 Paula Constanza Arriola Benitez1
Paula Constanza Arriola Benitez1 Ayelén Ivana Pesce Viglietti1
Ayelén Ivana Pesce Viglietti1 Marco Tulio R. Gomes2
Marco Tulio R. Gomes2 Sergio Costa Oliveira2
Sergio Costa Oliveira2 Jorge Fabián Quarleri3
Jorge Fabián Quarleri3 Guillermo Hernán Giambartolomei1
Guillermo Hernán Giambartolomei1 María Victoria Delpino1*
María Victoria Delpino1*- 1Instituto de Inmunología, Genética y Metabolismo (INIGEM), Universidad de Buenos Aires, CONICET, Buenos Aires, Argentina
- 2Department of Biochemistry and Immunology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil
- 3Instituto de Investigaciones Biomédicas en Retrovirus y Sida (INBIRS), Universidad de Buenos Aires, CONICET, Buenos Aires, Argentina
In human brucellosis, the liver is frequently affected. Brucella abortus triggers a profibrotic response on hepatic stellate cells (HSCs) characterized by inhibition of MMP-9 with concomitant collagen deposition and TGF-β1 secretion through type 4 secretion system (T4SS). Taking into account that it has been reported that the inflammasome is necessary to induce a fibrotic phenotype in HSC, we hypothesized that Brucella infection might create a microenvironment that would promote inflammasome activation with concomitant profibrogenic phenotype in HSCs. B. abortus infection induces IL-1β secretion in HSCs in a T4SS-dependent manner. The expression of caspase-1 (Casp-1), absent in melanoma 2 (AIM2), Nod-like receptor (NLR) containing a pyrin domain 3 (NLRP3), and apoptosis-associated speck-like protein containing a CARD (ASC) was increased in B. abortus-infected HSC. When infection experiments were performed in the presence of glyburide, a compound that inhibits NLRP3 inflammasome, or A151, a specific AIM2 inhibitor, the secretion of IL-1β was significantly inhibited with respect to uninfected controls. The role of inflammasome activation in the induction of a fibrogenic phenotype in HSCs was determined by performing B. abortus infection experiments in the presence of the inhibitors Ac-YVAD-cmk and glyburide. Both inhibitors were able to reverse the effect of B. abortus infection on the fibrotic phenotype in HSCs. Finally, the role of inflammasome in fibrosis was corroborated in vivo by the reduction of fibrotic patches in liver from B. abortus-infected ASC, NLRP, AIM2, and cCasp-1/11 knock-out (KO) mice with respect to infected wild-type mice.
Introduction
Human brucellosis is a zoonosis that induces a chronic and debilitating disease caused by Brucella species that manifests itself with a broad clinical spectrum (1, 2). Liver involvement in human brucellosis is usually documented, given the well-characterized tropism of Brucella for the reticuloendothelial system (1, 2). The incidence of liver involvement in active brucellosis has ranged from 5 to 53% or more (2).
Inflammasome activation has been documented in several liver diseases. Accordingly, it has been postulated that the upregulation of IL-1β and IL-18 secretion leads to myofibroblast differentiation with concomitant increase of collagen and TGF-β expression (3). In addition, it was established that inflammasome components are present in hepatic stellate cells (HSCs) and could regulate their function (3). The consequences of activation of inflammasome pathway were also confirmed in vivo, demonstrating its key role in liver fibrosis (4).
The activation and release of IL-1β and IL-18 requires two distinct signals. TLR engagement by pathogen or endogenous signal induces the expression of the precursor forms of these cytokines (pro–IL-1β and pro–IL-18), after which NLR-dependent activation of caspase-1 regulates their proteolytic processing and release (5). Activation of inflammasome by Brucella abortus infection has been previously demonstrated in bone marrow-derived macrophages and dendritic cells (6, 7). In these cells, B. abortus induces the secretion of IL-1β, in a process in which NLRP3 is necessary for activation of ASC inflammasome and the concomitant activation of caspase-1 and maturation and secretion of IL-1β (6, 7). In addition, ASC inflammasomes are also essential for IL-1β secretion induced by B. abortus infection in astrocytes and microglia (8). The first signal can be triggered by various pathogen-associated molecular patterns (PAMPs) via TLR activation. In the case of B. abortus infection inflammasome activation, the second signal involved the presence of a functional type 4 secretion system (T4SS) and DNA-sensing inflammasome receptor AIM2, in bone marrow-derived macrophages, and Mal/TIRAP and TLR-2 are the main signaling involved in astrocytes and microglia (8).
Previously, we have demonstrated that upon infection of HSCs, B. abortus triggers a profibrotic response characterized by inhibition of MMP-9 secretion inducing concomitant collagen deposition and transforming growth factor (TGF)-β1 secretion in a way that involves a functional T4SS and its effectors protein BPE005 (9). Taking into account that inflammasome has been documented to be necessary to induce activation to a fibrotic phenotype of HSCs, we hypothesized that Brucella infection might create a microenvironment that would promote inflammasome activation and concomitant profibrogenic phenotype in HSCs. The results of the study are presented here.
Materials and Methods
Bacterial Culture
Brucella abortus S2308 DsRed-expressing B. abortus S2308 or the isogenic B. abortus virB10 polar mutants were grown overnight in 10 ml of tryptic soy broth (Merck, Buenos Aires, Argentina) with constant agitation at 37°C. Bacteria were harvested and the inocula were prepared as described previously (10).
To obtain heat-killed B. abortus (HKBA), bacteria were washed five times for 10 min each in sterile PBS, heat killed at 70°C for 20 min, aliquoted, and stored at −70°C until they were used. The total absence of B. abortus viability after heat killing was verified by the absence of bacterial growth on tryptose soy agar.
All live Brucella manipulations were performed in biosafety level 3 facilities located at the Instituto de Investigaciones Biomédicas en Retrovirus y SIDA (INBIRS).
Cell Culture
LX-2 cell line, a spontaneously immortalized human HSC line, was kindly provided by Dr. Scott L. Friedman (Mount Sinai School of Medicine, New York, NY). LX-2 cells were maintained in DMEM (Life Technologies–Invitrogen, Carlsbad, CA, USA) and supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2% (v/v) fetal bovine serum (FBS; Gibco–Invitrogen, Carlsbad, CA, USA). All cultures were grown at 37°C and 5% CO2.
Cellular Infection
LX-2 cells were seeded in 24-well-plates and infected with B. abortus S2308, DsRed-expressing B. abortus S2308, or its isogenic mutants at multiplicities of infection (MOI) of 100 and 1000. After the bacterial suspension was dispensed, the plates were centrifuged for 10 min at 2,000 rpm and then incubated for 2 h at 37°C under a 5% CO2 atmosphere. Cells were extensively washed with DMEM to remove extracellular bacteria and incubated in medium supplemented with 100 μg/ml gentamicin and 50 μg/ml streptomycin to kill extracellular bacteria. LX-2 cells were harvested at different times to determine cytokine production, MMP secretion, and collagen deposition.
Neutralization Experiments
Neutralization experiments were performed with 5 μM of Bay 11-7082, an inhibitory compound of the nuclear factor-κB (NF-κB), 50 μM of glybenclamide (glyburide), an inhibitor of the NLRP3 inflammasome, 50 μM of general caspase inhibitor Z-VAD-FMK, or 50 μM of caspase 1 inhibitor Y-VAD-FMK (all inhibitors were from Sigma-Aldrich). The cells were treated for 1 h with each inhibitor before infection.
