Faunal Assemblages From Lower Bed I (Oldupai Gorge, Tanzania)
 Pamela Akuku1,2*
Pamela Akuku1,2*  Palmira Saladié1,2,3
Palmira Saladié1,2,3  Abdallah Mohamed4
Abdallah Mohamed4  Aloyce Mwambwiga5,6
Aloyce Mwambwiga5,6  Pastory Bushozi4
Pastory Bushozi4  Julio Mercader1,5,7,8
Julio Mercader1,5,7,8- 1Institut Catalá de Paleoecología Humana i Evolució Social (IPHES-CERCA), Tarragona, Spain
- 2Área de Prehistòria, Universitat Rovira i Virgili (URV), Tarragona, Spain
- 3Departamento de Paleobiologia, Museo Nacional de Ciencias Naturales, Madrid, Spain
- 4Department of Archaeology and Heritage Studies, University of Dar es Salaam, Dar es Salaam, Tanzania
- 5Department of Anthropology and Archaeology, University of Calgary, Calgary, AB, Canada
- 6National Natural History Museum, Arusha, Tanzania
- 7Max Planck Institute for the Science of Human History, Jena, Germany
- 8Department of Geoscience, University of Calgary, Calgary, AB, Canada
Palaeobiological and archeological excavations at the site of Ewass Oldupa, found in the western Plio-Pleistocene rift basin of Oldupai Gorge (also Olduvai Gorge), Tanzania, revealed rich fossiliferous levels and the earliest remains of human activity at Oldupai Gorge, dated to 2 million years ago. This paper provides zooarchaeological taxonomic, taphonomic, and behavioral analyses, applying several methods to explore the setting in which the assemblage was formed. We identified agency behind bone surface modifications, such as cut, tooth and percussion marks, and determined the frequency of carnivore tooth marks as well as their distribution on both discrete specimens and across species. In addition, our work revealed co-occurrence of modifications to include butchering marks and carnivore tooth marks. Ravaging levels were estimated as percentage. The faunal accumulation from Ewass Oldupa contains two cut marked specimens, together with low degrees of percussion and carnivore tooth marks, moderate ravaging, and diagenetic changes suggestive of water flow. Thus, multiple lines of evidence indicate a palimpsest accumulation. Taxonomic diversity is high, with up to 22 taxa representing diverse habitats, ranging from open grassland to wooded bushlands, as well as moist mosaics during Bed I. Overall, this archaeo-faunal assemblage speaks to increased behavioral versatility among Oldowan hominins and interactions with the carnivore guild.
Introduction
Oldowan archeological sites record hominin adaptive behavior through faunal and cultural remains, such as stone tools. The relationship between technological and feeding behavior is at the center of subsistence strategies perceived to be uniquely human, especially meat consumption during the early Pleistocene, more than 2.5 million years ago. However, establishing a solid link between hominin processing of animal tissue and technological manipulation for dietary purposes requires heavy proof to overcome the equifinality and ambiguity that have protracted the debate surrounding habitual meat-eating among early Homo species for decades (Bunn, 1986a; Blumenschine and Cavallo, 1992; Domínguez-Rodrigo, 2002; Pante, 2010, 2013; Pobiner, 2015; Organista et al., 2016; Domínguez-Rodrigo and Cobo-Sánchez, 2017; Dominguez-Rodrigo and Pickering, 2017; Pante et al., 2018).
The importance of this debate goes beyond dietary preference alone to include the emergence of novel landscape adaptations like long distance provisioning, new ways of structuring habitation sites around focal spots, archeological site formation, and avoidance strategies in the carnivore guild (Egeland, 2012; Pickering et al., 2013; Aramendi et al., 2017). Taphonomic studies carried out in Oldupai Gorge indicate that only a small number of anthropogenic sites dated to the Pleistocene exhibit a functional association between stone tools and faunal remains (Bunn, 1986a,b; Pickering et al., 2004; Pobiner et al., 2008; Yravedra and Domínguez-Rodrigo, 2009). There is a consensus of an existing disparity in the Pleistocene sites at Oldupai concerning hominin-faunal association despite the Bed I Oldowan material (Leakey et al., 1951; Toth, 1993; Kimura, 2002; Proffitt, 2018) and Bed II Acheulean (Leakey, 1971; McHenry et al., 2016). Taphonomy based studies record anthropogenic signals such as cut marks and percussion marks as well as zoogenic marks in the form of carnivore tooth marks from Bed I, e.g., FLK (Bunn et al., 1986; Blumenschine, 1995; Domínguez-Rodrigo and Barba, 2006) and II sites, e.g., Bells Korongo (Egeland and Domínguez-Rodrigo, 2008; Yravedra et al., 2017b). Studies conducted to understand the formation (accumulation and modification of bones) processes and relationship between hominins and carnivores in Pleistocene contexts indicate mixed resource access scenarios involving hominins and other large predators (Capaldo, 1997, 1998; Selvaggio, 1998; Domínguez-Rodrigo, 1999; Egeland et al., 2004; Baquedano et al., 2012; Arriaza and Dominguez-Rodrigo, 2016).
The abundant fauna from Bed I (Faith et al., 2009) supports taphonomic and palaeoecological studies used in reconstruction of the sites, e.g., FLK-NN and ‘Amin Mturi Korongo’ AMK (Domínguez-Rodrigo et al., 2014; Aramendi et al., 2017) and inferences of hominin habitat use (Plummer and Bishop, 1994; Domínguez-Rodrigo et al., 2007, 2019). Taxon-based reconstructions indicate the presence of mixed environments; open grasslands, wetlands, and wooded closed habitats in Bed I (Ashley et al., 2010; Aramendi et al., 2017) with hominins preferring riparian woodlands as residential camps (Domínguez-Rodrigo et al., 2007).
Our goal is to present the results of the taphonomic studies carried out on the recently excavated site of Ewass Oldupa; which currently preserves the earliest evidence of human occupation in the basin, boasting Oldupai’s oldest lithic remains in the Western Gorge, within the Ngorongoro Formation and Lower Bed I (Mercader et al., 2021). The lithics, which are mostly quartzite, predate the tool bearing deposits from ‘Douglas Korongo’ (DK) located in the Eastern basin, which represented the earliest Oldowan in the region at ∼1.848 ± 0.003 ma. Ewass Oldupa is located 350 m northeast of Geolocality 64 bracketed between the Naabi ignimbrite 2.0 ma and the Ng’eju tuff 1.818 ± 0.006 ma (Hay, 1963). It was primarily excavated in 2018 by the OGSDS (Olduvai Gorge Stone Tools, Diet and Sociality) project with a follow up season in 2019 leading to the excavation of 5 trenches: 2, 3, 5, 6, and 7 (Figure 1). Our initial studies (Mercader et al., 2021) implied Ewass Oldupa was characterized by open habitats with mosaics of bushland/woodland based on the presence of open and closed habitat species as well water dependent individuals. The archaeological record of Ewass Oldupa indicates the Oldowan hominins (2–1.9 ma) ability to exploit mixed environments and diverse biomes. In this study, we further examine hominin behavior in association with faunal datasets with the aim of understanding hominin and fauna relations, subsistence strategies, site formation processes, and paleo environmental reconstruction.
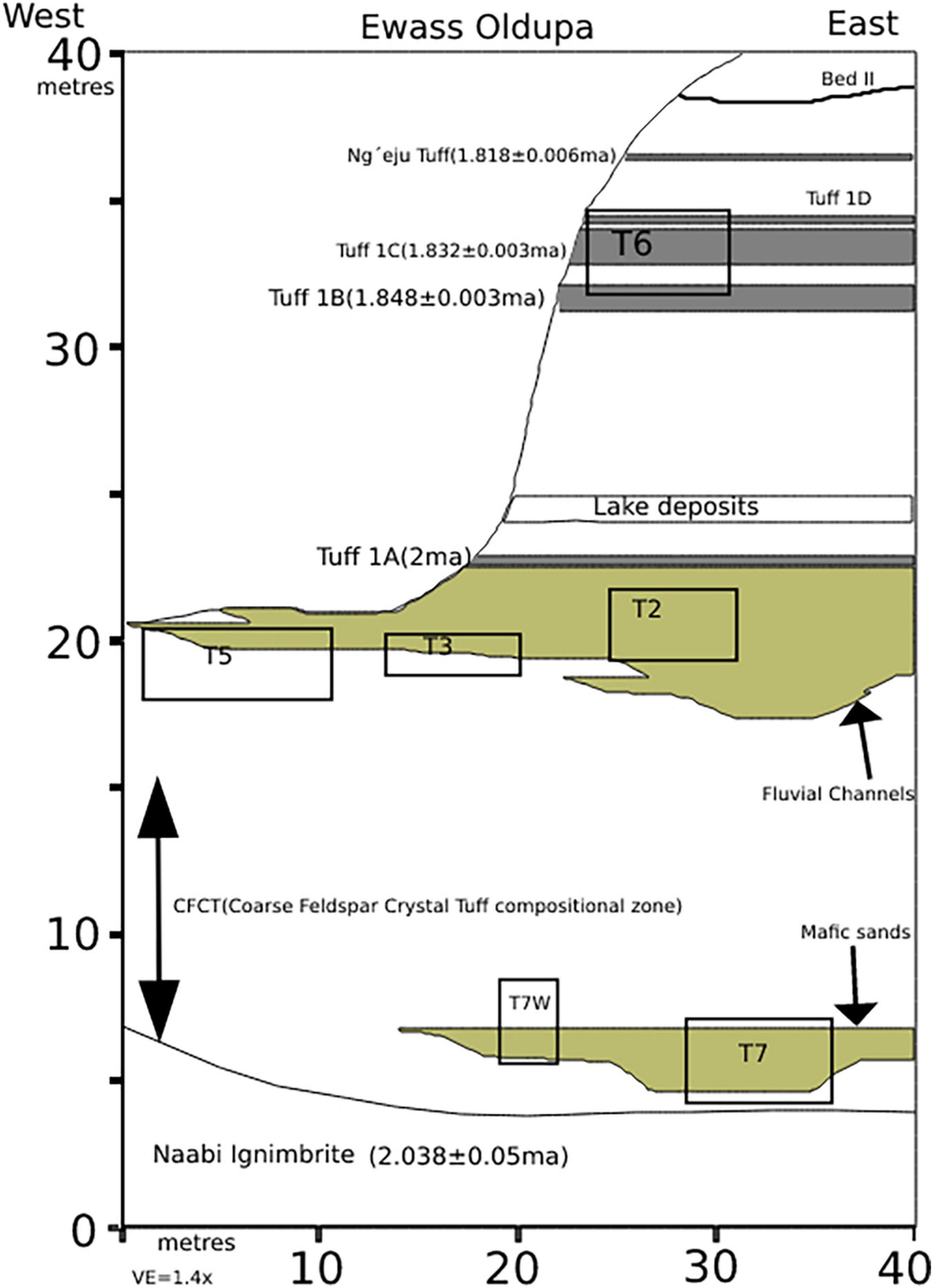
Figure 1. Tuff dates and locations, sampled stratigraphic sections and excavated trenches at Ewass Oldupa. Figure indicating the date and location of tuffs, stratigraphic sections sampled and excavation trenches from Ewass Oldupa (modified from Mercader et al., 2021).
Materials and Methods
The materials for this study came from Ewass Oldupa which occurs in the Oldupai group that has a thickness of 100 m and is divided into seven units; Bed I–IV, Masek, Ndutu, and Naisiusiu beds (Hay, 1976; Scoon, 2018). These units are recognized for their preservation of remains and evidence of hominin activities (Day and Molleson, 1976; Hay, 1976; Domínguez-Rodrigo, 1997; Fernández-Jalvo et al., 1998; McHenry et al., 2007; Blumenschine et al., 2012; de la Torre et al., 2012; Pante, 2013; Díez-Martín et al., 2015; Hlusko et al., 2015; Organista et al., 2016; Yravedra et al., 2017a). A densely deposited sequence with ages obtained from existing 40Ar/39Ar dates of tuffs was exposed at six localities Geolocalities 61, 62, 63, 64, 64-3, and 66 along a 2 km section (McHenry, 2012; Habermann et al., 2016; Favreau et al., 2020). At Ewass Oldupa it was established that the lowermost stratigraphic unit is the Naabi ignimbrite of the Ngorongoro Formation above this is the Coarse Feldspar Crystal Tuff (CFCT) dating to about 2.015 ± 0.006 ma (Favreau et al., 2020; Soto et al., 2020). Lower Bed I which overlays the CFCT contains Tuff 1A dating to 2.0 ma and is characterized by carbonate beds as well as green waxy clays. Tuff IB (1.848 ± 0.003 ma) occurs in Upper Bed I and is overlain directly by waxy claystones above these clays, is a layered siltstone with the geochemical fingerprint of Tuff IC (1.832 ± 0.003/1.848 ± 0.008 ma). On top of it, is a layered sandstone congruent with Tuff ID, capped by a thin bed with carbonate nodules identified as the Ng’eju Tuff dating to 1.818 ± 0.006 ma (Favreau et al., 2020; Soto et al., 2020; Mercader et al., 2021). A total of five trenches were excavated; trench2 measuring 21 m2, trench3 31 m2, trench5 40 m2, trench6 42 m2, and trench7 82 m2(Figure 1). The faunal assemblage studied came from three trenches; 2, 3, and 5 as there was no fauna recorded in trenches 6 and 7.
