Cross-Kingdom Extracellular Vesicles EV-RNA Communication as a Mechanism for Host–Pathogen Interaction
 Isadora Filipaki Munhoz da Rocha1
Isadora Filipaki Munhoz da Rocha1  Rafaela Ferreira Amatuzzi1
Rafaela Ferreira Amatuzzi1  Aline Castro Rodrigues Lucena2
Aline Castro Rodrigues Lucena2  Helisson Faoro2
Helisson Faoro2  Lysangela Ronalte Alves1*
Lysangela Ronalte Alves1*- 1Gene Expression Regulation Laboratory, Carlos Chagas Institute, Oswaldo Cruz Foundation (FIOCRUZ), Curitiba, Brazil
- 2Laboratory for Applied Science and Technology in Health, Carlos Chagas Institute, Oswaldo Cruz Foundation (FIOCRUZ), Curitiba, Brazil
The extracellular vesicle (EVs) traffic has been highlighted as a very important pathway of cellular communication. EVs are produced by prokaryotes and eukaryotes organisms and can carry molecules to help maintain homeostasis, responding to general disbalance, infections, and allowing rapid modulation of the immune system. In the context of infection, EVs from both the host and the pathogen have been identified as playing roles in the recruitment of immunological molecules that can lead to the resolution of the infection or the host’s defeat. Bacterial vesicles RNA cargo play roles in the host cell by regulating gene expression and modulating immune response. In fungi the RNA molecules present in EVs are diverse and participate in communication between the host and pathogenic fungi. Little is known about how cross-kingdom sRNA trafficking occurs, although in recent years, there has been an increase in studies that relate EV participation in sRNA delivery. This review aims to elucidate and update the reader concerning the role of extracellular vesicles, with emphasis in the RNA content. We describe the EVs during infection from the host point-of-view, as well as the bacteria and fungi pathogens producing EVs that help the establishment of the disease.
Introduction
Cell communication is crucial for organisms to maintain homeostasis and respond to adverse conditions, as during an infection. Distinct pathways are involved in cell communication, such as direct cell contact and molecular secretion and the transfer of extracellular vesicles (Raposo and Stoorvogel, 2013). Extracellular vesicles (EVs) are defined as cell-derived particles delimited by a lipid bilayer that cannot replicate and can carry different cargos across the organism; EVs include entities such as apoptotic bodies, exosomes, and microvesicles (Koliha et al., 2016; Théry et al., 2018).
EVs are produced by several organisms, ranging from prokaryotes to mammals and plants (Yáñez-Mó et al., 2015), and every known cell is capable of producing them (Mardahl et al., 2019). The secretion of extracellular vesicles is regulated by various mechanisms that culminate with shedding at the plasma membrane, which can occur spontaneously or in response to certain conditions (Théry et al., 2009). The EV content is closely related to cell-type specificity and is influenced by the physiological or pathological state of the cell (Kalluri and LeBleu, 2020)
Regarding EV classification, there is still no consensus. The difficulty in classifying EVs is attributed to their overlapping sizes, the similar composition of EV subclasses and the lack of knowledge about their biogenesis as well as isolation technique limitations. Currently, EVs are classified into three different subgroups: apoptotic bodies, microvesicles, and exosomes (Théry et al., 2018). Microvesicles and apoptotic bodies are released by plasma membrane budding and their size varies from approximately 50 nm to 1 μm in diameter (Kalluri and LeBleu, 2020). Exosomes have endosomal origin; therefore, they are involved in specific intracellular regulatory processes that determine the exosome content. The size ranges from approximately 40 to 200 nm in diameter (Kalluri and LeBleu, 2020). The biogenesis of exosomes depends on several cell stages (Yáñez-Mó et al., 2015). First, the plasma membrane undergoes invagination associated with soluble molecules from the extracellular milieu, and then, the early-sorting endosome forms. Next, early endosomes mature into late-sorting endosomes, which form multivesicular bodies with intraluminal vesicles in the lumen of the organelle (Yáñez-Mó et al., 2015). When multivesicular bodies fuse with the plasma membrane, they release the exosomes into the extracellular milieu (Hessvik and Llorente, 2018). ESCRT (endosomal sorting complexes required for transport) machinery and tetraspanins are important for exosome biogenesis (Colombo et al., 2013). Their depletion is related to exosome-secretion reduction (Hurwitz et al., 2016; Hurwitz et al., 2017). Other molecules, such as ceramides and sphingomyelinases, are also essential in vesicular transport and are involved in exosome biogenesis processes, such as membrane deformation, fission and fusion (McMahon and Boucrot, 2015). The origin of an extracellular vesicle may be traced by the surface proteins that resemble the cell. Among them is the tetraspanin family, whose proteins are commonly used as EV markers, for example, CD63, CD81, CD9 (Raposo and Stoorvogel, 2013), and the more recently discovered HSP70 (Van Niel et al., 2018). Importantly, so far, there is not a single marker—or a set of markers—that can identify all types of vesicles, and the distribution of tetraspanins among these subtypes is variable (Koliha et al., 2016). The characteristics and composition of EVs differ even depending on the cellular and environmental state (Yáñez-Mó et al., 2015).
