletion does not coincide with the maximum intensity of the thermal stratification (Figure 4). In fact, deoxygenations are more pronounced when the duration of the stratification (number of days with SST > 12.5°C) is longer (Figure 4). The stratification duration, and thus the residence time of bottom waters, therefore have a major influence on the intensity of deoxygenation. This is consistent with previous observations in other RiOMar systems such as the East China Sea (Wang et al., 2021) and the Gulf of Mexico (Rabouille et al., 2008).
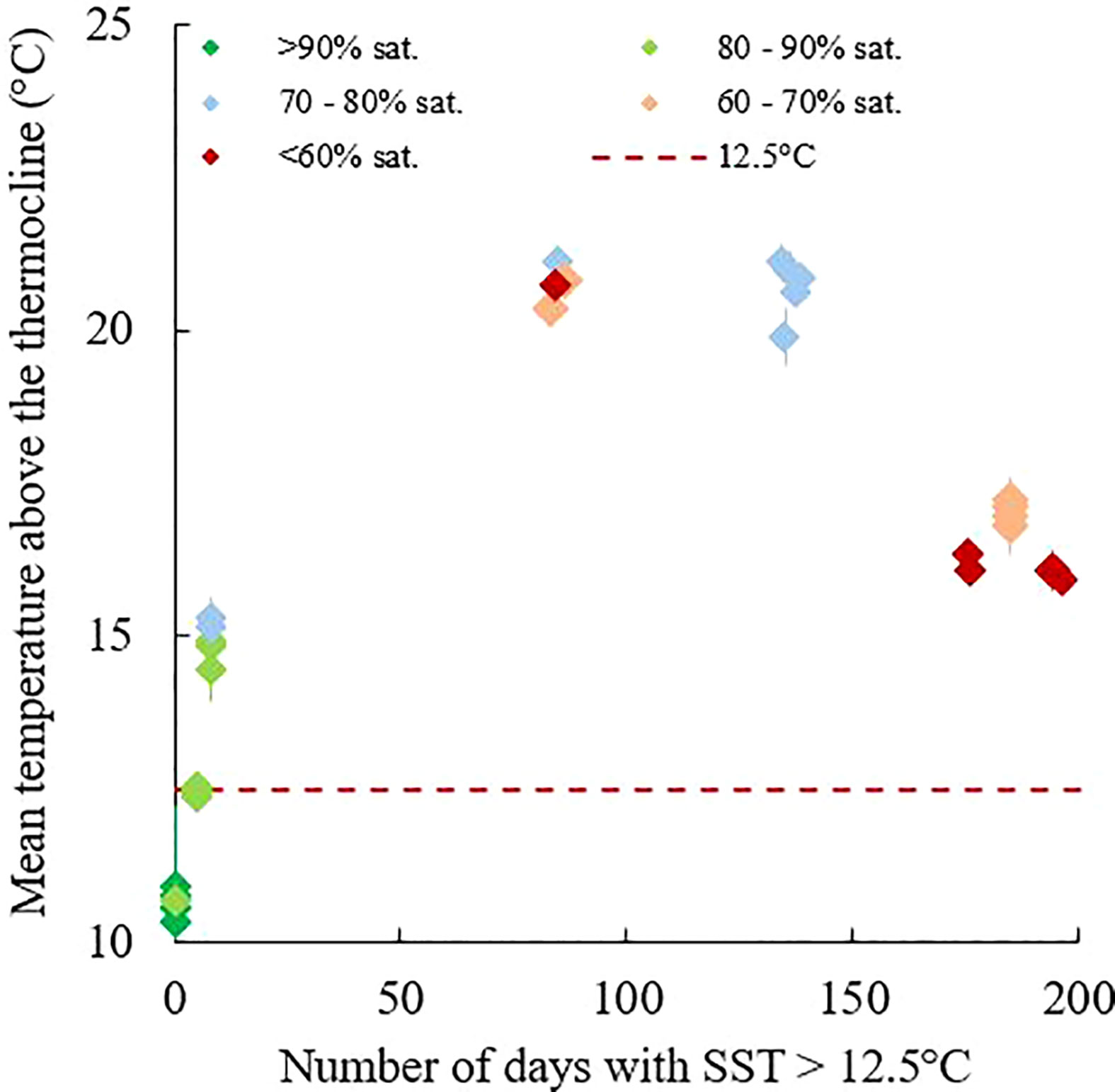
Figure 4 Relationships of the mean temperature above the thermocline, the duration of the thermal stratification (number of days with SST > 12.5°C) and the dissolved oxygen saturations of bottom waters in the water column of the continental shelf off the Gironde for each cruise and stations 1, 2, 3, 8 and 4. Dissolved oxygen saturations were average over the 5 meters above the bottom.