AIM2 inflammasome complex formation was prevented using 3 μM of A151, a DNA sequence that inhibits in a competitive manner the immunostimulatory DNA. A151 (5-TTAGGGTTAGGGTTAGGGTTAGGG-3) and the control C151 (5-TTCAAATTCAAATTCAAATTCAAA-3) constructs were synthesized with a phosphorothioate backbone. To determine the implication of IL-1β, neutralization experiments were performed by adding 50 ng/ml of ANAKINRA, the inhibitor of IL-1 receptor, and the natural antagonist IL-1Ra (R&D Systems). Recombinant human IL-1β (rIL-1β, R&D Systems) at a concentration of 50 ng/ml was used as a positive control.
mRNA Preparation and Quantitative PCR
RNA from LX-2 cells was isolated using the Quick-RNA MiniPrepKit (Zymo Research) and 1 μg of RNA was subjected to reverse transcription using Improm-II Reverse Transcriptase (Promega). PCR analysis was performed with StepOne real-time PCR detection system (Life Technology) using SYBR Green as fluorescent DNA binding dye. The primer sets used for amplification were as follows: β-actin sense: 5′-AACAGTCCGCCTAGAAGCAC-3′, β-actin antisense: 5′-CGTTGACATCCGTAAAGACC-3′; NLRP3 sense: 5′-CCACAAGATCGTGAGAAAACCC-3′; NLRP3 antisense: 5′-CGGTCCTATGTGCTCGTCA-3′; IL-1β sense: 5′-AGCTACGAATCTCCGACCAC-3′; IL-1β antisense: 5′-CGTTATCCCATGTGTCGAAGAA-3′; ASC sense: 5′-TGGATGCTCTGTACGGGAAG-3′; ASC antisense: 5′-CCAGGCTGGTGTGAAACTGAA-3′; Capase-1 sense: 5′-TTTCCGCAAGGTTCGATTTTCA-3′ Caspase-1 antisense: 5′-GGCATCTGCGCTCTACCATC-3′; AIM2 sense: 5′-TGGCAAAACGTCTTCAGGAGG-3′; AIM2 antisense: 5′-AGCTTGACTTAGTGGCTTTGG-3′.
The amplification cycle for Caspase-1, ASC, and β-actin was 95°C for 15 s, 58°C for 30 s, and 72°C for 60 s; the amplification cycle for IL-1β, NLRP3, and AIM2 was 95°C for 15 s, 61°C for 30 s, and 72°C for 60 s. All primer sets yielded a single product of the correct size. Relative expression levels were normalized against β-actin.
Immunofluorescence
LX-2 cells were infected with B. abortus, and after 24 h, cells were fixed in 4% paraformaldehyde for 10 min at room temperature, permeabilized with 0.3% Triton X-100 (Roche Diagnostics GmbH, Mannheim, Germany) for 10 min, and blocked with PBS containing 1% BSA for 1 h. Infected cells were stained with mouse anti-ASC (Santa Cruz Biotechnology) diluted in 0.1% PBS–Tween 20 overnight at 4°C. Cells then were incubated with rabbit anti-mouse Alexa Fluor 488 (Molecular Probes, Life Technologies) diluted in 0.1% PBS–Tween for 4 h at room temperature. 4,6-Diamidine-2-phenylindole (DAPI) was used for nuclear staining, and cells were stained for 30 min at room temperature. After washing in PBS, cells were mounted and then analyzed by fluorescence microscopy. Confocal images were analyzed using FV-1000 confocal microscope with an oil immersion Plan Apochromatic 60 × NA1.42 objective (Olympus).
Zymography
Gelatinase activity was assayed by the method of Hibbs et al. with modifications, as described (11–13). Briefly, a total of 20 μl of cell culture supernatants from infected LX-2 cells cultured in the presence or not of the inhibitors Bay 11-7082, glyburide, Y-VAD-FMK, A151, control C151, and ANAKINRA at the concentrations mentioned above was mixed with 5 μl of 5 × loading buffer [0.25 M Tris (pH 6.8), 50% glycerol, 5% SDS, and bromophenol blue crystals] and loaded onto 10% SDS-PAGE gels containing 1 mg/ml gelatin (Sigma-Aldrich, Buenos Aires, Argentina). After electrophoresis, gels were washed with a solution containing 50 mM Tris–HCl (pH 7.5) and 2.5% Triton X-100 (buffer A) for 30 min and with buffer A added with 5 mM CaCl2 and 1 μM ZnCl2 for 30 min and were later incubated with buffer A with an additional 10 mM CaCl2 and 200 mM NaCl for 48 h at 37°C. This denaturation/renaturation step promotes MMP activity without the proteolytic cleavage of pro-MMP. Gelatin activity was visualized by the staining of the gels with 0.5% Coomassie blue. Unstained bands indicated the presence of gelatinase activity, and their positions in the gel indicate the molecular weights of the enzymes involved.
Measurement of Cytokine Concentrations
Secretion of TGF-β1 and IL-1β in the supernatants was quantified by ELISA (BD Biosciences).
Assessment of Collagen Deposition—Sirius Red Staining
Collagen deposition was quantified using Sirius red (Sigma-Aldrich, Argentina SA), a strong anionic dye that binds strongly to collagen molecules (14). Sirius red staining was performed as was described (15). Briefly, Sirius red was dissolved in saturated aqueous picric acid at a concentration of 0.1%. Bouin's fluid (for cell fixation) was prepared by mixing 15 ml saturated aqueous picric acid with 5 ml of 35% formaldehyde and 1 ml of glacial acetic acid. Cell layers were fixed with 1 ml of Bouin's fluid for 1 h. Afterwards, culture plates were washed three times with deionized water. Culture dishes were air dried before adding 1 ml of Sirius red dye reagent. Cells were stained for 18 h with mild shaking. The stained cell layers were extensively washed with 0.01 N hydrochloric acid to remove all unbound dye. The stained material was dissolved in 0.2 ml of 0.1 N sodium hydroxide by shaking for 30 min. The dye solution was transferred to microtiter plates, and OD was measured using a microplate reader (Thermo Scientific) at 550 nm against 0.1 N sodium hydroxide as a blank.
Lipoproteins and LPS
Brucella abortus lipidated outer membrane protein 19 (LOmp19) and unlipidated Omp19 (U-Omp19) were obtained as described (16). Both contained <0.25 endotoxin U/μg of protein as assessed by Limulus Amebocyte Lysates (Associates of Cape Cod). B. abortus S2308 LPS and Escherichia coli O111k58H2 LPS were provided by I. Moriyon. The synthetic lipohexapeptide (tripalmitoyl S-glyceryl-Cys-Ser- Lys4-OH [Pam3Cys]) was purchased from Boehringer Mannheim (Mannheim, Germany).
DNA From B. abortus
Brucella abortus DNA was purified using the kit Wizard® Genomic DNA (Promega) following the instructions of the manufacturer. Brucella DNA was measured spectrophotometrically. Transient transfections of LX-2 cells with 2 μg/ml of B. abortus DNA were carried out using Lipofectamine 2000 (Invitrogen), following the manufacturer's instructions. After the purification step, an aliquot of 100 μg of DNA was treated with DNase I (1 U/mg) DNA (Zymo Research) according to the instructions of the manufacturer.
Hepatic Fibrosis in a Mouse Model
Mouse strains used in this study included apoptosis-associated speck-like protein containing a CARD (ASC), Nod-like receptor (NLR) containing a pyrin domain 3 (NLRP3), absent in melanoma 2 (AIM2), and caspase-1 (Casp-1)/11 knock-out (KO) mice, as described previously (8), and C57BL/6 wild-type (WT) mice (provided by Federal University of Minas Gerais, Belo Horizonte, Brazil). Six- to eight-week-old mice were infected through the intraperitoneal route with 5 × 105 CFU of B. abortus S2308. Mice were born from breeding pairs that were housed under controlled temperature (22 ± 2°C) and artificial light under a 12-h cycle period. Mice were kept under specific pathogen-free conditions in positive-pressure cabinets and provided with sterile food and water ad libitum.
