A Case Study of a Near Vessel Strike of a Blue Whale: Perceptual Cues and Fine-Scale Aspects of Behavioral Avoidance
 Angela R. Szesciorka1*†
Angela R. Szesciorka1*†  Ann N. Allen1
Ann N. Allen1  John Calambokidis1
John Calambokidis1  James Fahlbusch1,2
James Fahlbusch1,2  Megan F. McKenna3
Megan F. McKenna3  Brandon Southall4,5
Brandon Southall4,5- 1Cascadia Research Collective, Olympia, WA, United States
- 2Hopkins Marine Station, Department of Biology, Stanford University, Pacific Grove, CA, United States
- 3National Park Service, Natural Sounds and Night Skies Division, Fort Collins, CO, United States
- 4Southall Environmental Associates, Inc., Aptos, CA, United States
- 5Institute of Marine Sciences, Long Marine Laboratory, University of California, Santa Cruz, Santa Cruz, CA, United States
Despite efforts to aid recovery, Eastern North Pacific blue whales faces numerous anthropogenic threats. These include behavioral disturbances and noise interference with communication, but also direct physical harm – notably injury and mortality from ship strikes. Factors leading to ship strikes are poorly understood, with virtually nothing known about the cues available to blue whales from nearby vessels, behavioral responses during close encounters, or how these events may contribute to subsequent responses. At what distance and received levels (RLs) of noise whales respond to potential collisions is difficult to observe. A unique case study of a close passage between a commercial vessel and a blue whale off Southern California is presented here. This whale was being closely monitored as part of another experiment after two suction-cup archival tags providing acoustic, depth, kinematic, and location data were attached to the whale. The calibrated, high-resolution data provided an opportunity to examine the sensory information available to the whale and its response during the close encounter. Complementary data streams from the whale and ship enabled a precise calculation of the distance and acoustic cues recorded on the tag when the whale initiated a behavioral response and shortly after at the closest point of approach (CPA). Immediately before the CPA, the whale aborted its ascent and remained at a depth sufficient to avoid being struck for ∼3 min until the ship passed. In this encounter, the whale may have responded to a combination of cues associated with the close proximity of the vessel to avoid a collision. Long-term photo-identification records indicate that this whale has a long sighting history in the region, with evidence of previous ship encounters. Therefore, experiential factors may have facilitated the avoidance of a collision. In some instances these factors may not be available, which may make some blue whales particularly susceptible to deadly collisions, rendering efforts for ship-strike reduction even more challenging. The fine-scale information made available by the integration of these methods and technologies demonstrates the capacity for detailed behavioral studies of blue whales and other highly mobile marine megafauna, which will contribute to more informed evaluation and mitigation strategies.
Introduction
Like most baleen whales, blue whales (Balaenoptera musculus) were greatly depleted by commercial whaling (Monnahan et al., 2014). Abundance estimates from mark-recapture data suggest no evidence of an increase in this population since the early 1990s (Calambokidis, 2013), with the population currently estimated at 1,647 individuals. With pre-whaling abundance estimates modeled at between 1,823 and 3,721 individuals, this has led some to the conclusion that blue whales had returned to carrying capacity (Monnahan et al., 2014). However, the coastal habitats where blue whales feed on euphausiid aggregations (Rice, 1974; Croll et al., 1998; Fiedler et al., 1998; Calambokidis et al., 2009, 2015) overlap with human activities. As a result, these whales are vulnerable to many anthropogenic threats, including ship strikes.
Ship-strikes off California have resulted in the death of at least nine blue whales from 2007 to 2011 (Berman-Kowalewski et al., 2010; Carretta et al., 2013), though this is an underestimate of the true number due to the small proportion of large whale mortality that is documented (Heyning and Dahlheim, 1990; Kraus et al., 2005; Williams et al., 2011). A recent model estimated a true mortality of 18 blue whales per year off the United States West Coast (Rockwood et al., 2017). That is nearly eight times greater than the potential biological removal limit (Carretta et al., 2011), defined under the United States Marine Mammal Protection Act of 1972 as the maximum number of animals, not including natural mortalities, that may be removed while allowing that stock to reach or maintain its optimum sustainable population. The factors leading to a ship strike are poorly understood, difficult to predict, and subsequently difficult to prevent. Despite mitigation efforts, including ship speed limits and adjustments to the size and location of the major shipping lanes (DeAngelis et al., 2010; McKenna et al., 2012a; Redfern et al., 2013), ship strikes continue, and questions remain about the role the behavioral response of the animal plays in ship-strike risk.
Previous research found that during nine close encounters with large commercial ships, blue whales did not respond by moving horizontally, but may have altered their diving behavior. These dives were only observed when ships were within a few hundreds of meters of the whales, a range that might not allow for much avoidance time (McKenna et al., 2015). Their constrained response time may result from external cues that are only detectable – or interpreted as a threat – at limited distances, making them vulnerable to ship strikes. The detectable perceptual cues (e.g., visual and acoustic) corresponding to the presence of close-range vessels that provoke these types of avoidance responses are unknown. It is hypothesized that blue whales use visual cues to identify prey patches on the surface (Goldbogen et al., 2013a; Friedlaender et al., 2017) and could conceivably use vision to identify a large ship. Although whales may be able to visually detect ships at or below the surface over short ranges and under ideal ambient light conditions, sound propagates much further in water than light, likely making sound the primary sensory cue for whales orienting to their surroundings. Blue whales are acoustically active animals (Oleson et al., 2007) and noise from commercial ships directly overlaps with their vocalization frequency range. These ships emit a significant amount of low-frequency underwater noise (<1,000 Hz), which poses additional threats to this endangered population (e.g., masking whale communication, increasing stress, and resulting in habituation to ship presence, potentially limiting avoidance responses and times) (McKenna et al., 2012b).
A unique incident involving a well-documented close passage between a large ship and a tagged blue whale arose during an experimental study of blue whale behavioral response to military sonar (see: Southall et al., 2019). Fine scale movement and acoustic data were collected, including estimated distances between the whale and ship, vessel noise received levels (RLs) on the tag, and three-dimensional fine-scale kinematic behavioral response. We use this unique event to gain insights into the various perceptual cues that may be used by whales to avoid ships, and to evaluate implications for ship strike risk.
Materials and Methods
Data Collection
On September 13, 2014, a blue whale was dual tagged with a TDR10 tag (Wildlife Computers, Redmond, WA, United States) and a digital acoustic recording tag (DTAG-3; Woods Hole Oceanographic Institution, Woods Hole, MA, United States; Johnson and Tyack, 2003), in the Santa Barbara Channel (SBC) (33.66°N, 118.30°W). Both tags were simultaneously attached via suction cups in a single tagging approach at 0848 (local time henceforth). The animal was tagged as part of ongoing studies of whale behavior in shipping lanes (McKenna et al., 2015) and the Southern California Behavioral Response Study (SOCAL-BRS), a multi-year study of the response of different cetaceans to exposure of Navy sonar sounds conducted in the Southern California Bight (see Southall et al., 2019). As part of the SOCAL-BRS experiment, the animal was exposed to a 30-min experiment involving simulated mid-frequency (3–4 kHz) active sonar (MFAS), which ended 62 min prior to the close encounter with a large commercial ship.