The main functions of EVs related to cell communication include molecular transfer, changes in gene expression and cell surface rearrangement (Raeven et al., 2018). In humans, EVs are involved in several physiological processes, such as tissue regeneration (Teng et al., 2015), reproductive biology (Simon et al., 2018) and blood coagulation (Heijnen et al., 2015). They can also act in presenting antigens, stimulating immune responses and tolerogenic effects, immunosuppression, angiogenesis, tumor progression, and the transmission of virulence factors (Raposo and Stoorvogel, 2013). Moreover, they have already been linked with the aging process, cancer (Joncas et al., 2019) and stem cells differentiation (Tatischeff, 2019). Animal and plant pathogen/parasite EV secretion is used to aid survival, cell communication and pathogenesis (Yáñez-Mó et al., 2015). Inside the host, EVs derived from the parasite are related to host modulation, allowing the recruitment of specific immune cells and contributing to the parasite’s life cycle and reproduction (Yáñez-Mó et al., 2015; Dong et al., 2019).
EVs are also studied for drug delivery purposes, due to their biocompatible composition and availability in the organism, as it occurs with encapsulated compounds. The variety of compounds carried inside EVs also makes them attractive in biomarker research, especially in liquid biopsies, which are less invasive and methodologically demanding to execute than other diagnostic procedures (Kalluri and LeBleu, 2020).
Many different biomolecules are carried by EVs, including proteins, lipids and nucleic acids (Yáñez-Mó et al., 2015). In mammalian cells, RNA molecules present in the EVs have been highlighted for their capacity to be internalized by the recipient cell and regulate gene expression (O’Brien et al., 2020). The RNAs identified in EVs include mRNA (messenger RNA), microRNA, rRNA (ribosomal RNA), tRNA (transfer RNA), sRNA (small RNA), and lncRNA (long noncoding RNA) (Im et al., 2018; Turchinovich et al., 2019). Differences in the RNA composition of EVs are based on the cellular state and the producing cell, such as cancer and infection, and can, therefore, be used as biomarkers of certain cellular conditions (Mittelbrunn et al., 2011; Baglio et al., 2015; Shah et al., 2018; Turchinovich et al., 2019). The mechanism that directs the RNA species into the EVs is not understood. The RNA could be passively incorporated due to an RNA abundance in the cytosol; by RNA-binding proteins (RBPs) through motif recognition or RNA secondary structure recognition, and by specific modifications in RNA or RBPs, such as uridylation, ubiquitylation, sumoylation, and phosphorylation (Gibbings et al., 2009; McKenzie et al., 2016; Mateescu et al., 2017; Ragusa et al., 2017).
In this review, we provide an overview of RNAs enclosed in extracellular vesicles, with an emphasis on both the host and the bacteria and fungi pathogens, that are common agents of infection worldwide.
EVs and Infection: The Host Response
In an infectious environment, host-originated EVs can promote both immunostimulant and immunosuppressive events; they can carry antigenic molecules as well as Major Histocompatibility Complex (MHC) molecules (Mardahl et al., 2019). Antigen presenting cells (APCs) are at the center of studies involving host-originated extracellular vesicles. They are believed to mediate interactions with T and B cells to: a) activate naïve T cells through MHC molecules, b) serve as exogenous antigens for APCs, c) present antigens directly to CD4+ T cells, and d) present antigens to dendritic cells (DCs), which will be loaded into their own MHCs. EVs originating from host cells have also been reported to inhibit IL-8 and TNF, reducing inflammation. Additionally, the immune response can be promoted by EV content through antigen-independent mechanisms (Mardahl et al., 2019; Raposo and Stahl, 2019). On the other hand, EVs can mediate protective messages between cells during stress situations (Tatischeff, 2019). In a model of Conidium infection, the authors identified different EV cargo compositions derived from PMN cells enriched with antimicrobial molecules to respond to the infection (Shopova et al., 2020). It has been shown that the microRNA signature of EVs is different from the in-microRNA signature of the cell (Mittelbrunn et al., 2011; Roderburg et al., 2013). In the context of the immune system, some micro-RNAs, like miR-760, miR-632, miR-654-5p, and miR-671-5p, were loaded into activated T cell EVs and they were more abundant in the EVs compared to the parental cell during immune synapsis (Mittelbrunn et al., 2011). Additionally, vesicles from different species can lead to distinct protection against diseases. For example, EVs from Nippostrongylus brasiliensis, a hookworm that causes chronic gastrointestinal infection in humans, were isolated and injected in mice. After the EV injection, colitis was induced and the EVs protected the animals against the inflammation. However, EVs from the whipworm Trichuris muris did not present the same protective pattern. From RNA-Seq data of N. brasiliensis, 52 differentially expressed microRNAs were identified, and it was shown that they play an important role in inflammatory cytokine suppression and secretion of anti-inflammatory IL-10 in the host cell (Eichenberger et al., 2018).