All animal procedures were performed according to the rules and standards for the use of laboratory animals of the National Institutes of Health. Animal experiments were approved by the Institutional Committee for the Care and Use of Laboratory Animals (CICUAL, permit number: 287/2015). Histological examination of liver was carried out at week 4 post-infection after routine fixation and paraffin embedding. Five-micrometer-thick sections were cut and stained with Masson's trichrome stain. Masson's trichrome staining was conducted according to the manufacturer's instructions (Sigma-Aldrich). Collagen-positive areas were visualized by light microscopy and quantified using Image Pro-Plus 6.0 software (Media Cybernetics, Inc.).
Statistical Analysis
Statistical analysis was performed with one-way ANOVA, followed by post-hoc Tukey test (a single-step multiple comparison statistical test that finds means that are significantly different from each other) using GraphPad Prism 4.0 software. Data were presented as mean ± SEM.
Results
B. abortus Infection Induces IL-1β Secretion by LX-2 Cells via T4SS
It has been established that inflammasome components are present in HSCs and could regulate their function (3). To determine if B. abortus infection induces inflammasome activation, LX-2 cells were infected with B. abortus and the secretion of IL-1β was evaluated in culture supernatants by ELISA 24 h post-infection. B. abortus infection induces the secretion of IL-1β by LX-2 cells (Figure 1A). The T4SS encoded by virB genes was first involved in the capacity of Brucella to establish an intracellular replication niche in several cell types (17). In addition, this system has been involved in the induction of inflammatory response during B. abortus infection (18) and also in the inflammasome signaling activation (7). Therefore, experiments were conducted to determine if the T4SS could be involved in the secretion of IL-1β induced by B. abortus in LX-2 cells. To this end, LX-2 cells were infected with B. abortus and its isogenic B. abortus virB10 mutant, and IL-1β secretion induced by B. abortus was dependent on the expression of a functional T4SS, since the levels or IL-1β did not differ significantly between LX-2 cells infected with B. abortus virB10 mutant and uninfected controls (Figure 1A). Taken together, our results indicated that B. abortus infection induces IL-1β secretion in a mechanism that is dependent on the presence of a functional T4SS.
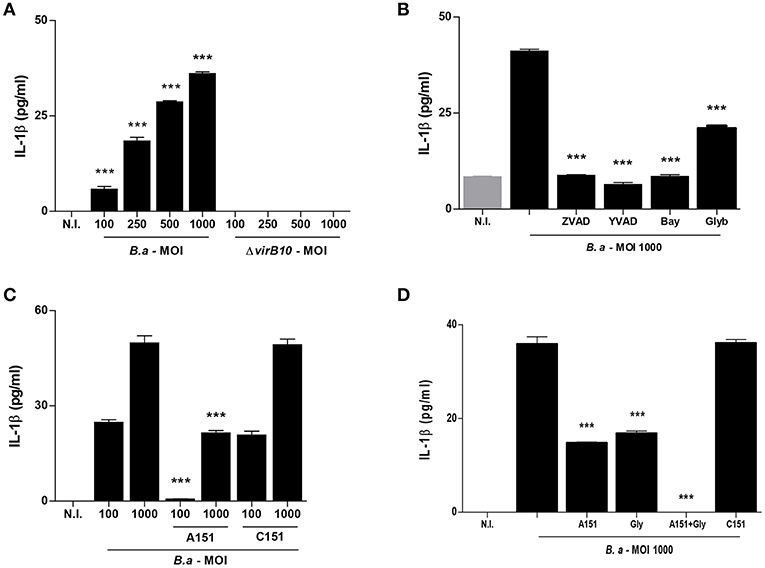
Figure 1. B. abortus infection induces IL-1β in a VirB-dependent manner. LX-2 cells were infected with B. abortus (B.a) and its virB10 isogenic mutant (ΔvirB10) at an MOI of 100–1000 and 24 h post-infection; IL-1β secretion was determined by ELISA in culture supernatants (A). Effect of Z-VAD-FMK (ZVAD), Y-VAD-FMK (YVAD), Bay 11-7082 (Bay) and glybenclamide (Glyb) during B. abortus infection (MOI 1000) on IL-1β secretion (B). Effect of A151 and the control 151 (C151) during B. abortus infection on IL-1β secretion (C). Effect of Glyb plus A151 on IL-1βsecretion during B. abortus infection (MOI 1000). Data are given as the means ± SD from at least three individual experiments. ***P < 0.001 vs. cells infected with ΔvirB10 (A) or vs. infected and untreated cells (B–D).
IL-1β Secretion Induced by B. abortus Infection Is Dependent on Caspase-1 and NLRP3
Caspase-1 plays a fundamental role in innate immunity as the protease that activates the pro-inflammatory cytokines pro-IL-1β and pro-IL-18. Caspase-1 itself is activated in different inflammasome complexes; however, activation of the NLRP3 inflammasome has been frequently implicated in the development of fibrosis (19). To determine the role of caspase-1 and NLRP3 in IL-1β secretion by B. abortus-infected LX-2 cells, we performed the infection of LX-2 cells in the presence of specific pharmacological inhibitors. Inhibition of caspase-1 using the general caspase inhibitor Z-VAD-FMK or the specific caspase-1 inhibitor Ac-YVAD-cmk completely abrogated the secretion of IL-1β secretion induced by B. abortus infection of LX-2 cells (Figure 1B). When infection experiments were performed in the presence of Bay compound that inhibits NFκB, the secretion of IL-1β was significantly inhibited. However, glyburide, a compound that inhibits NLRP3, partially inhibits IL-1β secretion with respect to untreated cells (Figure 1B). Taken together, these results indicated that caspase-1 and NLRP3 are involved in the secretion of IL-1β by B. abortus-infected LX-2 cells.
IL-1β Secretion Induced by B. abortus Infection Is Also Dependent on AIM2
AIM2 inflammasome has been previously involved in the induction of IL-1β secretion in a T4SS-dependent manner during B. abortus infection of bone marrow derived macrophages and dendritic cells (6, 7). Since the inhibition of NLRP3 did not completely abrogate the secretion of IL-1β in response to B. abortus infection, experiments were conducted to determine whether AIM2 inflammasome is also involved in caspase-1 activation. To this end, LX-2 cells were infected in the presence of A151, the oligodeoxinucleotide sequence that inhibits AIM2 or the sequence control C151. Our results indicated that AIM2 inflammasome contributes to IL-1β production by B. abortus-infected LX-2 cells, since the secretion of IL-1β was significantly inhibited when cells were treated with A151 with respect to untreated cells or cells treated with C151 (Figure 1C). When infection experiments were performed in the presence of A151 and glyburide administered in conjunction, the production of IL-1β was completely abrogated (Figure 1D). Taken together, these results indicated that NLRP3 and AIM2 inflammasomes are involved in the secretion of IL-1β by B. abortus-infected LX-2 cells.