Excavations at Ewass Oldupa recovered 3,661 fossil specimens and 565 stone artifacts of which flakes are the dominant product (Mercader et al., 2021) with trench 2 recording the highest density of faunal materials producing 2,065 specimens from a 2 m thick sequence. During excavation, the location of material larger than 2 cm was recorded on a Leica Total Station 09, placed within an X–Y Cartesian system, with Z-values also recorded and sediments from each trench dry sieved using a 0.5 cm mesh.
Quantification and Analysis
All specimens of macro fauna [defined here as any specimen with a live weight exceeding 4.5 kg in accordance with Brain’s (Brain, 1974) size classes] greater than 2 cm including both identifiable and unidentifiable specimens were analyzed. Elements less than 2 cm were weighed (500 g) and no further analyses conducted on them. We recorded the element, taxon (when possible), lateralization (left or right), portion, face, and age. Identifiable and non-identifiable specimens were classified into small, medium and large size classes according to the live weight size classes proposed by Brain (1974), Klein (1976), and Milo (1998) and described as long, flat, or irregular in relation to their morphology. Relative frequencies such as Number of Identified Specimens (NISP), Minimum Number of Elements (MNE) and Minimum Number of Individuals (MNI) were used to determine and compare relative abundance of skeletal elements and species (Binford, 1984; Grayson, 1984).
We created skeletal profiles for Ewass Oldupa using the normed NISP, MNE, and %MAU. Normed NISP (Nnisp) was calculated using the NISP numbers from the specimens with all elements included representing the skull, forelimbs, hindlimbs and distal parts (Faith, 2007; Reynard et al., 2016b). The MAU and %MAU was calculated using the MNE long bone portions represented in the assemblage (Binford, 1984). Correlations were used to establish the relationship between Minimal Animal Units (%MAU) and bone mineral density in addition to their food utility by comparing values to %MGUI (Binford, 1978) and Lam’s (Lam et al., 1999) Rangifer tarandus values. %MAU profiles were created for Ewass Oldupa using long bone portions for the small and medium size classes. There was disparity in the representation of long bone portions for the large size classes and as such, they were not included in the profile.
The age of the animals was described by the replacement and wear pattern of teeth following Bunn and Pickering (2010) with specimens classified as sub-adult juveniles (young and sub-adult juveniles), adults (early and late prime adults), old adults and unknown where age could not be determined. We grouped the age classes into the three main classes to perform ternary plots according to Steele and Weaver (2002) comparing them with Olduvai datasets from Bunn and Pickering (2010).
The complete surface of all the bones and teeth were inspected with a stereomicroscope (OPTHEC 120 Hz model) with between 10 and 45× magnification. The Ewass Oldupa assemblage recorded various anthropogenic, zoogenic and diagenetic modifications. We assessed and categorized them into nutritional, subaerial and post depositional phases (Capaldo, 1998).
Cut marks (Potts and Shipman, 1981) in the assemblage are scarce and in the form of slicing marks, which occur when the tool is used to apply force parallel to the long axis at the edge of the tool (Lyman, 2008). A hirox kh 7800 microscope was used to assess the cut marked specimens, take the cut mark measurements, and create the cut mark profiles. Their location on different bones in the skeleton is important and can help in the inference and understanding the mode of access and carcass utility (Galán et al., 2009; Yravedra and Domínguez-Rodrigo, 2009; Reynard et al., 2016a).
Tooth marks identified were scores, punctures, pits and furrowing on the cancellous tissue (Binford, 1981). Their location on the bone was noted and their measurements taken using digital calipers following the proposals of Andrés et al. (2012). Tooth mark measurements: mean lengths and widths were expressed in box plots with a 95% confidence interval and compared with living and known carnivore measurements (Selvaggio, 1994; Delaney-Rivera et al., 2009; Andrés et al., 2012). To assess the possible role of carnivores and their ravaging effect on assemblages for instance the removal of epiphyses we calculated the percentage of change (Marean and Spencer, 1991; Blumenschine, 1993) using the formula: MNE before ravaging-MNE after ravaging)/(MNE before ravaging) *100. The percentage change indicates the change in MNE portions before and after ravaging. High percentage change values suggest a high degree of attrition with low accuracy in estimating the original abundance of the elements (Marean and Spencer, 1991; Blumenschine, 1993; Domínguez-Rodrigo et al., 2002). These ratios are also useful to evaluate the integrity of the sample. Taking into the account the scarce anthropic modifications, the levels of ravaging present in the assemblage were assessed using the rations proposed by Domínguez-Rodrigo and Organista (2007). These ratios proposed were (ribs + vertebrae) to (limb bones), (proximal humeri + distal radii) to (distal humeri + proximal radii). Lower values obtained from these ratios would indicate intensive ravaging whereas a ratio value of one indicates lower levels or absence of ravaging.
We recorded the presence and incidence of post-depositional modifications such as weathering (Behrensmeyer, 1978), trampling (Fiorillo, 1988; Gaudzinski-Windheuser et al., 2010; Reynard, 2014), abrasion (Brain, 1967; Shipman and Rose, 1983; Olsen and Shipman, 1988) and rounding (Fernandez-Jalvo and Andrews, 2016) as these modifications allow us to assess the accumulation and modification processes of the fauna sets. Statistical tests such as r Pearson correlation and Simpson evenness tests were applied to determine relationships between the different taphonomic and taxonomic aspects analyzed. We used Spatstats package v. 3.2.3 (Renner et al., 2015) in R for the statistical spatial analysis. A non-parametric method of Kernel maps was also used to produce graphic smoothing estimations of intensity. Density maps were made by using bandwidths selected by sigma values, which control the degree of smoothing. Corrections for window-edge effects were considered. To select the optimal bandwidth, Diggle and Berman’s mean square error cross-validation method and the likelihood cross-validation method were used (Berman and Diggle, 1989). A test to evaluate of point inter-dependence between lithics and bones in the T2, T3, and T5 was a random labeling test [using the Kij(r) function]. A test to evaluate of point inter-dependence between lithics and bones in the T2, T3, and T5 was a random labeling test [using the Kij(r) function]. We used (α = 0.01) as the level of significance due to our relatively small datasets through Mont Carlo sampling of (n = 199) (Renner et al., 2015).
Breakage patterns were classified based on Villa and Mahieu (1991) considering the delineation, angle and surface of the fracture edges. We recorded fracture patterns for both ends of any given long bone fragment in relation to delineations (curve/V Shape, longitudinal, or transverse), angle (straight, oblique, or mixed) and surface (smooth or irregular). The breakage patterns provide information on the state of the bone (fresh vs. dry) at the time of fragmentation. Fresh fractures aid in understanding either hominin or carnivore access to carcasses in any given assemblage.
Results
A total of 30.3% of the specimens were taxonomically identified (NISP = 1,157) comprising 22 taxa with the most represented taxonomic group being bovids of different weight sizes. The presence of bovid tribes Tragelaphini, Hippotragini, and Antilopini was observed (Table 1) as well as families such as Equidae and Carnivora indicating the taxonomic diversity of the assemblage. Fragmented pieces of plastron (n = 47) and carapace (n = 41) belonging to Chelonia sp. as well as ostrich eggshells (n = 27) were also abundant in the assemblage (Table 2). Simpson diversity Index for trenches 2, 3, and 5 (0.7, 0.7, and 0.6, respectively) suggest high levels of taxonomic diversity across all three trenches with evenness values of 0.3. According to the bovid size classes, the more abundant identified specimens belong to the medium size weight class (which corresponds to Brain’s size classes 2 and 3) as well as most of the non-id specimens (Figure 2).
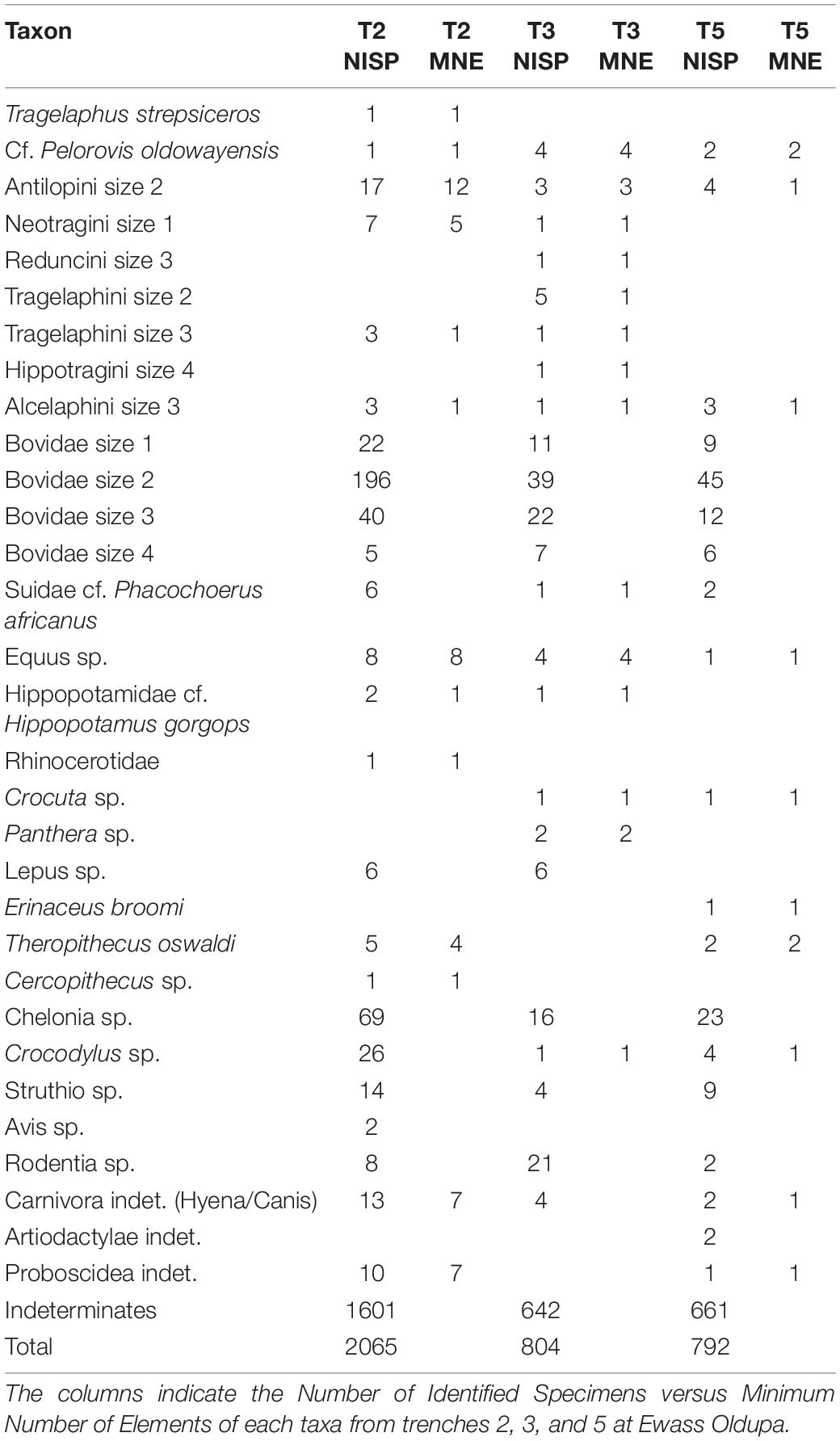
Table 1. Represented taxa in the trenches 2, 3, and 5 at Ewass Oldupa.