Within the gut microbiome, host vesicles have been reported to enter microorganisms and regulate their gene expression and growth (Lee, 2019). The gut microbiota is composed of 100-200 distinct bacterial species. Depending on genetics, diet and disease state this composition may vary, and a series of correlations have been described regarding the microRNA-mediated control of the gut microbiota. Liu et al. (2016) identified the most abundant miRNAs present in the feces of human and mice and compared to those observed inside the Evs. The most abundant miRNAs observed in the EVs were miR-1224, miR-2146, miR-2134, miR-483, miR-710, miR-2141, miR-720, miR-155, and miR-34c. The authors showed that some miRNAs were able to enter the bacterial cells and regulate gene expression by RNA alignment. The host miRNA targeted rRNA and ribozyme (RNaseP), but the expression could be induced or repressed, depending on the species analyzed. In addition, using mice defective in the miRNA pathway (Dicer knockouts), it was shown that the KO mice were more susceptible to induced colitis than the Wild type and that fecal miRNA transplantation could help to restore the gut microbiota (Liu et al., 2016). The EV traffic also helps communication between components of the microbiota through biofilm formation or aggregation-derived quorum sensing, promoting the homeostasis of the host and even prevent and fight infections (Morales and Hogan, 2010). The quorum sensing process uses extracellular signals to communicate and coordinate social activities. In Pseudomonas aeruginosa the molecule PQS (Pseudomonas quinolone signal) is an important quorum sensing molecule that is transported into vesicles. When this molecule was removed, it led to problems with cell-cell communication (Mashburn and Whiteley, 2005).
EVs are recognized as important players in the pathological process of sepsis, influencing aspects such as coagulation and hyperinflammation disturbances. A study showed that several microRNAs were dysregulated in septic shock, highlighting exosomal miR-125b-5p that was validated as a survival predictor and miR-26b-5p and miR-199b-5p, which were able to differentiate healthy individuals from septic patients (Reithmair et al., 2017). Bacterial vesicles were introduced intraperitonially in mice that afterwards presented symptoms of sepsis-like inflammation and eventually died, highlighting that the EVs were sufficient to trigger the host inflammatory response (Park et al., 2018). Additionally, it has been described that the pathogen vesicle formation can be affected by the antibiotic choice in the treatment of sepsis—a great contributor to treatment success—making it even more important to understand the phenomena surrounding the EV-cargo-microbe-host system (Dagnelie et al., 2019).
In addition to the host cell response to infections, one must consider the role of the EVs being shed by the pathogens; the next topics are focused on the EVs produced by common pathogens identified in worldwide infections—bacteria and fungi.
Microorganisms—EV Shedding and Their Roles in Cell Communication and Disease
Bacterial Membrane Vesicles
The bacterial membrane vesicles (MVs) are composed of a lipid bilayer and have a size ranging from 20 to 400 nm. As described for eukaryotes, there are different MV categories, varying according to their structure, composition and origin (Toyofuku et al., 2019). The outer membrane vesicles (OMVs) are a class of vesicles produced by gram-negative bacteria, derived from the outer membrane, and due to its origin, they are covered by lipopolysaccharides (LPS) (Schwechheimer and Kuehn, 2015). The OMVs biogenesis is still unclear, however some models are proposed to explain the process. One of these models is based on the dissociation of the covalent linkage between the outer membrane and the peptidoglycan layer, being the absence of these bonds associated with the growth of the outer membrane leading to the formation of OMVs (Kulp and Kuehn, 2010; Schwechheimer and Kuehn, 2015). Another model proposes that OMVs are formed by protuberances that appear on the outer membrane due to an increased pressure in the periplasmic space caused by the accumulation of misfolded proteins and fragments of peptidoglycans (Kulp and Kuehn, 2010; Schwechheimer and Kuehn, 2015). A third model is related to an enrichment of curvature-inducing molecules, such as quinolone PQS of P. aeruginosa (Mashburn-Warren et al., 2008). Roier and coworkers proposed a novel method based on phospholipid accumulation as a result of deletion or downregulation of vacJ and/or ybr genes (Roier et al., 2016). More recently a group of genes involved in OMVs biogenesis in Salmonella enterica Serovar Typhi were identified (Nevermann et al., 2019). This group contains some genes related to envelope stability, LPS synthesis, peptidoglycan synthesis and remodeling, stress sensor and transcription regulator. Gram-negative bacteria also produce outer-inner membrane vesicles (OIMVs), that are originated from the inner membrane and were firstly observed in Shewanella vesiculosa M7T and other pathogenic bacteria (Pérez-Cruz et al., 2013; Pérez-Cruz et al., 2015). These OIMVs contain both the outer membrane and the inner membrane, as well as cytoplasmic components. Gram-positive bacteria also produce MVs; however, the mechanisms of generation and release through the cell wall are not well known (Brown et al., 2015; Liu et al., 2018). Although the internal content is similar, the MVs of gram-positive and gram-negative bacteria have different glycoconjugates (Gill et al., 2019).