B. abortus Infection Induces ASC, NLRP3, AIM2, and Caspase-1 mRNA Expression and ASC Speck Formation in LX-2 Cells
The basal AIM2 expression was sufficient to initiate inflammasome activation (20), but NLRP3 upregulation is necessary to initiate the activation of inflammasome (21, 22). Then, experiments were conducted to determine whether expression of inflammasome components could be upregulated during B. abortus infection. To this end, we determine the mRNA transcription of ASC, NLRP3, AIM2, and caspase-1 by RT-qPCR. Our results indicated that B. abortus infection induces an increase in ASC, NLRP3, AIM2, and caspase-1 mRNA transcription in LX-2 cells (Figures 2A–D). Most inflammasomes require oligomerization of ASC and thus the presence of ASC specks is a direct evidence of inflammasome activation. After infection with B. abortus, the formation of ASC specks was detected using specific antibodies by a fluorescence microscope. ASC specks were formed in B. abortus-infected LX-2 cells, but these structures were not detectable in non-infected cells. This indicates that B. abortus infection induces inflammasome assembly and consequently ASC speck formation (Figure 2E).
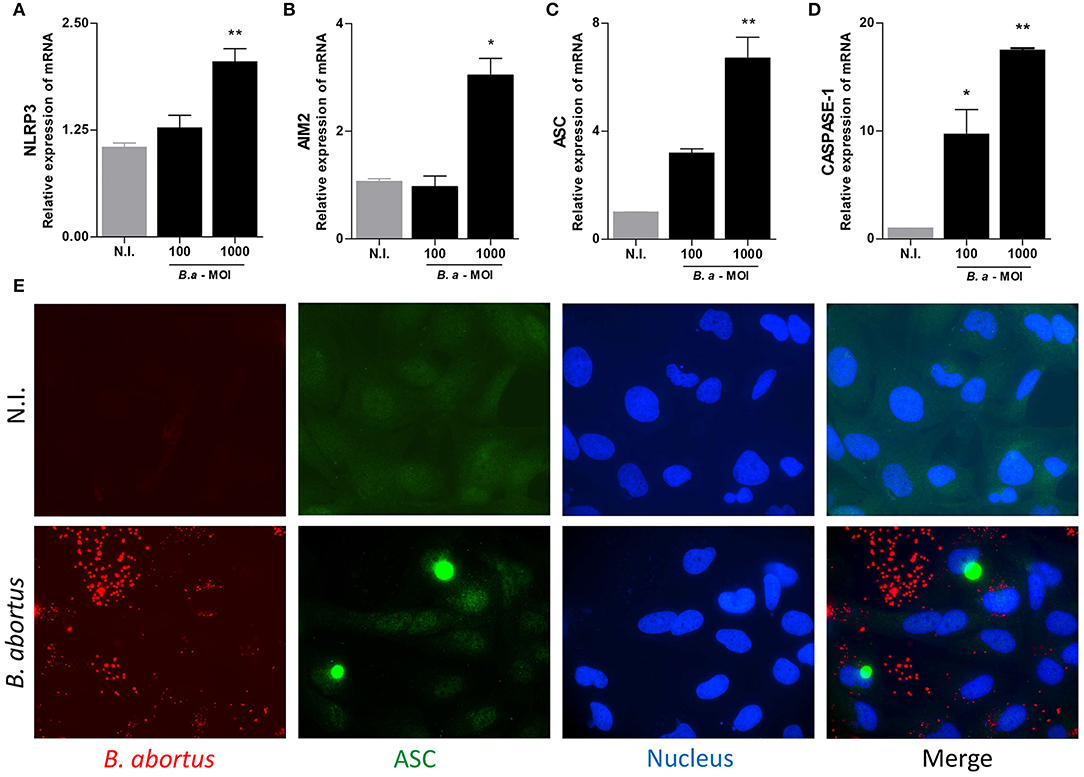
Figure 2. B. abortus induces inflammasome in LX-2 cells. LX-2 cells were infected with B. abortus at MOI of 100 and 1000; at 24 h, post-infection levels of NLRP3 (A), AIM2 (B), ASC (C), Caspase 1 (D), and IL-1β (E) were determined by RT-qPCR. LX-2 cells were infected with Ds-Red B. abortus and ASC speck were revealed by immunofluorescence with a specific antibody labeled with Alexa 488. Nuclei were stained with DAPI. Data are given as the means ± SD from at least three individual experiments. *P < 0.05; **P < 0.01 vs. non-infected cells (N.I.).
Brucella DNA Induces IL-1β Secretion in LX-2 Cells
IL-1β secretion was dependent on bacteria viability since stimulation of LX-2 cells with heat-killed B. abortus (HKBA) was unable to induce IL-1β (Figure 3A).
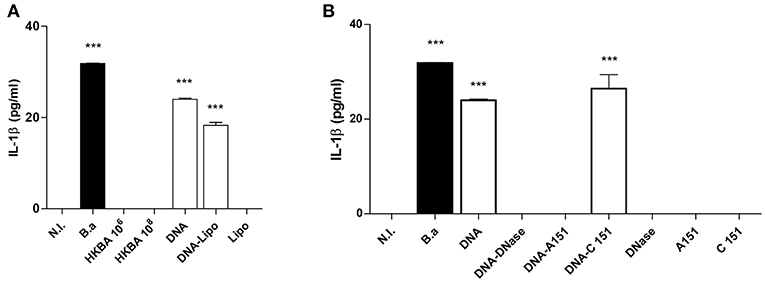
Figure 3. DNA from B. abortus induces IL-1β. (A) LX-2 cells were infected with B. abortus (B.a) at MOI 1000 or treated with heat-killed B. abortus (HKBA) (1 × 106 and 1 × 108 bacteria/ml), 2 μg/ml of DNA, DNA and lipofectamine (DNA + Lipo), and lipofectamine alone as control (Lipo); IL-1β secretion was measured in culture supernatant after 24 h by ELISA. Determination of IL-1β in culture supernatants from LX-2 cells treated with DNA, DNA and A151, DNA and C151 as control, DNA treated with DNAse I, and DNAse I alone as control (B). Data are given as the means ± SD from at least three individual experiments. ***P < 0.001 vs. non-infected cells (N.I.).
It has been recently demonstrated that Brucella DNA is involved in IL-1β secretion via activation of caspase-1 through the AIM2 inflammasome in macrophages and dendritic cells (6, 7). Therefore, we decided to test whether Brucella genomic DNA could be the putative ligand for AIM2 in the context of the inflammasome activation in LX-2 cells. To this end, Brucella DNA was transfected into LX-2 cells using lipofectamine or added to the culture medium to determine IL-1β secretion. Brucella DNA induced IL-1β secretion by LX-2 cells when it was added to the culture medium and also in transfected cells (Figure 3A). To determine if AIM2 inflammasome is involved in the secretion of IL-1β induced by Brucella DNA, experiments were performed in the presence of A151, the oligodeoxinucleotide that inhibits AIM2 or its oligodeoxinucleotide control C151. AIM2 is involved in the secretion of IL-1β induced by Brucella DNA, since A151 abrogated its secretion (Figure 3B). Additionally, DNase I treatment significantly reduced or abrogated Brucella DNA-induced IL-1β secretion (Figure 3B), demonstrating that bacterial DNA participates or is a major agonist that activates the inflammasome.
PAMPs Associated to Inflammasome Activation
For the production of IL-1β, PAMPs via TLRs and NLRs function in concert. PAMPs induce the expression of the precursor form of this cytokine (pro-IL-1β), and NLR-dependent CASP-1 activation induces its proteolytic processing and release (5). Hence, to assess the role of Brucella PAMPs, LX-2 cells were stimulated with HKBA and mRNA expression of IL-1β was determined by RT-qPCR. Our results indicated that HKBA was able to induce IL-1β expression at the mRNA level (Figure 4A) without IL-1β release, which demonstrated that viability is crucial for IL-1β protein production.
