 Alessandra M. da Fonseca-Martins1,2,3
Alessandra M. da Fonseca-Martins1,2,3 Phillipe de Souza Lima-Gomes2
Phillipe de Souza Lima-Gomes2 Maísa Mota Antunes4Renan Garcia de Moura5
Maísa Mota Antunes4Renan Garcia de Moura5 Luciana P. Covre5,6Carolina Calôba7Vivian Grizente Rocha7Renata M. Pereira7Gustavo Batista Menezes4
Luciana P. Covre5,6Carolina Calôba7Vivian Grizente Rocha7Renata M. Pereira7Gustavo Batista Menezes4 Daniel Claudio Oliveira Gomes5
Daniel Claudio Oliveira Gomes5 Elvira M. Saraiva2*†
Elvira M. Saraiva2*† Herbert L. de Matos Guedes1,3,8*†
Herbert L. de Matos Guedes1,3,8*†- 1Laboratório de Imunofarmacologia, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
- 2Departamento de Imunologia, Laboratório de Imunobiologia das Leishmanioses, Instituto de Microbiologia Paulo de Góes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
- 3Departamento de Imunologia, Laboratório de Imunobiotecnologia, Instituto de Microbiologia Paulo de Góes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
- 4Center for Gastrointestinal Biology, Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Minas Gerais, Brazil
- 5Núcleo de Doenças Infecciosas, Universidade Federal do Espírito Santo, Vitória, Brazil
- 6Division of Medicine, University College London, London, United Kingdom
- 7Departamento de Imunologia, Laboratório de Imunologia Molecular, Instituto de Microbiologia Paulo de Góes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
- 8Laboratório Interdisciplinar de Pesquisas Médicas, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
Neutrophils play an important role in the outcome of leishmaniasis, contributing either to exacerbating or controlling the progression of infection, a dual effect whose underlying mechanisms are not clear. We recently reported that CD4+ and CD8+ T cells, and dendritic cells of Leishmania amazonensis-infected mice present high expression of PD-1 and PD-L1, respectively. Given that the PD-1/PD-L1 interaction may promote cellular dysfunction, and that neutrophils could interact with T cells during infection, we investigated here the levels of PD-L1 in neutrophils exposed to Leishmania parasites. We found that both, promastigotes and amastigotes of L. amazonensis induced the expression of PD-L1 in the human and murine neutrophils that internalized these parasites in vitro. PD-L1-expressing neutrophils were also observed in the ear lesions and the draining lymph nodes of L. amazonensis-infected mice, assessed through cell cytometry and intravital microscopy. Moreover, expression of PD-L1 progressively increased in neutrophils from ear lesions as the disease evolved to the chronic phase. Co-culture of infected neutrophils with in vitro activated CD8+ T cells inhibits IFN-γ production by a mechanism dependent on PD-1 and PD-L1. Importantly, we demonstrated that in vitro infection of human neutrophils by L braziliensis induced PD-L1+ expression and also PD-L1+ neutrophils were detected in the lesions of patients with cutaneous leishmaniasis. Taken together, these findings suggest that the Leishmania parasite increases the expression of PD-L1 in neutrophils with suppressor capacity, which could favor the parasite survival through impairing the immune response.
Importance
Neutrophils outnumber leukocytes in healthy human blood, rapidly migrate to infected or inflamed sites and have powerful mechanisms to eliminate pathogens. These cells secrete cytokines and chemokines critical for the initiation and amplification of the inflammatory response. Neutrophils also have the ability to modulate adaptive immune cells through a variety of receptors, including the programmed death ligand-1 (PD-L1), a cell surface protein that suppresses T cell activation. We report that Leishmania amazonensis, which can cause cutaneous leishmaniasis, severe anergic diffuse cutaneous and even visceral leishmaniasis, upon interaction with murine and human neutrophils induces the expression of PD-L1. Neutrophils expressing PD-L1 were observed in the ear lesions and draining lymph nodes of infected mice and in human cutaneous leishmaniasis biopsies. Our findings suggest that Leishmania could modulate PD-L1 expression in neutrophils, weakening the immune response to favor its survival, which opens up new possibilities of targeting PD-L1 for therapy.
Introduction
Neutrophils, the most abundant white blood cells in the human circulation, play a crucial role in eliminating pathogens by multiple mechanisms, and participating in the development of the inflammatory reaction. Thus, neutrophils are key components of the innate immune response and in the elimination of infectious agents. They are recognized for their broad defense repertoire that includes the production of reactive oxygen species, phagocytosis, degranulation and the release of the antimicrobial neutrophil extracellular traps (1, 2).
Within the inflammatory environment, neutrophils interact with other immune cells, and secrete cytokines and chemokines critical for the development and establishment of the necessary conditions required for the interface with the adaptive immune response (3, 4). On the other hand, these cells can also favor the progression of various diseases, such as rheumatoid arthritis, systemic lupus erythematosus, and cystic fibrosis, as well as sepsis, HIV-1 infection, and malaria (5–8).
Neutrophils may also present opposing roles in infections caused by Leishmania sp. In fact, it has been shown that neutrophils can both exacerbate the disease or protect the host, an outcome that is dependent on a fine balance pertaining to the Leishmania species and host genetic background (9).
Furthermore, neutrophils are endowed with the ability to induce T cell suppression (10). Analyses of patients with visceral leishmaniasis (VL) showed an increased frequency of CD15+ neutrophils expressing high levels of arginase, an enzyme associated with immunosuppression, which, after successful treatment, returned to basal levels (11). In addition, co-culture of low-density HLA-DR+-neutrophils and lymphocytes, both from VL patients, with Leishmania antigen increased the expression of programmed death ligand-1 (PD-L1, also called B7-H1 and CD274) in neutrophils, and of the programmed death receptor 1 (PD-1, also called CD279) in lymphocytes. These findings led to the hypothesis that low-density HLA-DR+ neutrophils may be involved in promoting T-cell exhaustion (12).
PD-1 is a receptor found in Natural Killer cells, T cells, B cells and activated monocytes (13–15), while the PD-L1 ligand can be found in neutrophils, B cells, dendritic cells, macrophages, mesenchymal stem cells and in non-hematopoietic cells, such as epithelial cells, muscle cells and hepatocytes (16, 17). The function of this pathway has been widely studied revealing a dichotomous activity depending on the model in which it is applied, where often it can inhibit T cell proliferation and cytokine production or increase T cell activation (18, 19). Thus, the formation of the PD-1/PD-L1 complex can be seen as a mechanism used by pathogens to evade the immune response or as a possible regulator of tissue damage mediated by the immune system (20–22). The dysfunctional state characterized by increased expression of inhibitory receptors, like the PD-1/PD-L1 complex, and by progressive loss of function, is commonly known as cellular exhaustion (23).
The use of immunotherapy to reverse dysfunction due to T-cell exhaustion has been used successfully to treat several tumors (24, 25) and also in viral and parasitic infections (26, 27). Concerning leishmaniasis, we recently reported that mice infected with L. amazonensis presented high expression of PD-1 in CD4+ and CD8+ T cells, and PD-L1 in dendritic cells, and that treatment with anti-PD-1 or anti-PD-L1 antibodies reduced the parasite load and increased IFN-gamma production by CD4+ and CD8+ T cells, thus, reversing cell suppression (28).
It was demonstrated that neutrophils depletion using 1A8 increased T cells response against Leishmania (29). Here, we aimed to investigate whether the PD-L1 expression in murine and human neutrophils could be modulated after in vitro interaction with Leishmania, and identified PD-L1-expressing neutrophils in murine and human cutaneous lesions. We also evaluated in vitro the participation of PD-L1 expression by neutrophils in suppressing IFN-γ production by T cells.
Materials And Methods
Experimental Animals
Female BALB/c mice and C57BL/6, 6-8 weeks old, from the Central Bioterium (Centro de Ciências da Saúde – Universidade Federal do Rio de Janeiro, Brazil), were housed in ventilated mini-isolators (Alesco, Brazil) and kept under controlled temperature and light conditions. All animal experiments were performed in strict accordance with the Brazilian animal protection law (Lei Arouca, Number: 11.794/08) of the National Council for the Control of Animal Experimentation (CONCEA, Brazil). The protocol was approved by the Committee for Animal Use of the Universidade Federal do Rio de Janeiro (Permit Number: 161/18).
Parasite Culture
For in vivo infection, L. amazonensis promastigotes (MHOM/BR/75/Josefa) were used. Parasites were first obtained from the macerated lesions of infected BALB/c mice and then grown at 26°C in M-199 medium (Cultilab) with 20% heat inactivated fetal bovine serum (FBS, Cultilab). Parasites were used for the experimental infections until the fifth passage in culture.
For in vitro infection, L. amazonensis promastigotes (RAT/BA/74/LV78) and L. braziliensis (MHOM/BR/2005/RPL5) were cultured at 26°C in Schneider medium (Invitrogen) with 20% FBS and 50 µg/mL gentamicin (Sigma). The parasites were used in assays until the fifth passage.
Amastigotes of L. amazonensis (RAT/BA/74/LV78) and L. braziliensis (MHOM/BR/2005/RPL5) were obtained from a culture of promastigotes maintained at 32°C in Grace’s medium (Invitrogen), pH 5.3 supplemented with 20% FBS and 25 µg/ml gentamicin. The amastigotes were used until the third passage in culture.
Recruitment and Isolation of Murine Peritoneal Neutrophils
Neutrophils were recruited in BALB/c or C57BL/6 mice 3 h after intraperitoneal injection of 1 ml of 9% casein solution (Sigma). Mice were euthanized and the peritoneal cavity was washed with RPMI 1640 (Sigma) without serum at room temperature. A fraction of the obtained cells was first characterized by flow cytometry using anti-Ly6G-PerCP (murine, eBioscience) revealing >95% Ly6G+ cells (neutrophils). Remaining cells were centrifuged at 400 g for 5 min, resuspended in RPMI and used in the following assays.
Purification of Murine Neutrophils From Bone Marrow
Bone marrow cells were flushed from the femur and tibia of BALB/c mice with RPMI/10% FBS into a 15 ml conical tube through a 100 μm cell strainer, and then were centrifuged at 400 g for 7 min at 24°C. The cell pellet was resuspended in RPMI/10% FBS and centrifuged at 1500 g for 30 min at 24°C in a Percoll gradient (100%, 72%, 65% and 58% v/v GE Healthcare). Neutrophils collected at the interface of 65% and 72%, were washed with PBS at 400 g for 10 min at 15°C, resuspended in RPMI and used in the following assays. Cell yield was ≥80% Ly6G+ cells (neutrophils) as analyzed by flow cytometry using anti-Ly6G-PerCP (murine, eBioscience).
Isolation of Dermal Cells From Mouse Ears
Mice were infected intradermally in the right ear with 2x106 stationary-phase promastigotes of L. amazonensis in 20 µl PBS. After 18 h, 15 days and 60 days post-infection, the ears were collected. Control was performed with ears from naïve mice. The dermal sheets were opened and added to the wells of a 24-well plate in DMEM (Sigma) with 1% penicillin/streptomycin and 100 µg/ml each of collagenase I and II (Sigma), followed by 90 min incubation at 37°C. The tissue was then macerated with a tissue mixer for 3 min, filtered through a 70 µm filter and washed with PBS at 400 g for 5 min at 15°C. Cells were resuspended in RPMI/10% FBS for further use.
Isolation of Cells From the Lymph Nodes Draining the Infected Lesion
Mice were infected intradermally in the right footpad with 2x106 L. amazonensis stationary-phase promastigotes in 20 µl PBS. After approximately 60 days post-infection, draining lymph nodes were removed, individually macerated and the cell suspensions were centrifuged at 400 g for 5 min at 4°C. The cell pellet was resuspended in 2 ml RPMI/10% FBS for further use.
Purification of Human Blood Neutrophils
Human neutrophils from healthy blood were obtained by centrifugation on Ficoll Histopaque density gradient (Sigma-Aldrich), followed by hypotonic lysis (155 mM NH4Cl, 10 mM KHCO3, 0.1 mM EDTA, pH 7.4) to remove red blood cells. Isolated neutrophils (≥95% of the cells) were washed in PBS and resuspended in RPMI. All the procedures dealing with human blood were performed in accordance with the guidelines of the Research Ethics Committee (Hospital Universitário Fraga Filho, UFRJ, Brazil), protocol number: 4261 015400005257.