A tagging boat (5.9 m rigid-hull inflatable boat; RHIB) was used to deploy the tags with a ∼5-m carbon fiber pole. The whale exhibited no visible reaction during tagging and resumed the behavior observed prior to tagging (i.e., consistent traveling). The animal was photographed and compared with known individuals in the Cascadia Research photograph identification catalog database (Calambokidis et al., 2009, 2015). While a skin sample was collected via biopsy, the sex of the animal was identified as female from a previous biopsy of this individual. The tagged animal’s positions were recorded during a focal follow in order to provide georeferenced positions for the pseudotrack generated from tag data (see section “Distance Calculations”). In the focal follow two vessels were involved in observing the tagged whale. The RHIB stayed 100–200 m away until the whale made its terminal dive, then slowly approached the location to record the exact dive position from the whale’s footprint. A larger (22 m) vessel remained at distances of 362 to 2,750 m (on average 500–1,500 m) from the whale when it was at the surface and provided visual tracking support. Both vessels followed the methodology developed for the SOCAL-BRS experiment to ensure the presence of small boats would not impact behavior (see: Southall et al., 2012, 2016).
The DTAG-3 recorded dual-channel acoustics at a 240-kHz sampling rate, while pressure, temperature, and a tri-axial accelerometer and magnetometer were sampled at 250 Hz. The TDR10’s pressure sensor recorded at 1 Hz and the FastGPS sensor took sub-second instantaneous satellite position snapshots when the tag emerged from the water during surfacings of the whale. Both tags were deployed with VHF transmitters used for locating the tagged whale and for tag recovery. The DTAG-3 remained attached to the animal for 5.7 h while the TDR10 remained attached for 15 hr. The data from the two tags were synchronized based on the timestamps.
Kinematic Analysis
The three-axis accelerometer and magnetometer data from the DTAG-3 were down-sampled to 5 Hz and corrected in MATLAB (Mathworks, Natick, MA, United States) so the axes aligned with the “whale frame” using periods of known orientation (Johnson and Tyack, 2003). Animal orientation (i.e., pitch, roll, and heading) was calculated using custom-written MATLAB scripts (Johnson and Tyack, 2003; Cade et al., 2016). Animal speed was determined from the root-mean-square (RMS) amplitude of flow noise from tag acoustics (Goldbogen et al., 2006; Simon et al., 2009). Lunges indicative of feeding were detected from the DTAG-3 data using a custom-developed lunge detection algorithm [similar to Allen et al. (2016)]. Depths recorded by the TDR10 pressure sensor were assessed in R (R Core Team, 2019) using the package “diveMove” (Luque, 2007) to determine the number of dives and maximum depth per dive performed by the tagged whale. Dives recorded only on the TDR10 were manually audited for the presence of vertical lunges as a coarse determination of presumed feeding. Dives were classified as lunge-feeding or non-lunge feeding based on the presence or absence of lunges during each dive. This gave us four generalized behavioral states for each dive.
Distance Calculations
Ship positions from the Automatic Identification System (AIS), the global ship tracking system used by vessel traffic services, were obtained for the period when the whale was tagged from an AIS receiver on Santa Cruz Island (33.995°N, 119.632°W). Whale surface locations were resolved from satellite position snapshots for surfacings detected on the TDR10’s FastGPS sensor during which an adequate number of satellites (>4) were identified. We generated a georeferenced pseudotrack at 1 Hz sampling rate using the depth, pitch, speed, and known geographic reference points of the tagged animal (GPS positions from the TDR10 and focal follow positions) (Wilson et al., 2007). Ship positions were interpolated to 1-s intervals with the “ST_Line_Interpolate_Point” function in PostGIS assuming a constant speed and course over ground. The PostGIS “ST_Distance_Sphere” function was used to calculate horizontal distances from the tagged whale to every ship present in the AIS data. Three-dimensional straight-line distances were calculated as the hypotenuse of the horizontal and vertical distance between the ship and the whale and rounded to 10-m intervals. Horizontal distances were calculated as distance between the whale and the closest point to the ship after accounting for the location of the AIS transmitter on the ship and orientation relative to the whale. Vertical distances were calculated as the distance between the ship’s reported draft and the whale’s depth (determined from the TDR10’s pressure sensor).
Acoustic Analysis
The acoustic data from the DTAG-3 were initially viewed as 60-s spectrograms calculated from 10 Hz to 120 kHz in MATLAB using Triton, custom-written software (Wiggins, 2003), to identify ship noise. To extract sound levels from the DTAG-3, the acoustic data were first decimated to 48 kHz, and the broadband (0 Hz–48 kHz) RMS received sound pressure levels (dB re 1 μPa) were calculated in 1-s intervals. Additionally, the power spectral density was calculated at a 1s-resolution and then summed over 1/3-octave band sound pressure levels (dB re 1 μPa) for bands with center frequencies ranging from 160 Hz to 20 kHz, using methods described in Merchant et al. (2015).
Noise generated from water flowing over the tag hydrophone (flow noise) can contribute to acoustic measurements of actual noise in the environment at frequencies up to 1 kHz. Flow noise highly correlates with whale swim speed and fluking (Goldbogen et al., 2006; Simon et al., 2009) and the noise tends to predominate at frequencies below 100 Hz (Fletcher et al., 1996). Therefore, this study excluded 1/3-octave bands below 140 Hz from the calculations of noise levels associated with the vessel. Flow noise above 140 Hz, to the extent it was present, was considered to be a relatively constant element of overall noise and included as part of the noise level calculations.
Controlled Exposure Experiment
As part of the SOCAL-BRS project, the animal was exposed to simulated MFAS from 1045 to 1115 PDT (local), during which a stationary experimental sound source (deployed from the M/V Truth) was positioned at ranges from ∼800 m to >2 km from the whale. Prior to the controlled exposure experiment (CEE), prey mapping with a calibrated multi-beam echosounder occurred from 0910 to 1008 [as in Friedlaender et al. (2016)]. From tag deployment, until the CEE began (117 min), the animal’s baseline behavior was recorded during focal follow. After 30 min of MFAS exposure, post-exposure focal follow and prey mapping began, which ended at 1238. The animal was feeding before, during, and after the CEE and while behavioral changes were identified as a result of the experiment CEEs (Southall et al., 2019), these were ephemeral in nature. The animal exhibited typical deep feeding dives for the 62 min-period following the CEE and prior to the vessel encounter. The DTAG-3 detached from the whale at 1416 and the TDR10 detached at 2346.
Photograph Identification
Based on the identification of the whale from matches in Cascadia Research’s catalog and database, the animal was a known female that had been seen previously 23 times off the California coast in eight different years beginning in 1987. Most of the sightings were in the Southern California Bight in the vicinity of Palos Verdes Peninsula, a region near the shipping lanes leading to the port of Los Angeles/Long Beach and near where the animal was tagged in this study. The animal was also sighted off Pt. Reyes, California, a region near the northbound shipping lanes leaving San Francisco Bay. The animal was previously tagged during the 2011 SOCAL-BRS on August 3, 2011, however the tag remained attached for only 1 hr and therefore no playback experiment occurred. This whale was also sighted when the tagged whale and another whale were involved in the capsizing of a 23-foot private vessel off San Diego on July 2, 2014 (∼2 months prior to the encounter described here), after the boat approached the whales to take photographs. There were no reports of injury to the whales following the incident.
Results
The R package “diveMove” detected 118 dives from the TDR10 pressure sensor data (Figure 1). The DTAG-3 pressure sensor captured the first 33 of these dives. Of the 118 dives detected, 12 were deep lunge-feeding dives, 35 were deep non-lunge feeding dives, 4 were shallow lunge-feeding dives, and 67 were shallow non-lunge feeding dives. At the onset of tagging, the whale was making a series of deep non-lunge feeding dives interspersed with lunge-feeding dives as she traveled southeast along the 200-m contour line (Figure 1). Two lunge-feeding periods were identified, one from 0910 to 1057, which occurred during the CEE and included 1 deep and 3 shallow lunge-feeding dives, and one from 1613 to 1930, which included 8 deep and 4 shallow lunge-feeding dives. Sunset occurred at 1854. From 1930, the onset of civil twilight, until the TDR10 tag detached at 2346, the dive record suggested a resting bout of 4 h and 15 min during which the whale stayed shallower than 35 m and no lunges were detected.