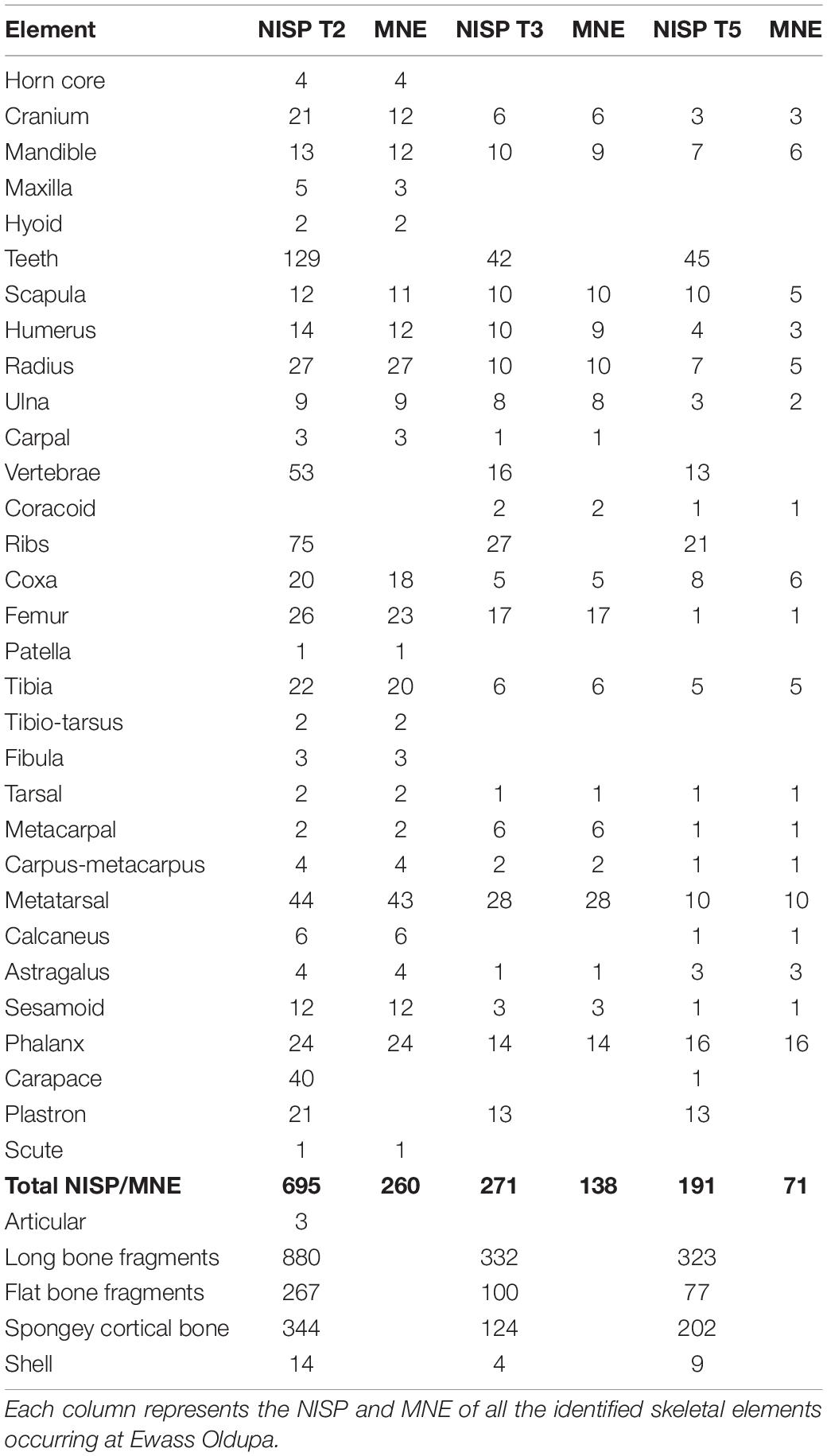
Table 2. Representation of elements and quantification through NISP and MNE.

Figure 2. Mortality pattern of the bovids from Ewass trenches 2, 3, and 5. Actualistic data from Stiner (1990) and modified by Bunn and Pickering (2010). (A) Triangular mortality diagram showing the different mortality patterns described by Stiner (1990). Panels (B,C) modified triangular graphs comparing with actualistic data of several predators.
The results of the MNI indicate a total of 291 individuals are represented in our assemblage with Bovidae accounting for (53.6%, n = 156), Suidae (3.4%, n = 10), Equidae (3%, n = 9), Carnivora (2.7%, n = 8), Lagomorph (2.4%, n = 7), Primates (2.4%, n = 7), Aves (2.1%, n = 6), Proboscidea (1%, n = 3) the remaining 85 individuals represent crocodiles and indeterminate species. Age profiles from the bovid age of death indicate a dominance of prime adult individuals in the assemblage resulting from a catastrophic mortality (Figure 3).
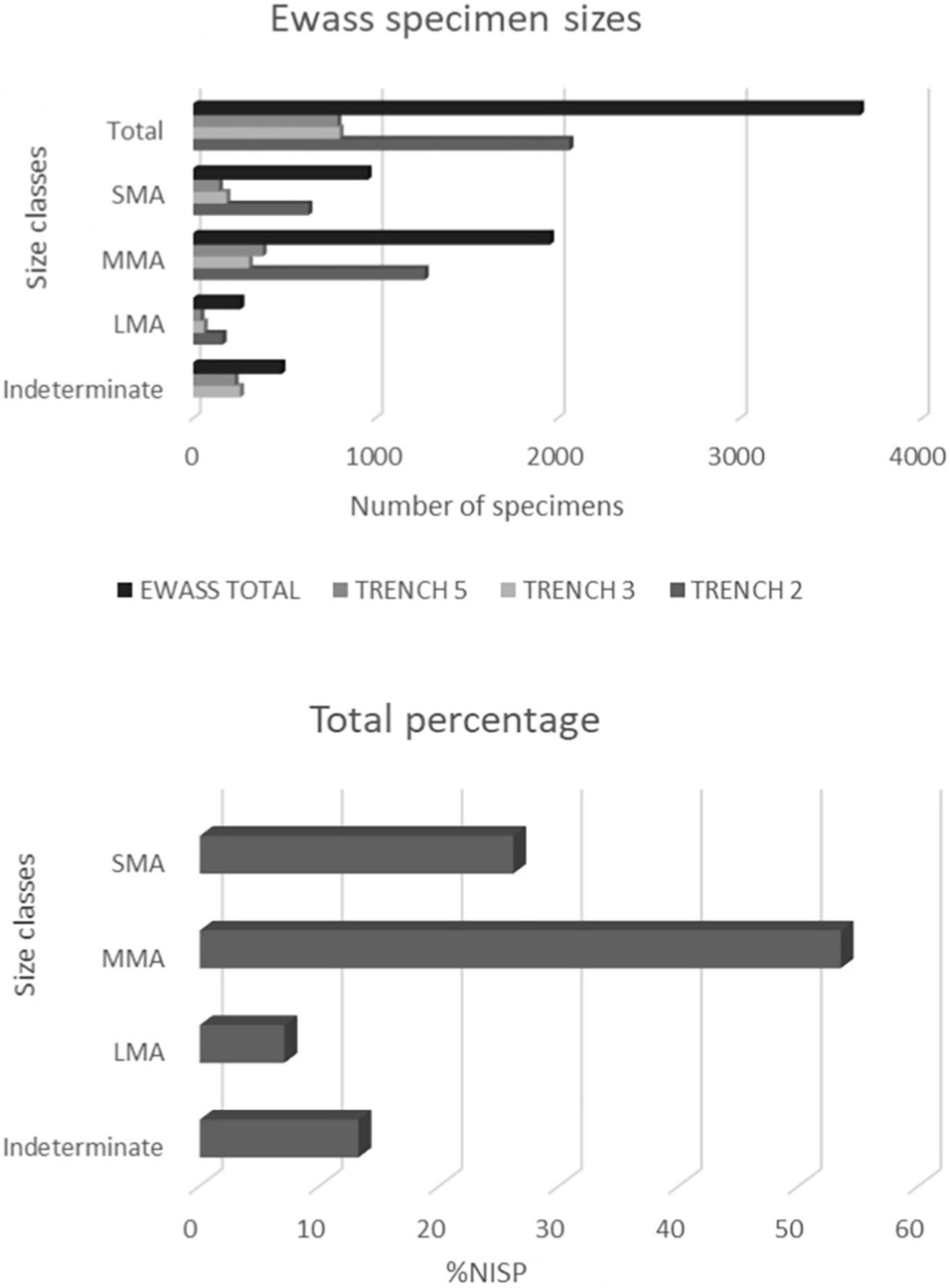
Figure 3. Frequency bar chart representation of Ewass size classes across the trenches, %NISP of size classes across the trenches.
The most dominant elements based on the MNE across the trenches were metatarsals, phalanges and radii (Table 3). The medium size class recorded high preservation of radius and femur.
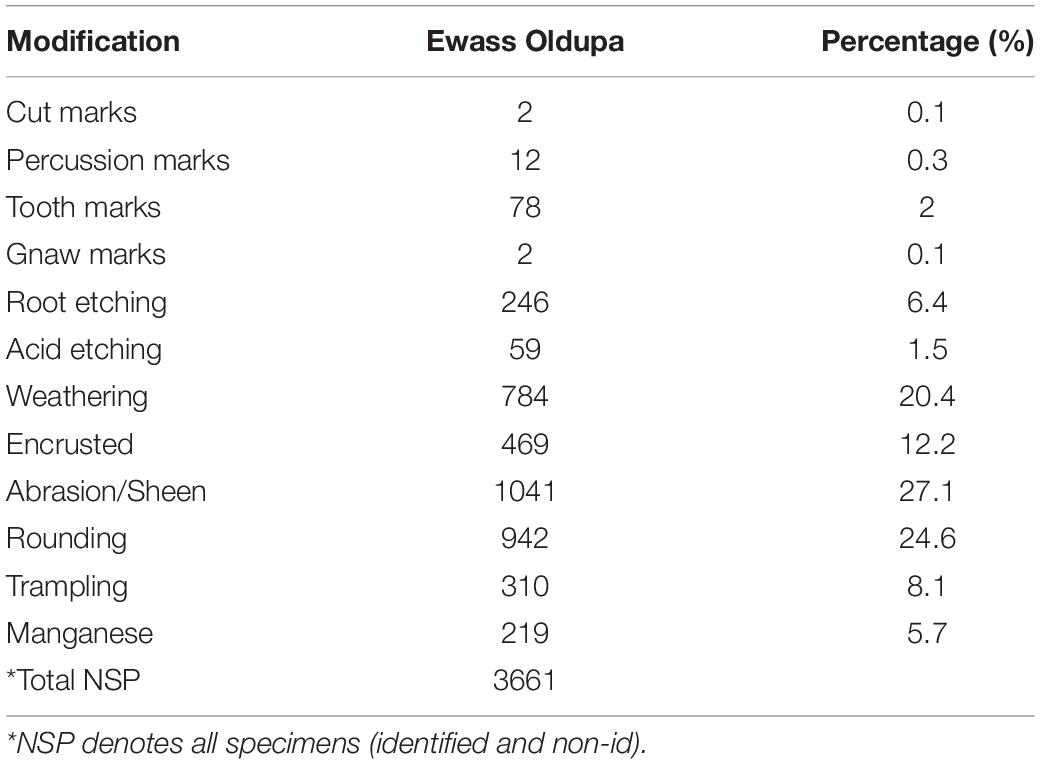
Table 3. Shows the number and percentage of the various anthropogenic, zoogenic, and diagenic modifications recorded in the Ewass assemblage.
Normed NISP profiles for Ewass Oldupa indicate equitable distribution of skeletal portions in the small and medium classes whereas the large sizes show dominant preservation of skull elements (Figures 3, 4). The results using the %MAU and Lam’s values indicate no significant statistical correlation [rs = 0.13511, p (2-tailed) = 0.64515]. Pearson correlation tests carried out on the %MAU and %MGUI show no significant statistical correlation (r = 0.4836, P-Value = 0.111189).