In Vivo Infection
BALB/c mice were infected with 2x106 L. amazonensis stationary-phase promastigotes in 20 µl PBS, either intradermally in the right ear or subcutaneously in the right footpad. The lesions were followed weekly for around two months by measuring the thickness with a pachymeter.
Leishmania-Neutrophil Interaction In Vitro
Parasites were washed twice with PBS and incubated with 0.5 µM CFSE (carboxyfluorescein succinimidyl ester - Invitrogen) at 37°C. After 15 min, RPMI/20% FBS was added, and parasites incubated for a further 15 min on ice, followed by three washes with PBS, and resuspension in PBS. Murine neutrophils were infected with CFSE-labeled promastigotes or amastigotes of L. amazonensis (cell:parasite-MOI 1: 10) for 4 h at 35°C and human neutrophils were infected with CFSE-labeled promastigotes or amastigotes of L. amazonensis or L. braziliensis (MOI 1:5 or 1:10) for 4 h at 35°C.
Cell Staining for Flow Cytometry
Cells from lymph nodes (5x105) or ear homogenates (1x106) were washed with PBS at 400 g for 5 min at 4°C and blocked with Human FcX (BioLegend) for 15 min, followed by staining with the antibody cocktail for 30 min at 4°C. Cells were then washed with a cytometry buffer (PBS with 5% FBS) at 400 g for 5 min and 4°C), then fixed with 4% formaldehyde (Sigma) for 15 min at 4°C. Cells were washed and resuspended in the cytometry buffer and stored in the dark at 4°C until acquisition. The following antibodies were used: anti-PD-L1-APC, anti-CD10-APC-780 (human, eBioscience); anti-CD45-APCcy7, anti-CD11b-FITC, anti-CD11b-PE, anti-CD11b-Pecy7, anti-Ly6G-PerCP, anti-Ly6G-FITC and anti-PD-L1-APC (murine, eBioscience). Acquisition of events (lymph node, 100,000 events; ear, all cells) was performed on a BD FACSAria™. The gate strategy was performed based on the selection of cell size (FSC) and composition (SSC). After identifying the main population, a gate of FSC-A (area) and FSC-H (weight) was used, where cellular doublets were excluded. Gates for positive events were established through Fluorescence Minus One (FMO) control. The data analyzes were performed using the FlowJo software.
Imaging PD-L1 Expression by Intravital Microscopy
L. amazonensis ear-infected BALB/c mice (naïve-not infected; control with saline; 1 h, 7 days and 15 days and 60 days post-infection) were injected on the ear with 8-12 µl/animal of anti-PD-L1-APC and Ly6G-FITC antibodies. After 2 h, mice were anesthetized with ketamine and xylazine intraperitoneally. Images were obtained using a Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x. Analysis was performed using Volocity 6.3 software (PerkinElmer).
CD8+ T Cells Activation And Culture In Vitro
Spleens were harvested from 6 to 8-week-old mice. Naïve CD8+ T cells (CD44lo CD62Lhi) T cells were purified (>95% purity) by negative selection (Dynabeads™ Untouched™ Mouse CD8 Cells Kit, Invitrogen) from RBC-lysed single-cell suspensions from spleen followed by cell sorting. For stimulation, purified CD8+ T cells were plated at 106 cells/ml in 24-well plates precoated with 0,3 mg/mL goat anti-hamster and coated with anti-CD3 (clone 2C11) and anti-CD28 (clone 37.51) (1 μg/mL), as previously described (30, 31). After, 48 h, cells were removed from the TCR signal and re-cultured by diluting 1:2 in media (DMEM (11995-065, Gibco) supplemented with 10% FBS (26140079, Gibco), 1x Penicillin/Streptomycin (15140-122, Gibco), 1x L-Glutamine (25030-081, Gibco), 1x MEM Vitamins (11120-052, Gibco), 1x Sodium Pyruvate (11360-070, Gibco), 1x Essential Amino Acids (M5550, Sigma),1x Non-Essential Amino Acids (11140-050, Gibco), 10 mM HEPES (15630-080, Gibco) and 50 μM β-Mercaptoethanol (M3148, Sigma), containing 200 U/ml final concentration of recombinant murine IL-2 (Peprotech). Every 24 h, cells were diluted 1:2 with fresh media containing IL-2. On day 5, cells were stained with anti-CD25-PE, anti-CD44-FITC, anti-CD62L-APC and anti-CD127-PEcy7 antibodies (Biolegend) for phenotypic analysis and were assessed by flow cytometry.
Coculture of Murine Neutrophils and Activated CD8+ T Cells
C57BL/6 neutrophils (5x105) recruited to the peritoneum with casein were infected with promastigotes of L. amazonensis (cell:parasite-MOI 1:5) by 4h. After that, we added 5 µg/ml anti-PD-1 (CD279, clone RMP1-14, catalog # BE0146, Bioxcell) or anti-PD-L1 (BMS-936559, Bristol-Myers Squibb) or isotype (IgG2a, SouthernBiotech) for 15 min in the culture. Following, activated CD8+ T cells were added (5x105) to the culture and the media was supplemented with 200 U/ml recombinant murine IL-2 (Peprotech) for 18h. The supernatants were collected and IFN-gamma quantified by specific ELISA using a standard protocol (Peprotech), detection limits are 12 - 3000 pg/ml.
Skin Biopsies and Study Subjects
Punch biopsies (8 mm in diameter) from the border of skin lesions were obtained from 7 untreated patients with cutaneous leishmaniasis (CL) attended at the University Hospital (HUCAM) of Universidade Federal do Espírito Santo, Brazil. The diagnosis of CL was based on clinical criteria including differential diagnosis, skin lesion analysis and parasite identification by microscopy. In addition, all patients in this study tested positive for the PCR/restriction fragment length polymorphism of L. braziliensis and reported no prior infections or treatment. The CL group consisted of 5 males and 2 females with illness duration ranging from 30 to 120 days, lesion sizes ranging from 50–200 mm2 and 1.42± 0.53 lesion per patient. All patients involved in this study progressed to clinical cure after Meglumine antimoniate treatment. Control skin punch biopsy specimens from 12 healthy volunteers living in a non-endemic area with no history of leishmaniasis were also obtained. Biopsy specimens were frozen in OCT compound (Sakura). Sections of 6 µM were obtained longitudinally to expose all skin layers and were placed on poly-L-lysine-coated slides (Star Frost®). Tissues were then fixed in acetone and ethanol and stored at -80°C until use. All participants (patients and healthy volunteers) were seronegative for HIV, HBV and HCV infections, and had no history of chemotherapy, radiotherapy or treatment with immunosuppressive medications within the last 6 months. They provided written informed consent, and study procedures were performed in accordance with the principles of the Declaration of Helsinki. This study was registered with the HUCAM Ethical Committee under the number 735.274.
Immunofluorescence Staining and Analysis of Human Tissues
Briefly, sections were hydrated with PBS, blocked with a 1% bovine serum albumin (BSA- Sigma) solution for 20 min and incubated with the following primary antibodies: anti-elastase (1:400; US1481001; Merk KGaA) or anti-PD-L1 (1:200; clone: ABM4E54; ab210931; Abcam) overnight at 4°C. Afterwards, slides were washed with PBS and incubated with goat anti-rabbit Alexa Fluor 488 (1:400; A11008) or goat anti-mouse Alexa Fluor 568 (1:400; A21124 or A21134) secondary antibodies (Thermo Fischer Scientific) for 1 h at room temperature. Slides were mounted with Fluoroshield Mounting Medium with DAPI (Abcam, ab104139) and analyzed by manually selecting regions of interest (ROI) using the Chromoplex Staining Detection system (Leica Biosystems). Cell and marker frequencies were defined according to the evaluated marker and cell nuclei of each ROI. Slides were imaged with a 20x objective (200× magnification) on a fluorescence microscope (Leica DMi8) with a 710 Metahead (Zeiss) by z-stack tile-scans and counted manually by using computer-assisted image analysis (National Institutes of Health Image Software ImageJ 1.52j; https://imagej.nih.gov/ij/).
Data Analysis
Results are expressed as mean ± SEM with confidence level p ≤ 0.05. For lesion development analysis, a two-way ANOVA with Bonferroni post-test was used. For multiple comparisons, a one-way ANOVA followed by Tukey pairing was performed. Cell analyzes were performed using the paired Student’s t test or Mann-Whitney test (indicated p value in the graph and legend). Skin biopsies were analyzed using Student’s t test with Welch correction or Mann-Whitney test. Data analysis was performed using GraphPad Prism® 8.00 software.
Results
PD-L1 Expression on Neutrophils Upon Interaction With Leishmania
Initially, we tested whether casein-recruited peritoneal neutrophils interacted equally with CSFE-labeled promastigotes and amastigotes of L. amazonensis. Our results evidenced that neutrophils internalized both parasite stages at similar levels (Figure 1A). Next, we analyzed the expression of PD-L1 on the CSFE+ neutrophils, which were the neutrophils that had internalized the parasites. We observed that uninfected neutrophils (medium) constitutively expressed PD-L1 at low levels, while both forms of the parasite induced PD-L1 expression in the infected casein-recruited peritoneal neutrophils (Figure 1B). It is important to note that within the neutrophil population, PD-L1 expression in cells that did not internalize the parasites (bystander CSFE- neutrophils) was not augmented, expressing similar PD-L1 levels to the control (Figure 1C). Similarly, we evaluated the parasite interaction with neutrophils isolated from the bone marrow. Amastigotes were internalized significantly more than promastigotes by these neutrophils (Figure 1D). Again, internalization of both parasite forms significantly increased PD-L1 expression in relation to the uninfected control (Figure 1E). Interestingly, in the bone marrow neutrophils, amastigotes induced significantly more PD-L1 expression than promastigotes (Figure 1E). Like the casein-recruited neutrophils, constitutive PD-L1 expression of bystander bone marrow neutrophils, was not modulated by the presence of either form of the parasite (Figure 1F). Comparing the levels of PD-L1 expression of the peritoneal and bone marrow neutrophils, our results demonstrated that PD-L1 expression was higher in peritoneal neutrophils, independent of the parasite infection (Figure S1), suggesting that the inflamed milieu or the maturation stage of the neutrophils could influence PD-L1 expression.

Figure 1 Increased expression of PD-L1 in murine and human neutrophils exposed to challenge with L. amazonensis. BALB/c neutrophils (5x105) recruited to the peritoneum with casein, or purified from bone marrow, and human neutrophils (5x105) from healthy donors were incubated with CFSE-stained promastigotes (Pro) and amastigotes (Ama) of L. amazonensis (murine 1:10; human 1:5), for 4 h. Control was performed with neutrophils on medium. Cells were then analyzed by flow cytometry using Ly6G+-Percp (neutrophils), CFSE+ (L. amazonensis) and PD-L1+-APC. (A) Percentage of peritoneal neutrophils infected with CFSE-labeled parasites. (B) Percentage of PD-L1+ peritoneal neutrophils upon interaction with parasites. (C) Percentage of PD-L1 expression on CFSE- (bystander) and CFSE+ casein-recruited neutrophils. (D) Percentage of bone marrow (BM) neutrophils infected with CFSE-labeled parasites. (E) Percentage of PD-L1+ bone marrow neutrophils upon interaction with parasites. (F) Percentage of PD-L1 expression on CFSE- and CFSE+ bone marrow neutrophils. (G) Percentage of human neutrophils infected with CFSE-labeled parasites. (H) Percentage of PD-L1+ human neutrophils upon interaction with parasites. (I) Percentage of PD-L1 expression on CFSE- and CFSE+ human neutrophils. Data are mean ± SEM (N=10-11) and human donors (N=7). Each point represents a donor. *p < 0.01, **p < 0.001, ***p < 0.001.
We next tested the interaction of human neutrophils with promastigotes and amastigotes of L. amazonensis (MOI 1:5). Unlike mouse neutrophils, human neutrophils internalized promastigotes more than the amastigotes (Figure 1G). However, PD-L1 induction occurred similarly to murine neutrophils, being significantly more expressed after interaction with amastigotes (Figure 1H). Interestingly, the PD-L1 constitutive expression (medium) in human neutrophils (Figure 1I) was lower than in mouse peritoneal (Figure 1C) and bone marrow (Figure 1F) neutrophils. Surprisingly, a reduction in PD-L1 expression was observed after human neutrophil infection with promastigotes and amastigotes at a higher MOI, although maintaining the same pattern of higher PD-L1 expression induced by amastigotes in comparison to promastigotes (Figure S2). Together, these results suggest that PD-L1 expression in neutrophils is modulated upon parasite interaction.