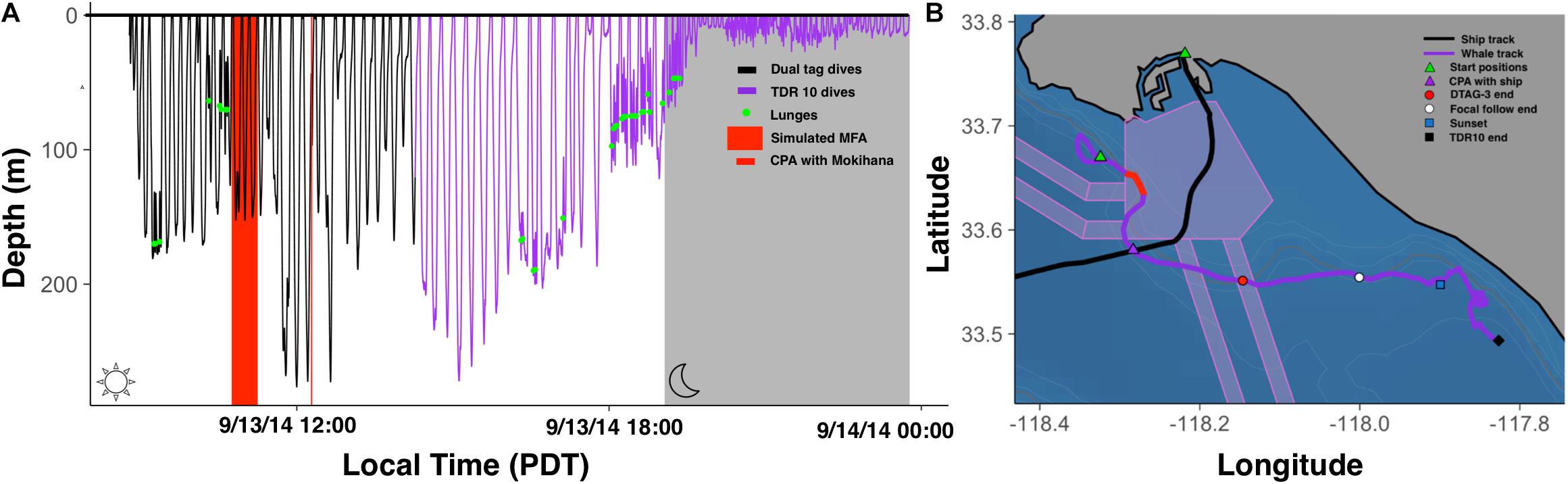
Figure 1. (A) Dive record from DTAG-3 data (black) and TDR10 data (purple). Detected lunges are indicated by green circles. Red shading indicates simulated mid-frequency (3–4 kHz) active sonar playback as part of the SOCAL-BRS CEE. Red line indicates closest CPA with container ship Mokihana. (B) Horizontal tracks of Mokihana (black) and tagged whale (purple). Green triangles represent start positions for ship and whale. Red shading indicates period of the CEE. Purple triangle indicates the whale location during the CPA. Red circle indicates end of DTAG-3 recording, white circle indicates conclusion of focal follow, the blue square indicates sunset, and black square indicates end of TDR10 attachment. Shipping lanes are pink polygons and contour lines are represented in light gray from 50 to 500 m (in 50 m increments), with the 200-m contour in dark gray.
The TDR10 collected 122 resolvable GPS locations. Distance calculations between the ship and whale tracks revealed three instances where an underway ship was within 2 km of the tagged whale. The closest point of approach (CPA) between the Mokihana, a 263-m container ship traveling at 11.3 knots, and the tagged whale occurred at a horizontal distance of 93 m while the whale was at a depth of 67.5 m (Figures 1, 2). The corrected horizontal distance from the AIS transmitter on the boat at the starboard side closest to the whale was 77 m and the corrected vertical distance between the whale and the reported draft of the ship (10 m) was 57.5 m. The 3D straight-line distance between the Mokihana and the tagged female blue whale was approximately 100 m. The other two ships passed at horizontal distances greater than 1.5 km from the whale and occurred after the MFAS CEE during the post-exposure focal follow and prey mapping.
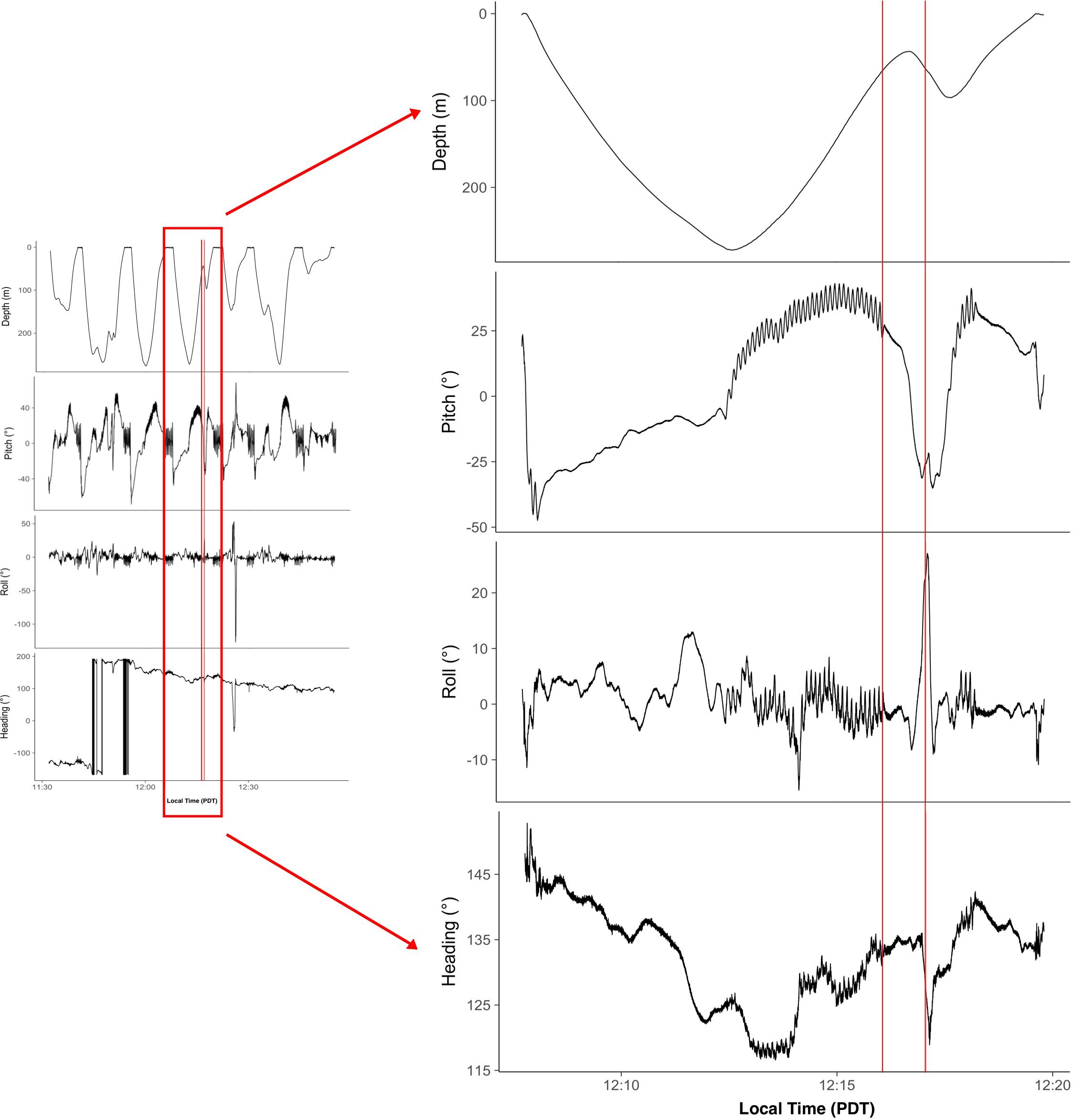
Figure 2. Close up of fine scale kinematic data recorded during the CPA with the Mokihana. The first panel illustrates the depth (meters), the second panel shows pitch (degrees), the third panel illustrates roll (degrees), and the fourth panel illustrates heading (degrees). The first solid red line indicates the onset of a behavioral response by the whale. The second solid red line indicates the CPA with the Mokihana.