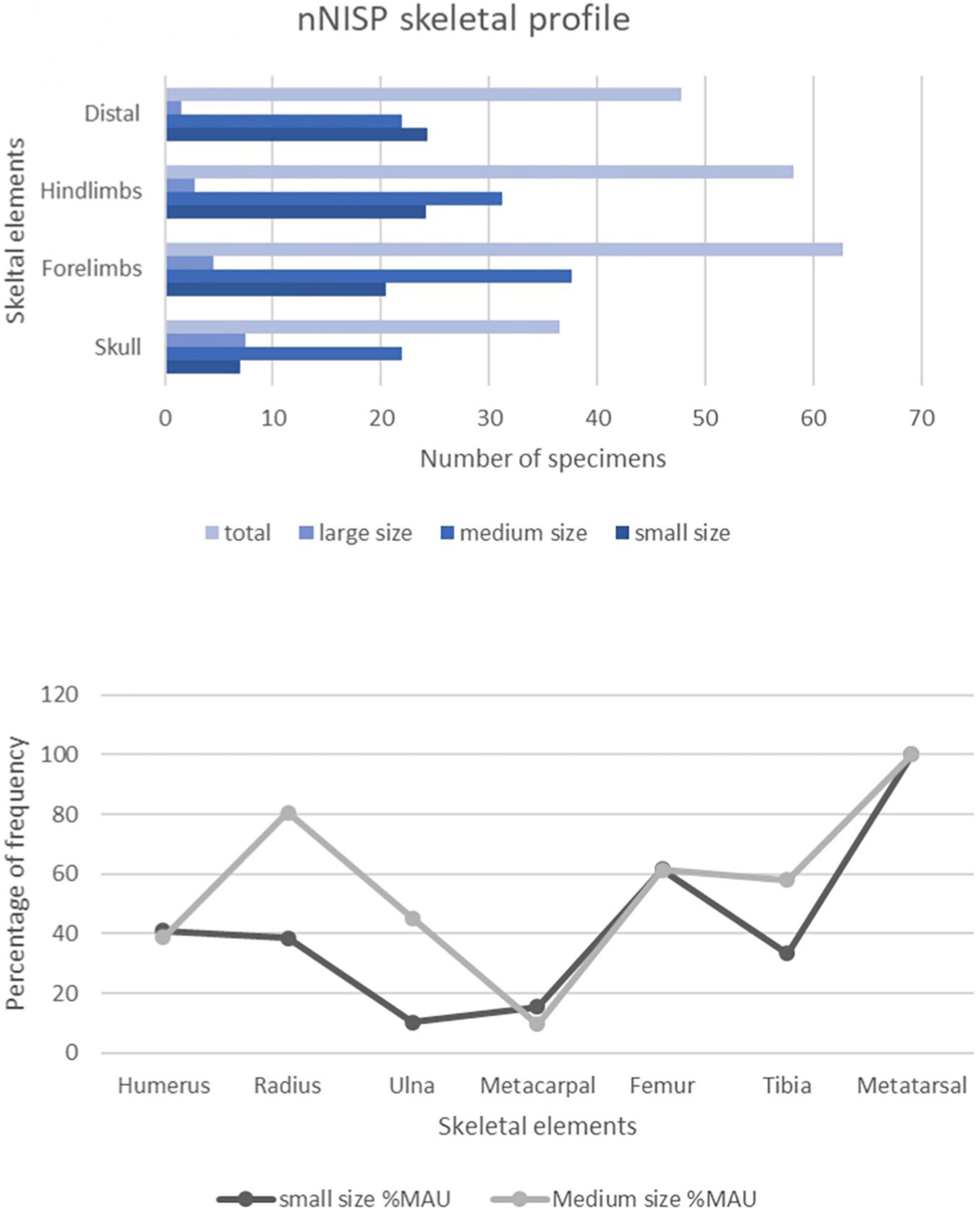
Figure 4. Skeletal profiles of Ewass Specimens created from normed NISP (nNISP). Skeletal profiles of Ewass Oldupa specimens created from the normed Number of Identified Specimens compared with %MAU profiles using long bone portions of small and medium size classes. Percentage Minimum Animal Units (%MAU) profile created using the long bone portions of small and medium size classes calculated from the MNE based on Binford (1984).
Smoothing Kernel density maps of fauna and lithic remains show no overlapping in T2. However, the maps of T3 and T5 visually indicate that the materials are found in nearby areas. In T3 there is a zone of greater accumulation, although fauna and lithic industry are not completely overlapping. At T5 the overlap appears larger according to visual examination of the Kernel density maps. The results of the Kij(r) function indicate that the accumulation processes of fauna and lithic tools are aleatory and independent in T2, there is a tendency to segregation between the material types in T3, and in T5, although there could be some spatial dependence, there is little statistical significance.
Bone Breakage
The nature of the assemblage sampled is somewhat fragmentary with 91% of long bone fragments retaining less than 1/4 in length and approximations could not be made of 12.6% of the sample. 90% of the assemblage can be classified as type 1 based on (Bunn, 1986a) as they retain <50% of the shaft circumference. Among the rest of the specimens, 27.6% are diaphysis of long bones with a scarcity of epiphyses. Long bone fragments and fracture patterns indicate majority of the bones were broken when dry. In the set of long bones, 3% of the fracture edges were transverse with right angles, 13% recorded irregular fractures with mixed angles and 86% had smooth fracture/eroded fractures with oblique angles indicating a high presence of diagenetic fractures, when the bones were already dry. However, a small set 2.5% with green spiral fractures was located, where two out of the 12 elements exhibiting percussion marks preserved conchoidal fracture notches, hinting at minimal anthropic action. The assemblage also records a set of phalanges that exhibit longitudinal and transverse fractures. The fractures are very smooth and precise perhaps denoting green fractures smoothed by post depositional processes over time.
Bone Surface Modifications
Nutritional Phase
The modifications of the nutritional phase of the carcasses are scarce. Modifications were documented in the form of cut marks, based on their section, delineation and internal micro striations (Figure 5). Two specimens belonging to medium size class animals (a rib shaft fragment and long bone shaft fragment) exhibit the slice marks. In the first specimen the cuts occur on the ventral side of the rib fragment and on the cortical surface of the long bone fragment in the second case. The average length of the cut marks was 2 mm and the width was 0.2 mm. The long bone shaft records two scores as well adjacent to the cut marks.

Figure 5. Two cut marks from two specimens recorded in the Ewass Oldupa assemblage. (Top) Cut marks (with white arrows) and scores (yellow double arrow) on long bone fragment. (Bottom) Cut mark (white arrow) on rib shaft fragment. Measurements in mm on X- and Y-axis.
The anthropogenic bone breakage is identified through 12 (0.3%) percussion marks occurring on mostly on long bones and a single indeterminate flat bone. Majority of these modifications are located on the non-id long bone fragments (n = 8) with long bone mid-shafts belonging to the medium size class recording 3 percussion marks as well as an indeterminate flat bone fragment. One of these mid-shaft specimens preserves a conchoidal fracture. Peeling is not recorded in the assemblage, however, 4 of the non-id long bone fragments preserve both percussion and tooth marks.
Seventy-eight specimens (2%) in the sampled assemblage exhibit tooth marks. Carnivore modifications in the assemblage occur in the form of pits (n = 31), scores (n = 23), punctures (n = 16), and furrowing (n = 8) as well as acid etching in 1.5% (n = 59) of the specimens. The tooth marks occur in variety of skeletal elements representing different species such as Bovidae, Theropithecus, Testudinae and Proboscidea. These modifications are more common on long bones (n = 43) than on flat bones (n = 21) with 50% occurring on the diaphysis and 5.1% on cancellous areas near the epiphysis portions while in the flat bones 19.2% are recorded on the diaphysis with 7.7% near the cancellous edges. The diversity in the specimens exhibiting tooth marks may indicate the action of more than one carnivore on the assemblage.
One of the specimens (SF54) recorded 15 pits, its measurements were compared to recorded measurements from actualistic samples dimensions (Figure 6). Results indicate that the measurements of tooth pits from the Ewass sample were the result of medium to large sized carnivores.
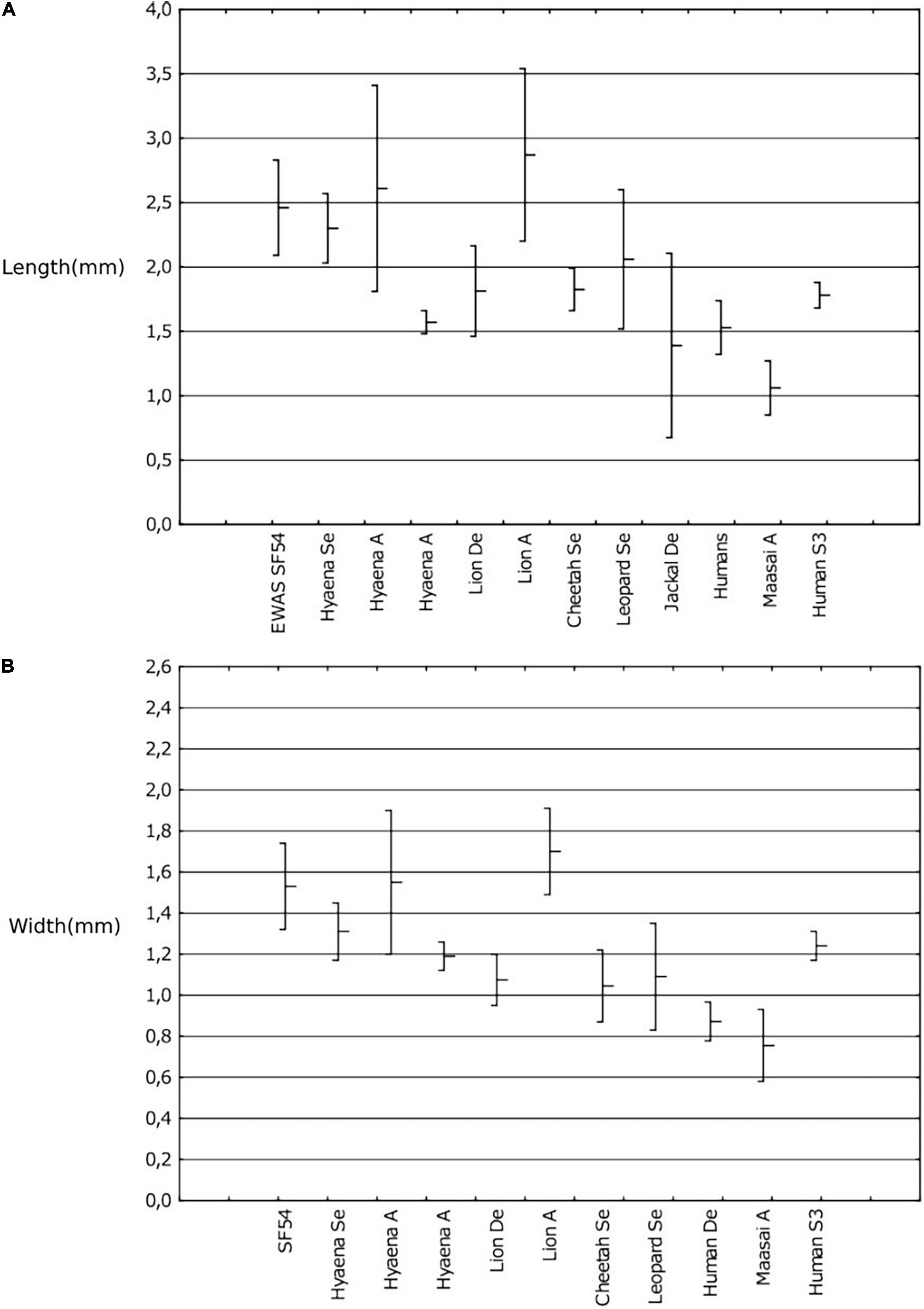
Figure 6. Measurements [mean and 95% confidence intervals in mm; length: (A); width: (B)] compared with known carnivore measurements. Indicates Measurements of specimen SF54 compared with living and known carnivore measurements (length: (A), width: (B), mean and 95% confidence intervals in mm) Pit lengths- Selvaggio (1994), Delaney-Rivera et al. (2009), Andrés et al. (2012), Ewass (2019) and Humans S3-Saladié et al. (2013).
Ravaging levels ratios indicate more intense ravaging in the ribs, vertebrae and limb bones (Figure 7) whereas elements such as humeri and radii, femur and tibia provided a ratio of one suggesting lower levels of ravaging. The Ewass assemblage records stages 1–2 ravaging levels based on Marean and Spencer (1991) as recorded in sites such as FLKNN 3 and FLK Zinj. The assemblage recorded relatively high percentage change values (Table 4) with r Pearson correlation testing [r = 0.8911, p < 0.05(0.042428)] indicating probable intense modification of the Ewass assemblage. This is evidenced by the long bone specimen that exhibits both cut marks and carnivore tooth marks in the form of scores (Figure 5).