The bottom deoxygenation is most likely amplified by the input of organic matter from local primary production. Indeed, the higher DO depletion in autumn 2016 compared to 2021 may be due to the higher primary production of surface waters in 2016 (Figure 1C), whose sedimentation induced a higher DO consumption in the underlying waters for its degradation. The pH measurements in October 2021 revealed a positive correlation between pH and DO (r=0.903, p<0.001, Supplementary Figure 2). This indicates that aerobic respiration, which consumes oxygen and produces carbon dioxide, is the main process inducing the observed deoxygenation. Such a relationship between DO and pH was commonly observed in other oceanic environments (Cai et al., 2011; Mucci et al., 2011; Zhai et al., 2019; Steckbauer et al., 2020; Yang et al., 2021).
Primary production off the Gironde estuary is partly controlled by nutrients inputs and by winter haline stratification due to high river flows (Labry et al., 2001). The inter-annual variability of river flows therefore partly controls that of the primary production, resulting in higher surface Chl–a concentrations in years with longer periods of high flow. The higher surface Chl-a concentrations in the proximal than distal WGMP (Figure 1) support this hypothesis, suggesting that Gironde flows indirectly influence the intensity of deoxygenation. However, the rivers of the northern Bay of Biscay, the Loire and the Vilaine, are also likely to influence the waters off the Gironde, although they are more remote. A recent study based on tritium signatures of the Loire and the Gironde rivers suggested a significant input of water from the Loire off the Gironde in summer and autumn (Oms, 2019). As recurrent summer hypoxia occur in the lower Loire estuary (Schmidt et al., 2019) and the Vilaine Bay (Retho et al., 2020; Ratmaya et al., 2022), the southward spreading of these oxygen-depleted waters may reinforce the deoxygenation off the Gironde. In contrast, the Gironde does likely not exacerbate deoxygenations by supplying deoxygenated waters to the shelf as seasonal hypoxia is episodic and limited to the upper estuary (Lanoux et al., 2013; Schmidt et al., 2019).
4.3 Potential futures responses to global warming
Simulations of the evolution of European continental shelves until 2100 predict that waters of the Bay of Biscay will become fresher and warmer due to global warming (Wakelin et al., 2020). The salinity decrease is an indirect effect of climate changes related to a modification of hydrological cycles and of an increase in river flows (Gröger et al., 2013; Wakelin et al., 2020). This will result in an increase of the intensity and duration of the stratification. Simultaneously, the net primary production is expected to decrease on the Bay of Biscay continental shelf. These changes will result in an estimate decrease of bottom water DO concentrations of 10.9 ± 2.8 µM between the periods 1980 – 2009 and 2070 – 2090 (Wakelin et al., 2020), mainly related to a decrease of DO solubility induced by temperature and salinity changes. Variations in stratification or net primary production only have a significant effect on annual minimums of DO concentrations. However, this effect is low compared to that of solubility (-1.3 ± 3.1 against -8.8 ± 2.2 µM, respectively).
These results are quite surprising because our data highlights that the duration of the stratification is one of the main factors controlling the deoxygenation (Figure 4). Therefore one would have expected that the increase in the duration of stratification would reinforce deoxygenation. One explanation could be that the authors have grouped the impacts of stratification and primary production under the term “ecosystem effect”. Thus, although stratification enhanced deoxygenation, it also limits the supply of recycled nutrients from bottom to surface waters and thus reduces primary production and the DO consumption for its degradation (Wakelin et al., 2020). However, the impact of climate changes on riverine freshwater and nutrient inputs was not included in the simulation while nutrient loads and primary production can widely influence deoxygenations (Li et al., 2016). The relative effects of stratification and river flow on primary production must therefore be assessed to better estimate their respective impact on the future deoxygenations. At last, as the continental shelf off the Gironde is influenced by water advection from the north of the Bay of Biscay (Oms, 2019), it is possible that future changes of hypoxia intensities in the lower Loire estuary and the Vilaine Bay impact the deoxygenation.