PD-L1 Expression in Cells From Ear Lesions of BALB/c Mice Infected With L. amazonensis
Following our observations of neutrophils in vitro, we assessed whether cells present at the lesion site in vivo would also express PD-L1 (Figure S10). As the peak of neutrophil infiltration has been reported to occur at 12 h after Leishmania infection in mice (29), we started by analyzing PD-L1 expression at this time-point. Our data shows increase in the percentage of Ly6G+ cells in infected group, but, while there is an increase in the number of neutrophils, it was not statistically different from the other groups (Figures 2A–C). Although the number of PD-L1+Ly6G+ increased in the infected group (Figure 2F), no differences were observed in the frequency of PD-L1+Ly6G+ or in the MFI between the groups (Figures 2D, E, G). Evaluating the same parameters after 18 h post infection, although the frequency of CD11b+ cells was a little higher in the infected ear, it was not significantly different from the naïve (non-infected) control; a similar observation was noted regarding the numbers of CD11b+ cells in naïve and infected mouse ears (Figures S3A, B and S4A). In addition, no differences were observed between the ears of naïve and infected mice in the frequency and number of Ly6G+CD11b+ cells (Figures S3C, D and S4B) and the expression of PD-L1 in the Ly6G+ cells (Figures S3E–H).
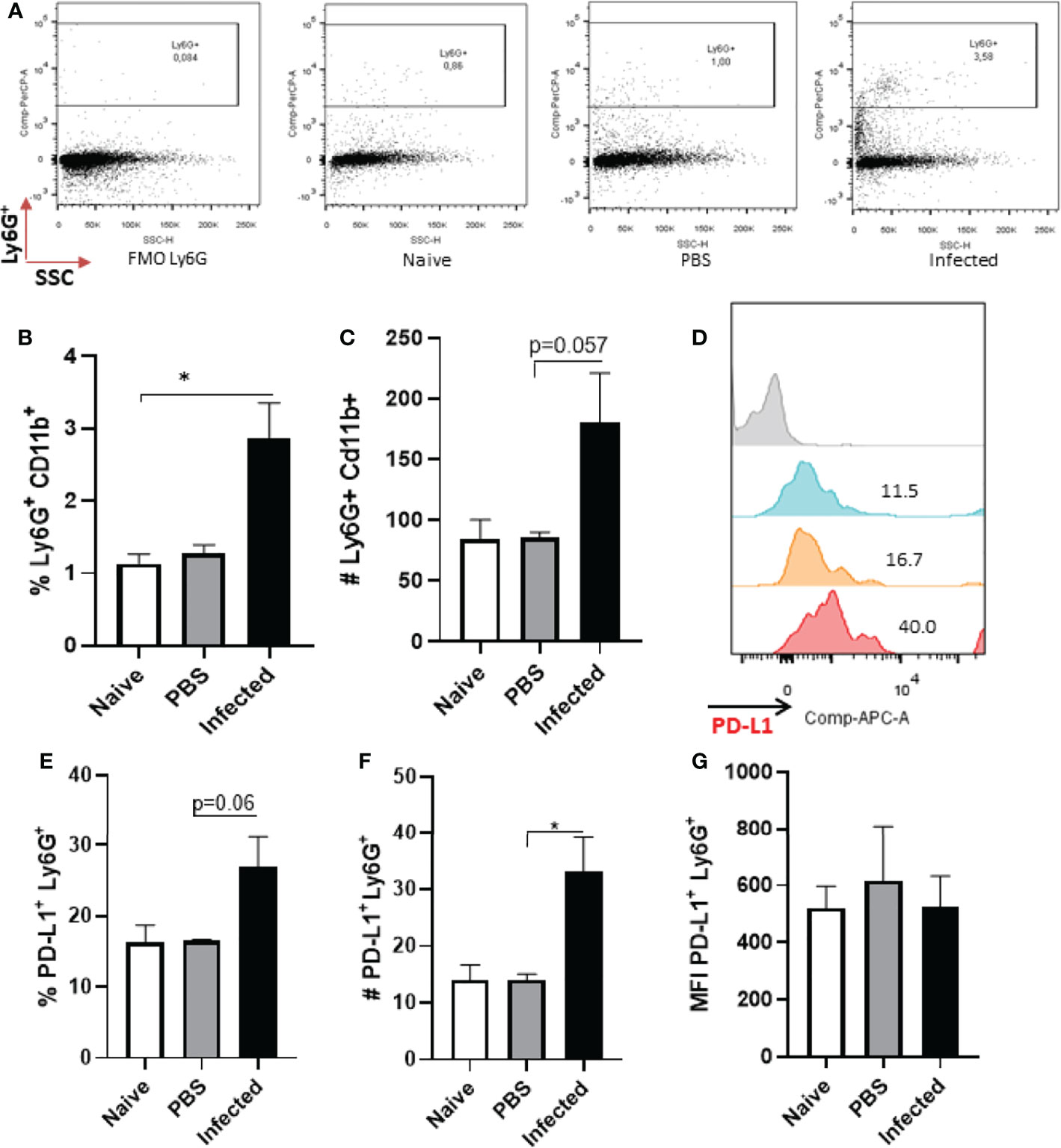
Figure 2 Analysis of murine ear neutrophils after 12h of infection. Cells were collected from L. amazonensis-infected ears after 12h then submitted to flow cytometry. Controls were performed with uninfected (naïve) and PBS injected mice. (A) Dot plot showing Ly6G+ cells (Ly6G-PerCP, SSC). (B) Percentage of Ly6G+ CD11b+ cells. (C) Number of Ly6G+ CD11b+ cells, p=0.057 (Mann-Whitney). (D) Histogram of PD-L1 expression. Grey = Fluorescence minus one control (FMO) PD-L1, Blue = Naïve, Orange = PBS and Red = Infected. (E) Percentage of PD-L1+ Ly6G+ cells, p=0,06 (Mann-Whitney). (F) Number of PD-L1+ Ly6G+ cells, p=0.06. (G) MFI of PD-L1+ Ly6G+. Data are mean ± SEM from cells of individual mice (N = 4-7 mice/group). *p < value 0,0323.
A second wave of neutrophils has been described to migrate to the infection site after 15 days (29), therefore we also evaluated PD-L1 expression at this time-point. Our results showed no statistical difference in the frequency of neutrophils between naïve and Leishmania-infected ear lesions (Figure 3A), however, a significant increase in the number of neutrophils was observed in infected mice compared with the naïve control (Figure 3B). Concomitantly, increased PD-L1 expression was detected in infected mice (Figures 3C, D, F) as well as an increased number of PD-L1-expressing neutrophils (Figure 3E).
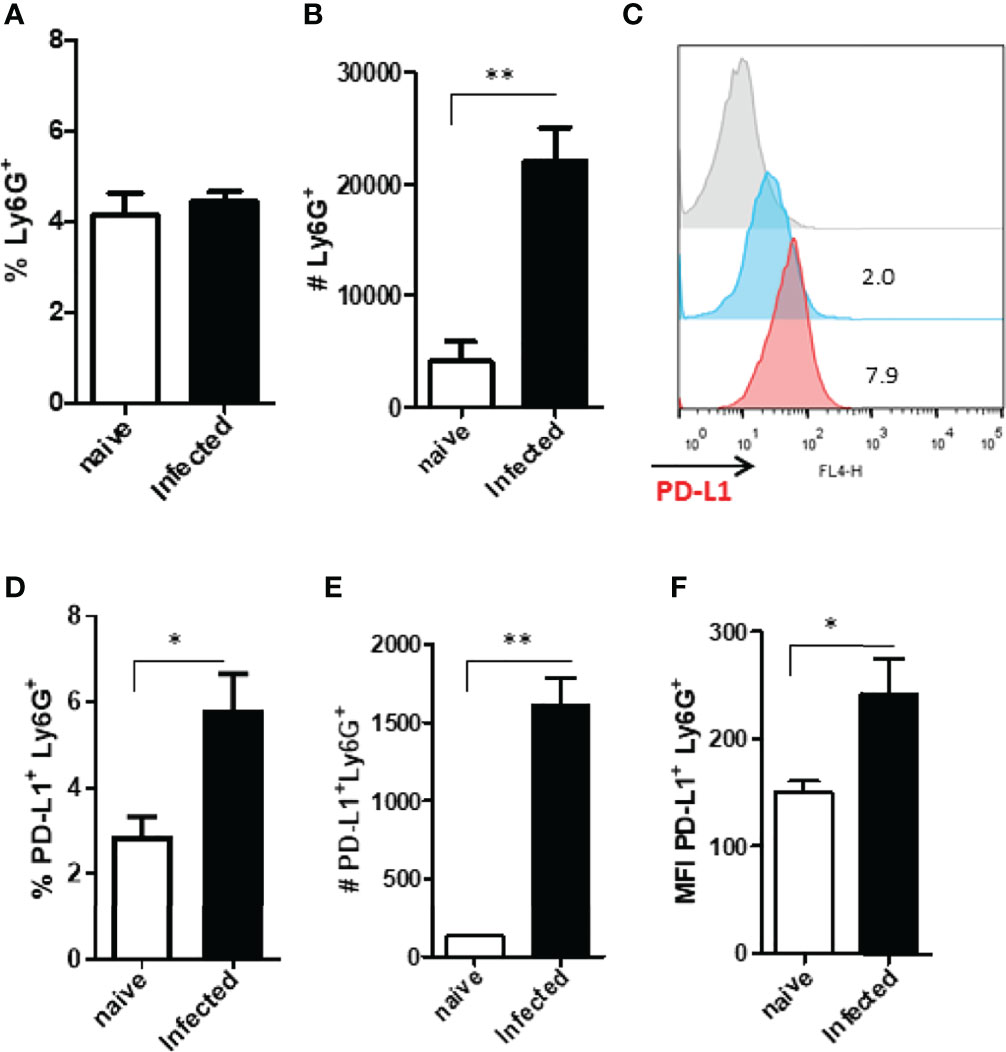
Figure 3 Analysis of murine ear neutrophils after 15 days of infection. Cells were collected from L. amazonensis-infected ears after 15 days then submitted to flow cytometry. Controls were performed with uninfected mice (naïve). (A) Percentage of Ly6G+ cells. (B) Number of Ly6G+ cells. (C) Histogram of PD-L1 expression. Grey = Fluorescence minus one control (FMO) PD-L1+, Blue = Naïve and Red = Infected. (D) Percentage of PD-L1+ Ly6G+ cells. (E) Number of PD-L1+ Ly6G+ cells. (F) MFI of PD-L1+ Ly6G+. Data are mean ± SEM from cells of individual mice (N = 4-5/group). *p < 0.03, **p < 0.005.
The analysis of cells from infected animals in the chronic phase, after 60 days of infection, displayed an increase in the neutrophil population (Figures 4A–C and S5). The expression of PD-L1 on neutrophils from 60-day lesions followed the same pattern with increased expression of PD-L1 in the cells from the infected animals (Figures 4D–G). These data show that as the lesion progresses, there is an increase in the population of neutrophils that express PD-L1, and this strengthens our hypothesis that a constant Leishmania stimulus promotes this induction.