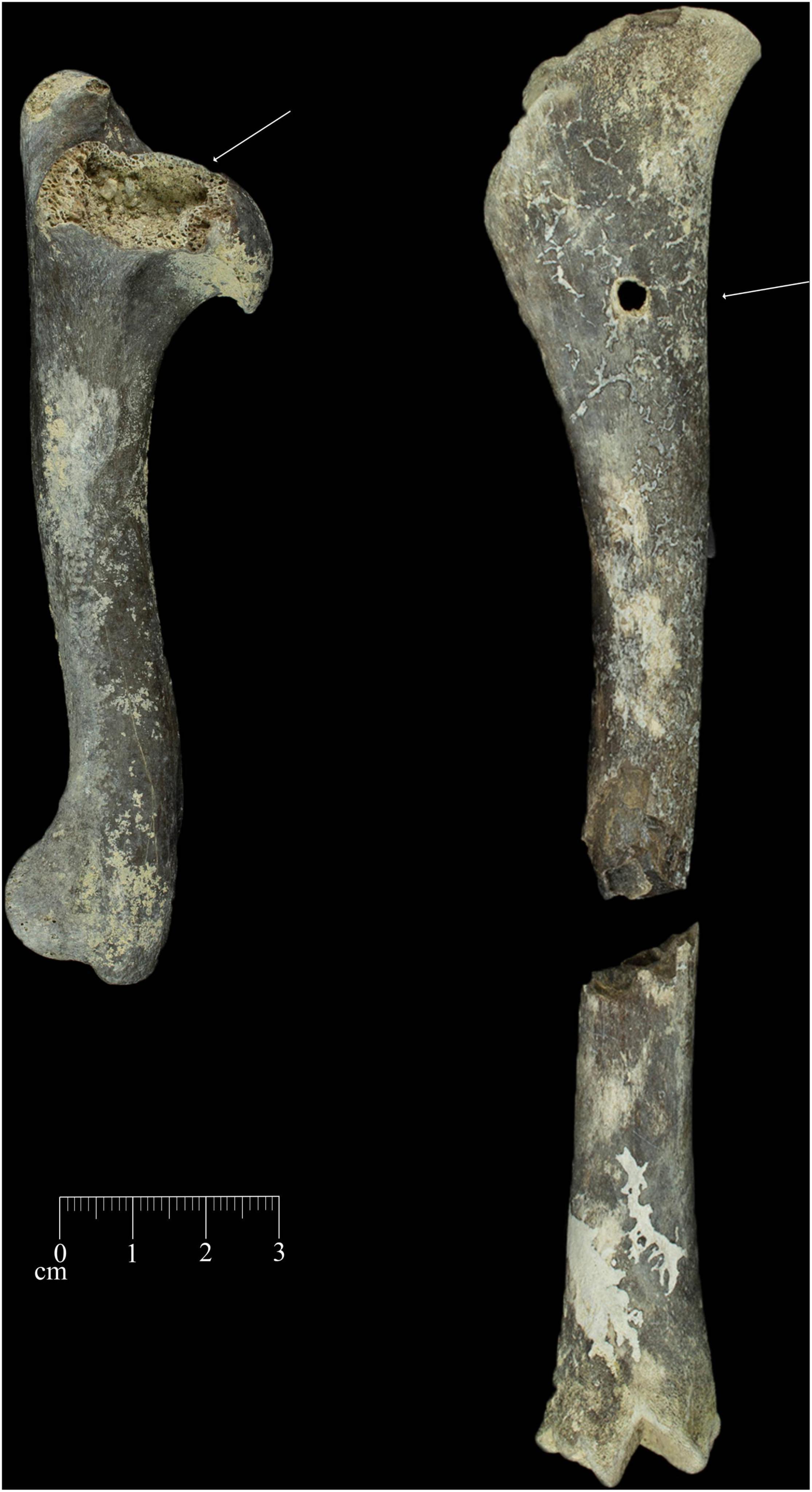
Figure 7. Example of carnivore modifications; tooth marks present at Ewass Oldupa. Carnivore modifications present on specimens. (Right) Arrow denoting furrowing present on humeral proximal end. (Left) Arrow indicating puncture mark present on proximal shaft of tibia.
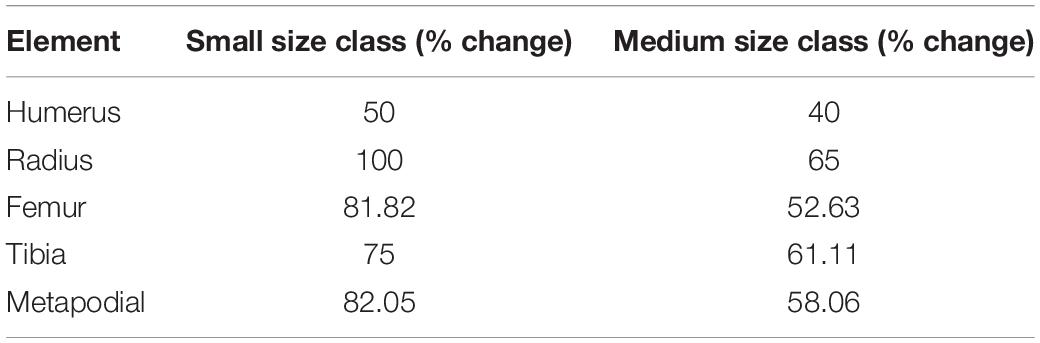
Table 4. Percentage change values of long bone specimens from the small and medium size classes.
Subaerial Phase
The modifications produced during the subaerial phase in the formation of Ewass Oldupa sample are the most abundant. These are rounding (24.6%), weathering (20.4%) encrusted bones (12.2%) and trampling (8.1%). 12.3% of the bones of this sample exhibit rounding of the entire surface while 6.1% had their original morphology completely modified. This indicates that at least part of the fauna set may have been incorporated in the form of bioclasts. Weathering was observed in many degrees (Behrensmeyer, 1978) degree 1 was present in 14.1%, degree 2 in 0.9% and degree 3 in 0.1% of the specimens. Part of the diagenetic bone breakage in Pleistocene sites may be related to this taphonomic process, however, the Ewass assemblage records minimal degrees of weathering.
Several specimens from Ewass Oldupa were affected by other diagenic modifications such as root etching 6.4% with 2% of the root-etched specimens recording biochemical corrosion through moss/lichen and fungi action. Staining on the specimens through manganese action was present in 5.7% of the specimens.
Discussion
The taphonomic signals are essential to know how archeological and paleontological assemblages formed, and the influence of the different taphonomic agents in these processes and in the modification processes themselves. It is also important to understand the relationship between the different objects in the assemblages to gauge formational and functional site types during formation. For instance, Leakey (1971) and Isaac (1978) established site typologies inclusive of stone caches, living floors, home bases, and butchering/kill sites near water sources. The results of our taphonomic analysis at the oldest occupation site from Oldupai Gorge show that, in spite of great taxonomic diversity (Figure 8), conflicting taphonomic scenarios arise from the data. For instance, there is no doubt that hominins accumulated stone tools at the excavated locality, but anthropogenic bone surface modifications are extremely rare. Thus, two specimens alone exhibit cut marks, and therefore no functional association can be made between lithics and animal bones. The site’s bone surfaces exhibit poor preservation in 38.5% of the assemblage in the form of encrustation, rounding, and exfoliation. These processes denote subaerial exposure and water flow, affecting the preservation of evidence for anthropic activity at the site (Pineda and Saladié, 2019).

Figure 8. Specimens recovered expressing diversity in the Ewass Oldupa assemblage. (A) Crocodylus sp. Teeth and Chelonia sp. plastron fragment with puncture mark. (B) Occlusal view of tooth belonging to Proboscidae indet.p and Theropithecus oswaldi ulna, radius and premolar.
The ravaging ratio indices are based on the presence/absence of low survival elements like epiphysis portions or postcranial axial elements (Marean and Cleghorn, 2003). However, their absence is common in archeological assemblages (Lyman and Lyman, 1994) and can be attributed to carnivore ravaging, hominin activity, mechanical, and chemical post depositional processes (e.g., fluvial transport, bone weathering, trampling, microbial attack, or chemical attrition) or by a mixture of all these processes (Saladié et al., 2014). The low frequency of anthropogenic and zoogenic modifications suggests that other post nutritional processes influenced the final anatomical composition of the assemblage.
Statistical spatial analyses suggest the assemblage was the product of independent accumulative processes (Figure 9). However, the water flows tend to disperse the elements in relation to their density (among other characters such as the size, shape, and weight) (Behrensmeyer, 1975; Petraglia and Potts, 1994; Coard, 1999; Domínguez-Rodrigo et al., 2019), a case is evident in T3, a set in which there is a large volume of rounded remains.
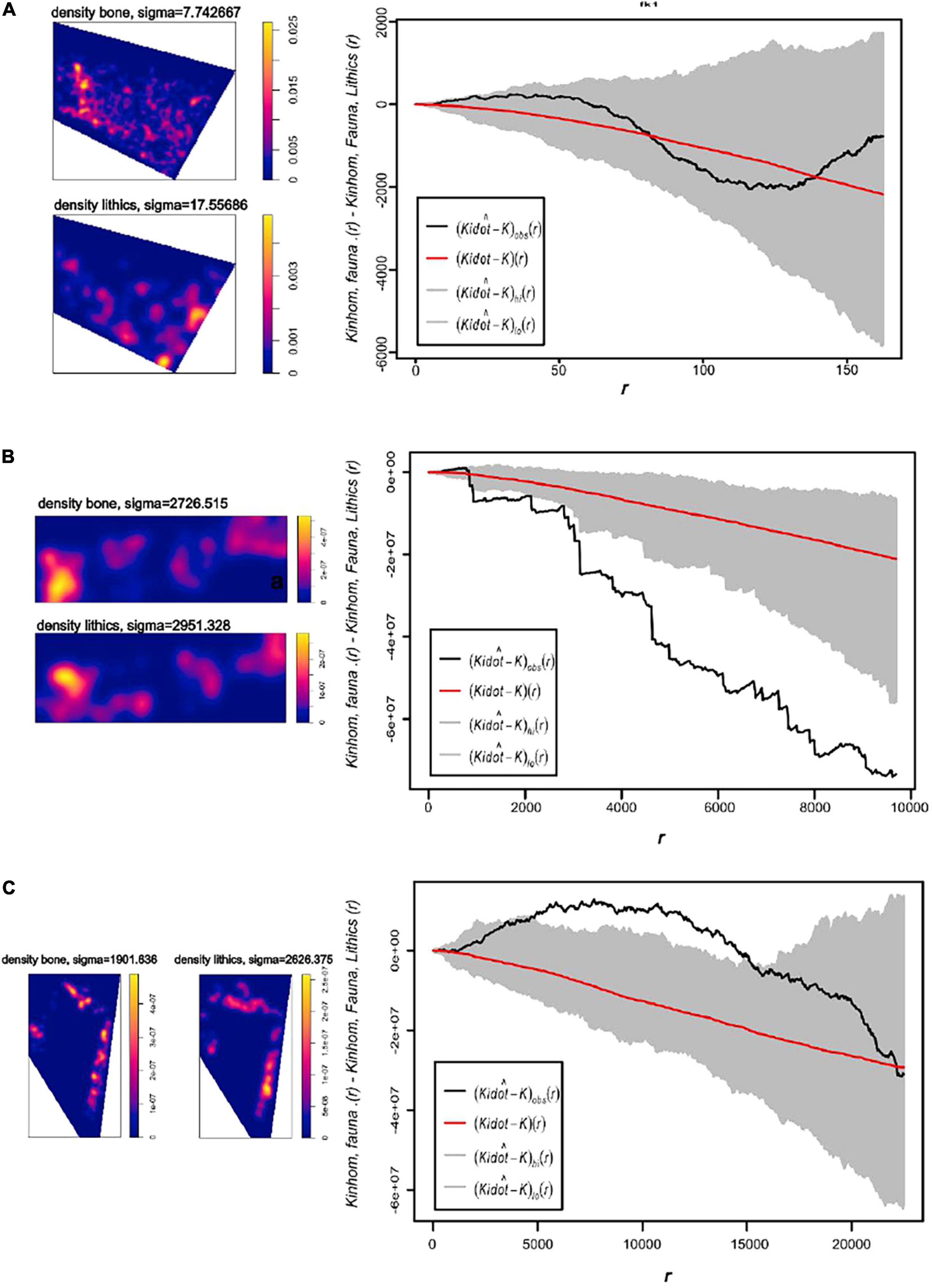
Figure 9. (A) Kernel density maps of the distribution of faunal and lithics remains from trench 2 (left), pointwise envelopes of the homogeneous bivariate Kij(r) function, random labeling cases/controls of lithics and faunal remains from trench 2. (B) Kernel density maps of the distribution of faunal and lithics remains from trench 3 (left), Pointwise envelopes of the homogeneous bivariate Kij(r) function, random labeling cases/controls of lithics and faunal remains from trench 3. (C) Kernel density maps of the distribution of faunal and lithics remains from trench 5 (left), pointwise envelopes of the homogeneous bivariate Kij(r) function, random labeling cases/controls of lithics and faunal remains from trench 5.
The presence of weathering and rolled bones in the assemblage suggests they were affected by hydraulic currents, probably of varying intensities. The presence of rounded bones, mainly in trench 3 implies the existence of a water flow, this is consistent with previous paleo geographic studies (Mercader et al., 2021) indicating an overlapping paleo lake shore expansion in trench 3, fluvial floodplain paleosols in trench 5 and layered fluvial channel belt deposits in trench 2. Biochemical modifications resulting from fungi/bacteria are also present on the fauna an aspect common in bones deposited in moist areas (Child, 1995; Sharmin et al., 2003). In addition, weathering could contribute to the reduction of bones into small fragments until they disappear (Behrensmeyer, 1978). On the other hand, the water currents tend to segregate the bones in relation to their density and shape, thus separating the diaphyses from the epiphyses (Yravedra et al., 2017a), therefore both processes could contribute to trench 3 assemblage formation. However, as the weathering identified is mixed both in the lower and medium-high grades its influence cannot be definitively determined.
Diagenetic processes recorded on the specimens such as varying weathering levels indicate perhaps different periods of accumulation and exposure which may have played a role in the material reduction processes. The time of sub aerial exposure of the specimens recorded is minimal and unlikely to have caused the breakage recorded in the sample. There is minimal plant action preserved in the form of root etching and moss/lichen corrosion and a portion of the assemblage is encrusted pointing to the possibility of being accumulated as bioclasts or sedimentary particles (Behrensmeyer, 1975). These factors suggest that the fauna set correspond to different depositional events, some of them independent, which may have lasted several years. Ewass represents a spatial and accumulative palimpsest, in which various independent events of different natures and characteristics are recorded in the same place, overlaying one another. Overall, neither hominin nor carnivore bone modifications are common though it must be considered that a number of bones exhibit poor preservation due to influence from diagenetic influences. Several indices show high ravaging and even competition (Marean and Spencer, 1991; Blumenschine, 1993; Domínguez-Rodrigo et al., 2002, 2007). However, it must be pointed out that attrition can cause the disappearance of lighter bone elements with interosseous fat content.