4.4 The need to establish a long-term in situ monitoring of the WGMP
This study highlighted that summer and autumnal deoxygenation occurs on the continental shelf off the Gironde. This seems to be mainly related to a thermal stratification from spring to autumn inducing an isolation of the bottom waters that are progressively depleted in DO. The intensity of deoxygenation appears to be influenced by the duration of the stratification, the intensity of primary production, which is itself partly controlled by the duration of the high flow period, and by a probable advection of deoxygenated waters from the north of the Bay of Biscay. However, these hypotheses need to be reinforced. DO variability depends on several factors (e.g. consumption by respiration; physical transport; production by photosynthesis; Fennel and Testa, 2019) and is affected by different temporal cycles (e.g. daily and tidal, seasonal, decadal climatic cycles; Gilbert et al., 2010). Thus, more severe deoxygenations probably occur, but only episodically, as observed in the Gironde estuary (Schmidt et al., 2019). Furthermore, due to the multiple factors influencing the DO dynamics, it is difficult to predict its long-term evolution merely based on these first results. To better document the DO variability on large spatial and temporal scales, satellite data are a powerful tool (Kim et al., 2017). However as such data must be validated with in situ measurements (Kim et al., 2020; Triana and Wahyudi, 2021), an in situ monitoring must first be performed. The implementation of such a long-term and continuous monitoring would allow to characterize the duration and intensity of deoxygenation events and their controlling factors. These measurements coupled to satellite data would finally allow improving model outputs to anticipate the system evolution in response to climate changes and its impacts on ecosystems and fisheries. Indeed, as the common sole presents a reduced activity when DO saturations drops below 40% (Van den Thillart et al., 1994), an exacerbation of deoxygenation could question the long-term presence of this commercial species, affecting the local economy.
Considering that deoxygenation is observed throughout the whole WGMP and that the influence of the Gironde flow decreases seaward, we recommend monitoring only a single site close to the mud patch center. The site should be chosen on the basis of recent bathymetry maps and avoiding known trawling areas. Because such a dynamic area is challenging to instrument due to winter storminess, swells and currents, one recommendation would be to ensure at least a monitoring of DO, temperature, salinity and pressure in the bottom waters in order to document the occurrences, durations and intensities of deoxygenations. However, if possible, it would be ideal to perform these measurements also in surface waters to better characterize the temporal variability of the water column. In that case, the addition of chlorophyll-a, pH and pCO2 sensors would allow to precise the influence of primary production on deoxygenations and to document the pH dynamics, closely related to that of DO.
5 Conclusions
This work enables to document for the first time the DO dynamics and its controlling factors on seasonal to inter-annual scales in the water column of the continental shelf off the Gironde. Results validate the occurrence of bottom water deoxygenations that were suggested by models (Ciavatta et al., 2016; Wakelin et al., 2020). Although deoxygenation has already been reported in RiOMars (Rabouille et al., 2008; Cai et al., 2011; Fennel and Testa, 2019; Wang et al., 2021), this was quite surprising that a thermal stratification can isolate bottom waters and trigger deoxygenation in such an energetic environment. If the stratification seems to be the main factor inducing these deoxygenations, they are potentially reinforced by inputs of organic matter from local production and by the advection deoxygenated water from the northern Bay of Biscay. This work enables us to alert about the deoxygenation issues in this ecosystem of a great economic interest in a context of global warming, the potential effects of which being still uncertain. A better understanding of DO dynamics undoubtedly requires the establishment of a long-term and continuous in situ monitoring.
Data availability statement
The complete dataset of dissolved oxygen and ancillary parameters acquired during the 7 cruises between 2016 and 2021 in the water column of the West Gironde Mud Patch is freely available on the SEANOE data repository at: https://www.seanoe.org/data/00783/89508/; DOI: 10.17882/89508.
Author contributions
BD and AG conceived the projects and performed funding acquisition. SS conceived of the presented idea. ND, BD, SS, BL, SR, and MD performed in situ measurements and data qualification; ND, SS, and BD analyzed data. JS extracted and analyzed satellite data. ND wrote the manuscript with support from SS and BD. All co-authors commented on and approved the final manuscript.