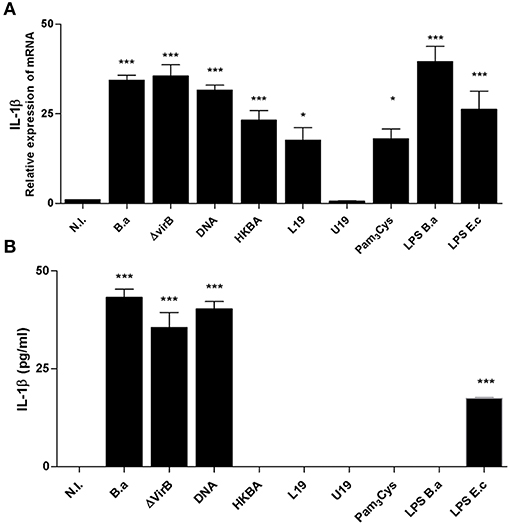
Figure 4. L-Omp19 induces mRNA of IL-1β. (A) LX-2 cells were infected with B. abortus (B.a) and its virB10 isogenic mutant (ΔvirB10) at MOI 1000, or incubated with DNA (2 μg/ml), heat-killed B. abortus (HKBA) (1 × 106 and 1 × 108 bacteria/ml), L-Omp19 (1,000 ng/ml), U-Omp19 (1,000 ng/ml), B. abortus LPS (1,000 ng/ml), Pam3Cys (50 ng/ml), or E. coli LPS (100 ng/ml). Determination of IL-1β mRNA by RT-qPCR (A) and IL-1β secretion by ELISA (B). Data are given as the means ± SD from at least three individual experiments. *P < 0.05; ***P < 0.001 vs. non-infected cells (N.I.).
Previous observations indicated that LPS and lipoproteins from B. abortus are crucial for inflammatory responses induced by B. abortus in different models in vivo and in vitro (16, 23–26). Cells were then incubated with LPS from B. abortus and lipidated Omp19 (L-Omp19) as a Brucella lipoprotein model, and the expression of IL-1β mRNA was determined by RT-qPCR. Our results indicated that L-Omp19 and LPS induce an increase in IL-1β mRNA expression in LX-2 cells. IL-1β mRNA expression induced by Omp19 was dependent on the lipid moiety of the molecule because unlipidated Omp19 (U-Omp19) did not induce IL-1β mRNA expression. The requirement for lipidation was further supported by the fact that Pam3Cys, a lipohexapeptide with an irrelevant peptide sequence, also induced the production of mRNA of IL-1β (Figure 4A). As expected, the presence of IL-1β at the protein level was not detected in supernatants of cultures of LX-2 cells treated with LPS or L-Omp19 (Figure 4B). Taken together, these results indicated that B. abortus lipoproteins and LPS induce mRNA of IL-1β in LX-2 cells.
The Inflammasome Pathway Is Involved in the Profibrogenic Response of LX-2 Cells Upon B. abortus Infection
It has been demonstrated that inflammasome activation has a variety of functional consequences for HSCs, including enhanced collagen 1 and TGF-β expression (4). Previously, we have demonstrated that upon infection of LX-2 cells, B. abortus inhibits MMP-9 secretion and induces concomitant collagen and TGF-β1 secretion (15). Therefore, experiments were conducted to determine the role of the inflammasome in the induction of a fibrogenic phenotype in LX-2 cells during B. abortus infection. To this end, the levels of secretion of MMP-9, TGF-β, and collagen deposition were determined in LX-2 cells infected with B. abortus in the presence of different inflammasome inhibitors. When infection experiments were performed in the presence of YVAD, glyburide, or A151, the effect of B. abortus infection on MMP-9 expression, collagen deposition, and TGF-β secretion on LX-2 cells was partially reversed with respect to untreated infected cells (Figure 5). These results indicated that NLRP3 and AIM2 inflammasome are involved in the induction of profibrogenic phenotype in LX-2 cells.
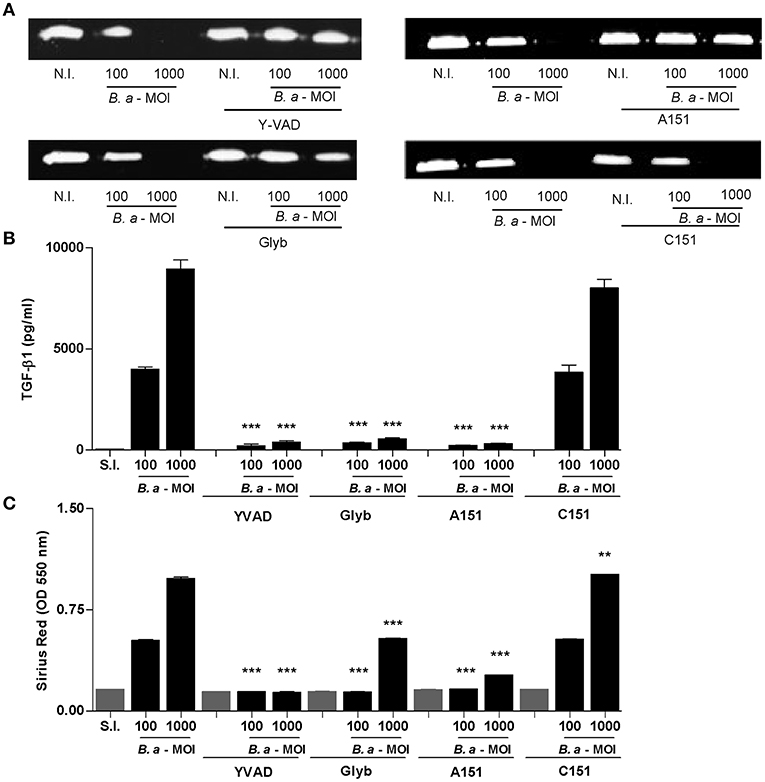
Figure 5. Inflammasome is involved in profibrogienic response of LX-2 cells. LX-2 cells were infected with B. abortus at MOI of 100 and 1000 in the presence or not of Z-VAD-FMK (ZVAD), Y-VAD-FMK (YVAD), Bay 11-7082 (Bay), and glybenclamide (Glyb). At 24 h after infection, supernatants were harvested to analyze MMP-9 production by zymography (A) and TGF-β1 secretion by ELISA (B). Quantification of collagen deposition was revealed by Sirius red staining by OD readings at 550 nm at 10 days post-infection (C). Data are given as the means ± SD of duplicates. **P < 0.01; ***P < 0.001 vs. non-infected cells (N.I.).
IL-1β Is Involved in the Induction of a Profibrogenic Phenotype
Caspase-1 is required not only for IL-1β secretion but also for IL-18 secretion. To determine the role of IL-1β in the induction of a fibrotic phenotype, infection experiments were performed in the presence of the inhibitor of IL-1 receptor, the natural antagonist IL-1Ra (ANAKINRA). As shown in Figure 6, ANAKINRA abrogated the ability of B. abortus to inhibit MMP-9 and to induce collagen deposition. These results indicate that IL-1β could be a key cytokine during inflammasome activation involved in the fibrogenic phenotype triggered by B. abortus infection.