We note that anthropic activity at early Pleistocene sites, though scarce at first sight, may have been greater than what has been identified through surface modifications alone (Pineda and Saladié, 2022). The extremely low representation of anthropic marks renders further inferences about carnivore/hominin relations unreliable (cf. Blumenschine, 1986; Capaldo, 1997; Selvaggio, 1998; Domínguez-Rodrigo, 2001; Domínguez-Rodrigo and Barba, 2006; Pante, 2013; Hodgkins et al., 2016), although the existence of cut marks on a rib indicates early hominin access (Bunn and Ezzo, 1993) to at least one carcass. Carnivore impact on the faunal assemblage is also scarce, with a mere 2% of the remains showing damage. Modifications on long bone mid shafts and vertebrae suggest primary access, as they have high flesh content (Blumenschine, 1986; Domínguez-Rodrigo and Barba, 2006). Modification distribution on bones and bone portions conflicts with the proposal by Capaldo (1995), for whom tooth mark percentages <15% signify secondary access to meat. Bailey and Galanidou (2009), however, state that mixed agency in spatial palimpsests gives rise to datasets unable to resolve access rank by hominins versus carnivores.
The presence of hominin groups and other predators at Ewass Oldupa is consistent with other Pleistocene sites, which have indicated hominin access to resources (Egeland et al., 2004; Domínguez-Rodrigo and Barba, 2006; Baquedano et al., 2012), with some sites, such as Bouri (Sahle et al., 2017) in Ethiopia and FwJj14A, FwJj14B, and GaJi14 in Koobi Fora, recording cut marked assemblages with no lithic association (Pobiner et al., 2008). The presence of minimal anthropic action may also point to a brief hominin occupation of the site as studies of Pleistocene sites in Koobi Fora (Pobiner et al., 2008; Braun et al., 2010) indicate that factors such as raw material location/distance for lithics influenced choice of occupation sites. Lithic studies at Ewass indicate that the Oldowan hominins were utilizing both local (about 400 m) and outsourced materials (12 km away). This could have played a role in the utility of the site resulting in the preservation of minimal anthropogenic signals (Pineda and Saladié, 2022). Though it is difficult to determine the level of influence of the agents involved in the accumulation of the assemblage, the presence of Oldowan hominins in an environment with levels of competition suggests they had control over the environment.
Faunal analysis also shows an indication of open habitats with mosaics of bushland/woodland occurring around Ewass Oldupa. This is supported by the presence of both open and closed habitat species tribes such as Reduncini and Tragelaphini, as well as Theropithecus, along with water dependent taxa, such as Crocodylae sp. Previous faunal studies in Ewass indicated mosaics of grassy and wooded environments as well as perennial water sources. Phytoliths indicated an abundance of woody dicots and grassland encroachment on woodlands (Mercader et al., 2021). This not only corresponds to studies from other Bed I sites (Domínguez-Rodrigo et al., 2007, 2010, 2014; Aramendi et al., 2017), but also from other Pleistocene sites in the Omo-Turkana basin which record faunal turnovers around 2.5–1.8 ma indicating presence of diverse environments and biomes (Bobe and Leakey, 2009; Beyene et al., 2013) and their exploitation by archaic hominins. Taxonomic diversity in Ewass may point to the existence of various habitats or changing climates perhaps suggesting the ability of Oldowan hominins to exploit and utilize mixed biomes. This adaptability can be considered as an aspect of behavioral complexity and flexibility in Oldowan hominins.
Hominins have shown preference for riparian woodlands (Domínguez-Rodrigo et al., 2007) as recorded in other Bed I sites such as FLK 22(Zinj) and FLK N (Ashley et al., 2010) to establish the residential sites. Egeland (2014) argues that Olduvai Pleistocene hominins were likely to use and store lithic materials in wooded habitats due to the cover they provide. However, it is plausible that this preference was also influenced by the presence of water bodies and number of taxa supported by mixed habitats evidenced by the diverse hominin diet recorded in Pleistocene sites in East Africa (Blumenschine and Pobiner, 2007; Braun et al., 2010; Lüdecke et al., 2018). Despite the independence between the deposition of the faunal remains and hominin activity at Ewass, an aspect present in other Bed I sites such as AMK (Aramendi et al., 2017) the site was used or traveled by hominins.
Behavioral complexity and flexibility present in Oldowan hominins imply that they may have created more than the typological sites currently in use to categorize Pleistocene land use (Egeland, 2014). However, the human presence recorded through the lithic industry, but also cut and percussion marks, indicates the reutilization of the area by foragers. The deposition of a large part of the fauna specimens seems to have occurred in independent episodes, which would also support the existence of independence with the deposition and accumulation of the lithic industry. The mixture of depositional events at Ewass may be a representation of landscape utility influenced by Oldowan flexibility that archeologists may not have categorized yet.
Conclusion
The taxonomic, zooarchaeological, and taphonomic analyses presented here reconstruct faunal assemblages from the early Pleistocene, 2–1.8 ma. While contemporaneous lithics demonstrate hominin occupation, anthropogenic modification of faunal remains is minimal, and so is carnivore access. These mixed features occur in contemporaneous East African assemblages. Human ecology reveals capability to exploit diverse habitats, as inferred from trophic competition levels, palaeoenvironments, and the adaptive versatility to exploit diverse taxa from mixed habitats. The taphonomic signals recorded speak to different periods of accumulation, even though site functionality over time remains unknown. It is possible that it was occupied by Oldowan hominins briefly, yet repeatedly, due to the presence of water and woodland. At present, we cannot demonstrate a link between lithic and faunal accumulations, although other sites at the Gorge display a similar concurrence, perhaps indicating that Bed I sites can fall under typologies yet to be understood for what they tell us about Oldowan behavioral complexity.
Data Availability Statement
All datasets used in this study have been provided as Supplementary Tables 4–6.
Author Contributions
PA, PS, and JM conceptualized and designed the experiments. PA performed the experiments. PA, PS, JM, and AMo analyzed the data and statistics. AMo, AMw, and PB revised existing literature and interpreted the data. All authors contributed to the writing of this article and the final submittal.
Funding
This work was supported by the Canadian Social Sciences and Humanities Research Council under its Partnership Grant Program No. 895-2016-1017. The Tanzania Commission for Science and Technology authorized this work under permit no. 2018-112-NA-2018-36. The Tanzanian Ministry of Natural Resources and Tourism, through its Antiquities Division, granted us permission to carry out this work (14/2017/2018) and authorities at the Ngorongoro Conservation Area allowed us to enter the protected area (BE.504/620/01/53). The export license for the materials presented in this study were obtained from the Antiquities Division (EA.150/297/01: 5/2018/2019) and the Tanzanian Executive Secretary from the Mining Commission (00001258). PA and PS are supported by AGAUR (project no. 2017 SGR-1040) and the URV (2018PFR-URV-B2-91). PA’s doctoral program was funded by the Canadian Social Sciences and Humanities Research Council as well as the Leakey Foundation Franklin Mosher Baldwin Fellowship.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors wish to acknowledge the important contributions to the knowledge and information presented here by the Maasai communities in Oldupai Gorge.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.895305/full#supplementary-material
Supplementary Figures 1 and 2 | Cut mark furrow depth and shape images created using a hirox kh 7800 microscope. The graphs indicate the cutmark profile for the two cutmarks present in the long bone specimen (Figure 5 top).
Supplementary Figure 3 | Cut mark furrow depth and shape images created using a hirox kh 7800 microscope. The graphs indicate the cutmark profile for the rib (Figure 5 bottom).
Supplementary Table 1 | Ewass normed NISP (nNisp) calculations for various skeletal elements. The workbook contains the calculations for the normed NISP values for all skeletal remains from Ewass Oldupa.
Supplementary Table 2 | Ewass Oldupa tooth mark measurements. The columns in the excel sheet contain information on the specimen number, and measurements for lengths abbreviated L and widths W. Various specimens record more than one tooth mark.
Supplementary Table 3 | Species, tribe and age of Ewass Oldupa dental materials analyzed for ternary plot. The excel workbook contains the information on all tooth elements recorded at Ewass Oldupa as well as the dental remains used for the ternary plot.
Supplementary Tables 4–6 | Workbooks containing datasets for specimens from Ewass Oldupa trenches 2, 3, and 5 presented in this study.
References
Andrés, M., Gidna, A. O., Yravedra, J., and Domínguez-Rodrigo, M. (2012). A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol. Anthropol. Sci. 4, 209–219. doi: 10.1007/s12520-012-0093-4
Aramendi, J., Uribelarrea, D., Arriaza, M. C., Arráiz, H., Barboni, D., Yravedra, J., et al. (2017). The paleoecology and taphonomy of AMK (Bed I, Olduvai Gorge) and its contributions to the understanding of the “Zinj” paleolandscape. Palaeogeography Palaeoclimatol. Palaeoecol. 488, 35–49. doi: 10.1016/j.palaeo.2017.02.036
Arriaza, M. C., and Dominguez-Rodrigo, M. (2016). When felids and hominins ruled at Olduvai Gorge: a machine learning analysis of the skeletal profiles of the non-anthropogenic Bed I sites. Quaternary Sci. Rev. 139, 43–52. doi: 10.1016/j.quascirev.2016.03.005
Ashley, G. M., Barboni, D., Dominguez-Rodrigo, M., Bunn, H. T., Mabulla, A. Z., Diez-Martin, F., et al. (2010). Paleoenvironmental and paleoecological reconstruction of a freshwater oasis in savannah grassland at FLK North, Olduvai Gorge, Tanzania. Quaternary Res. 74, 333–343. doi: 10.1016/j.yqres.2010.08.006
Bailey, G., and Galanidou, N. (2009). Caves, palimpsests and dwelling spaces: examples from the Upper Palaeolithic of south-east Europe. World Archaeol. 41, 215–241.
Baquedano, E., Domínguez-Rodrigo, M. U., and Musiba, C. (2012). An experimental study of large mammal bone modification by crocodiles and its bearing on the interpretation of crocodile predation at FLK Zinj and FLK NN3. J. Archaeol. Sci. 39, 1728–1737.
Behrensmeyer, A. K. (1975). The taphonomy and paleoecology of Plio-Pleistocene vertebrate assemblages east of Lake Rudolf, Kenya. Bull. Museum Comp. Zool. 146, 473–578.
Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology 4, 150–162.
Berman, M., and Diggle, P. (1989). Estimating weighted integrals of the second-order intensity of a spatial point process. J. R. Statistical Soc. Series B (Methodological) 51, 81–92.
Beyene, Y., Katoh, S., WoldeGabriel, G., Hart, W. K., Uto, K., Sudo, M., et al. (2013). The characteristics and chronology of the earliest Acheulean at Konso, Ethiopia. Proc. Natl. Acad. Sci U S A. 110, 1584–1591. doi: 10.1073/pnas.1221285110
Binford, L. R. (1978). Dimensional analysis of behaviour and site structure: learning from an Eskimo hunting stand. Am. Antiquity 43, 330–361. doi: 10.2307/279390
Binford, L. R. (1984). Bones of contention: a reply to Glynn Isaac. Am. Antiquity 49, 164–167. doi: 10.2307/280522
Blumenschine, R. J. (1986). Carcass consumption sequences and the archaeological distinction of scavenging and hunting. J. Hum. Evol. 15, 639–659.
Blumenschine, R. J. (1993). “A carnivore’s view of archaeological bone assemblages,” in From Bones to Behaviour, ed. J. Hudson (Carbondale, IL: The Center for Archaeological Investigations at Southern Illinois University). doi: 10.1016/j.jhevol.2017.08.004
Blumenschine, R. J. (1995). Percussion marks, tooth marks, and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 29, 21–51. doi: 10.1006/jhev.1995.1046
Blumenschine, R. J., and Cavallo, J. A. (1992). Scavenging and human evolution. Sci. Am. 267, 90–97. doi: 10.1038/scientificamerican1092-90
Blumenschine, R. J., and Pobiner, B. L. (2007). “Zooarchaeology and the ecology of Oldowan hominin carnivory,” in Evolution of the Human Diet: the Known, the Unknown, and the Unknowable, ed. P. Ungar (Oxford: Oxford University Press).
Blumenschine, R. J., Stanistreet, I. G., Njau, J. K., Bamford, M. K., Masao, F. T., Albert, R. M., et al. (2012). Environments and hominin activities across the FLK Peninsula during Zinjanthropus times (1.84 Ma), Olduvai Gorge, Tanzania. J. Hum. Evol. 63, 364–383. doi: 10.1016/j.jhevol.2011.10.001
Bobe, R., and Leakey, M. G. (2009). “Ecology of Plio-Pleistocene mammals in the Omo—Turkana Basin and the emergence of Homo,” in The First Humans–Origin and Early Evolution of the Genus Homo, eds F. E. Grine, J. G. Fleagle, and R. E. Leakey (Dordrecht: Springer).