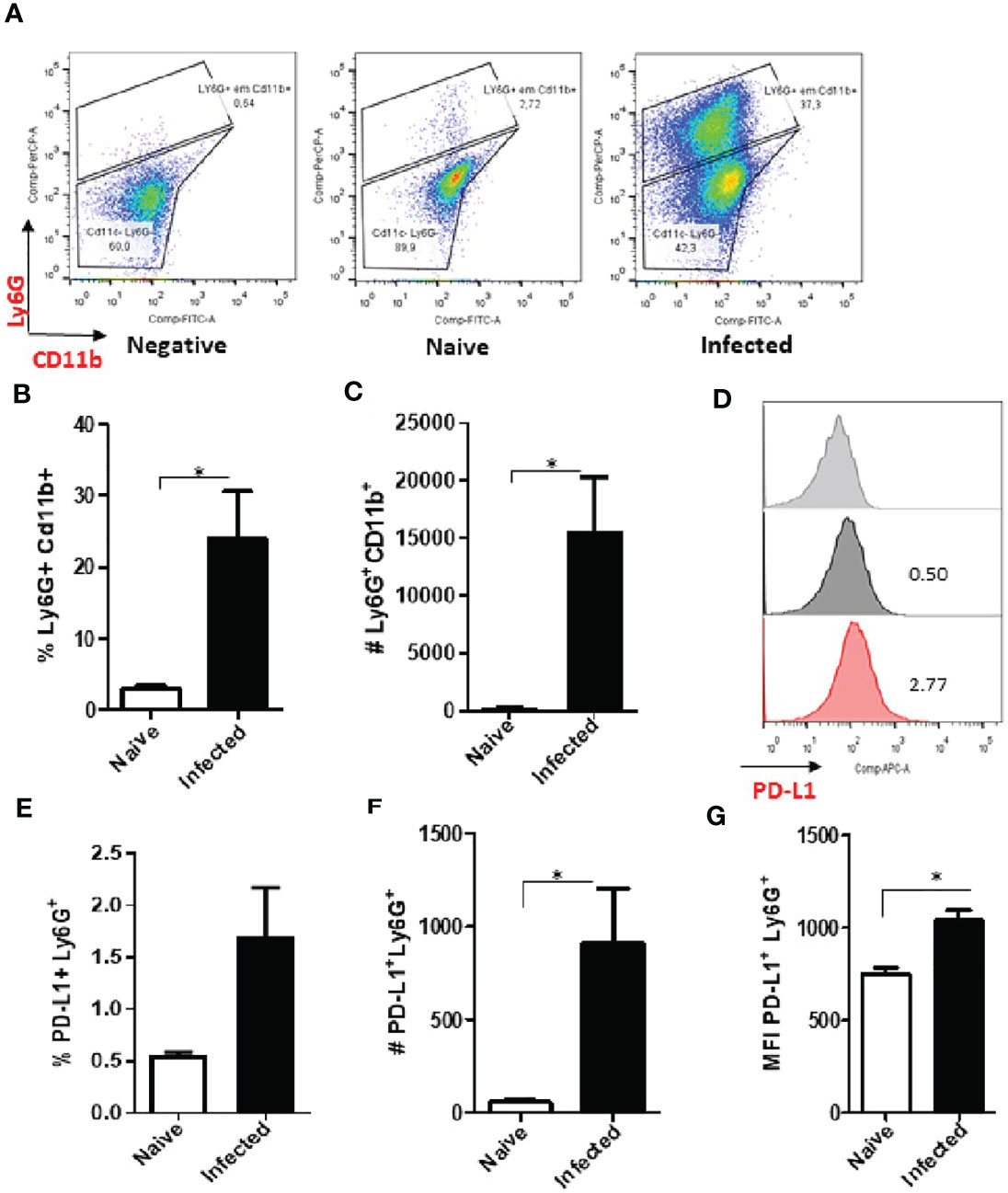
Figure 4 Increase of PD-L1+ Ly6G+ cells in L. amazonensis-infected ear lesions. Cells were collected from L. amazonensis-infected mouse ears after around 50 days. Controls were performed with uninfected mice (naïve). (A) Dot plot showing Ly6G+ CD11b+ cells (Ly6G-PerCP, CD11b-FITC). (B) Percentage of Ly6G+ CD11b+ cells. (C) Number of Ly6G+ CD11b+ cells. (D) Histogram of PD-L1 expression. Grey = Fluorescence minus one control (FMO) for PD-L1, Dark grey = Naïve and Red = Infected ear cells. (E) Percentage of PD-L1+ Ly6G+ cells, p=0.057. (F) Number of PD-L1+ Ly6G+ cells. (G) MFI of PD-L1+ Ly6G+. Data are mean ± SEM from two independent experiments (7-8 mice/group). *p < 0.02.
Intravital Images of Neutrophils Expressing PD-L1 in Infected Lesions but Not in Circulation
To validate our findings, we evaluated PD-L1 expression in neutrophils present in the lesions of L. amazonensis-infected mice directly in vivo. Initially, we analyzed the animals within 1 h of infection, since it is at this time that neutrophils begin the process of migrating to the infected site (32), in addition to the chronic lesion (60 days post-infection). Some migrating neutrophils in the tissue were expressing PD-L1, but interestingly, no expression was observed in the circulating neutrophils (Figure S11, Movies S1, S2). The absence of PD-L1 expression was observed in naïve and in saline-injected control mice (Figures S11A, B). After 1h post-infection, some migrating neutrophils in the tissue were expressing PD-L1, but interestingly, no expression was observed in the circulating neutrophils (Figure S11C; Movies S1 and S2). Importantly, chronic lesions showed a high presence of neutrophils expressing PD-L1 (Figure S11D). In fact, the presence of PD-L1-expressing neutrophils, at times, formed a mass (Movie S3) that we could image only after cutting the lesion tissue. After this step, we could see the circulation contralateral to the injury (Movie S4). Interestingly, this contralateral circulation did not have circulating neutrophils expressing PD-L1. This observation is important because it demonstrates that the infection environment is conducive to the induction of the suppressive neutrophil without affecting the circulating neutrophils.
PD-L1-Expressing Neutrophils in the Draining Lymph Nodes of Mice
To better map PD-L1-expressing neutrophils we investigated these cells in the lymph nodes that drain the footpad lesions caused by L. amazonensis. Our results show that the neutrophil population was increased in the lymph nodes of the infected mice at 60 days (Figures 5A–C). The frequency and numbers of PD-L1-expressing neutrophils was also significantly increased (Figures 5D–F), however, the median intensity of fluorescence (MFI) was similar to the naïve control (Figure 5G). This data reveals that the neutrophils not only express PD-L1 at the infection site, but these cells, expressing the ligand, also have the ability to be drained to the lymph nodes.
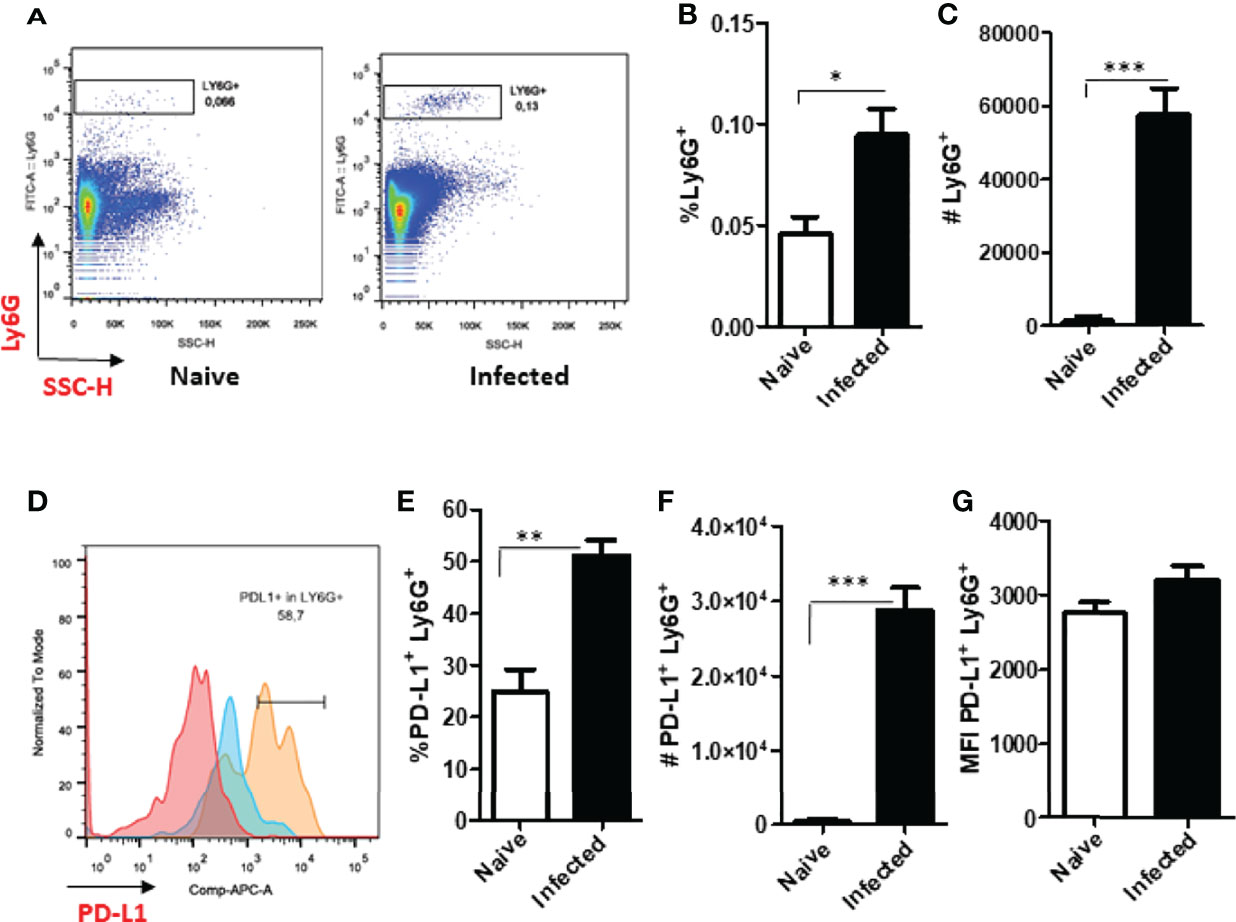
Figure 5 Increased expression of Ly6G+ and PD-L1+ in the draining lymph nodes of infected BALB/c mice. Mice were infected in the footpad with L. amazonensis promastigotes (2×106). After approximately 60 days, the draining lymph nodes were collected, macerated and the cells analyzed by flow cytometry. Controls were performed with uninfected mice (naïve). (A) Dot plot showing Ly6G+ cells (Ly6G-FITC x SSC-H). (B) Percentage of Ly6G+ cells. (C) Number of Ly6G+ cells. (D) Histogram of PD-L1 (PD-L1-APC) expression. Red = Fluorescence minus one control (FMO) PD-L1; Blue = Naïve lymph nodes; Orange = Infected lymph nodes. (E) Percentage of PD-L1+ Ly6G+ cells. (F) Number of PD-L1+ Ly6G+ cells. (G) MFI of PD-L1+Ly6G+. Data are mean ± SEM of individual mice (5 mice/group) shown as a representative experiment of three producing the same result profile. *p < 0.01, **p < 0.001, ***p < 0.0001.
Neutrophils Expressing PD-L1 Suppress IFN-γ Production by T Cell
First, the expression of PD-L1 was evaluated in infected C57BL6 peritoneal neutrophils with L. amazonensis promastigotes (MOI 1:1; 1:5; 1:10). We observed 50% of infected neutrophils in 1:1 and 90% in 1:5 and 1:10 MOIs (Figure 6A). The expression of PD-L1 was not modulated by the infection rate, and similarly to the BALB/c neutrophils, bystander CSFE- neutrophils expressed significantly less PD-L1 (Figures 6B, C). Following, the capacity of PD-L1+-neutrophils to inhibit the production of IFN-γ by CD8+ T was evaluated by co-culturing infected neutrophils with activated CD8+ T cells (Figure 6D). Characterization of isolated CD8+ T cells is shown in Figure S9. We observed a significant reduction in the IFN-γ production by CD8+ T cells co-cultured with infected neutrophil (1:5) compared to the non-infected neutrophils (Neu). Importantly, addition of anti-PD-L1 or anti-PD-1 to the co-culture restored the production of IFN-γ. Moreover, the isotype control was unable to reverse IFN-γ inhibition mediated by infected neutrophils. This result demonstrated that inhibition of IFN-γ production is dependent of PD-L1/PD-1 (Figure 6D).