Several functions are attributed to bacterial MVs, including communication with other bacteria. The distribution of antimicrobial resistance genes is an example of this interaction, being considered a type of horizontal gene transfer (Chatterjee et al., 2017; Domingues and Nielsen, 2017). The presence of DNA in MVs has been reported in several cases, and even the acquisition of resistance in sensitive bacteria has been observed after exposure to OMVs from resistant bacteria in E. coli (Kim et al., 2018). MVs can also mediate the host cell invasion process and act in competition with other pathogens, due to the presence of several virulence factors and toxins. In P. aeruginosa OMVs, multiple virulence factors that are involved with the host colonization process were identified, such as β-lactamase associated with host peptide degradation, alkaline phosphatase involved in biofilm formation, hemolytic phospholipase C, and Cif related to P. aeruginosa virulence (Bomberger et al., 2009). Toxins involved in cell death induction were found in OMVs from enterohemorrhagic Escherichia coli (EHEC) O157, such as cytolethal distending toxin V and EHEC-hemolysin (Bielaszewska et al., 2017). In addition, MVs can induce the host immune response through interaction with pathogen-associated molecular pattern (PAMP) receptors. Molecules such as LPS, peptidoglycan, lipoprotein, DNA, and RNA are recognized by host cell receptors, such as Toll-like receptors (TLRs), resulting in the induction of signaling cascades and the production of proinflammatory molecules (Pathirana and Kaparakis-Liaskos, 2016). OMVs from P. aeruginosa stimulate the production of IL-8 in A549 human lung epithelial carcinoma cells (Bauman and Kuehn, 2012).
As already mentioned, interaction between the host vesicles and the microbiome in the intestine have been described. It was observed that the MVs contribute with gut homeostasis by enhancing innate immunity, since OMVs of the microbiota are involved in the activation of NOD1 signaling pathways in intestinal epithelial cells (Cañas et al., 2018). In fact, it was demonstrated that OMVs from B. fragilis induce immunomodulatory effects and prevent colitis. Inside OMVs, capsular polysaccharide A was detected, which interacts with dendritic cells via TLR2, enhancing regulatory T cells and the production of cytokine IL-10 (Shen et al., 2012). However, it is also suggested that vesicles play a role in nutrient acquisition for the entire microbiota community, since they are enriched with hydrolytic enzymes, such as glycosidases and proteases (Elhenawy et al., 2014). The mechanism of vesicle content packaging has not yet been elucidated; however, it seems to be a regulated process. When the RNA profile of Salmonella OMVs was analyzed under different environmental conditions, it was observed that some mRNAs were enriched in OMVs when compared to intracellular fractions, reinforcing the concept that MVs packaging is not a passive process, but tightly regulated (Malabirade et al., 2018).
Bacterial MVs can transport different types of RNA molecules, including mRNA, tRNA, rRNA and ncRNA (Dauros-Singorenko et al., 2018) (Figure 1 and Table 1). Studies characterizing the MV and extracellular RNA-content of Escherichia coli K-12 substrain MG1655 (OMVs and OMVs-free) identified mainly tRNA and rRNA fragments (3S rRNA, 16S rRNA, and 5S rRNA), as well ncRNAs (Ghosal et al., 2015). In OMVs from Vibrio cholerae strain A1552, ncRNA was the most abundant class of RNA identified (Sjöström et al., 2015). In V. cholerae O395, the enrichment of rRNA and sRNA, including CsrB1, CrsB2 and CrsB3 was also observed (Langlete et al., 2019).