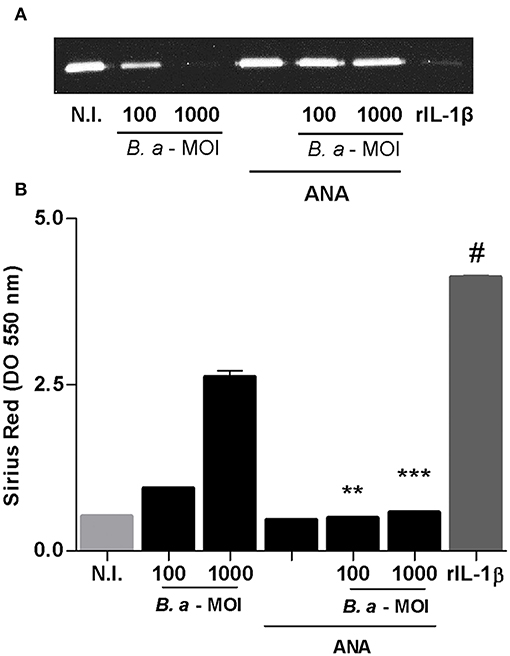
Figure 6. IL-1β is involved in the induction of a profibrogenic phenotype. LX-2 cells were infected with B. abortus at MOI of 100 and 1000 in the presence or not of ANAKINRA (ANA). Determination of MMP-9 at 24 h post-infection in culture supernatants by zymography (A). Recombinant human IL-1-β, (rIL-1β) (50 ng/ml) was used as a positive control. Quantification of collagen deposition was revealed by Sirius red staining by OD readings at 550 nm at 10 days post-infection (B). Data are given as the means ± SD of duplicates. **P < 0.01; ***P < 0.001 vs. untreated cells. #P < 0.001 vs. non-infected cells (N.I.).
NLRP3 and AIM2 Influence Liver Fibrosis in Livers From B. abortus-Infected Mice
Finally, to verify the in vivo significance of our hypothesis, Casp-1, ASC, NLRP3, and AIM2 KO mice and WT mice, as control, were infected with B. abortus, and 4 weeks later, animals were sacrificed to determine the role of inflammasome in the liver fibrosis. Accordingly with our previous results, Masson's trichrome staining revealed the presence of fibrotic patch in livers from B. abortus-infected mice with respect to uninfected control (9, 15). In contrast, Casp-1, ASC, NLRP3, and AIM2 KO animals presented a significant reduction in the fibrotic patch (Figure 7). No fibrotic patch was observed in mice inoculated with saline (data not shown). These results indicated that inflammasomes NLRP2 and AIM2 play a key role in the modulation of fibrosis during B. abortus infection.
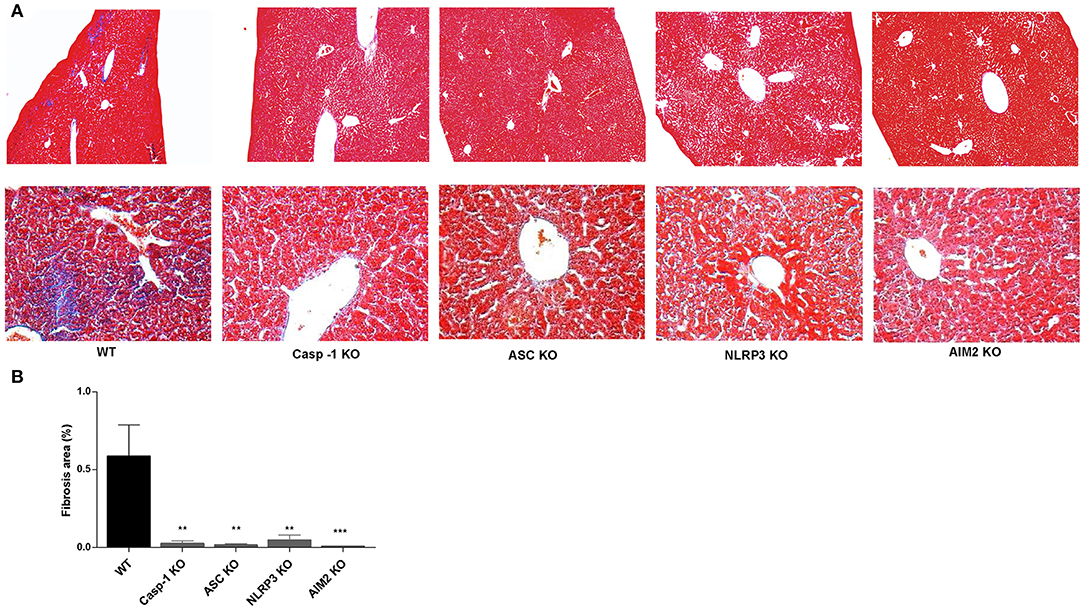
Figure 7. NLRP3 and AIM2 influence liver fibrosis in livers from B. abortus-infected mice. Representative photomicrographs of liver sections from control mice, B. abortus-infected mice WT, Casp-1, ASC, NLRP3, and AM2 KO mice (n5) stained with Masson's trichrome staining. The top panels show images taken at the original magnification (×100), and the bottom panels show a detail of collagen patches (magnification, ×400) (A). Collagen-positive areas were quantified using Image Pro-Plus 6.0 software (B). Data are given as the means ± SD from at least three individual experiments. **P < 0.01; ***P < 0.001 vs. wild type (WT).
Discussion
The liver is frequently affected in patients with active brucellosis, as revealed by the presence of histopathology lesions, such as granulomas, inflammatory infiltrates, and necrosis of liver parenchyma (27, 28).
Fibrosis is an intrinsic response to chronic persistent liver injury that results in a wound-healing process to mitigate the damage, but can also lead to scar formation. Inflammasome activation may play an important role in this process (19).
In the present study, we demonstrated that B. abortus infection activates the inflammasome with concomitant secretion of IL-1β in HSC leading to upregulation of a profibrogenic phenotype.
Inflammasomes have emerged as critical signaling molecules of innate immune system involved in liver fibrosis. Inflammasomes are intracellular multiprotein complexes that act as regulators of inflammation and cell destiny. They respond to several danger signals by activating caspase-1 by the release of proinflammatory cytokines IL-1β and IL-18 (29). In particular, the NLRP3 inflammasome has been frequently implicated in the pathogenesis of chronic inflammatory liver diseases that causes liver fibrosis (30).
HSCs are the main cells involved in extracellular matrix deposition during liver fibrosis (31). In this process, the NLRP3 inflammasome has been involved in the functional changes in HSCs, including upregulation of the expression of collagen and TGF-β (4, 32), in findings that were confirmed by performing the knocking in NLRP3 (33).
Previous studies performed in dendritic cells, macrophages, and glial cells indicate that Brucella is sensed by ASC-dependent inflammasomes, mainly NLRP3 and AIM2, that induce caspase-1 activation with pro-inflammatory response (6, 8, 34, 35). Accordingly, in Brucella-infected HSCs, the secretion of IL-1β depends on NLRP3 and AIM2 inflammasomes. In this context, the participation of PAMPs in the activation of the inflammasome and the secretion of IL-1β must be discussed. The activation and release of IL-1β requires two distinct signals. The first signal can be triggered by various pathogen-associated molecular patterns via TLR activation, which induces the synthesis of pro–IL-1β. The second signal is provided by the activation of the inflammasome and caspase-1 leading to IL-1β processing. During B. abortus infection, the induction of IL-1β at mRNA level was independent of bacterial viability and induced at least by two structural bacterial components, including lipoproteins and LPS. However, the second signal requires bacterial viability and the presence of a functional T4SS and B. abortus DNA. The T4SS is encoded by virB genes that play a main role in Brucella intracellular replication (17), and it has also been involved in the immune response to Brucella infection (7, 18). Bacteria utilize the T4SS to deliver effectors to eukaryotic cells. In LX-2 cells, B. abortus virB10 mutant was unable to induce IL-1β secretion, and it could suggest that Brucella T4SS is involved in the transport of effector molecules that act via NLRP3 and/or AIM2 to activate inflammasomes. This is in agreement with our other finding in which we demonstrate that HKBA does not induce the secretion of IL-1β by LX-2 cells.