Behavioral Response During CPA With Mokihana
Prior to the CPA with the Mokihana, the tagged whale was ascending from a deep non-lunge feeding dive (max depth = 277.5 m). The whale began to slow its ascent ∼90 s before the CPA. Forty-seconds before the CPA, while the ship was at an approximate 3D straight line distance (hypotenuse between the ship and the whale) of 300 m from the whale, the tagged whale reversed into a descent. Kinematic data from the DTAG-3 shows a change in pitch, which corresponds to the switch to descent. The CPA occurred as the whale was at a depth of 57.5 m from the ship’s draft. By this time the ship was approximately 100 m away from the whale at a 3D straight line distance. The data also indicate that the whale rolled to the left and changed its heading quickly at the CPA. The tagged whale resumed its ascent and surfaced after a ∼3-min delay from the previous projected surfacing time (Figure 2).
Before the close approach of the vessel, the broadband (RMS) ambient noise was generally ∼125–130 dB re 1 μPa (Figures 3, 4). The overall ambient conditions in this environment were likely strongly influenced by aggregate vessel noise in the general area, including the Mokihana. However, as the ship approached, there was a rapid increase in the acoustic energy at higher frequencies (>1 kHz) with a typical spectral and temporal pattern associated with large vessels (McKenna et al., 2012b). The lower frequency bands (<1 kHz) exhibited an initial drop, associated with the cessation of fluking by the whale. These lower frequency bands then exhibited a rapid increase in levels, with no concurrent increase in fluking activity. The increase was instead associated with the passing of the ship within 100 m of the whale. The broadband sound level at CPA peaked at 135 dB re 1 μPa compared to ∼125 dB re 1 μPa at the last approximate point with similarly no fluking activity (Figure 4), representing a 10-dB increase over ambient broadband levels. Higher frequency (>1 kHz) 1/3-octave levels increased by up to 40 dB over pre-ship ambient levels. Additionally, as indicated in the noise spectra (Figure 3) and the broadband RMS RLs (Figure 4), there was a relatively abrupt change in the received sound levels around the point at which the whale initiated a change in behavior. There was a subsequent peak in the noise in all frequencies corresponding to the CPA of the Mokihana. The 1/3-octave band sound levels (Figure 4) indicate that the whale initiated a response dive when higher frequency (>1 kHz) RLs were only a few dB above ambient levels, just prior to reaching their maximum values. The whale only resurfaced after the vessel passed and was moving away, at which point RLs and the prevalence of higher frequency noise energy from the vessel were decreasing. The broadband RMS sound levels indicate a second peak after the passage of the ship, which corresponds to the resumption of fluking (evident in pitch, Figure 2) as the whale ascends. This peak in acoustic energy is only evident in the low frequency components of the 1/3-octave band levels, further indicating the second peak in broadband sound levels is due to increased flow noise associated with fluking.
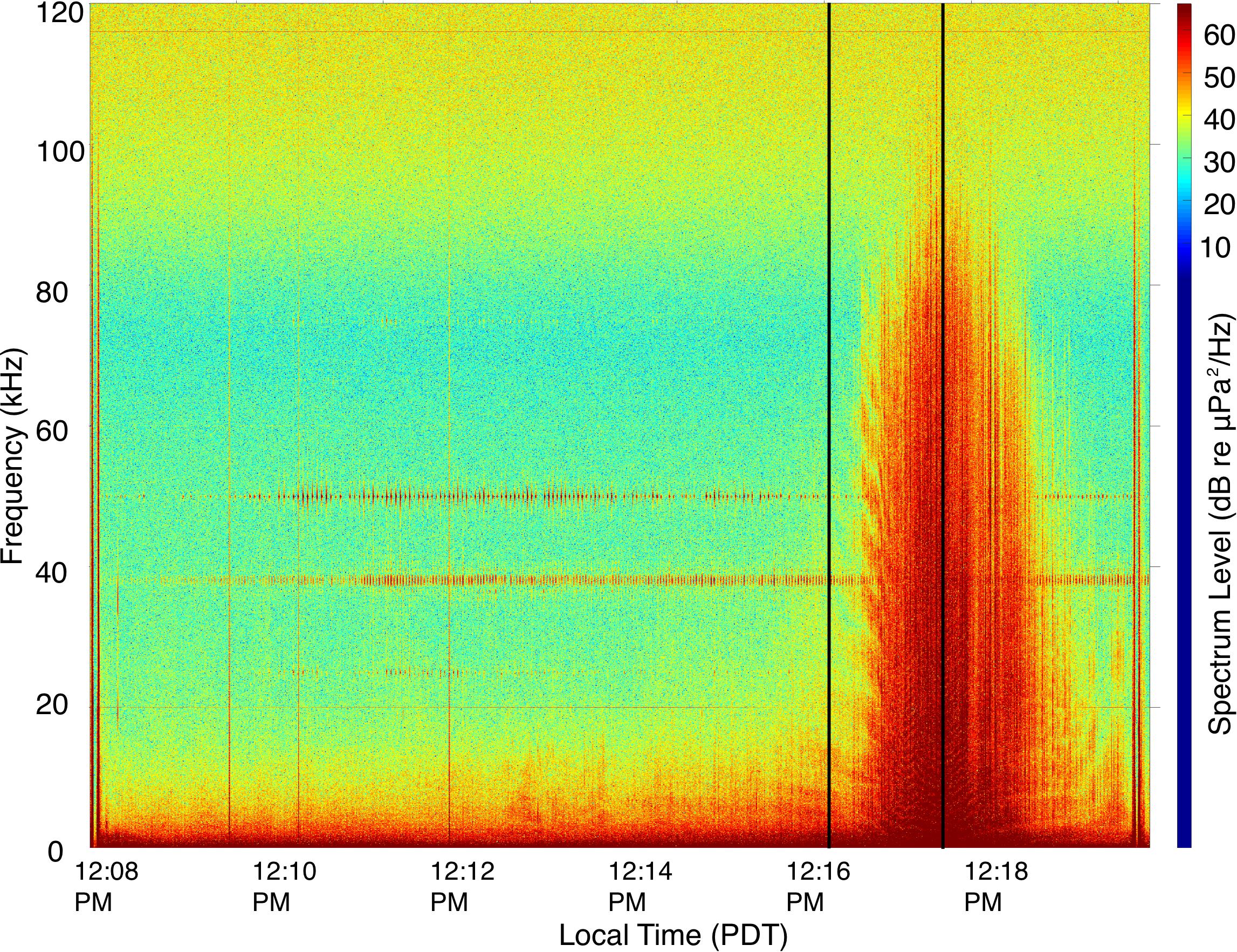
Figure 3. Spectrogram showing the acoustic signal of the Mokihana during the CPA as recorded on the DTAG-3 hydrophone. The first black vertical line indicates the onset of a behavioral response by the whale. The second black vertical line indicates the CPA with the Mokihana. Spectrogram parameters: NFFT = 240000, 90% overlap, Hanning window.