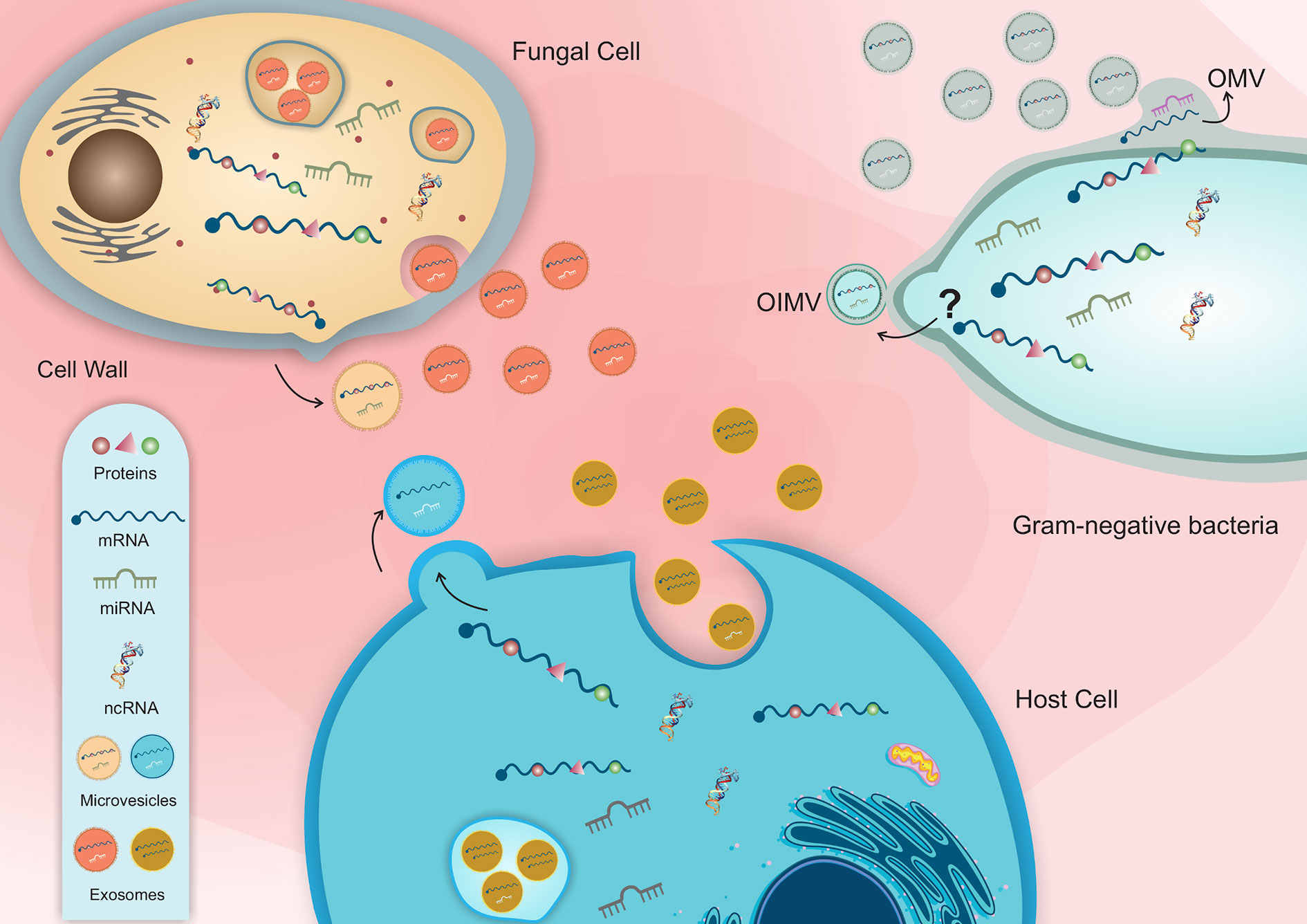
Figure 1 Schematic representation of host-pathogen EVs interaction and the distinct classes of RNA molecules in bacterial and fungal vesicles.
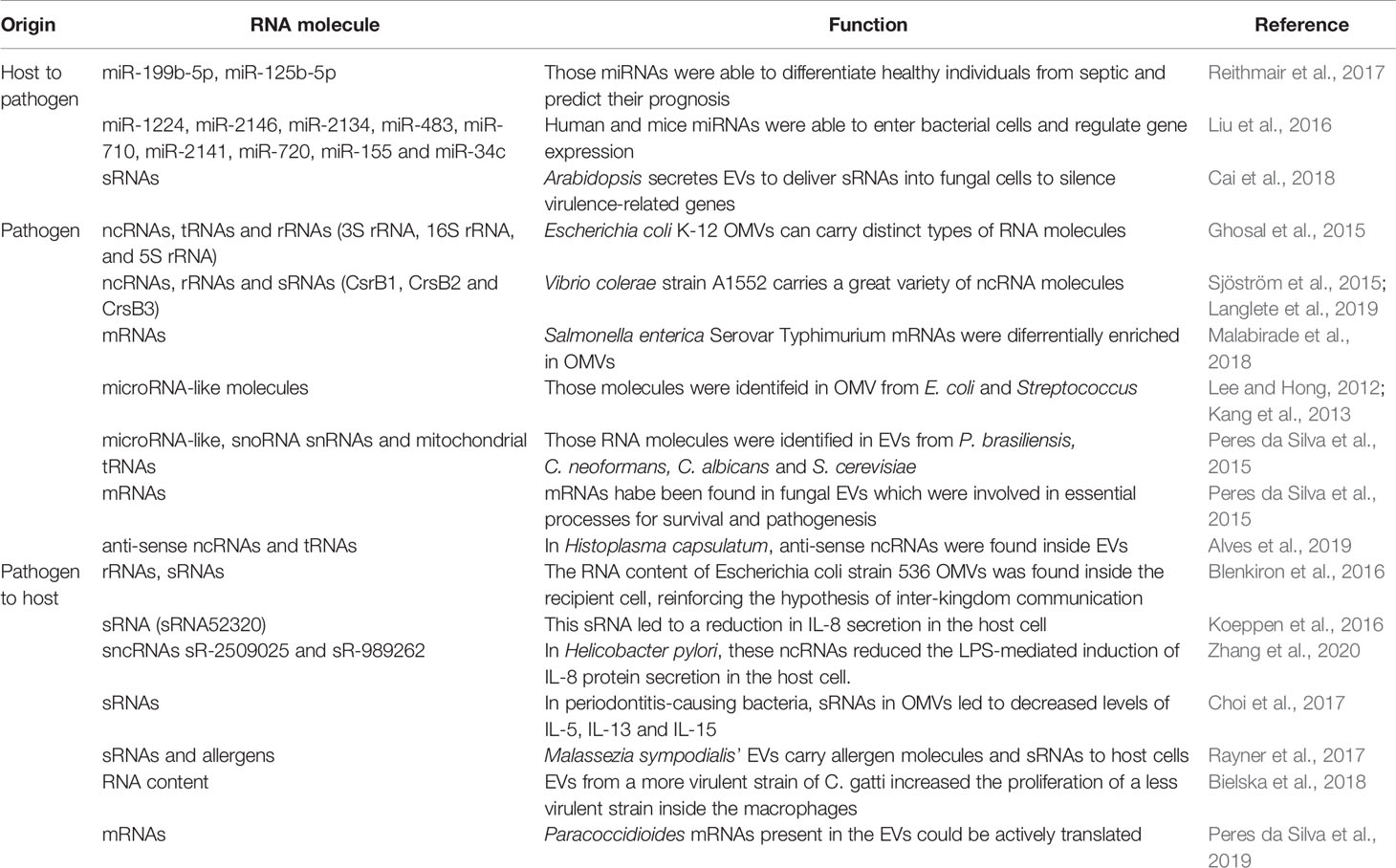
Table 1 Summary of the role of RNA molecules present in extracellular vesicles on pathogen and host communication.
Blenkiron and coworkers demonstrated that bacterial MVs RNA content is transferred to the recipient cell and directed to the cytoplasm of epithelial cells and could be also found in the nucleus, reinforcing the hypothesis of inter-kingdom communication (Blenkiron et al., 2016). In fact, MVs also show regulatory activity in the host through MV-RNA molecules that are functional in the host’s cell. RNA sequencing analysis identified “microRNA-like” in E. coli and Streptococcus mutans, speculating that these small RNA molecules may play a role in bacteria similar to miRNAs in eukaryotes (Lee and Hong, 2012; Kang et al., 2013). As mentioned, MVs can trigger immune response in the host, however, in P. aeruginosa it was identified an interaction between an sRNA molecule present in the OMV and the host cell that led to a reduction in IL-8 secretion (Koeppen et al., 2016). Using RNA-Seq, Koeppen and coworkers identified the sRNA5320 present in OMVs from P. aeruginosa. Different assays were performed in vitro and in vivo, showing that this sRNA is transferred to cells via OMVs and plays a role in modulating the immune response by decreasing IL-8 secretion. Similarly, in Helicobacter pylori, sncRNAs sR-2509025, and sR-989262 can reduce the LPS-mediated induction of IL-8 protein secretion in human gastric adenocarcinoma cells (Zhang et al., 2020).