In vitro studies have shown that IL-1β promotes the proliferation and myofibroblast transdifferentiation of HSCs with substantial increased levels of their fibrogenic markers (36–38). Activated caspase-1 could also cleave pro-IL-18 into its active form IL-18, and this cytokine has also been involved in fibrosis induction (38, 39). In a murine model of non-alcoholic steatohepatitis, fibrosis was not reversed by IL-1Ra treatment, indicating that other regulators of NLRP3 inflammasome are involved in fibrogenesis promotion (33). However, our experiments performed in B. abortus-infected HSCs in the presence of ANAKINRA, a version of the human interleukin-1 receptor antagonist, indicated that IL-1β secreted by HSCs has a main role in the induction of TGF-β with concomitant collagen deposition and inhibition of MMP-9 secretion. This role of IL-1β in the myofibroblast transdifferentiation with concomitant fibrosis is not exclusive for HSCs; accordingly, it has been previously described in endothelial and epithelial cell transdifferentiation and fibrosis (40–42).
These results and previous findings suggest that the interaction of Brucella with innate immunity in vivo may result in an increase of inflammatory response that results in liver fibrosis. Inflammasomes would dictate this fibrotic phenotype. Consequently, B. abortus infection induces fibrosis in mice that was reduced in mice lacking AIM2 or NLRP3.
Together, these results indicated that upon infection of HSCs, B. abortus triggers AIM2 and NLRP3 inflammasome activation with concomitant IL-1β secretion in a mechanism that is dependent on a functional T4SS and DNA. This IL-1β is implicated in the induction of fibrotic phenotype in HSC.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Ethics Statement
This animal study was reviewed and approved by CICUAL-Facultad de Medicina, Universidad de Buenos Aires.
Author Contributions
MD conceived and designed the experiments, supervised experiments, interpreted the data, and wrote the manuscript. PA, MG, and AP performed the experiments. PA analyzed the data and wrote sections of the manuscript. SO, JQ, and GG supported the work with key suggestions and helped with data interpretation. All authors reviewed the manuscript.
Funding
This work was supported by grants PICT 2014-1111 and PICT 2015-0316 from Agencia Nacional of Promoción Científica y Tecnológica (ANPCYT, Argentina) and by grant PIP 2015-2017-0200 from Consejo Nacional de Investigaciones científicas y técnicas (CONICET). Funding agencies had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Horacio Salomón and the staff of the Instituto de Investigaciones Biomédicas en Retrovirus y Sida (INBIRS) for their assistance with biosafety level 3 laboratory uses. PA and AP are recipients of a fellowship from CONICET. JQ, GG, and MD are members of the Research Career of CONICET.
References
1. Pappas G, Akritidis N, Bosilkovski M, Tsianos E. Brucellosis. N Engl J Med. (2005) 352:2325–36. doi: 10.1056/NEJMra050570
2. Madkour MM. Osteoarticular brucellosis. In: Madkour MM, editor. Madkour's Brucellosis. 2nd ed. Berlin: Springer-Verlag. (2001). p. 74–84. doi: 10.1007/978-3-642-59533-2_8
3. Alegre F, Pelegrin P, Feldstein AE. Inflammasomes in liver fibrosis. Semin Liver Dis. (2017) 37:119–27. doi: 10.1055/s-0037-1601350
4. Watanabe A, Sohail MA, Gomes DA, Hashmi A, Nagata J, Sutterwala FS, et al. Inflammasome-mediated regulation of hepatic stellate cells. Am J Physiol Gastrointest Liver Physiol. (2009) 296:G1248–57. doi: 10.1152/ajpgi.90223.2008
5. Miao EA, Andersen-Nissen E, Warren SE, Aderem A. TLR5 and Ipaf: dual sensors of bacterial flagellin in the innate immune system. Semin Immunopathol. (2007) 29:275–88. doi: 10.1007/s00281-007-0078-z
6. Costa Franco MMS, Marim FM, Alves-Silva J, Cerqueira D, Rungue M, Tavares IP, et al. AIM2 senses Brucella abortus DNA in dendritic cells to induce IL-1beta secretion, pyroptosis and resistance to bacterial infection in mice. Microbes Infect. (2018) 21:85–93. doi: 10.1016/j.micinf.2018.09.001
7. Gomes MT, Campos PC, Oliveira FS, Corsetti PP, Bortoluci KR, Cunha LD, et al. Critical role of ASC inflammasomes and bacterial type IV secretion system in caspase-1 activation and host innate resistance to Brucella abortus infection. J Immunol. (2013) 190:3629–38. doi: 10.4049/jimmunol.1202817
8. Miraglia MC, Costa Franco MM, Rodriguez AM, Bellozi PM, Ferrari CC, Farias MI, et al. Glial cell-elicited activation of brain microvasculature in response to Brucella abortus infection requires ASC inflammasome-dependent IL-1beta production. J Immunol. (2016) 196:3794–805. doi: 10.4049/jimmunol.1500908
9. Arriola Benitez PC, Rey Serantes D, Herrmann CK, Pesce Viglietti AI, Vanzulli S, Giambartolomei GH, et al. The effector protein BPE005 from Brucella abortus induces collagen deposition and matrix metalloproteinase 9 downmodulation via transforming growth factor beta1 in hepatic stellate cells. Infect Immun. (2016) 84:598–606. doi: 10.1128/IAI.01227-15
10. Scian R, Barrionuevo P, Rodriguez AM, Arriola Benitez PC, Garcia Samartino C, Fossati CA, et al. Brucella abortus invasion of synoviocytes inhibits apoptosis and induces bone resorption through RANKL expression. Infect Immun. (2013) 81:1940–51. doi: 10.1128/IAI.01366-12
11. Hibbs MS, Hasty KA, Seyer JM, Kang AH, Mainardi CL. Biochemical and immunological characterization of the secreted forms of human neutrophil gelatinase. J Biol Chem. (1985) 260:2493–500.