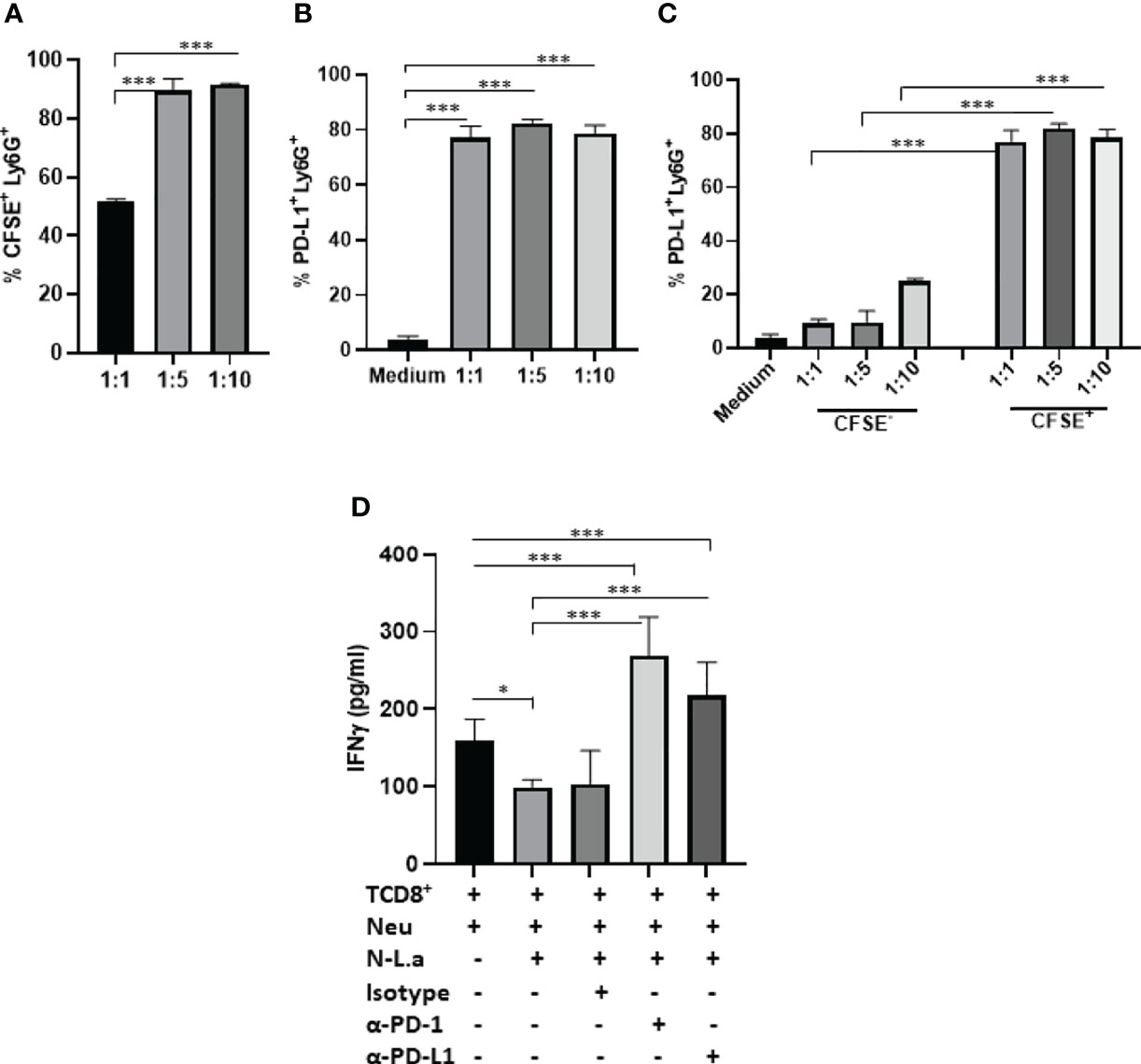
Figure 6 Coculture of murine neutrophils exposed to L. amazonensis with CD8+ T activated cells. C57BL/6 neutrophils (5x105) recruited to the peritoneum with casein were incubated with CFSE-stained promastigotes (Pro) of L. amazonensis (1:1, 1:5, 1:10), for 4 h. Control performed with neutrophils on medium. Cells were then analyzed by flow cytometry using Ly6G+-Percp (neutrophils), CFSE+ (L. amazonensis) and PD-L1+-APC. (A) Percentage of CFSE+ Ly6G+ cells. (B) Percentage of PD-L1+ Ly6G+ cells. (C) Percentage of PD-L1 expression on CFSE- (bystander) and CFSE+ cells. (D) Coculture of murine neutrophils (Neu) infected or not with promastigotes of L. amazonensis (Leish) with CD8+ T effectors cells. The coculture was treated or not with anti-PD-1 or anti-PD-L1 or isotype control for 18h. IFN-γ was analyzed in the culture supernatant by ELISA. Data are mean ± SEM. (A–C) N=3-4 and (D) N=7. *p=0.049, ***p < 0.001.
PD-L1-Expressing of Human Neutrophil In Vitro and in the Lesions Caused by Leishmania braziliensis
Finally, we evaluated the expression of PD-L1 in human neutrophils after Leishmania braziliensis infection. Differently from L. amazonensis, our results evidenced that independently of the MOI, neutrophils internalized both parasite stages of L. braziliensis at similar levels (Figure 7A). However, PD-L1 expression was higher in promastigotes infected-neutrophils and likewise L. amazonensis, PD-L1 was not expressed in the bystander neutrophils (Figures 7B, C).
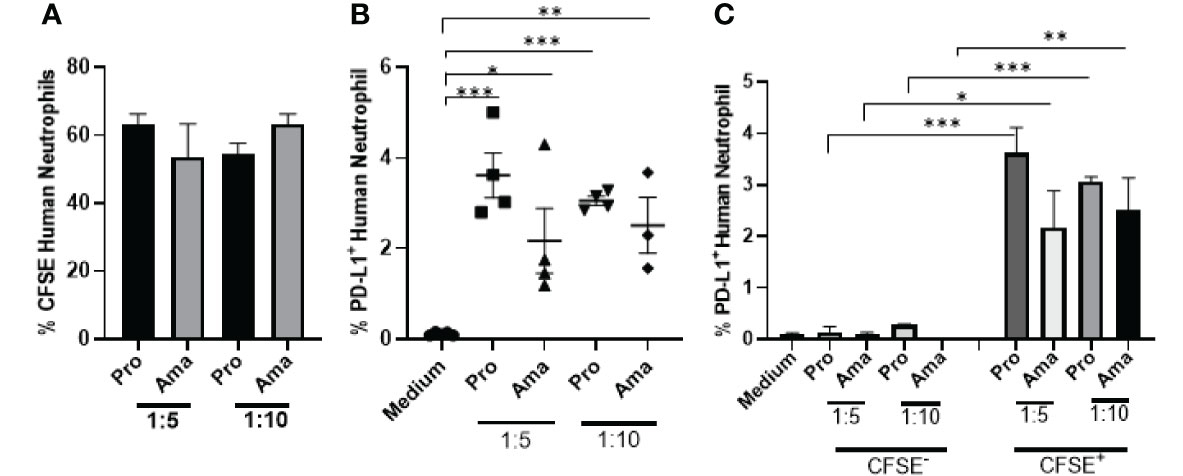
Figure 7 Increased expression of PD-L1 in human neutrophils exposed to L. braziliensis. Human neutrophils (5x105) from healthy donors were incubated with CFSE-stained promastigotes (Pro) and amastigotes (Ama) of L. braziliensis (1:5 and 1:10), for 4 h. Control performed with neutrophils on medium. Cells were then analyzed by flow cytometry. (A) Percentage of neutrophils infected with CFSE-labeled parasites. (B) Percentage of PD-L1+ neutrophils upon interaction with parasites. (C) Percentage of PD-L1 expression on CFSE- (bystander) and CFSE+ neutrophils. Data are mean ± SEM (N=4-5). *p < 0.0303, **p < 0.0047 ***p < 0.0001.
We also investigated the PD-L1 expression on neutrophils from human cutaneous leishmaniasis biopsies obtained from untreated patients. It is already been demonstrated that cutaneous leishmaniasis lesions present hyperplasia with a dense inflammatory cellular infiltrate involving the dermal-epidermal junction, consisting mainly of lymphocytes, macrophages, neutrophils and plasma cells (33–38). Thus, we evaluated whether neutrophils in the lesions expressed PD-L1, which could directly impact on the skin immune balance during infection. Interestingly, lesional analysis revealed the expression of PD-L1 on neutrophils, that were respectively 7.44 and 3.77-fold higher than healthy controls (Figures 8A–C and S6), suggesting that neutrophils may have a role in human injuries caused by L. braziliensis, which still needs to be assessed.
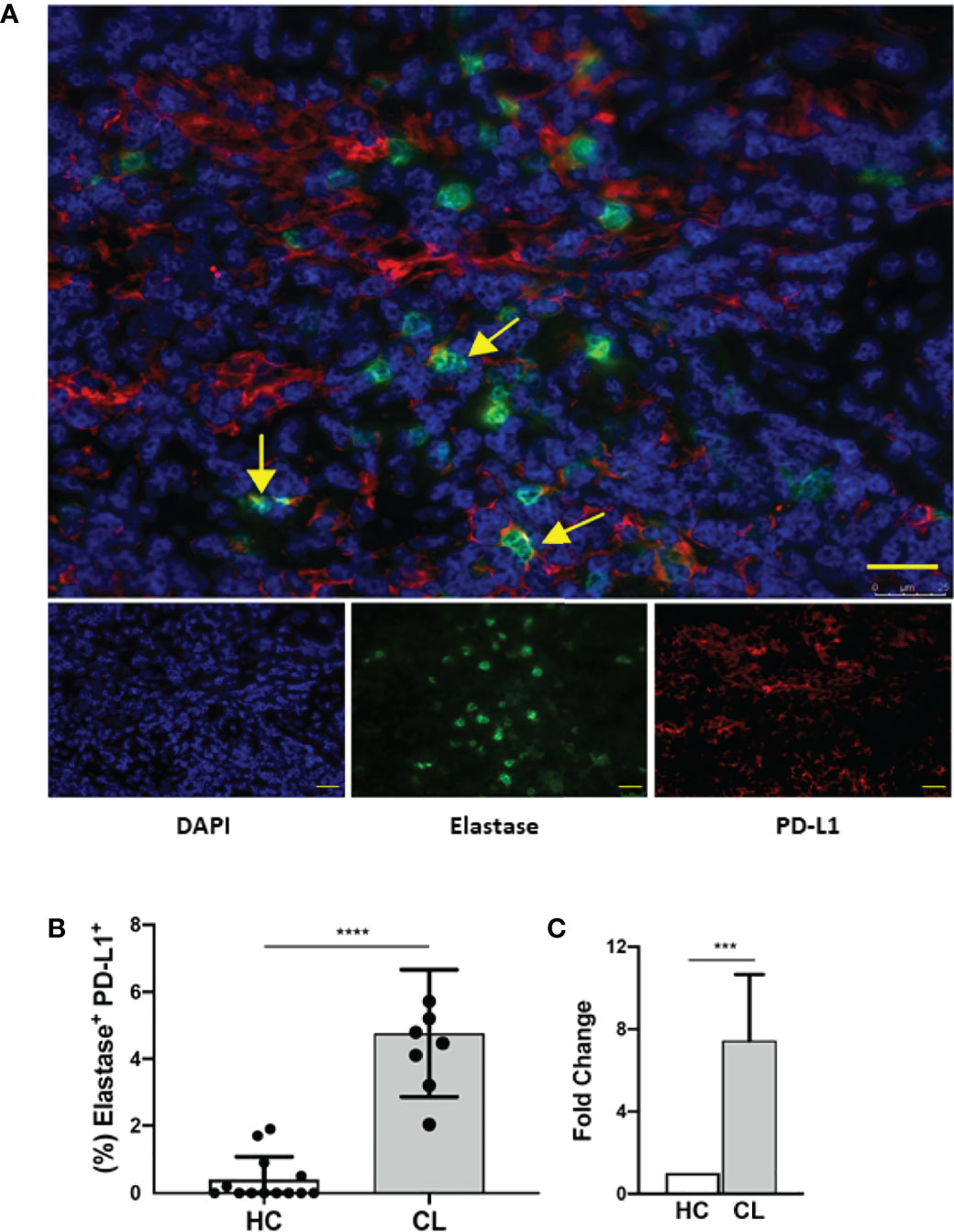
Figure 8 Human lesion neutrophils present a conspicuous expression of PD-L1. Healthy human skin and human cutaneous leishmaniasis skin lesions were stained with anti-elastase (green), anti-PD-L1 (red) and counter-stained with DAPI (blue). (A) Representative immunofluorescence staining of a cutaneous leishmaniasis lesion. Yellow arrows indicate anti-PD-L1 and anti-elastase double-stained cells. Bars = 50 µm. (B) Frequency of elastase-positive PD-L1+ cells of cumulative data of PD-L1 in neutrophils in healthy control skin (HC, N = 12) and in cutaneous leishmaniasis skin lesions (CL, N = 7). (C) Frequencies of double-stained neutrophils were normalized and data are expressed as fold over the control. Data shown as median with 95% confidence. The p values were calculated using Mann-Whitney test. ***p < 0.001.