In addition, distinct sRNAs have been identified in the vesicles of three different bacteria that cause periodontitis. The transfection of synthetic copies of these sRNAs into Jurkat T cells resulted in decreased expression levels of IL-5, IL-13, and IL-15 (Choi et al., 2017). Afterwards, for Aggregatibacter actinomycetemcomitans, which is one of the causes of periodontitis, it was observed that the sRNA of OMVs can be incorporated by the host's RISC system, contributing to the alteration in gene expression of the host (Han et al., 2019). The potential of sRNAs to act in the pathogen-host interaction has been proposed as a target to be explored for the discovery of new biomarkers for bacterial diseases (Wang and Fu, 2019).
Fungi EVs and the RNA Content
EVs have been described in several fungal species, both in yeast and in filamentous fungi such Saccharomyces cerevisiae (Zhao et al., 2019), Candida albicans (Vargas et al., 2015), Histoplasma capsulatum (Albuquerque et al., 2008), Paracoccidioides brasiliensis (Ganiko et al., 2011), Cryptococcus neoformans (Rodrigues et al., 2007), Aspergillus fumigatus (Souza et al., 2019), Sporothrix brasiliensis (Ikeda et al., 2018), Malassezia sympodialis (Rayner et al., 2017) and Alternaria infectoria (Silva et al., 2014).