Brain, C. K. (1967). Hottentot food remains and their bearing on the interpretation of fossil bone assemblages. Sci. Papers Namib Desert Res. Station 32, 1–7. doi: 10.1525/9780520955110-003
Brain, C. K. (1974). Some suggested procedures in the analysis of bone accumulations from southern African Quaternary sites. Ann. Transvaal Museum 29, 1–8.
Braun, D. R., Harris, J. W., Levin, N. E., McCoy, J. T., Herries, A. I., Bamford, M. K., et al. (2010). Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. Proc. Natl. Acad. Sci. 107, 10002–10007. doi: 10.1073/pnas.1002181107
Bunn, H. T. III (1986a). Meat-eating and Human Evolution: Studies on the Diet and Subsistence Patterns of Plio-Pleistocene Hominids in East Africa. Doctoral dissertation, Berkeley: University of California.
Bunn, H. T. (1986b). Patterns of skeletal representation and hominid subsistence activities at Olduvai Gorge, Tanzania, and Koobi Fora, Kenya. J. Hum. Evol. 15, 673–690. doi: 10.1016/s0047-2484(86)80004-5
Bunn, H. T., and Ezzo, J. A. (1993). Hunting and scavenging by Plio-Pleistocene hominids: nutritional constraints, archaeological patterns, and behavioural implications. J. Archaeol. Sci. 20, 365–398. doi: 10.1006/jasc.1993.1023
Bunn, H. T., and Pickering, T. R. (2010). Methodological recommendations for ungulate mortality analyses in paleoanthropology. Quaternary Res. 74, 388–394. doi: 10.1016/j.yqres.2010.07.013
Bunn, H. T., Kroll, E. M., Ambrose, S. H., Behrensmeyer, A. K., Binford, L. R., Blumenschine, R. J., et al. (1986). Systematic butchery by Plio/Pleistocene hominids at Olduvai Gorge, Tanzania [and comments and reply]. Curr. Anthropol. 27, 431–452. doi: 10.1086/203467
Capaldo, S. D. (1995). Inferring Hominid and Carnivore Behaviour from Dual-patterned Archaeofaunal Assemblages. Doctoral dissertation, New Brunswick, NJ: Rutgers The State University of New Jersey-New Brunswick.
Capaldo, S. D. (1997). Experimental determinations of carcass processing by Plio-Pleistocene hominids and carnivores at FLK 22 (Zinjanthropus), Olduvai Gorge, Tanzania. J. Hum. Evol. 33, 555–597. doi: 10.1006/jhev.1997.0150
Capaldo, S. D. (1998). Simulating the formation of dual-patterned archaeofaunal assemblages with experimental control samples. J. Archaeol. Sci. 25, 311–330. doi: 10.1006/jasc.1997.0238
Child, A. M. (1995). Microbial taphonomy of archaeological bone. Stud. Conserv. 40, 19–30. doi: 10.2307/1506608
Coard, R. (1999). One bone, two bones, wet bones, dry bones: transport potentials under experimental conditions. J. Archaeol. Sci. 26, 1369–1375. doi: 10.1006/jasc.1999.0438
Day, M. H., and Molleson, T. I. (1976). The puzzle from JK2—A femur and a tibial fragment (OH 34) from Olduvai Gorge, Tanzania. J. Hum. Evol. 5, 455–465. doi: 10.1016/0047-2484(76)90086-5
de la Torre, I. G., McHenry, L. J., Njau, J., and Pante, M. D. (2012). The origins of the Acheulean at Olduvai Gorge (Tanzania): a new paleoanthropological project in East Africa. Archaeol. Int. 15, 89–98.
Delaney-Rivera, C., Plummer, T. W., Hodgson, J. A., Forrest, F., Hertel, F., and Oliver, J. S. (2009). Pits and pitfalls: taxonomic variability and patterning in tooth mark dimensions. J. Archaeol. Sci. 36, 2597–2608. doi: 10.1016/j.jas.2009.08.001
Díez-Martín, F., Sánchez Yustos, P., Uribelarrea, D., Baquedano, E., Mark, D. F., Mabulla, A., et al. (2015). The origin of the Acheulean: the 1.7 million-year-old site of FLK West, Olduvai Gorge (Tanzania). Sci. Rep. 5:17839. doi: 10.1038/srep17839
Domínguez-Rodrigo, M. (1997). Meat-eating by early hominids at the FLK 22Zinjanthropussite, Olduvai Gorge (Tanzania): an experimental approach using cut-mark data. J. Hum. Evol. 33, 669–690. doi: 10.1006/jhev.1997.0161
Domínguez-Rodrigo, M. (1999). Flesh availability and bone modifications in carcasses consumed by lions: palaeoecological relevance in hominid foraging patterns. Palaeogeography Palaeoclimatol. Palaeoecol. 149, 373–388. doi: 10.1016/s0031-0182(98)00213-2
Domínguez-Rodrigo, M. (2001). A study of carnivore competition in riparian and open habitats of modern savannas and its implications for hominid behavioural modelling. J. Hum. Evol. 40, 77–98. doi: 10.1006/jhev.2000.0441
Domínguez-Rodrigo, M. (2002). Hunting and scavenging by early humans: the state of the debate. J. World Prehistory. 16, 1–54. doi: 10.1038/s41598-020-63250-x
Domínguez-Rodrigo, M., and Barba, R. (2006). New estimates of tooth mark and percussion mark frequencies at the FLK Zinj site: the carnivore-hominid-carnivore hypothesis falsified. J. Hum. Evol. 50, 170–194. doi: 10.1016/j.jhevol.2005.09.005
Domínguez-Rodrigo, M., and Cobo-Sánchez, L. (2017). A spatial analysis of stone tools and fossil bones at FLK Zinj 22 and PTK I (Bed I, Olduvai Gorge, Tanzania) and its bearing on the social organization of early humans. Palaeogeography Palaeoclimatol. Palaeoecol. 488, 21–34. doi: 10.1016/j.palaeo.2017.04.010
Domínguez-Rodrigo, M., and Organista, E. (2007). “Natural background bone assemblages and their ravaging stages in Olduvai Bed I,” in Deconstructing Olduvai: a Taphonomic Study of the Bed I Sites (Dordrecht: Springer), 201–215. doi: 10.1007/978-1-4020-6152-3_11
Dominguez-Rodrigo, M., and Pickering, T. R. (2017). The meat of the matter: an evolutionary perspective on human carnivory. Azania: Archaeol. Res. Africa 52, 4–32. doi: 10.1080/0067270x.2016.1252066
Domínguez-Rodrigo, M., Baquedano, E., Barba, R., Uribelarrea, D., and Gidna, A. (2019). The river that never was: fluvial taphonomy at Olduvai Bed I and II sites and its bearing on early human behavior. Quaternary Int. 526, 26–38. doi: 10.1016/j.quaint.2019.09.038
Domínguez-Rodrigo, M., Barba, R., and Egeland, C. P. (2007). Deconstructing Olduvai: a Taphonomic Study of the Bed I Sites. Berlin: Springer Science & Business Media.
Domínguez-Rodrigo, M., de La Torre, I., De Luque, L., Alcalá, L., Mora, R., Serrallonga, J., et al. (2002). The ST site complex at Peninj, West Lake Natron, Tanzania: implications for early hominid behavioural models. J. Archaeol. Sci. 29, 639–665. doi: 10.1006/jasc.2001.0768
Domínguez-Rodrigo, M., Diez-Martín, F., Yravedra, J., Barba, R., Mabulla, A., Baquedano, E., et al. (2014). Study of the SHK Main Site faunal assemblage, Olduvai Gorge, Tanzania: implications for Bed II taphonomy, paleoecology, and hominin utilization of megafauna. Quaternary Int. 322, 153–166. doi: 10.1016/j.quaint.2013.09.025
Domínguez-Rodrigo, M., Pickering, T. R., and Bunn, H. T. (2010). Configurational approach to identifying the earliest hominin butchers. Proc. Natl. Acad. Sci. U S A. 107, 20929–20934. doi: 10.1073/pnas.1013711107
Egeland, C. P. (2012). “Model hominid lifeways during the Oldowan,” in Stone Tools and Fossil Bones: Debates in the Archaeology of Human Origins, ed. M. Domínguez-Rodrigo (Cambridge: Cambridge University Press).
Egeland, C. P. (2014). Taphonomic estimates of competition and the role of carnivore avoidance in hominin site use within the Early Pleistocene Olduvai Basin. Quaternary Int. 322, 95–106. doi: 10.1016/j.quaint.2013.11.021
Egeland, C. P., and Domínguez-Rodrigo, M. (2008). Taphonomic perspectives on hominid site use and foraging strategies during Bed II times at Olduvai Gorge, Tanzania. J. Hum. Evol. 55, 1031–1052. doi: 10.1016/j.jhevol.2008.05.021
Egeland, C. P., Pickering, T. R., Domínguez-Rodrigo, M., and Brain, C. K. (2004). Disentangling Early Stone Age palimpsests: determining the functional independence of hominid-and carnivore-derived portions of archaeofaunas. J. Hum. Evol. 47, 343–357. doi: 10.1016/j.jhevol.2004.08.004
Faith, J. T. (2007). Changes in reindeer body part representation at Grotte XVI, Dordogne, France. J. Archaeol. Sci. 34, 2003–2011. doi: 10.1016/j.jas.2007.01.014
Faith, J. T., Domínguez-Rodrigo, M., and Gordon, A. D. (2009). Long-distance carcass transport at Olduvai Gorge? a quantitative examination of Bed I skeletal element abundances. J. Hum. Evol. 56, 247–256. doi: 10.1016/j.jhevol.2008.12.008
Favreau, J., Soto, M., Nair, R., Bushozi, P. M., Clarke, S., DeBuhr, C. L., et al. (2020). Petrographic characterization of raw material sources at oldupai gorge, Tanzania. Front. Earth Sci. 8:158. doi: 10.3389/feart.2020.00158
Fernandez-Jalvo, Y., and Andrews, P. (2016). Atlas of taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification. Berlin: Springer.
Fernández-Jalvo, Y., Denys, C., Andrews, P., Williams, T., Dauphin, Y., and Humphrey, L. (1998). Taphonomy and palaeoecology of Olduvai bed-I (Pleistocene, Tanzania). J. Hum. Evol. 34, 137–172. doi: 10.1006/jhev.1997.0188
Fiorillo, A. R. (1988). Taphonomy of hazard homestead quarry (Ogallala Group), Hitchcock County, Nebraska. Rocky Mountain Geol. 26, 57–97.
Galán, A. B., Rodríguez, M., De Juana, S., and Domínguez-Rodrigo, M. (2009). A new experimental study on percussion marks and notches and their bearing on the interpretation of hammerstone-broken faunal assemblages. J. Archaeol. Sci. 36, 776–784. doi: 10.1016/j.jas.2008.11.003
Gaudzinski-Windheuser, S., Kindler, L., Rabinovich, R., and Goren-Inbar, N. (2010). Testing heterogeneity in faunal assemblages from archaeological sites. Tumbling and trampling experiments at the early-Middle Pleistocene site of Gesher Benot Ya’aqov (Israel). J. Archaeol. Sci. 37, 3170–3190. doi: 10.1016/j.jas.2010.07.018
Grayson, D. K. (1984). Quantitative Zooarchaeology: Topics in the Quantification of Archaeofaunas. Madhya Pradesh: Orlando Acad.
Habermann, J. M., McHenry, L. J., Stollhofen, H., Tolosana-Delgado, R., Stanistreet, I. G., and Deino, A. L. (2016). Discrimination, correlation, and provenance of Bed I tephrostratigraphic markers, Olduvai Gorge, Tanzania, based on multivariate analyses of phenocryst compositions. Sedimentary Geol. 339, 115–133. doi: 10.1016/j.sedgeo.2016.03.026
Hay, R. L. (1963). Stratigraphy of Beds I through IV, Olduvai Gorge, Tanganyika. Science 139, 829–833. doi: 10.1126/science.139.3557.829
Hay, R. L. (1976). Geology of the Olduvai Gorge: A Study of Sedimentation in a Semiarid Basin. Oakland, CA: Univ of California.