Funding
This work was supported by: (1) the JERICO-NEXT project (European Union’s Horizon 2020 Research and Innovation program under grant agreement no. 654410), (2) the LEFE- EC2CO VOG project), and (3) the MAGMA project cofounded by the COTE Cluster of Excellence (ANR-10-LABX-45), the Nouvelle-Aquitaine Regional Council, and the French Biodiversity Agency. Cruises were funded by the French Oceanographic Fleet. Doctoral fellowships were provided to ND and BL by the french Ministry of Higher Education, Research and Innovation.
Acknowledgments
We sincerely acknowledge the captains and crews of the R/V “Côtes de la Manche” (CNRS-INSU) for their great help during the cruises.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1006453/full#supplementary-material
References
Breitburg D., Levin L. A., Oschlies A., Grégoire M., Chavez F. P., Conley D. J., et al. (2018). Declining oxygen in the global ocean and coastal waters. Science 359. doi: 10.1126/science.aam7240
Cai W.-J., Hu X., Huang W.-J., Murrell M. C., Lehrter J. C., Lohrenz S. E., et al. (2011). Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 4, 766–770. doi: 10.1038/ngeo1297
Castaing P., Froidefond J. M., Lazure P., Weber O., Prud’homme R., Jouanneau J. M. (1999). Relationship between hydrology and seasonal distribution of suspended sediments on the continental shelf of the bay of Biscay. Deep Sea Res. Part II: Topical Stud. Oceanogr. 46, 1979–2001. doi: 10.1016/S0967-0645(99)00052-1
Ciavatta S., Kay S., Saux-Picart S., Butenschön M., Allen J. I. (2016). Decadal reanalysis of biogeochemical indicators and fluxes in the north West European shelf-sea ecosystem. J. Geophysical Res.: Oceans 121, 1824–1845. doi: 10.1002/2015JC011496
Deflandre B., Schafer J., Coynel A., Mauffret A., Burgeot T. (2021). “MAGMA cruises,” in Côtes de la manche R/V. doi: 10.17600/18000861
Diaz R. J., Rosenberg R. (2008). Spreading dead zones and consequences for marine ecosystems. Science 321, 926–929. doi: 10.1126/science.1156401
Dubosq N., Schmidt S., Walsh J. P., Grémare A., Gillet H., Lebleu P., et al. (2021). A first assessment of organic carbon burial in the West gironde mud patch (Bay of Biscay). Continental Shelf Res. 221, 104419. doi: 10.1016/j.csr.2021.104419
Fennel K., Testa J. M. (2019). Biogeochemical controls on coastal hypoxia. Annu. Rev. Mar. Sci. 11, 105–130. doi: 10.1146/annurev-marine-010318-095138
Fontanier C., Deflandre B., Rigaud S., Mamo B., Dubosq N., Lamarque B., et al. (2022). Live (stained) benthic foraminifera from the West-gironde mud patch (Bay of Biscay, NE atlantic): Assessing the reliability of bio-indicators in a complex shelf sedimentary unit. Continental Shelf Res. 232, 104616. doi: 10.1016/j.csr.2021.104616
Gilbert D., Rabalais N. N., Díaz R. J., Zhang J. (2010). Evidence for greater oxygen decline rates in the coastal ocean than in the open ocean. Biogeosciences 7, 2283–2296. doi: 10.5194/bg-7-2283-2010
Gröger M., Maier-Reimer E., Mikolajewicz U., Moll A., Sein D. (2013). NW European Shelf under climate warming: implications for open ocean – shelf exchange, primary production, and carbon absorption. Biogeosciences 10, 3767–3792. doi: 10.5194/bg-10-3767-2013
Kim H.-C., Son S., Kim Y. H., Khim J. S., Nam J., Chang W. K., et al. (2017). Remote sensing and water quality indicators in the Korean West coast: Spatio-temporal structures of MODIS-derived chlorophyll-a and total suspended solids. Mar. pollut. Bull. 121, 425–434. doi: 10.1016/j.marpolbul.2017.05.026
Kim Y. H., Son S., Kim H.-C., Kim B., Park Y.-G., Nam J., et al. (2020). Application of satellite remote sensing in monitoring dissolved oxygen variabilities: A case study for coastal waters in Korea. Environ. Int. 134, 105301. doi: 10.1016/j.envint.2019.105301
Koutsikopoulos C., Le Cann B. (1996). Physical processes and hydrological structures retated to the bay of Biscay anchovy. Scientia Marina 9–19.