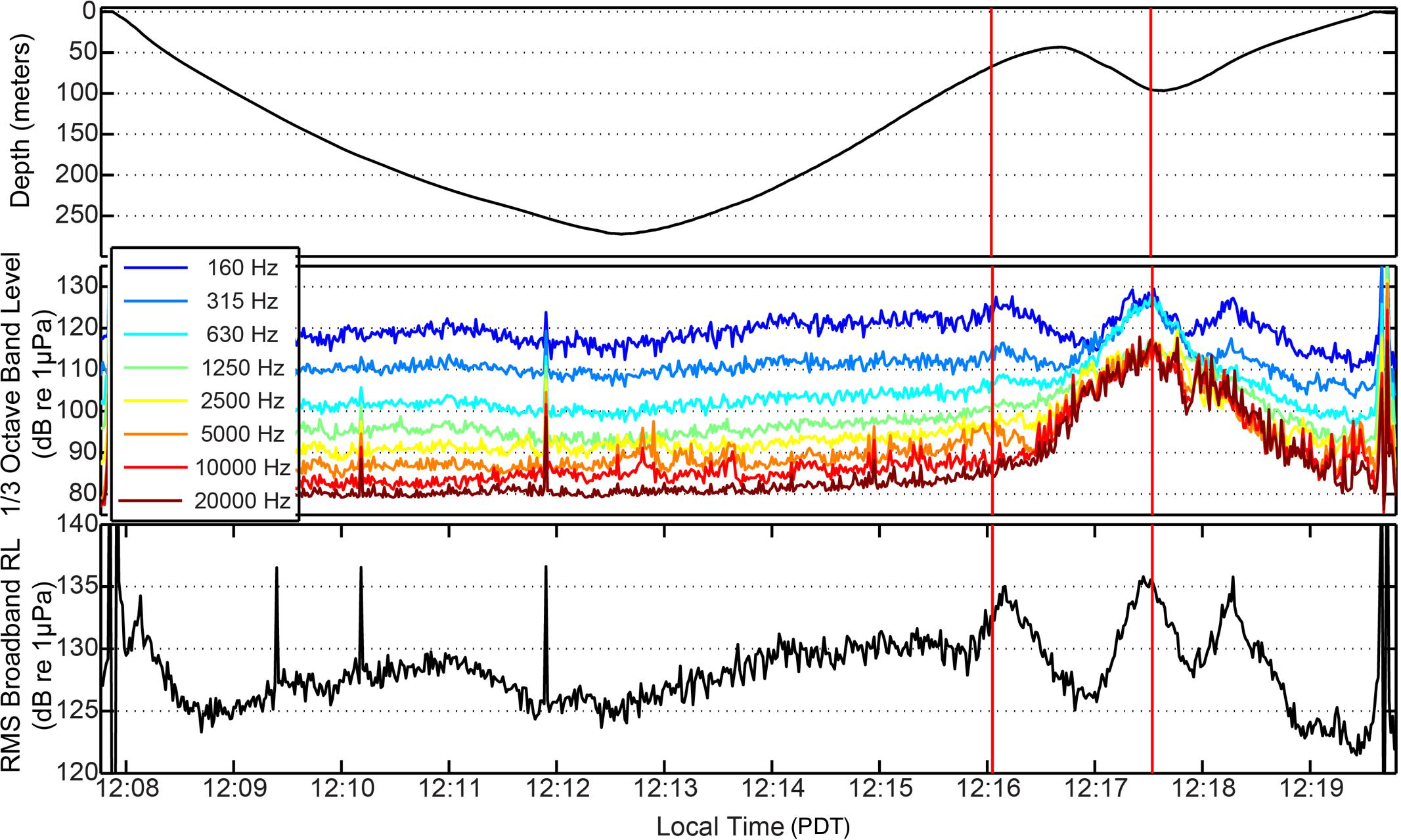
Figure 4. Received levels (RLs) recorded on the DTAG-3 during the passage and CPA of Mokihana. Both 1/3-octave band sound levels (middle panel) and broadband (RMS) measurements (bottom panel) are shown. The upper panel shows the depth of the tagged whale during the same time period. The first red vertical line indicates the onset of a behavioral response by the whale. The second red vertical line indicates the CPA with the Mokihana.
Discussion
The unique dataset from this case study provides a detailed account of the closest documented encounter between a large commercial vessel and a blue whale. The exact cues used to facilitate the successful avoidance in this close encounter case were unknown. However, contemporaneous data from multiple platforms (i.e., fine-scale kinematic, acoustic, movement, position, demographic, and long-term sighting history data) available in this study provided a comprehensive picture of the interaction, allowing us to explore the potential visual and acoustic cues available to the whale. It is likely that the observed behavioral response to the close ship passage resulted from some integration of these multi-modal indicators of close presence rather than any single parameter (e.g., maximum RL) driving the avoidance response.
The observed response behavior of the whale in this study occurred during an ascent from a deep non-lunge feeding dive when the whale aborted its ascent to the surface in order to descend back down to a deeper, and potentially safer depth, until the ship had passed overhead. There appeared to be no change in the direction of the whale as it traveled along the shelf edge perpendicular to the course of the ship. This mirrored the behavioral response previously described by a blue whale in McKenna et al. (2015). The whale also performed a 25-degree left-hand roll as the ship passed overhead.
The focal follow of the whale was a consistent part of the observation/tracking of this whale and lasted from 0736 (nearly 1 h prior to tagging) through 1436 (with a small number of follow-up observations through 1756). The only exceptions from this routine involving other types of approaches were well before or after the ship close approach and included approaches by the RHIB to deploy tags at 0848, an approach to conduct a unmanned aircraft system flight over the whale around 1000 (and ending by 1010), and two approaches to collect biopsy and fecal samples between 1340 and 1436. No obvious strong reactions were noted to these approaches (a potential acceleration was noted as a reaction to the biopsy collection at 1340). There were no sudden changes or close approaches to the tagged whale immediately before, during, and after the close approach with the Mokihana, allowing us to reliably detect changes during close encounters. Given that these approaches were not within an hour of the ship close approach and did not elicit a response, we are confident the specific and unusual observed response documented around the time of the ship CPA and described here is primarily related to the encounter with the Mokihana.
The cues whales use to detect the presence of a ship will likely influence how they respond and the amount of time they may have to react before a potential collision. Although cetacean vision is monochromatic, they do have adaptations for better underwater vision, including large, flattened eyeballs; enlarged pupils; and a tapetum lucidum, which translates to increased light intake and clearer images (Dawson, 1980; Mass and Supin, 2007). Deep-diving whales also have higher rhodopsin, a light-sensitive protein in the rod cells that confer greater sensitivity toward blue-shifted underwater light (Jacobs, 1993; Southall et al., 2002; Dungan et al., 2016). This suggests that in a clear ocean, whales could make use of any available light within the euphotic zone. In turbid waters, reduced visibility may increase the risk of ship strike; however, in our study, the Beaufort Sea State was reported as a 4, and the whale was 67.5 m from the surface. The whale may have been close enough to the surface to see the downwelling light blocked by the nearly 300-m cargo ship, similar to how they would assess prey distribution. Additionally, rolling 25 degrees, an uncommon response for blue whales near the surface (Segre et al., 2018), is suggestive of deliberate behavior, and would enhance panoramic vision (120–130° visual field) in multiple dimensions (Goldbogen et al., 2013a), allowing the whale to watch the ship pass overhead. Because cetacean vision functions in air and water (Supin et al., 2001), this whale also may have seen the ship approaching when the whale was at the surface.