Discussion
In the present work we investigated the expression of PD-L1 in human and murine neutrophils, showing that upon interaction with promastigotes or amastigotes of L. amazonensis, these host cells express PD-L1, a molecule involved in T cell exhaustion. Interestingly, a higher number of murine bone marrow and human peripheral neutrophils expressing PD-L1 was evidenced after interaction with amastigotes than with promastigotes. Although, human neutrophils were less infected by L. amazonensis amastigotes than the murine neutrophils, this parasite stage induced higher numbers cells expressing PD-L1. However, these effects were not observed in casein-recruited murine neutrophils, where similar numbers of PD-L1+ neutrophils were stimulated by both parasite developmental stages, and which did not display any differences in neutrophil infection between the two parasite forms studied. A higher number of casein-recruited neutrophils expressed PD-L1 than bone marrow neutrophils upon interaction with both parasite stages. This result could be due to the inflammation induced by casein, since it has been shown that inflammation could contribute to a higher PD-L1 expression (16). The same lack of inflammatory stimuli could explain the low frequency of human PD-L1+ neutrophils in comparison with the murine neutrophils, but it should be noted that the frequency of PD-L1-expressing-neutrophils increased 8 times in relation to control (medium) after only 4h of interaction with the amastigotes. This high expression of PD-L1 in amastigote-infected neutrophils may be the consequence of its increased survival, consistent with the study highlighting the resistance of L. amazonensis amastigotes against microbicidal mechanisms of neutrophils, such as the oxidative burst (39).
Regardless of the source of neutrophils, the expression of PD-L1 was not affected in bystander cells irrespective of the presence of parasite forms in the co-culture. Taken together, this suggests that upon infection of neutrophils, the parasite induces this suppression marker in the host cell. Expression of PD-L1 has been described in monocytes from a patient presenting diffuse cutaneous leishmaniasis (40), and only in low-density neutrophils (normal-density neutrophils did not expressed PD-L1) from human visceral leishmaniasis patients (12), but, to the best of the authors’ knowledge, this is the first description of PD-L1 expression upon in vitro interaction with Leishmania. Interestingly, similar to our results, it was shown that incubation of neutrophils ex vivo with LPS for 6 h also induced PD-L1 expression (41).
A swarm of neutrophils are recruited to the Leishmania infection site and the presence of these cells has been reported in chronic lesions as well (32, 42–45). Thus, we investigated PD-L1 expression in neutrophils from a L. amazonensis cutaneous lesion in mouse ears at both these early and late time-points. Remarkably, although PD-L1+ neutrophils could already be observed after 12h or 18 h post-infection, and a significant number of PD-L1+ neutrophils was detected after 15 days post-infection, the number and frequency of which decreased a little in 2-month chronic lesions. However, the mean fluorescence intensity greatly enhanced (around 4 times) in the chronic lesion, compared to the lesion at 15 days. It was recently demonstrated that in the beginning of the infection (1h-24h) (46), resident macrophages are the predominant infected cells. In the absence of infection, neutrophils are not stimulated to express PD-L1 as observed in the frequency in 12 or 18 hours post in vivo infection. In the chronic phase the combination of infected neutrophils and the inflammatory cytokines can prime the express of PD-L1 as observed in 15, 30 and 60 days post infection.
Next, we sought to image the environment of an L. amazonensis infection in vivo, and our results showed that as the infection progressed, there was an accumulation of neutrophils at the site of infection, and some of them expressing PD-L1. The presence of neutrophils is in agreement with reports describing these cells in chronic lesions (60 days post infection) of animals naturally or experimentally infected with Leishmania (42–44). Importantly, PD-L1 expression was not seen in neutrophils circulating in vessels close to the infected site, as observed by intravital microscopy. Instead, in human diffuse cutaneous leishmaniasis and in visceral leishmaniasis PD-L1-expressing cells have been detected in the blood circulation (12, 40). Similarly, higher levels of blood PD-L1+ neutrophils have been reported in tuberculosis and HIV-1-infected patients (7, 47), in patients with cancer (48), severe sepsis (49), while in rheumatoid disease, patients have neutrophils expressing PD-L1 in the synovial fluid, which can be related to the severity of the disease (50).
Another aspect identified in our study was the presence of PD-L1-expressing neutrophils in the draining lymph nodes of L. amazonensis infected mice. Upon analyzing the lymph nodes draining the chronic lesion we evidenced an increased number of neutrophils in this organ, and half of this population was expressing PD-L1. It has been reported that neutrophils have the ability to migrate to the lymph nodes, transporting and presenting antigens to T cells and inducing the activation and expansion of CD4+ T cells (51–54). We recently described in the draining lymph nodes of a non-healing L. amazonensis infection in BALB/c mice that both, CD4+ and CD8+ T cells expressed PD-1, and dendritic cells PD-L1 (28). Importantly, treatment of the L. amazonensis-infected mice with anti-PD-1 and anti-PD-L1 antibodies significantly increased IFN-γ production by T cells and decreased the parasite load (28). Our results in Leishmania-infected mice confirmed the presence of neutrophils in the lesion site in the first and second waves of neutrophil infiltration as well as in the chronic phase, and we perceived a relationship between PD-L1 expression and the cutaneous disease progression in the mouse.
It has been shown that neutrophil depletion with 1A8 monoclonal antibody enhanced T cell production of IFN-γ, suggesting a suppressor function of neutrophis (29). Here, we show the direct effect of Leishmania in the induction of PD-L1 on neutrophils, which interacting with CD8+ T cells, reduce IFN-γ production. Here, we demonstrated PD-L1 expression was detected in the skin of dogs severely infected with L. infantum (54), and in the spleen and in B cells of dogs infected with L. infantum (55–57). Several reports have identified the presence of neutrophils in chronic inflammatory infiltrate of patients with localized cutaneous (58, 59), diffuse cutaneous (60) and mucocutaneous leishmaniasis (61, 62).
Likewise to L. amazonensis induction of PD-L1 in human neutrophils, L. braziliensis infection in these cells shares the same capacity. Both promastigotes and amastigotes induced PD-L1 expression in neutrophils indicating an ability of this parasite to this phenotype. Finally, we analyzed lesions from patients with cutaneous leishmaniasis caused by L. braziliensis, evidencing an enhanced frequency of neutrophils expressing PD-L1 than in healthy controls. Similar increased PD-L1 transcripts have been reported in the lesions of patients infected with L. braziliensis (62). Our study demonstrated that neutrophils contributed to PD-L1 expression in the Leishmania lesion site.
Recently, it has been demonstrated by RNA-seq and immunohistochemistry that PD-1 and PD-L1 were highly up regulated in L. braziliensis skin lesion (63). Moreover, indicating an exhaustion process, anti-PD-L1/PD-L2 added to cultures of CD4+ and CD8+ T cells from cutaneous leishmaniasis patients increased the response to L. braziliensis antigen, and restored their IFN-γ response (63).
In summary, we show for the first time that L. amazonensis infection induces the expression of PD-L1 in neutrophils and that this expression increases with disease progression associated to chronic development of the disease. The induction of PD-L1 in neutrophils could favor the suppressive milieu that is important for the persistence of the parasite in human and experimental infections. Our finding is relevant because it opens new possibilities for therapeutic targets, and for understanding the local environment of the infection that may favor Leishmania growth.
Conclusion
The present study demonstrated that both amastigotes and promastigotes of L. amazonensis and L. braziliensis are capable of inducing the expression of PD-L1 in murine and human neutrophils. It was also observed that, in a murine model, the expression is not present in the first wave of neutrophil infiltration, however, the expression increases in the lesion and in the draining lymph nodes as the disease progress. Importantly, we demonstrated the capacity of Leishmania-infected neutrophils to inhibit IFN-γ production by effector TCD8+ cells, which was reversed by anti-PD-L1. Overall, our findings strongly suggest that PD-L1 expressing neutrophils could participate in the modulation of the immune response, favoring Leishmania survival and persistence.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by the HUCAM Ethical Committee and this study was registered under the number 735.274. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by Committee for Animal Use of the Universidade Federal do Rio de Janeiro (Permit Number: 161/18).
Author Contributions
AF-M: Conceived and designed the analysis, data collection, data analysis and interpretation, scientific discussion, wrote and revised the manuscript. PL-G: Data collection. MA: Data collection, data analysis and interpretation. RG Data collection, data analysis, interpretation and critical revision of the article. LC: Data collection, data analysis, interpretation and critical revision of the article. GM: Scientific discussion. DG: Scientific discussion and critical revision of the article and funding. ES: Conceived and designed the analysis, scientific discussion, wrote and critical revised the manuscript and funding. HM: Conceived and designed the analysis, scientific discussion, IDEM acima and funding. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the kind donations by Dr. Milton A.P. Oliveira (Universidade Federal de Goiás, GO, Brazil), Dr. André M. Vale (Universidade Federal do Rio de Janeiro, RJ, Brazil) and Bristol-Myers Squibb (Nova York, NY, EUA), respectively of L. braziliensis (MHOM/BR/2005/RPL5) strain, anti-IgG2A and anti-PD-L1. The work was supported by the following Brazilian agencies: Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ); Programa Jovem Cientista do Nosso Estado (FAPERJ); Productivity Fellowships from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) Finance code 001. The funders had no role in the study design, data collection, analysis, decision to publish or preparation of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.598943/full#supplementary-material
Supplementary Figure 1 | Comparison of PD-L1 expression in murine neutrophils from bone marrow and peritonium. Neutrophils recruited to the peritoneum with casein (Per) for 3 h or isolated from the bone marrow (BM) of mice. Cells were infected (1:10) with promastigotes (Pro) or amastigotes (Ama) of L. amazonensis for 4 h. Controls were performed with the same cells without infection (control). Data shown percentage of PD-L1+ cells are mean + SEM (N = 9-11). **p < 0.03, ***p < 0.0001.
Supplementary Figure 2 | PD-L1 expression in human neutrophils infected with L. amazonensis. Human neutrophils (5x105) from healthy donors were incubated with CFSE-stained promastigotes (Pro) and amastigotes (Ama) of L. amazonensis (1:5 and 1:10), for 4 h. Control performed with neutrophils on medium. Cells were then analyzed by flow cytometry. Data shown as percentage of PD-L1 expression on human neutrophils, mean ± SEM (N= 5-6). ***p < 0.0001.
Supplementary Figure 3 | Analysis of murine ear neutrophils after 18 h of infection. Cells were collected from L. amazonensis-infected ears after 18 h then submitted to flow cytometry. Controls were performed with uninfected mice (naïve). (A) Percentage of CD11b+ cells. (B) Number of CD11b+ cells. (C) Percentage of Ly6G+ CD11b+ cells. (D) Number of Ly6G+ CD11b+ cells. (E) Histogram of PD-L1 expression. Grey = Fluorescence minus one control (FMO) PD-L1, Blue = Naïve and Red = Infected. (F) Percentage of PD-L1+ Ly6G+ cells. (G) Number of PD-L1+ Ly6G+ cells, p=0.055. (H) MFI of PD-L1+ Ly6G+. Data are mean ± SEM from cells of individual mice (N = 4 mice/group).
Supplementary Figure 4 | Dot plots of murine cells from lesions infected with L. amazonensis. Cells were collected from L. amazonensis-infected ears after 18 h. Controls were performed with uninfected mice (naïve) and FMO for (A) CD11b and (B) Ly6G (negative). (A) Dot plot of CD11b+ cells. (B) Dot plot of Ly6G+ CD11b+ cells. Data of individual mice (4 mice/group).
Supplementary Figure 5 | Expression of PD-L1+ Ly6G+ cells in L. amazonensis-infected lesions. Cells were collected from L. amazonensis-infected ears after approximately 60 days. Controls were performed with uninfected mice (naïve). (A) Dot plot showing CD11b+ cells (PE-CD11b x SSC) on FMO for CD11b (negative), naïve and infected mice. (B) Percentage of CD11b+ cells. (C) Number of CD11b+ cells. (D) Dot plot showing PD-L1+ Ly6G+ and PD-L1+ Ly6G- gates (PD-L1-APC x Ly6G-PerCP) on FMO Ly6G and PD-L1, naïve and infected mice. **p<0.008 Data are mean ± SEM of individual mice (4-5 mice/group) and are representative of three experiments producing the same result profile.
Supplementary Figure 6 | Analyses of lesion neutrophils present a conspicuous expression of PD-L1. PD-L1 expression in neutrophils from healthy skin (HC, N = 8) and cutaneous leishmaniasis lesions (CL, N = 9). (A) Neutrophils are a percentage of nucleated cells. (B) Percentage PD-L1+ neutrophils. The P values were calculated using Student’s t test with Mann-Whitney test. ***p < 0.001.