Labry C., Herbland A., Delmas D., Laborde P., Lazure P., Froidefond J., et al. (2001). Initiation of winter phytoplankton blooms within the gironde plume waters in the bay of Biscay. Mar. Ecol. Prog. Ser. 212, 117–130. doi: 10.3354/meps212117
Lamarque B., Deflandre B., Galindo Dalto A., Schmidt S., Romero-Ramirez A., Garabetian F., et al. (2021). Spatial distributions of surface sedimentary organics and sediment profile image characteristics in a high-energy temperate marine RiOMar: The West gironde mud patch. J. Mar. Sci. Eng. 9, 242. doi: 10.3390/jmse9030242
Lanoux A., Etcheber H., Schmidt S., Sottolichio A., Chabaud G., Richard M., et al. (2013). Factors contributing to hypoxia in a highly turbid, macrotidal estuary (the gironde, France). Environ. Sci.: Processes Impacts 15, 585–595. doi: 10.1039/c2em30874f
Lesueur P., Tastet J. P., Weber O. (2002). Origin and morphosedimentary evolution of fine-grained modern continental shelf deposits: the gironde mud fields (Bay of Biscay, France). Sedimentology 49, 1299–1320. doi: 10.1046/j.1365-3091.2002.00498.x
Lesueur P., Tastet J.-P., Weber O., Sinko J.-A. (1991). Modèle faciologique d’un corps sédimentaire pélitique de plate-forme : la vasière ouest-gironde (France). Oceanologica Acta, 11, 143–153.
Levin L. A. (2018). Manifestation, drivers, and emergence of open ocean deoxygenation. Annu. Rev. Mar. Sci. 10, 229–260. doi: 10.1146/annurev-marine-121916-063359
Li M., Lee Y. J., Testa J. M., Li Y., Ni W., Kemp W. M., et al. (2016). What drives interannual variability of hypoxia in Chesapeake bay: Climate forcing versus nutrient loading? Geophysical Res. Lett. 43, 2127–2134. doi: 10.1002/2015GL067334
Ménesguen A., Dussauze M., Dumas F., Thouvenin B., Garnier V., Lecornu F., et al. (2019). Ecological model of the bay of Biscay and English channel shelf for environmental status assessment part 1: Nutrients, phytoplankton and oxygen. Ocean Model. 133, 56–78. doi: 10.1016/j.ocemod.2018.11.002
Mucci A., Starr M., Gilbert D., Sundby B. (2011). Acidification of lower st. Lawrence estuary bottom waters. Atmosphere-Ocean 49, 206–218. doi: 10.1080/07055900.2011.599265
NASA Ocean Color. (USA: NASA) Available at: https://oceancolor.gsfc.nasa.gov/.
Oms P.-E. (2019) Transferts multi-échelles des apports continentaux dans le golfe de gascogne. Available at: https://tel.archives-ouvertes.fr/tel-02354264.
Oschlies A., Brandt P., Stramma L., Schmidtko S. (2018). Drivers and mechanisms of ocean deoxygenation. Nat. Geosci. 11, 467–473. doi: 10.1038/s41561-018-0152-2
Pond S., Pickard G. L. (2007). Introductory dynamical oceanography. 3. ed (Oxford: Butterworth-Heinemann).
Puillat I., Lazure P., Jégou A. M., Lampert L., Miller P. I. (2004). Hydrographical variability on the French continental shelf in the bay of Biscay, during the 1990s. Continental Shelf Res. 24, 1143–1163. doi: 10.1016/j.csr.2004.02.008
Rabouille C., Conley D. J., Dai M. H., Cai W.-J., Chen C. T. A., Lansard B., et al. (2008). Comparison of hypoxia among four river-dominated ocean margins: The changjiang (Yangtze), Mississippi, pearl, and rhône rivers. Continental Shelf Res. 28, 1527–1537. doi: 10.1016/j.csr.2008.01.020
Ratmaya W., Laverman A. M., Rabouille C., Akbarzadeh Z., Andrieux-Loyer F., Barillé L., et al. (2022). Temporal and spatial variations in benthic nitrogen cycling in a temperate macro-tidal coastal ecosystem: Observation and modeling. Continental Shelf Res. 235, 104649. doi: 10.1016/j.csr.2022.104649
Retho M., Quemener L., Le Gall C., Repecaud M., Souchu P., Gabellec R., et al. (2020). MOLIT vilaine data and metadata from Coriolis data centre. (France:SEANOE) doi: 10.17882/46529
Roman M. R., Brandt S. B., Houde E. D., Pierson J. J. (2019). Interactive effects of hypoxia and temperature on coastal pelagic zooplankton and fish. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00139
Rossignol-Strick M. (1985). A marine anoxic event on the Brittany coast, July 1982. J. Coast. Res. 1, 11–20.