At the time the whale initiated its response, there was only a minimal increase in the overall ship noise level above background levels (as detected on the tag) although there was a rapid increase in relative levels of high-frequency noise. This indicates that the whale may have reacted to the these changes in acoustic cues of the vessel’s proximity soon after they were available. However, the ship was only audible on the tag above background levels once it was within extremely close range (∼300 m). Additionally, the main source of noise – the propeller – is located at the stern of the ship, so at the maximum received sound level, hundreds of meters of ship had already passed overhead. This suggests that a whale ahead of a ship may have very little acoustic information to indicate its approach and therefore only extremely limited time to initiate an appropriate behavioral response. Several factors can affect the ability of whales to detect and locate the sounds of approaching ships, including acoustical shadowing if the propellers are located shallower than keel depth, masking of ship noise by ambient sound from other ships, and the Lloyd’s Mirror Effect whereby refraction of lower frequency sounds from the surface leads to extreme sound attenuation at shallow depths (Gerstein et al., 2005).
Additionally, the maximum RMS broadband received sound levels exceeded pre-ship sound levels by ∼10 dB, a value well below those associated with avoidance and diving behavioral responses of shallow-diving blue whales to active sonar sounds (see: Southall et al., 2019). While these have different contexts than continuous noise associated with vessels, the data are consistent with the observation that the response was not necessarily driven by an aversive reaction to a perceived loud sound. Rather, the increase in ship noise above ambient conditions, and other factors we were unable to measure (e.g., Doppler shifts indicating relative motion), were potentially integrated with visual information to indicate the close proximity of the ship to the whale that resulted in the observed response. However, as background ocean noise levels increase, particularly driven by greater shipping traffic (Ross, 1993; Andrew et al., 2002; Chapman and Price, 2011; Southall et al., 2018), it may prove to be even more difficult for a blue whale to detect acoustic cues in order to locate and avoid passing ships. If blue whales are not detecting acoustic cues, or the acoustic cues are below individual hearing thresholds, they must rely solely on visual detection, which greatly reduces the range that they can detect an oncoming ship.
The whale’s behavioral state at the time of the close encounter may have played a role in its behavioral response. Lunge feeding was not detected in the dive recorded by the DTAG-3 during the CPA. However, lunges were detected in dives before and after the CPA. The dive occurring during the CPA may have been part of a larger foraging bout or constituted traveling in search of a new prey patch. Behavioral state has been shown to influence the context-dependent behavioral response of tagged blue whales, including during playback experiments with ship noises and navy sonar (Goldbogen et al., 2013b; Southall et al., 2018, 2019). Feeding whales may be distracted (Chatterton, 1926; Horwood, 1981; Watkins, 1986) and thus be less capable of detecting – and, therefore, avoiding – approaching vessels. They may also ignore ships in favor of their current behavior (e.g., feeding, socializing, migrating) or due to habituation (Laist et al., 2001; Nowacek et al., 2004; Silber et al., 2010).
The avoidance of a collision between the tagged whale and large vessel may not have been solely due to the animal’s behavior. Specifically, the ship’s speed may have played a role by giving the whale enough time to respond. At the time of the close passage and onset of the observed behavioral response by the whale, the ship was going 11.3 knots. This ship had recently left the Precautionary Area of the SBC Traffic Separation Scheme. Matson, Inc., which owns the Mokihana, was participating in a vessel speed reduction trial incentive program, which aimed to slow ships in the SBC from 14–18 knots to 12 knots. In addition to reducing air pollution, slowing ships to 12 knots has been shown to greatly reduce the chances of a lethal ship strike (Vanderlaan and Taggart, 2007; Gende et al., 2011; Wiley et al., 2011; Conn and Silber, 2013; McKenna et al., 2015). The Mokihana had not yet picked up speed, which may have allowed the additional reaction time for the animal to arrest its ascent and avoid a potential collision. The behavioral action may not have been as effective if the vessel was traveling at greater speeds (McKenna et al., 2015), and the whale could have been struck at the surface or gotten close enough to the ship’s draft that the propeller suction effect created by the ship’s hydrodynamic flow could pull the whale toward the hull (Silber et al., 2010) resulting in a ship strike.
One of the hypotheses to arise from the research of McKenna et al. (2015) is that because the evolutionary history of blue whales did not include threats at the surface, whales have not developed an effective behavioral response strategy for this surface hazard. Our study confirms that there are some sensory cues available to the whale, but only at relatively close ranges (<300 m) and under certain oceanographic conditions. This may mean that even experienced individuals cannot always effectively adapt to the threat of shipping traffic. However, this may be further compounded by potential habituation to the presence of ships in important habitats. We know from the long sighting history of the tagged whale that it spent large amounts of time in high ship traffic areas, was exposed to military sonar, and was even involved in the capsizing of a small boat. The whale in this study was able to make last minute behavioral changes in response to the ship when it was already extremely close. However, this response may not be effective in all situations, making blue whales particularly vulnerable to ship strikes. The two key data points from our study – distance and acoustic cues (including RLs and frequency content) – will aid future models in determining when animals would need to respond to avoid being hit by a ship.
The combination of the distinct methodologies and technologies presented in this case study allowed for the collection of high-resolution behavioral information to examine a blue whale’s response during a close encounter with a large vessels. Not only has this filled in gaps in our current understanding of blue whale exposure to anthropogenic threats, which will contribute to more informed evaluation and mitigation strategies, but this study provides an example of how multiple methodologies can be combined to conduct behavioral studies in other highly mobile marine megafauna. Future work will examine close encounters from multiple whales to determine if certain contextual factors lead to a higher rate of behavioral response. This information can be used by managers to reduce the risk of exposure to ships or increase the chances of a successful evasion during a ship encounter.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
This study was carried out in accordance with the recommendations of Institutional Animal Care and Use Committee protocols (#AUP-6) and National Marine Fisheries Service Authorizations and Permits for Protected Species (#14534-2). The protocol was approved by the Institutional Animal Care and Use Committee protocols (#AUP-6) and National Marine Fisheries Service Authorizations and Permits for Protected Species (#14534-2).
Author Contributions
JC and BS conducted the field work and collected the data. AS and JC conceived of the presented idea. AS processed and analyzed the kinematic dive data, conducted the distance calculations, and wrote the draft manuscript. AA and BS conducted the acoustic analysis. JF created the whale pseudotrack track file. AS, AA, JC, JF, MM, and BS discussed the results and contributed to the final manuscript.
Funding
This research was funded by the Office of Naval Research (Grant Number N00014-13-1-0772 to JC). Research funding for the overall BRS study was provided by the United States Navy’s Living Marine Resources program and the Office of Naval Research. All tagging was conducted under National Marine Fisheries Service Authorizations and Permits for Protected Species permit #14534-2 and Cascadia Research Collective’s IACUC AUP-6.
Conflict of Interest
BS was employed by the company Southall Environmental Associates, Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Miche Capone for his programing assistance, which was essential for the distance calculations, and for his helpful comments on earlier drafts of the manuscript. We thank the entire SOCAL-BRS field team, including the crew of the M/V Truth. We also thank the staff at Cascadia Research for assisting with photo-identification matching, and Scripps Institution of Oceanography for providing us access to the AIS receiver on Santa Cruz Island.
References
Allen, A. N., Goldbogen, J. A., Friedlaender, A. S., and Calambokidis, J. (2016). Development of an automated method of detecting stereotyped feeding events in multisensor data from tagged rorqual whales. Ecol. Evol. 6, 7522–7535. doi: 10.1002/ece3.2386
Andrew, R. K., Howe, B. M., Mercer, J. A., and Dzieciuch, M. A. (2002). Ocean ambient sounds: comparing the 1960’s with the 1990’s for a receiver off the California coast. Acoust. Res. Lett. Online 3, 65–70. doi: 10.1121/1.1461915
Berman-Kowalewski, M., Gulland, F., Wilkin, S., Calambokidis, J., Mate, B., Cordaro, J., et al. (2010). Association between blue whale mortality and ship strikes along the California coast. Aquat. Mamm. 36, 59–66. doi: 10.1578/AM.36.1.2010.259
Cade, D. E., Friedlaender, A. S., Calambokidis, J., and Goldbogen, J. A. (2016). Kinematic diversity in rorqual whale feeding mechanisms. Curr. Biol. 26, 2617–2624. doi: 10.1016/j.cub.2016.07.037
Calambokidis, J. (2013). Updated Abundance Estimates of Blue and Humpback Whales off the US West Coast Incorporating Photo-Identifications From 2010 and 2011. Final Report Document PSRG-2013-13. Silver Spring, MD: NOAA.