As described for mammalian cells, fungal EVs are also classified according to their biogenesis, being able to release both exosomes and microvesicles (Kwon et al., 2019). The EVs, biogenesis process in fungi and how the sorting of their cargo occurs remains unknown. However, there are preliminary studies that report the relevance of EVs regulation pathways in molecular traffic across the cell wall (Bielska et al., 2018). In C. albicans EVs are key players to biofilm matrix production. ESCRT (endosomal sorting complexes required for transport) defective mutations caused reduced EV production and consequently biofilm thickness reduction and increased sensitivity to the antifungal drug (Mitchell et al., 2018). Mutations in genes that encode components of ESCRT machinery in S. cerevisiae also led to the reduction in EVs population and changes in the EVs proteomic profile (Zhao et al., 2019). The deletion of Sec6, a component of the exocyst complex involved with vesicles fusion with the plasma membrane, prevents EVs production and laccase secretion to extracellular milieu, and decreased virulence in mice (Panepinto et al., 2009). In addition, mutations in protein of the ESCRT complex, Vps27, led to MVB accumulation and decreased laccase transport to the cell wall (Park et al., 2020).
Fungal EVs have been related to several functions, such as biofilm matrix production (Mitchell et al., 2018), the delivery of virulence factors (Bielska et al., 2018; Ikeda et al., 2018; Konečná et al., 2019), cell wall remodeling (Zhao et al., 2019), host response and host–pathogen interaction (Rayner et al., 2017; Bitencourt et al., 2018; Johansson et al., 2018) (Figure 1 and Table 1). The EVs are composed of polysaccharides (Rodrigues et al., 2007), lipids (Vallejo et al., 2012), allergens (Johansson et al., 2018), pigments (Frases et al., 2009), cytosolic and membrane proteins (Gil-Bona et al., 2015), and nucleic acids such RNA molecules (Peres da Silva et al., 2015; Rayner et al., 2017; Alves et al., 2019; Peres da Silva et al., 2019).
Several studies have identified different RNA species loaded in fungal EVs that could perform different functions in recipient cells. The commensal yeast Malassezia sympodialis, which colonizes human skin and is associated with common skin disorders, can secrete vesicles ranging from 50 to 600 nm, which carry allergens related to inflammatory responses and small RNAs ranging from 16 to 22 nucleotides in length (Rayner et al., 2017). For the human pathogenic fungi P. brasiliensis, C. neoformans, C. albicans, and S. cerevisiae EVs, several miRNA-like sequences, as well as small nucleolar RNAs (snoRNAs) and nuclear RNA and mitochondrial tRNAs, were identified in high abundance (Peres da Silva et al., 2015). Messenger RNAs have also been found in fungal EVs; they are involved in essential processes, such as vesicle-mediated transport, metabolic pathways, cellular responses to stress, transcriptional regulation and cell cycle control (Peres da Silva et al., 2015). The pathogenic fungi H. capsulatum also produces EVs enriched with different ncRNA populations; the most abundant were tRNA fragments. It was also identified anti-sense ncRNAs, with 25 nt in length, that aligned with specific regions of the transcripts and could act in gene expression regulation as a silencing mechanism (Alves et al., 2019). In fungi, ncRNAs are important players in gene expression regulation in the fungal cells. For example, during the transition phase in dimorphic fungi, a process already recognized as important in the context of infection and virulence. Antisense transcription of ncRNAs have been linked to hyphae and spore formation in Ustilago maydis, and the loss of such RNAs resulted in virulence attenuation (Morrison et al., 2012).
In an elegant work performed by Bielska and coworkers, it was shown that EVs from a more virulent strain of C. gatti could increase the proliferation of less virulent fungal cells inside the macrophages, thus promoting pathogen survival instead of clearance by the host cell. The protein and RNA fractions were required for this transference process (Bielska et al., 2018).
Although it is not clear whether EV mRNAs are translated into functional peptides, the presence of functional mRNAs in fungal EVs was confirmed from two species of Paracoccidioides by in vitro translation (Peres da Silva et al., 2019). The presence of these transcripts suggests that they can be internalized by fungal or host cells and alter gene expression regulation and play a role in the host-fungal interaction. Most of the mRNAs present in the EVs are associated with virulence, like heat shock proteins Hsp 70 and Hsp 90-like, that have a role during infection of dimorphic fungi (Peres da Silva et al., 2019). It is possible to speculate that EV mRNA can be translated into the host cell, inducing gene expression alterations that could aid pathogen infection and survival.
Small RNAs can induce gene silencing by binding to argonaute proteins, directing the RNA-induced silencing complex (RISC) to target mRNAs for their repression. In host and pathogenic fungi interactions, sRNA molecules can participate in cross-kingdom communication (Weiberg et al., 2013; Chen et al., 2014; Weiberg et al., 2015; Cai et al., 2018). In plants, the EVs can act as effectors that suppress fungal pathogens (Cai et al., 2018). Phytophogenic (Weiberg et al., 2013; Chen et al., 2014; Wang et al., 2016) and entomopathogenic fungi (Cui et al., 2019) interfere in plant and insect immunity, respectively. They play a role in silencing host immunity genes by hijacking the host's RNAi machinery to facilitate infection (Weiberg et al., 2013; Wang et al., 2016). The entomopathogenic fungus Beauveria bassiana exports miRNA-like molecules loaded in vesicles to the host's mosquito Anopheles stephensi, attenuating host immunity, and facilitating infection (Cui et al., 2019).