Schmidt S., Diallo I. I., Derriennic H., Fallou H., Lepage M. (2019). Exploring the susceptibility of turbid estuaries to hypoxia as a prerequisite to designing a pertinent monitoring strategy of dissolved oxygen. Front. Mar. Sci. 6, 352. doi: 10.3389/fmars.2019.00352
Steckbauer A., Klein S. G., Duarte C. M. (2020). Additive impacts of deoxygenation and acidification threaten marine biota. Global Change Biol. 26, 5602–5612. doi: 10.1111/gcb.15252
Testa J. M., Kemp W. M. (2011). "Oxygen – Dynamics and Biogeochemical Consequences". in Treatise on Estuarine and Coastal Science Eds. Wolanski E., McLusky D. S. (Waltham: Academic Press), 163–199.
Triana K., Wahyudi A. J. (2021). Dissolved oxygen variability of Indonesian seas over decades as detected by satellite remote sensing. IOP Conf. Series: Earth Environ. Sci. 925, 012003. doi: 10.1088/1755-1315/925/1/012003
Van den Thillart G., Dalla Via J., Vitali G., Cortesi P. (1994). Influence of long-term hypoxia exposure on the energy metabolism of solea solea. i. critical 02 levels for aerobic and anaerobic metabolism. Mar. Ecol. Prog. Ser. 104, 109–117. doi: 10.3354/meps104109
Vincent A., Kurc G. (1969). Hydrologie, variations saisonnières de la situation thermique dans le golfe de gascogne en 1967. Rev. Des. travaux l'Institut Des. pêches Maritimes 33, 79–96.
Wakelin S. L., Artioli Y., Holt J. T., Butenschön M., Blackford J. (2020). Controls on near-bed oxygen concentration on the Northwest European continental shelf under a potential future climate scenario. Prog. Oceanogr. 187, 102400. doi: 10.1016/j.pocean.2020.102400
Wang K., Cai W.-J., Chen J., Kirchman D., Wang B., Fan W., et al. (2021). Climate and human-driven variability of summer hypoxia on a Large river-dominated shelf as revealed by a hypoxia index. Front. Mar. Sci. 8, 634184. doi: 10.3389/fmars.2021.634184
Yang B., Gao X., Zhao J., Liu Y., Xie L., Lv X., et al. (2021). Summer deoxygenation in a bay scallop (Argopecten irradians) farming area: The decisive role of water temperature, stratification and beyond. Mar. pollut. Bull. 173, 113092. doi: 10.1016/j.marpolbul.2021.113092
Keywords: deoxygenation, continental shelf, seasonal evolution, stratification, Bay of Biscay, dissolved oxygen
Citation: Dubosq N, Schmidt S, Sudre J, Rigaud S, Lamarque B, Danilo M, Grémare A and Deflandre B (2022) First observations of seasonal bottom water deoxygenation off the Gironde estuary (Bay of Biscay, North East Atlantic). Front. Mar. Sci. 9:1006453. doi: 10.3389/fmars.2022.1006453
Received: 29 July 2022; Accepted: 20 September 2022;
Published: 07 October 2022.
Edited by:
Ivica Vilibic, Rudjer Boskovic Institute, CroatiaReviewed by:
Yun Li, University of Delaware, United StatesKarlina Triana, Indonesian Institute of Sciences, Indonesia
Copyright © 2022 Dubosq, Schmidt, Sudre, Rigaud, Lamarque, Danilo, Grémare and Deflandre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicolas Dubosq, nicolas.dubosq@u-bordeaux.fr