Calambokidis, J., Barlow, J., Ford, J. K. B., Chandler, T. E., and Douglas, A. B. (2009). Insights into the population structure of blue whales in the eastern North Pacific from recent sightings and photographic identifications. Mar. Mamm. Sci. 25, 816–832. doi: 10.1111/j.1748-7692.2009.00298.x
Calambokidis, J., Steiger, G. H., Curtice, C., Harrison, J., Ferguson, M. C., Becker, E., et al. (2015). Biologically important areas for selected cetaceans within U.S. Waters – West Coast Region. Aquat. Mamm. 41, 39–53. doi: 10.1578/AM.41.1.2015.39
Carretta, J. V., Forney, K. A., Oleson, E., Martien, K., Muto, M. M., Lowry, M. S., et al. (2011). U.S. Pacific Marine Mammal Stock Assessments: 2010. Technical Memorandum, NMFS-SWFSC-476. La Jolla, CA: U.S. National Marine Fisheries Service.
Carretta, J. V., Wilkin, S. M., Muto, M. M., and Wilkinson, K. (2013). Sources of Human-Related Injury and Mortality for U.S. Pacific West Coast Marine Mammal Stock Assessments, 2007-2011. Washington, DC: U.S. department of commerce.
Chapman, N. R., and Price, A. (2011). Low frequency deep ocean ambient noise trend in the Northeast Pacific Ocean. J. Acoust. Soc. Am. 129, EL161–EL165. doi: 10.1121/1.3567084
Chatterton, E. K. (1926). Whalers and Whaling, the Story of Whaling Ships up to the Present Day. Philadelphia: JB Lippincott Co.
Conn, P. B., and Silber, G. K. (2013). Vessel speed restrictions reduce risk of collision related mortality for North Atlantic right whales. Ecosphere 4, 1–16. doi: 10.1890/ES13-00004.1
Croll, D. A., Tershy, B. R., Hewitt, R., Demer, D., Hayes, S., Fiedler, P., et al. (1998). An integrated approach to the foraging ecology of marine birds and mammals. Deep Sea Res. Part II Top. Stud. Oceanogr. 45, 1353–1371. doi: 10.1016/S0967-0645(98)00031-9
Dawson, W. W. (1980). “The cetacean eye,” in Cetacean Behavior, ed. L. M. Herman, (New York, NY: Wiley), 53–100.
DeAngelis, M., Fahy, C., and Cardaro, J. (2010). Report of a Workshop to Reduce Vessel Strikes of Large Whales in California, Long Beach, California, 19-20 May 2010. Silver Spring, MD: U.S. Department of Commerce.
Dungan, S. Z., Kosyakov, A., and Chang, B. S. (2016). Spectral Tuning of Killer Whale (Orcinus orca) Rhodopsin: evidence for positive selection and functional adaptation in a cetacean visual pigment. Mol. Biol. Evol. 33, 323–336. doi: 10.1093/molbev/msv217
Fiedler, P. C., Reilly, S. B., Hewitt, R. P., Demer, D., Philbrick, V. A., Smith, S., et al. (1998). Blue whale habitat and prey in the California Channel Islands. Deep Sea Res. Part II Top. Stud. Oceanogr. 45, 1781–1801. doi: 10.1016/S0967-0645(98)80017-9
Fletcher, S., Le Boeuf, B. J., Costa, D. P., Tyack, P. L., and Blackwell, S. B. (1996). Onboard acoustic recording from diving northern elephant seals. J. Acoust. Soc. Am. 100, 2531–2539. doi: 10.1121/1.417361
Friedlaender, A. S., Hazen, E. L., Goldbogen, J. A., Stimpert, A. K., Calambokidis, J., and Southall, B. L. (2016). Prey-mediated behavioral responses of feeding blue whales in controlled sound exposure experiments. Ecol. Appl. 26, 1075–1085. doi: 10.1002/15-0783
Friedlaender, A. S., Herbert-Read, J., Hazen, E. L., Cade, D. E., Calambokidis, J., Southall, B. L., et al. (2017). Context-dependent lateralized feeding strategies in blue whales. Curr. Biol. 27, R1206–R1208. doi: 10.1016/j.cub.2017.10.023
Gende, S. M., Hendrix, A. N., Harris, K. R., Eichenlaub, B., Nielsen, J., and Pyare, S. (2011). A Bayesian approach for understanding the role of ship speed in whale-ship encounters. Ecol. Appl. 21, 2232–2240. doi: 10.1890/10-1965.1
Gerstein, E. R., Blue, J. E., and Forysthe, S. E. (2005). “The acoustics of vessel collisions with marine mammals,” in Proceedings of OCEANS MTS/IEEE, Washington, DC, 1190–1197.
Goldbogen, J. A., Calambokidis, J., Friedlaender, A. S., Francis, J., DeRuiter, S. L., Stimpert, A. K., et al. (2013a). Underwater acrobatics by the world’s largest predator: 360° rolling maneuvers by lunge-feeding blue whales. Biol. Lett. 9:20120986. doi: 10.1098/rsbl.2012.0986
Goldbogen, J. A., Southall, B. L., DeRuiter, S. L., Calambokidis, J., Friedlaender, A. S., Hazen, E. L., et al. (2013b). Blue whales respond to simulated mid-frequency military sonar. Proc. R. Soc. Lond. B Biol. Sci. 280, 20130657. doi: 10.1098/rspb.2013.0657
Goldbogen, J. A., Calambokidis, J., Shadwick, R. E., Oleson, E. M., McDonald, M. A., and Hildebrand, J. A. (2006). Kinematics of foraging dives and lunge-feeding in fin whales. J. Exp. Biol. 209, 1231–1244. doi: 10.1242/jeb.02135
Heyning, J. E., and Dahlheim, M. E. (1990). “Strandings and incidental takes of gray whales,” Paper Presented at SC/A90/G2, International Whaling Commission Scientific Committee on the Assessment of Gray Whales, Cambridge.