Hlusko, L. J., Reiner, W. B., and Njau, J. K. (2015). A one-million-year-old hominid distal ulna from Olduvai Gorge, Tanzania. Am. J. Phys. Anthropol. 158, 36–42. doi: 10.1002/ajpa.22765
Hodgkins, J., Marean, C. W., Turq, A., Sandgathe, D., McPherron, S. J., and Dibble, H. (2016). Climate-mediated shifts in Neandertal subsistence behaviours at Pech de l’Azé IV and Roc de Marsal (Dordogne Valley, France). J. Hum. Evol. 96, 1–8. doi: 10.1016/j.jhevol.2016.03.009
Isaac, G. L. (1978). The Harvey Lecture series, 1977-1978. Food sharing and human evolution: archaeological evidence from the Plio-Pleistocene of east Africa. J. Anthropol. Res. 34, 311–325. doi: 10.1086/jar.34.3.3629782
Kimura, Y. (2002). Examining time trends in the Oldowan technology at Beds I and II, Olduvai Gorge. J. Hum. Evol. 43, 291–321. doi: 10.1006/jhev.2002.0576
Klein, R. G. (1976). The mammalian fauna of the Klasies River mouth sites, southern Cape Province, South Africa. South African Archaeol. Bull. 31, 75–98. doi: 10.2307/3887730
Lam, Y. M., Chen, X., and Pearson, O. M. (1999). Intertaxonomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. Am. Antiquity 64, 343–362.
Leakey, L. S., Hopwood, A. T., and Reck, H. (1951). Olduvai Gorge: A Report on the Evolution of the Hand-axe Culture in Beds I-IV. Cambridge: Cambridge University Press.
Leakey, M. D. (1971). Olduvai Gorge: Excavations in Beds I and II, 1960-1963. Cambridge: Cambridge University Press.
Lüdecke, T., Kullmer, O., Wacker, U., Sandrock, O., Fiebig, J., Schrenk, F., et al. (2018). Dietary versatility of Early Pleistocene hominins. Proc. Natl. Acad. Sci. U S A. 115, 13330–13335. doi: 10.1073/pnas.1809439115
Marean, C. W., and Cleghorn, N. (2003). Large mammal skeletal element transport: applying foraging theory in a complex taphonomic system. J. Taphonomy 1, 15–42.
Marean, C. W., and Spencer, L. M. (1991). Impact of carnivore ravaging on zooarchaeological measures of element abundance. Am. Antiquity 56, 645–658. doi: 10.2307/281542
McHenry, H. M., Brown, C. C., and McHenry, L. J. (2007). Fossil hominin ulnae and the forelimb of Paranthropus. Am. J. Phys. Anthropol. 134, 209–218. doi: 10.1002/ajpa.20656
McHenry, L. J. (2012). A revised stratigraphic framework for Olduvai Gorge Bed I based on tuff geochemistry. J. Hum. Evol. 63, 284–299. doi: 10.1016/j.jhevol.2011.04.010
McHenry, L. J., Njau, J. K., de la Torre, I., and Pante, M. C. (2016). Geochemical “fingerprints” for Olduvai Gorge Bed II tuffs and implications for the Oldowan–Acheulean transition. Quaternary Res. 85, 147–158.
Mercader, J., Akuku, P., Boivin, N., Bugumba, R., Bushozi, P., Camacho, A., et al. (2021). Earliest Olduvai hominins exploited unstable environments~ 2 million years ago. Nat. Commun. 12:3. doi: 10.1038/s41467-020-20176-2
Milo, R. G. (1998). Evidence for hominid predation at Klasies River Mouth, South Africa, and its implications for the behaviour of early modern humans. J. Archaeol. Sci. 25, 99–133.
Olsen, S. L., and Shipman, P. (1988). Surface modification on bone: trampling versus butchery. J. Archaeol. Sci. 15, 535–553.
Organista, E., Domínguez-Rodrigo, M., Egeland, C. P., Uribelarrea, D., Mabulla, A., and Baquedano, E. (2016). Did Homo erectus kill a Pelorovis herd at BK (Olduvai Gorge)? A taphonomic study of BK5. Archaeol. Anthropol. Sci. 8, 601–624.
Pante, M. C. (2010). The Larger Mammal Fossil Assemblages from Beds III and IV, Olduvai Gorge, Tanzania: Implications for the Feeding Behaviour of Homo Erectus. New Brunswick, NJ: Rutgers The State University of New Jersey-New Brunswick.
Pante, M. C. (2013). The larger mammal fossil assemblage from JK2, Bed III, Olduvai Gorge, Tanzania: implications for the feeding behaviour of Homo erectus. J. Hum. Evol. 64, 68–82. doi: 10.1016/j.jhevol.2012.10.004
Pante, M. C., Njau, J. K., Hensley-Marschand, B., Keevil, T. L., Martín-Ramos, C., Peters, R. F., et al. (2018). The carnivorous feeding behaviour of early Homo at HWK EE, Bed II, Olduvai Gorge, Tanzania. J. Hum. Evol. 120, 215–235.
Petraglia, M. D., and Potts, R. (1994). Water flow and the formation of Early Pleistocene artifact sites in Olduvai Gorge, Tanzania. J. Anthropol. Archaeol. 13, 228–254.
Pickering, T. R., Domínguez-Rodrigo, M., Egeland, C. P., and Brain, C. K. (2004). Beyond leopards: tooth marks and the contribution of multiple carnivore taxa to the accumulation of the Swartkrans Member 3 fossil assemblage. J. Hum. Evol. 46, 595–604. doi: 10.1016/j.jhevol.2004.03.002
Pickering, T. R., Domínguez-Rodrigo, M., Heaton, J. L., Yravedra, J., Barba, R., Bunn, H. T., et al. (2013). Taphonomy of ungulate ribs and the consumption of meat and bone by 1.2-million-year-old hominins at Olduvai Gorge. Tanzania. J. Archaeol. Sci. 40, 1295–1309.
Pineda, A., and Saladié, P. (2019). The Middle Pleistocene site of Torralba (Soria, Spain): a taphonomic view of the Marquis of Cerralbo and Howell faunal collections. Archaeol. Anthropol. Sci. 11, 2539–2556.
Pineda, A., and Saladié, P. (2022). Beyond the problem of bone surface preservation in taphonomic studies of early and middle pleistocene open-air sites. J. Archaeol. Method Theory. 1–41.
Plummer, T. W., and Bishop, L. C. (1994). Hominid paleoecology at Olduvai Gorge, Tanzania as indicated by antelope remains. J. Hum. Evol. 27, 47–75.
Pobiner, B. L. (2015). New actualistic data on the ecology and energetics of hominin scavenging opportunities. J. Hum. Evol. 80, 1–6. doi: 10.1016/j.jhevol.2014.06.020
Pobiner, B. L., Rogers, M. J., Monahan, C. M., and Harris, J. W. (2008). New evidence for hominin carcass processing strategies at 1.5 Ma, Koobi Fora, Kenya. J. Hum. Evol. 55, 103–130. doi: 10.1016/j.jhevol.2008.02.001
Potts, R., and Shipman, P. (1981). Cutmarks made by stone tools on bones from Olduvai Gorge. Tanzania. Nature 291, 577–580.
Proffitt, T. (2018). Is there a developed Oldowan A at Olduvai Gorge? a diachronic analysis of the Oldowan in bed I and lower-middle bed II at Olduvai Gorge, Tanzania. J. Hum. Evol. 120, 92–113. doi: 10.1016/j.jhevol.2018.01.006
Renner, I. W., Elith, J., Baddeley, A., Fithian, W., Hastie, T., Phillips, S. J., et al. (2015). Point process models for presence-only analysis. Methods Ecol. Evol. 6, 366–379.
Reynard, J. P. (2014). Trampling in coastal sites: an experimental study on the effects of shell on bone in coastal sediment. Quaternary Int. 330, 156–170.
Reynard, J. P., Discamps, E., Badenhorst, S., van Niekerk, K., and Henshilwood, C. S. (2016a). Subsistence strategies in the southern Cape during the Howiesons Poort: Taphonomic and zooarchaeological analyses of Klipdrift Shelter. South Africa. Quaternary Int. 404, 2–19.
Reynard, J. P., Discamps, E., Wurz, S., van Niekerk, K. L., Badenhorst, S., and Henshilwood, C. S. (2016b). Occupational intensity and environmental changes during the Howiesons Poort at Klipdrift Shelter, southern Cape, South Africa. Palaeogeography Palaeoclimatol. Palaeoecol. 449, 349–364.
Sahle, Y., El Zaatari, S., and White, T. D. (2017). Hominid butchers and biting crocodiles in the African Plio–Pleistocene. Proc. Natl. Acad. Sci. U S A. 114, 13164–13169. doi: 10.1073/pnas.1716317114
Saladié, P., Rodríguez-Hidalgo, A., Díez, C., Martín-Rodríguez, P., and Carbonell, E. (2013). Range of bone modifications by human chewing. J. Archaeol. Sci. 40, 380–397.
Saladié, P., Rodríguez-Hidalgo, A., Huguet, R., Cáceres, I., Díez, C., Vallverdú, J., et al. (2014). The role of carnivores and their relationship to hominin settlements in the TD6-2 level from Gran Dolina (Sierra de Atapuerca, Spain). Quaternary Sci. Rev. 93, 47–66.
Scoon, R. N. (2018). “Oldupai gorge and laetoli,” in In Geology of National Parks of Central/Southern Kenya and Northern Tanzania (Cham: Springer), 115–128.
Selvaggio, M. M. (1994). Carnivore tooth marks and stone tool butchery marks on scavenged bones: archaeological implications. J. Hum. Evol. 27, 215–228.
Selvaggio, M. M. (1998). Evidence for a three-stage sequence of hominid and carnivore involvement with long bones at FLKZinjanthropus, Olduvai Gorge, Tanzania. J. Archaeol. Sci. 25, 191–202.
Sharmin, S., Kishi, F., Sano, A., Kamei, K., Nishimura, K., and Miyaji, M. (2003). Direct invasion of bones by highly pathogenic fungi in an in vitro model and its ecological significance. Nippon Ishinkin Gakkai Zasshi 44, 17–23. doi: 10.3314/jjmm.44.17
Shipman, P., and Rose, J. (1983). Early hominid hunting, butchering, and carcass-processing behaviours: approaches to the fossil record. J. Anthropol. Archaeol. 2, 57–98. doi: 10.1016/0278-4165(83)90008-9
Soto, M., Favreau, J., Campeau, K., Carter, T., Durkin, P. R., Hubbard, S. M., et al. (2020). Systematic sampling of quartzites in sourcing analysis: intra-outcrop variability at Naibor Soit, Tanzania (part I). Archaeol. Anthropol. Sci. 12, 1–4.
Steele, T. E., and Weaver, T. D. (2002). The modified triangular graph: a refined method for comparing mortality profiles in archaeological samples. J. Archaeol. Sci. 29, 317–322. doi: 10.1006/jasc.2001.0733
Stiner, M. C. (1990). The use of mortality patterns in archaeological studies of hominid predatory adaptations. J. Anthropol. Archaeol. 9, 305–351.
Toth, N. (1993). “Early stone industries and inferences regarding language and cognition,” in Tools, Language and Cognition in Human Evolution, eds K. R. Gibson and T. Ingold (Cambridge: Univ.), 346–362.
Villa, P., and Mahieu, E. (1991). Breakage patterns of human long bones. J. Hum. Evol. 21, 27–48. doi: 10.1016/0047-2484(91)90034-s
Yravedra, J., and Domínguez-Rodrigo, M. (2009). The shaft-based methodological approach to the quantification of long limb bones and its relevance to understanding hominid subsistence in the Pleistocene: application to four Palaeolithic sites. J. Quaternary Sci. Published Quaternary Res. Assoc. 24, 85–96.
Yravedra, J., Diez-Martín, F., Egeland, C. P., Maté-González, M. Á, Palomeque-González, J. F., and Arriaza, M. C. (2017a). FLK West (Lower Bed II, Olduvai Gorge, Tanzania): a new early Acheulean site with evidence for human exploitation of fauna. Boreas 46, 816–830.
Yravedra, J., Maté-González, M. Á, Palomeque-González, J. F., Aramendi, J., Estaca-Gómez, V., San Juan Blazquez, M., et al. (2017b). A new approach to raw material use in the exploitation of animal carcasses at BK (Upper Bed II, Olduvai Gorge, Tanzania): a micro-photogrammetric and geometric morphometric analysis of fossil cut marks. Boreas 46, 860–873. doi: 10.1111/bor.12224
Keywords: faunal taphonomy, paleoanthropology, paleobiology, Oldowan, Oldupai Gorge, Tanzania
Citation: Akuku P, Saladié P, Mohamed A, Mwambwiga A, Bushozi P and Mercader J (2022) Faunal Assemblages From Lower Bed I (Oldupai Gorge, Tanzania). Front. Ecol. Evol. 10:895305. doi: 10.3389/fevo.2022.895305
Received: 13 March 2022; Accepted: 26 April 2022;
Published: 27 May 2022.
Edited by:
Pasquale Raia, University of Naples Federico II, ItalyReviewed by:
Carlo Meloro, Liverpool John Moores University, United KingdomJulien Louys, Griffith University, Australia
Copyright © 2022 Akuku, Saladié, Mohamed, Mwambwiga, Bushozi and Mercader. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pamela Akuku, pakuku@iphes.cat