12. Scian R, Barrionuevo P, Giambartolomei GH, De Simone EA, Vanzulli SI, Fossati CA, et al. Potential role of fibroblast-like synoviocytes in joint damage induced by Brucella abortus infection through production and induction of matrix metalloproteinases. Infect Immun. (2011) 79:3619–32. doi: 10.1128/IAI.05408-11
13. Scian R, Barrionuevo P, Giambartolomei GH, Fossati CA, Baldi PC, Delpino MV. Granulocyte-macrophage colony-stimulating factor- and tumor necrosis factor alpha-mediated matrix metalloproteinase production by human osteoblasts and monocytes after infection with Brucella abortus. Infect Immun. (2011) 79:192–202. doi: 10.1128/IAI.00934-10
14. Tullberg-Reinert H, Jundt G. In situ measurement of collagen synthesis by human bone cells with a sirius red-based colorimetric microassay: effects of transforming growth factor beta2 and ascorbic acid 2-phosphate. Histochem Cell Biol. (1999) 112:271–6. doi: 10.1007/s004180050447
15. Arriola Benitez PC, Scian R, Comerci DJ, Serantes DR, Vanzulli S, Fossati CA, et al. Brucella abortus induces collagen deposition and MMP-9 down-modulation in hepatic stellate cells via TGF-beta1 production. Am J Pathol. (2013) 183:1918–27. doi: 10.1016/j.ajpath.2013.08.006
16. Giambartolomei GH, Zwerdling A, Cassataro J, Bruno L, Fossati CA, Philipp MT. Lipoproteins, not lipopolysaccharide, are the key mediators of the proinflammatory response elicited by heat-killed Brucella abortus. J Immunol. (2004) 173:4635–42. doi: 10.4049/jimmunol.173.7.4635
17. Comerci DJ, Martinez-Lorenzo MJ, Sieira R, Gorvel JP, Ugalde RA. Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cell Microbiol. (2001) 3:159–68. doi: 10.1046/j.1462-5822.2001.00102.x
18. Roux CM, Rolan HG, Santos RL, Beremand PD, Thomas TL, Adams LG, et al. Brucella requires a functional Type IV secretion system to elicit innate immune responses in mice. Cell Microbiol. (2007) 9:1851–69. doi: 10.1111/j.1462-5822.2007.00922.x
19. Szabo G, Csak T. Inflammasomes in liver diseases. J Hepatol. (2012) 57:642–54. doi: 10.1016/j.jhep.2012.03.035
20. Rathinam VA, Jiang Z, Waggoner SN, Sharma S, Cole LE, Waggoner L, et al. The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat Immunol. (2010) 11:395–402. doi: 10.1038/ni.1864
21. Franchi L, Eigenbrod T, Nunez G. Cutting edge: TNF-alpha mediates sensitization to ATP and silica via the NLRP3 inflammasome in the absence of microbial stimulation. J Immunol. (2009) 183:792–6. doi: 10.4049/jimmunol.0900173
22. Bauernfeind FG, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. (2009) 183:787–91. doi: 10.4049/jimmunol.0901363
23. Zwerdling A, Delpino MV, Barrionuevo P, Cassataro J, Pasquevich KA, Garcia Samartino C, et al. Brucella lipoproteins mimic dendritic cell maturation induced by Brucella abortus. Microbes Infect. (2008) 10:1346–54. doi: 10.1016/j.micinf.2008.07.035
24. Zwerdling A, Delpino MV, Pasquevich KA, Barrionuevo P, Cassataro J, Garcia Samartino C, et al. Brucella abortus activates human neutrophils. Microbes Infect. (2009) 11:689–97. doi: 10.1016/j.micinf.2009.04.010
25. Garcia Samartino C, Delpino MV, Pott Godoy C, Di Genaro MS, Pasquevich KA, Zwerdling A, et al. Brucella abortus induces the secretion of proinflammatory mediators from glial cells leading to astrocyte apoptosis. Am J Pathol. (2010) 176:1323–38. doi: 10.2353/ajpath.2010.090503
26. Oliveira SC, de Oliveira FS, Macedo GC, de Almeida LA, Carvalho NB. The role of innate immune receptors in the control of Brucella abortus infection: toll-like receptors and beyond. Microbes Infect. (2008) 10:1005–9. doi: 10.1016/j.micinf.2008.07.005
27. Akritidis N, Tzivras M, Delladetsima I, Stefanaki S, Moutsopoulos HM, Pappas G. The liver in brucellosis. Clin Gastroenterol Hepatol. (2007) 5:1109–12. doi: 10.1016/j.cgh.2006.08.010
28. Madkour MM. Gastrointestinal brucellosis. Madkour MM, editor. Madkour's Brucellosis. 2nd ed. Berlin: Springer-Verlag (2001). p. 150–8. doi: 10.1007/978-3-642-59533-2_13
29. Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat Rev Immunol. (2013) 13:397–411. doi: 10.1038/nri3452
30. Henao-Mejia J, Elinav E, Jin C, Hao L, Mehal WZ, Strowig T, et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature. (2012) 482:179–85. doi: 10.1038/nature10809
32. Mederacke I, Hsu CC, Troeger JS, Huebener P, Mu X, Dapito DH, et al. Fate tracing reveals hepatic stellate cells as dominant contributors to liver fibrosis independent of its aetiology. Nat Commun. (2013) 4:2823. doi: 10.1038/ncomms3823
33. Wree A, McGeough MD, Pena CA, Schlattjan M, Li H, Inzaugarat ME, et al. NLRP3 inflammasome activation is required for fibrosis development in NAFLD. J Mol Med. (2014) 92:1069–82. doi: 10.1007/s00109-014-1170-1
34. Campos PC, Gomes MTR, Marinho FAV, Guimaraes ES, de Moura Lodi Cruz MGF, Oliveira SC. Brucella abortus nitric oxide metabolite regulates inflammasome activation and IL-1beta secretion in murine macrophages. Eur J Immunol. (2019) 49:1023–37. doi: 10.1002/eji.201848016
35. Costa Franco MM, Marim F, Guimaraes ES, Assis NRG, Cerqueira DM, Alves-Silva J, et al. Brucella abortus triggers a cGAS-independent STING pathway to induce host protection that involves guanylate-binding proteins and inflammasome activation. J Immunol. (2018) 200:607–22. doi: 10.4049/jimmunol.1700725
36. Yaping Z, Ying W, Luqin D, Ning T, Xuemei A, Xixian Y. Mechanism of interleukin-1beta-induced proliferation in rat hepatic stellate cells from different levels of signal transduction. Apmis. (2014) 122:392–8. doi: 10.1111/apm.12155
37. Reiter FP, Wimmer R, Wottke L, Artmann R, Nagel JM, Carranza MO, et al. Role of interleukin-1 and its antagonism of hepatic stellate cell proliferation and liver fibrosis in the Abcb4(-/-) mouse model. World J Hepatol. (2016) 8:401–10. doi: 10.4254/wjh.v8.i8.401
38. Artlett CM. Inflammasomes in wound healing and fibrosis. J Pathol. (2013) 229:157–67. doi: 10.1002/path.4116
39. Tanino A, Okura T, Nagao T, Kukida M, Pei Z, Enomoto D, et al. Interleukin-18 deficiency protects against renal interstitial fibrosis in aldosterone/salt-treated mice. Clin Sci. (2016) 130:1727–39. doi: 10.1042/CS20160183
40. Leaf IA, Nakagawa S, Johnson BG, Cha JJ, Mittelsteadt K, Guckian KM, et al. Pericyte MyD88 and IRAK4 control inflammatory and fibrotic responses to tissue injury. J Clin Invest. (2017) 127:321–34. doi: 10.1172/JCI87532
41. Fan JM, Huang XR, Ng YY, Nikolic-Paterson DJ, Mu W, Atkins RC, et al. Interleukin-1 induces tubular epithelial-myofibroblast transdifferentiation through a transforming growth factor-beta1-dependent mechanism in vitro. Am J Kidney Dis. (2001) 37:820–31. doi: 10.1016/S0272-6386(01)80132-3
Keywords: Brucella, inflammasome, fibrosis, hepatic stellate cells, IL-1β
Citation: Arriola Benitez PC, Pesce Viglietti AI, Gomes MTR, Oliveira SC, Quarleri JF, Giambartolomei GH and Delpino MV (2020) Brucella abortus Infection Elicited Hepatic Stellate Cell-Mediated Fibrosis Through Inflammasome-Dependent IL-1β Production. Front. Immunol. 10:3036. doi: 10.3389/fimmu.2019.03036
Received: 13 October 2019; Accepted: 11 December 2019;
Published: 21 January 2020.
Edited by:
Juarez Antonio Simões Quaresma, Instituto Evandro Chagas, BrazilReviewed by:
Leticia Cedillo-Barron, Center for Research and Advanced Studies (CINVESTAV), MexicoRebecca Leigh Schmidt, Upper Iowa University, United States
Copyright © 2020 Arriola Benitez, Pesce Viglietti, Gomes, Oliveira, Quarleri, Giambartolomei and Delpino. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María Victoria Delpino, mdelpino@ffyb.uba.ar