Horwood, J. W. (1981). Results from the IWC IDCR minke whale marking and sighting cruise 1979/80. Rep. Int. Whal. Commn. 31, 287–315. doi: 10.1017/S0032247400003995
Jacobs, G. H. (1993). The distribution and nature of color vision among the mammals. Biol. Rev. 68, 413–471. doi: 10.1111/j.1469-185x.1993.tb00738.x
Johnson, M. P., and Tyack, P. L. (2003). A digital acoustic recording tag for measuring the response of wild marine mammals to sound. IEEE J. Ocean. Eng. 28, 3–12. doi: 10.1109/JOE.2002.808212
Kraus, S. D., Brown, M. W., Caswell, H., Clark, C. W., Fujiwara, M., Hamilton, P. K., et al. (2005). North Atlantic right whale in crisis. Science 309, 561–562. doi: 10.1126/science.1111200
Laist, D. W., Knowlton, A. R., Mead, J. G., Collet, A. S., and Podesta, M. (2001). Collisions between ships and whales. Mar. Mamm. Sci. 17, 35–75. doi: 10.1111/j.1748-7692.2001.tb00980.x
Mass, A., and Supin, Y. A. (2007). Adaptive features of aquatic adaptive features of aquatic mammals’. Eye Anat. Rec. 290, 701–715. doi: 10.1002/ar.20529
McKenna, M. F., Calambokidis, J., Oleson, E. M., Laist, D. W., and Goldbogen, J. A. (2015). Simultaneous tracking of blue whales and large ships demonstrates limited behavioral responses for avoiding collision. Endanger. Species Res. 27, 219–232. doi: 10.3354/esr00666
McKenna, M. F., Katz, S. L., Condit, C., and Walbridge, S. (2012a). Response of commercial ships to a voluntary speed reduction measure: are voluntary strategies adequate for mitigating ship-strike risk? Coast. Manag. 40, 634–650. doi: 10.1080/08920753.2012.727749
McKenna, M. F., Ross, D., Wiggins, S. M., and Hildebrand, J. A. (2012b). Underwater radiated noise from modern commercial ships. J. Acoust. Soc. Am. 131, 92–103. doi: 10.1121/1.3664100
Merchant, N. D., Fristrup, K. M., Johnson, M. P., Tyack, P. L., Witt, M. J., Blondel, P., et al. (2015). Measuring acoustic habitats. Methods Ecol. Evol. 6, 257–265. doi: 10.1111/2041-210X.12330
Monnahan, C. C., Branch, T. A., and Punt, A. E. (2014). Do ship strikes threaten the recovery of endangered eastern North Pacific blue whales? Mar. Mamm. Sci. 31, 279–297. doi: 10.1111/mms.12157
Nowacek, D., Johnson, M. P., and Tyack, P. L. (2004). North Atlantic right whales (Eubalaena glacialis) ignore ships but respond to alerting stimuli. Proc. R. Soc. Lond. B Biol. Sci. 271, 227–231. doi: 10.1098/rspb.2003.2570
Oleson, E. M., Calambokidis, J., Burgess, W. C., McDonald, M. A., LeDuc, C. A., and Hildebrand, J. A. (2007). Behavioral context of call production by eastern North Pacific blue whales. Mar. Ecol. Prog. Ser. 330, 269–284. doi: 10.3354/meps330269
R Core Team, (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Redfern, J. V., McKenna, M. F., Moore, T. J., Calambokidis, J., Deangelis, M. L., Becker, E. A., et al. (2013). Assessing the risk of ships striking large whales in marine spatial planning. Conserv. Biol. 2, 292–302. doi: 10.1111/cobi.12029
Rice, D. W. (1974). “Whales and whale research in the eastern North Pacific,” in The Whale Problem: A Status Report, ed. W. E. Schevill, (Cambridge, MA: Harvard Press), 170–195.
Rockwood, R. C., Calambokidis, J., and Jahncke, J. (2017). High mortality of blue, humpback and fin whales from modeling of vessel collisions on the U.S. West Coast suggests population impacts and insufficient protection. PLoS One 12:e0183052. doi: 10.1371/journal.pone.0183052
Segre, P. S., Cade, D. E., Calambokidis, J., Fish, F. E., Friedlaender, A. S., Potvin, J., et al. (2018). Body flexibility enhances maneuverability in the world’s largest predator. Integr. Comp. Biol. 59, 48–60. doi: 10.1093/icb/icy121
Silber, G. K., Slutsky, J., and Bettridge, S. (2010). Hydrodynamics of a ship/whale collision. J. Exp. Mar. Biol. Ecol. 391, 10–19. doi: 10.1016/j.jembe.2010.05.013
Simon, M., Johnson, M., Tyack, P., and Madsen, P. T. (2009). Behaviour and kinematics of continuous ram filtration in bowhead whales (Balaena mysticetus). Proc. R. Soc. Lond. B Biol. Sci. 276, 3819–3828. doi: 10.1098/rspb.2009.1135
Southall, B. L., DeRuiter, S. L., Friedlaender, A., Stimpert, A. K., Goldbogen, J. A., Hazen, E., et al. (2019). Behavioral responses of individual blue whales (Balaenoptera musculus) to mid-frequency military sonar. J. Exp. Biol. 222:jeb190637. doi: 10.1242/jeb.190637
Southall, B. L., Hatch, L., Scholik-Schlomer, A., Bergmann, T., Jasny, M., Metcalf, K., et al. (2018). Reducing noise from large commercial ships: progress and partnerships. Proc. Mar. Saf. Sec. Council 1, 58–65.
Southall, B. L., Moretti, D., Abraham, B., Calambokidis, J., DeRuiter, S. L., and Tyack, P. L. (2012). Marine mammal behavioral response studies in southern California: advances in technology and experimental methods. MTS J. 46, 48–59. doi: 10.4031/mtsj.46.4.1
Southall, B. L., Nowacek, D. P., Miller, P. J. O., and Tyack, P. L. T. (2016). Experimental field studies to measure behavioral responses of cetaceans to sonar. Endang. Species Res. 31, 293–315. doi: 10.3354/esr00764
Southall, K. D., Oliver, G. W., Lewis, J. W., Le Boeuf, B. J., Levenson, D. H., and Southall, B. L. (2002). Visual pigment sensitivity in three deep diving marine mammals. Mar. Mamm. Sci. 18, 275–281. doi: 10.1111/j.1748-7692.2002.tb01033.x
Supin, A. Y., Popov, V. V., and Mass, A. M. Vision (2001). “Vision in aquatic mammals,” in The Sensory Physiology of Aquatic Mammals, Vol. 1091 (Boston, MA: Springer), 229–284. doi: 10.1007/978-1-4615-1647-7
Vanderlaan, A. S. M., and Taggart, C. T. (2007). Vessel collisions with whales: the probability of lethal injury based on vessel speed. Mar. Mamm. Sci. 23, 144–156. doi: 10.1111/j.1748-7692.2006.00098.x
Watkins, W. A. (1986). Whale reactions to human activities in Cape Cod waters. Mar. Mamm. Sci. 2, 251–262. doi: 10.1111/j.1748-7692.1986.tb00134.x
Wiggins, S. M. (2003). Autonomous acoustic recording package (ARPs) for long-term monitoring of whale sounds. MTS J. 37, 13–22. doi: 10.4031/002533203787537375
Wiley, D. N., Thompson, M., Pace, I. I. I. R. M., and Levenson, J. (2011). Modeling speed restrictions to mitigate lethal collisions between ships and whales in the Stellwagen Bank National Marine Sanctuary, USA. Biol. Conserv. 144, 2377–2381. doi: 10.1016/j.biocon.2011.05.007
Williams, R., Gero, S., Bejder, L., Calambokidis, J., Kraus, S. D., Lusseau, D., et al. (2011). Underestimating the damage: interpreting cetacean carcass recoveries in the context of the Deepwater Horizon/BP incident. Conserv. Lett. 4, 228–233. doi: 10.1111/j.1755-263X.2011.00168.x
Keywords: ship strike, blue whale, near collision, active avoidance, behavioral response, perceptual cues
Citation: Szesciorka AR, Allen AN, Calambokidis J, Fahlbusch J, McKenna MF and Southall B (2019) A Case Study of a Near Vessel Strike of a Blue Whale: Perceptual Cues and Fine-Scale Aspects of Behavioral Avoidance. Front. Mar. Sci. 6:761. doi: 10.3389/fmars.2019.00761
Received: 30 March 2019; Accepted: 22 November 2019;
Published: 10 December 2019.
Edited by:
Joshua Nathan Smith, Murdoch University, AustraliaReviewed by:
Gregory K. Silber, Smultea Environmental Sciences, United StatesMichael James Williamson, King’s College London, United Kingdom
Copyright © 2019 Szesciorka, Allen, Calambokidis, Fahlbusch, McKenna and Southall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angela R. Szesciorka, angela@szesciorka.com
†Present address: Angela R. Szesciorka, Scripps Institution of Oceanography, UC San Diego, La Jolla, CA, United States