Supplementary Figure 7 | Gating strategy of CFSE+ neutrophils expressing PD-L1. (A) Neutrophils FSC x SSC. (B) Single cells - FSC-A x FSC-H. (C) CFSE+ and CFSE- (CFSE x SSC). (D) Histogram of PD-L1 expression, Green = FMO (negative), Red= isotype, Orange = control not infected, Blue = infected.
Supplementary Figure 8 | Gating strategy of neutrophils expressing PD-L1 on the mice ear and on draining lymph nodes. Ear cells after processing were analyzed as follows: (A) Cells FSC x SSC. (B) Single cells - FSC-A x FSC-H. (C) CD45+ (APC-cy7 x FSC-H). (D) CD11b+ and Ly6G+ (FITC x Percp). (E) Histogram of PD-L1 expression, Grey = FMO (negative), Black= naïve control (not infected), Red = infected. Draining lymph node cells were analyzed as follows: (F) Cells FSC x SSC. (G) Single cells - FSC-A x FSC-H. (H) CD11b+ (Pe-cy7 x SSC-A). (I) Ly6G+ CD11b+ (FITC x SSC-H). (J) Histogram of PD-L1 expression, Red = FMO (negative), Blue= naïve control (not infected), Orange = infected.
Supplementary Figure 9 | Phenotype of CD8+ T cells differentiated in vitro. Spleen cells of C57BL/6 mice were processed, and CD8+ T cells purified by negative selection and sorted. (A) Pre-sort and Pos-sort SSC x CD8+, CD62L+ x CD44+. (B) On day 5, cells were stained with CD25+ x CD44+, CD127+ x CD44+ and CD62L+ x CD44+.
Supplementary Figure 10 | Lesion development in L. amazonensis-infected mice. Mice were infected in the ear with L. amazonensis stationary-phase promastigotes (2x106). Ear thickness was measured weekly using a thickness gauge.
Supplementary Figure 11 | PD-L1-expressing neutrophils in infected ear lesions. BALB/c mice infected in the ear with L. amazonensis were injected with anti-PD-L1-APC and anti-Ly6G-FITC antibodies and imaged by intravital microscopy. Controls were performed with mice injected with PBS and non-infected (naïve) mice. Arrows show neutrophils expressing PD-L1. (A) Naïve mouse. (B) Control mouse injected with PBS. (C) Mouse infected for 1 h. (D) Mouse with chronic infection (60 days). Autofluorescence of the skin fur can be observed (Arrow head). Images were obtained using Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation at: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x. Bars: 130 µm.
Movie S1 | Neutrophils in the blood vessels in the infected ear did not express PD-L1. Mice infected in the ears with L. amazonensis for 1 h were injected with anti-PD-L1-APC and anti-Ly6G-FITC antibodies. White arrows indicate circulating neutrophils with no expression of PD-L1. Images were obtained using a Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation at: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x. Presence of skin hair autofluorescence. Bar = 130 µm.
Movie S2 | Migration of neutrophils without expression of PD-L1 to the infected ear. Mice infected in the ear with L. amazonensis for 1 h were injected with anti-PD-L1-APC and anti-Ly6G-FITC antibodies. Presence of skin hair autofluorescence (Arrow head). Images were obtained using a Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation at four wavelengths: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x and 2x zoom. Bar = 62 µm.
Movie S3 | Neutrophils expressing PD-L1 in a chronic L. amazonensis ear infection. Mice infected in the ear with L. amazonensis in the chronic infection (60 days) were injected with anti-PD-L1-APC and anti-Ly6G-FITC antibodies. Images were obtained using a Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation at four wavelengths: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x. Bar =38,57 µm and 127,77 µm.
Movie S4 | Circulating neutrophils not expressing PD-L1 in a chronic L. amazonensis. Mice infected in the ear with L. amazonensis in the chronic infection (60 days) were injected with anti-PD-L1-APC and anti-Ly6G-FITC antibodies. Presence of skin hair autofluorescence (Arrow head). White arrows indicate circulating neutrophils with no expression of PD-L1. Images were obtained using a Nikon Eclipse Ti with an A1R confocal head equipped with four different lasers (excitation at four wavelengths: 405, 488, 546 and 647 nm) and emission bandpass filters at 450/50, 515/30, 584/50 and 663/738 nm. Objective Plan Apo λ 10x. Bar = 130 µm.
References
1. Nordenfelt P and Tapper H. Phagosome Dynamics During Phagocytosis by Neutrophils. J Leukoc Biol (2011) 90:271–84. doi: 10.1189/jlb.0810457
2. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil Extracellular Traps Kill Bacteria. Science (2004) 303: (5663):1532–5. doi: 10.1126/science.1092385
3. Amulic B, Cazalet C, Hayes GL, Metzler KD, Zychlinsky A. Neutrophil Function: From Mechanisms to Disease. Annu Rev Immunol (2012) 30:459–89. doi: 10.1146/annurev-immunol-020711-074942
4. Yang F, Feng C, Zhang X, Lu J, Zhao Y. The Diverse Biological Functions of Neutrophils, Beyond the Defense Against Infections. Inflammation (2017) 40(1):311–23. doi: 10.1007/s10753-016-0458-4
5. Castanheira FVS, Kubes P. Neutrophils and NETs in Modulating Acute and Chronic Inflammation. Blood (2019) 133:2178–85. doi: 10.1182/blood-2018-11-844530
6. Colón DF, Wanderley CW, Franchin M, Silva CM, Hiroki CH, Castanheira FVS, et al. Neutrophil Extracellular Traps (Nets) Exacerbate Severity of Infant Sepsis. Crit Care (2019) 23(1):113. doi: 10.1186/s13054-019-2407-8
7. Bowers NL, Helton ES, Huijbregts RP, Goepfert PA, Heath SL, Hel Z. Immune Suppression by Neutrophils in HIV-1 Infection: Role of PD-L1/PD-1 Pathway. PloS Pathog (2014) 10(3):e1003993. doi: 10.1371/journal.ppat.1003993
8. Kho S, Minigo G, Andries B, Leonardo L, Prayoga P, Poespoprodjo JR, et al. Circulating Neutrophil Extracellular Traps and Neutrophil Activation Are Increased in Proportion to Disease Severity in Human Malaria. J Infect Dis (2019) 219(12):1994–2004. doi: 10.1093/infdis/jiy661
9. Hurrell BP, Regli IB, Tacchini-Cottier F. Different Leishmania Species Drive Distinct Neutrophil Functions. Trends Parasitol (2016) 32:392–401. doi: 10.1016/j.pt.2016.02.003
10. Zemans RL. Neutrophil-Mediated T-Cell Suppression in Influenza: Novel Finding Raising Additional Questions. Am J Respir Cell Mol Biol (2018) 58(4):423–5. doi: 10.1165/rcmb.2017-0425ED
11. Abebe T, Takele Y, Weldegebreal T, Cloke T, Closs E, Corset C, et al. Arginase Activity - A Marker of Disease Status in Patients With Visceral Leishmaniasis in Ethiopia. PloS Neglected Trop Dis (2013) 7(3):e2134. doi: 10.1371/journal.pntd.0002134
12. Sharma S, Davis RE, Srivastva S, Nylén S, Sundar S, Wilson ME. A Subset of Neutrophils Expressing Markers of Antigen-Presenting Cells in Human Visceral Leishmaniasis. J Infect Dis (2016) 214(10):1531–8. doi: 10.1093/infdis/jiw394
13. Greenwald RJ, Freeman GJ, Sharpe AH. The B7 Family Revisited. Annu Rev Immunol (2005) 23:515–48. doi: 10.1146/annurev.immunol.23.021704.115611
14. Okazaki T, Honjo T. The PD-1-PD-L Pathway in Immunological Tolerance. Trends Immunol (2006) 27(4):195–201. doi: 10.1016/j.it.2006.02.001
15. Chen L. Co-Inhibitory Molecules of the B7-CD28 Family in the Control of T-cell Immunity. Nat Rev Immunol (2004) 4(5):336–47. doi: 10.1038/nri1349
16. Bankey PE, Banerjee S, Zucchiatti A, De M, Sleem RW, Lin CFL, et al. Cytokine Induced Expression of Programmed Death Ligands in Human Neutrophils. Immunol Lett (2010) 129:100–7. doi: 10.1016/j.imlet.2010.01.006
17. Kythreotou A, Siddique A, Mauri FA, Bower M, Pinato DJ. Pd-L1. J Clin Pathol (2018) 71:189–94. doi: 10.1136/jclinpath-2017-204853
18. Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, et al. Engagement of the PD-1 Immunoinhibitory Receptor by A Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J Exp Med (2000) 192(7):1027–34. doi: 10.1084/jem.192.7.1027
19. Dong H, Zhu G, Tamada K, Chen L. B7-H1, a Third Member of the B7 Family, Co-Stimulates T-Cell Proliferation and Interleukin-10 Secretion. Nat Med (1999) 5(12):1365–9. doi: 10.1038/70932
20. Jones D, Como CN, Jing L, Blackmon A, Neff CP, Krueger O, et al. Varicella Zoster Virus Productively Infects Human Peripheral Blood Mononuclear Cells to Modulate Expression of Immunoinhibitory Proteins and Blocking PD-L1 Enhances Virus-Specific CD8+ T Cell Effector Function. PloS Pathog (2019) 15(3):e1007650. doi: 10.1371/journal.ppat.1007650
21. de Mel S, Tan JZ-C, Jeyasekharan AD, Chng W-J, Ng S-B. Transcriptomic Abnormalities in Epstein Barr Virus Associated T/NK Lymphoproliferative Disorders. Front Pediatr (2019) 6:405. doi: 10.3389/fped.2018.00405
22. Sanjo N, Nose Y, Shishido-Hara Y, Mizutani S, Sekijima Y, Aizawa H, et al. A Controlled Inflammation and a Regulatory Immune System are Associated With More Favorable Prognosis of Progressive Multifocal Leukoencephalopathy. J Neurol (2019) 266(2):369–77. doi: 10.1007/s00415-018-9140-0
23. Bardhan K, Anagnostou T, Boussiotis VA. The PD1:PD-L1/2 Pathway From Discovery to Clinical Implementation. Front Immunol (2016) 7:550. doi: 10.3389/fimmu.2016.00550
24. Patel R, Bock M, Polotti CF, Elsamra S. Pharmacokinetic Drug Evaluation of Atezolizumab for the Treatment of Locally Advanced or Metastatic Urothelial Carcinoma. Expert Opin Drug Metab Toxicol (2017) 13(2):225–32. doi: 10.1080/17425255.2017.1277204
25. Wang C, Yu X, Wang W. A Meta-Analysis of Efficacy and Safety of Antibodies Targeting PD-1/PD-L1 in Treatment of Advanced Nonsmall Cell Lung Cancer. Med (Baltimore) (2016) 95(52):e5539. doi: 10.1097/MD.0000000000005539
26. Chamoto K, Al-Habsi M, Honjo T. Role of PD-1 in Immunity and Diseases. Curr Top Microbiol Immunol (2017) 410:75–97. doi: 10.1007/82_2017_67
27. Wang J, Jebbawi F, Bellanger AP, Beldi G, Millon L, Gottstein B. Immunotherapy of Alveolar Echinococcosis Via PD-1/PD-L1 Immune Checkpoint Blockade in Mice. Parasite Immunol (2018) 40(12):e12596. doi: 10.1111/pim.12596
28. da Fonseca-Martins AM, Ramos TD, Pratti JES, Firmino-Cruz L, Gomes DCO, Soong L, et al. Immunotherapy Using Anti-PD-1 and Anti-PD-L1 in L. amazonensis-Infected BALB/c Mice Reduce Parasite Load. Sci Rep (2019) 9(1):20275. doi: 10.1038/s41598-019-56336-8
29. Ribeiro-Gomes FL, Peters NC, Debrabant A, Sacks DL. Efficient Capture of Infected Neutrophils by Dendritic Cells in the Skin Inhibits the Early Anti-Leishmania Response. PloS Pathog (2012) 8(2):e1002536. doi: 10.1371/journal.ppat.1002536
30. Scott-Browne JP, López-Moyado IF, Trifari S, Wong V, Chavez L, Rao A, et al. Dynamic Changes in Chromatin Accessibility Occur in CD8+ T Cells Responding to Viral Infection. Immunity (2016) 45(6):1327–40. doi: 10.1016/j.immuni.2016.10.028
31. Pipkin ME, Sacks JA, Cruz-Guilloty F, Lichtenheld MG, Bevan MJ, Rao A. Interleukin-2 and Inflammation Induce Distinct Transcriptional Programs That Promote the Differentiation of Effector Cytolytic T Cells. Immunity (2010) 32(1):79–90. doi: 10.1016/j.immuni.2009.11.012
32. Peters NC, Egen JG, Secundino N, Debrabant A, Kimblin N, Kamhawi S, et al. In Vivo Imaging Reveals an Essential Role for Neutrophils in Leishmaniasis Transmitted by Sand Flies. Science (2008) 321(5891):970–4. doi: 10.1126/science.1159194
33. Schubach A, Cuzzi-Maya T, Oliveira AV, Sartori A, de Oliveira-Neto MP, Mattos MS, et al. Leishmanial Antigens in the Diagnosis of Active Lesions and Ancient Scars of American Tegumentary Leishmaniasis Patients. Mem Inst Oswaldo Cruz (2001) 96(7):987–96. doi: 10.1590/s0074-02762001000700018
34. Viana AG, Mayrink W, Fraga CA, Silva LM, Domingos PL, Bonan PR, et al. Histopathological and Immunohistochemical Aspects of American Cutaneous Leishmaniasis Before and After Different Treatments. Bras Dermatol (2013) 88(1):32–40. doi: 10.1590/s0365-05962013000100003
35. Mendes DS, Dantas ML, Gomes JM, Santos WL, Silva AQ, Guimarães LH, et al. Inflammation in Disseminated Lesions: An Analysis of CD4+, Cd20+, CD68+, CD31+ and vW+ Cells in non-Ulcerated Lesions of Disseminated Leishmaniasis. Mem Inst Oswaldo Cruz (2013) 108(1):18–22. doi: 10.1590/s0074-02762013000100003
36. Eid AA, Badr El Dine FM, Nabil IM. Histopathologic and Ultrastructural Changes in Seminiferous Tubules of Adult Male Albino Rats Following Daily Administration of Different Doses of Tadalafil. Urology (2016) 90:89–96. doi: 10.1016/j.urology.2015.12.038
37. Gomes AHS, Martines RB, Kanamura CT, Barbo MLP, Iglezias SD, Lauletta Lindoso JA, et al. American Cutaneous Leishmaniasis: In Situ Immune Response of Patients With Recent and Late Lesions. Parasite Immunol (2017) 39(4):1–10. doi: 10.1111/pim.12423
38. Covre LP, Devine OP, Garcia de Moura R, Vukmanovic-Stejic M, Dietze R, Ribeiro-Rodrigues R, et al. Compartmentalized Cytotoxic Immune Response Leads to Distinct Pathogenic Roles of Natural Killer and Senescent CD8+ T Cells in Human Cutaneous Leishmaniasis. Immunology (2020) 159(4):429–40. doi: 10.1111/imm.13173
39. Carlsen ED, Hay C, Henard CA, Popov V, Garg NJ, Soong L. L. amazonensis Amastigotes Trigger Neutrophil Activation But Resist Neutrophil Microbicidal Mechanisms. Infect Immun (2013) 81(11):3966–74. doi: 10.1128/IAI.00770-13
40. Barroso DH, Falcão SAC, da Motta JOC, Sevilha Dos Santos L, Takano GHS, Gomes CM, et al. Pd-L1 May Mediate T-Cell Exhaustion in A Case of Early Diffuse Leishmaniasis Caused By L. amazonensis. Front Immunol (2018) 9:1021. doi: 10.3389/fimmu.2018.01021
41. de Kleijn S, Langereis JD, Leentjens J, Kox M, Netea MG, Koenderman L, et al. Ifn-γ-Stimulated Neutrophils Suppress Lymphocyte Proliferation Through Expression of PD-L1. PloS One (2013) 8(8):e72249. doi: 10.1371/journal.pone.0072249
42. Souza-Lemos C, de-Campos SN, Teva A, Côrte-Real S, Fonseca EC, Porrozzi R, et al. Dynamics of Immune Granuloma Formation in a Leishmania Braziliensis-Induced Self-Limiting Cutaneous Infection in the Primate Macaca Mulatta. J Pathol (2008) 216(3):375–86. doi: 10.1002/path.2403
43. Lopez Kostka S, Dinges S, Griewank K, Iwakura Y, Udey MC, von Stebut E. Il-17 Promotes Progression of Cutaneous Leishmaniasis in Susceptible Mice. J Immunol (2009) 182(5):3039–46. doi: 10.4049/jimmunol.0713598
44. Verçosa BL, Melo MN, Puerto HL, Mendonça IL, Vasconcelos AC. Apoptosis, Inflammatory Response and Parasite Load in Skin of Leishmania (Leishmania) Chagasi Naturally Infected Dogs: A Histomorphometric Analysis. Veterinary Parasitology (2012) 189(2-4):162–70. doi: 10.1016/j.vetpar.2012.04.035
45. Morgado FN, Nascimento MT, Saraiva EM, de Oliveira-Ribeiro C, Madeira Mde F, da Costa-Santos M, et al. Are Neutrophil Extracellular Traps Playing a Role in the Parasite Control in Active American Tegumentary Leishmaniasis Lesions? PloS One (2015) 10(7):e0133063. doi: 10.1371/journal.pone.0133063
46. Chaves MM, Lee SH, Kamenyeva O, Ghosh K, Peters NC, Sacks D. The Role of Dermis Resident Macrophages and Their Interaction With Neutrophils in the Early Establishment of Leishmania Major Infection Transmitted by Sand Fly Bite. PloS Pathog (2020) 16(11):e1008674. doi: 10.1371/journal.ppat.1008674
47. McNab FW, Berry MP, Graham CM, Bloch SA, Oni T, Wilkinson KA, et al. Programmed Death Ligand 1 is Over-Expressed by Neutrophils in the Blood of Patients With Active Tuberculosis. Eur J Immunol (2011) 41(7):1941–7. doi: 10.1002/eji.201141421
48. He G, Zhang H, Zhou J, Wang B, Chen Y, Kong Y, et al. Peritumoural Neutrophils Negatively Regulate Adaptive Immunity Via the PD-L1/PD-1 Signalling Pathway in Hepatocellular Carcinoma. J Exp Clin Cancer Res (2015) 34:141. doi: 10.1186/s13046-015-0256-0
49. Fallon EA, Biron-Girard BM, Chung CS, Lomas-Neira J, Heffernan DS, Monaghan SF, et al. A Novel Role for Coinhibitory Receptors/Checkpoint Proteins in the Immunopathology of Sepsis. J Leukoc Biol (2018) 103:1151–64. doi: 10.1002/JLB.2MIR0917-377R
50. Luo Q, Huang Z, Ye J, Deng Y, Fang L, Li X, et al. Pd-L1-Expressing Neutrophils as A Novel Indicator to Assess Disease Activity of Rheumatoid Arthritis. Int J Clin Exp Med (2017) 10:7716–24. doi: 10.1186/s13075-016-0942-0
51. Abadie V, Badell E, Douillard P, Ensergueix D, Leenen PJ, Tanguy M, et al. Neutrophils Rapidly Migrate Via Lymphatics After Mycobacterium Bovis BCG Intradermal Vaccination and Shuttle Live Bacilli to the Draining Lymph Nodes. Blood (2005) 106(5):1843–50. doi: 10.1182/blood-2005-03-1281
52. Castell SD, Harman MF, Morón G, Maletto BA, Pistoresi-Palencia MC. Neutrophils Which Migrate to Lymph Nodes Modulate Cd4+ T Cell Response by a PD-L1 Dependent Mechanism. Front Immunol (2019) 10:105. doi: 10.3389/fimmu.2019.00105
53. Bogoslowski A, Wijeyesinghe S, Lee WY, Chen CS, Alanani S, Janne C, et al. Neutrophils Recirculate Through Lymph Nodes to Survey Tissues for Pathogens. J Immunol (2020) 204:2552–61. doi: 10.4049/jimmunol.2000022
54. Ordeix L, Montserrat-Sangrà S, Martínez-Orellana P, Baxarias M, Solano-Gallego L. Toll-Like Receptors 2, 4 and 7, Interferon-Gamma and Interleukin 10, and Programmed Death Ligand 1 Transcripts in Skin From Dogs of Different Clinical Stages of Leishmaniosis. Parasit Vectors (2019) 12(1):575. doi: 10.1186/s13071-019-3827-7
55. de Souza TL, da Silva AVA, Pereira LOR, Figueiredo FB, Mendes AAV Jr, Menezes RC, et al. Pro-Cellular Exhaustion Markers are Associated With Splenic Microarchitecture Disorganization and Parasite Load in Dogs With Visceral Leishmaniasis. Sci Rep (2019) 9:12962. doi: 10.1038/s41598-019-49344-1
56. Chiku VM, Silva KL, de Almeida BF, Venturin GL, Leal AA, de Martini CC, et al. PD-1 Function in Apoptosis of T Lymphocytes in Canine Visceral Leishmaniasis. Immunobiology (2016) 221(8):879–88. doi: 10.1016/j.imbio.2016.03.007
57. Schaut RG, Lamb IM, Toepp AJ, Scott B, Mendes-Aguiar CO, Coutinho JF, et al. Regulatory Igdhi B Cells Suppress T Cell Function Via IL-10 and PD-L1 During Progressive Visceral Leishmaniasis. J Immunol (2016) 196(10):4100–9. doi: 10.4049/jimmunol.1502678
58. Dabiri S, Hayes MM, Meymandi SS, Basiri M, Soleimani F, Mousavi MR. Cytologic Features of “Dry-Type” Cutaneous Leishmaniasis. Diagn Cytopathol (1998) 19(3):182–5. doi: 10.1002/(sici)1097-0339(199809)19:3<182::aid-dc5>3.0.co;2-f
59. Morgado FN, Schubach A, Rosalino CM, Quintella LP, Santos G, Salgueiro M, et al. Is the In Situ Inflammatory Reaction An Important Tool to Understand the Cellular Immune Response in American Tegumentary Leishmaniasis? Br J Dermatol (2008) 158(1):50–8. doi: 10.1111/j.1365-2133.2007.08255.x
60. Dantas ML, Oliveira JM, Carvalho L, Passos ST, Queiroz A, Guimarães LH, et al. Comparative Analysis of the Tissue Inflammatory Response in Human Cutaneous and Disseminated Leishmaniasis. Mem Inst Oswaldo Cruz (2014) 109(2):202–9. doi: 10.1590/0074-0276130312
61. Boaventura VS, Santos CS, Cardoso CR, de Andrade J, Dos Santos WL, Clarêncio J, et al. Human Mucosal Leishmaniasis: Neutrophils Infiltrate Areas of Tissue Damage That Express High Levels of Th17-related Cytokines. Eur J Immunol (2010) 40(10):2830–6. doi: 10.1002/eji.200940115
62. Christensen SM, Dillon LA, Carvalho LP, Passos S, Novais FO, Hughitt VK, et al. Meta-Transcriptome Profiling of the Human-Leishmania Braziliensis Cutaneous Lesion. PloS Negl Trop Dis (2016) 10(9):e0004992. doi: 10.1371/journal.pntd.0004992
Keywords: PD-L1, neutrophils, skin, Leishmania, human cutaneous leishmaniasis, murine leishmaniasis, supression
Citation: da Fonseca-Martins AM, de Souza Lima-Gomes P, Antunes MM, de Moura RG, Covre LP, Calôba C, Rocha VG, Pereira RM, Menezes GB, Gomes DCO, Saraiva EM and de Matos Guedes HL (2021) Leishmania Parasites Drive PD-L1 Expression in Mice and Human Neutrophils With Suppressor Capacity. Front. Immunol. 12:598943. doi: 10.3389/fimmu.2021.598943
Received: 26 August 2020; Accepted: 13 April 2021;
Published: 15 June 2021.
Edited by:
Fabienne Tacchini-Cottier, University of Lausanne, SwitzerlandReviewed by:
Jaqueline França-Costa, Federal University of Bahia, BrazilHira Nakhasi, Center of Biologics Evaluation and Research (FDA), United States
Copyright © 2021 da Fonseca-Martins, de Souza Lima-Gomes, Antunes, de Moura, Covre, Calôba, Rocha, Pereira, Menezes, Gomes, Saraiva and de Matos Guedes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elvira M. Saraiva, esaraiva@micro.ufrj.br; Herbert L. de Matos Guedes, herbert@biof.ufrj.br; herbert@micro.ufrj.br; herbert@ioc.fiocruz.br
†These authors share senior authorship