Conversely, the host can also suppress fungal pathogenesis. The extracellular vesicles of plants play an essential role in sRNA trafficking between Arabidopsis and the pathogen Botrytis cinerea. Arabidopsis secretes EVs to deliver sRNAs into fungal cells to silence virulence-related genes (Cai et al., 2018).
Concluding Remarks
In summary, this unconventional pathway of communication is gaining more attention because it is involved in all aspects of cell life regarding its important, if not essential, role in organism homeostasis. Regarding infection, it was shown that EVs of both hosts and pathogens play a role in either promoting or fighting it where the EVs influence antigen presentation, immunological stimulation and suppression as well as transmission of virulence factors. Additionally, one of the most widely studied aspects of EVs is their RNA cargos, mainly because of their capacity to regulate gene expression in the recipient cell, the microRNA pathway, leading to gene silencing that can favor the pathogen, and also the mRNA in the EVs that can be translated and influence the cell metabolism. In addition, some physiological aspects regarding pathogenesis and host response were studied and are helping to understand how the EVs can be used for diagnostic and therapeutical purposes, the RNA present in EVs derived from bacteria and fungi and their role in the host are recent and still need to be further addressed for most of the work has been descriptive. The EV biogenesis and how specific molecules are sorted and directed to them are questions to be answered. Nevertheless, the promising results obtained so far are paving a new path for the study of RNA present in the vesicles and their important role in cell communication and gene expression regulation highlighting the potential of EVs and their role in the host during infection.
Author Contributions
IM, RA, AL, HF, and LA discussed, wrote, and approved the manuscript in its current form. All authors contributed to the article and approved the submitted version.
Funding
This work received financial support from Inova Fiocruz/Fundação Oswaldo Cruz [Grant number VPPCB-07-FIO-18-2-52] and CNPq [Grant number 442317/2019-0]. LRA is a research fellow awardee from CNPq.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Wagner Nagib Birbeire for the schematic Figure 1.
References
Albuquerque P. C., Nakayasu E. S., Rodrigues M. L., Frases S., Casadevall A., Zancope-Oliveira R. M., et al. (2008). Vesicular transport in Histoplasma capsulatum: An effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell. Microbiol. 10, 1695–1710. doi: 10.1111/j.1462-5822.2008.01160.x
Alves L. R., Peres da Silva R., Sanchez D. A., Zamith-Miranda D., Rodrigues M. L., Goldenberg S., et al. (2019). Extracellular Vesicle-Mediated RNA Release in Histoplasma capsulatum. mSphere 4, 1–17. doi: 10.1128/msphere.00176-19
Baglio S. R., Rooijers K., Koppers-Lalic D., Verweij F. J., Pérez Lanzón M., Zini N., et al. (2015). Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 6, 127. doi: 10.1186/s13287-015-0116-z
Bauman S. J., Kuehn M. J. (2012). Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes Infect. 8, 2400–2408. doi: 10.1016/j.micinf.2006.05.001
Bielaszewska M., Rüter C., Bauwens A., Greune L., Jarosch K. A., Steil D., et al. (2017). Host cell interactions of outer membrane vesicle-associated virulence factors of enterohemorrhagic Escherichia coli O157: Intracellular delivery, trafficking and mechanisms of cell injury. PloS Pathog. 13 (2), e1006159. doi: 10.1371/journal.ppat.1006159
Bielska E., Sisquella M. A., Aldeieg M., Birch C., O’Donoghue E. J., May R. C. (2018). Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii. Nat. Commun. 9, 1–9. doi: 10.1038/s41467-018-03991-6
Bitencourt T. A., Rezende C. P., Quaresemin N. R., Moreno P., Hatanaka O., Rossi A., et al. (2018). Extracellular vesicles from the dermatophyte Trichophyton interdigitale modulate macrophage and keratinocyte functions. Front. Immunol. 9, 2343. doi: 10.3389/fimmu.2018.02343
Blenkiron C., Simonov D., Muthukaruppan A., Tsai P., Dauros P., Green S., et al. (2016). Uropathogenic Escherichia coli releases extracellular vesicles that are associated with RNA. PloS One 11, 1–16. doi: 10.1371/journal.pone.0160440
Bomberger J. M., MacEachran D. P., Coutermarsh B. A., Ye S., O’Toole G. A., Stanton B. A. (2009). Long-distance delivery of bacterial virulence factors by Pseudomonas aeruginosa outer membrane vesicles. PloS Pathog. 5, e1000382. doi: 10.1371/journal.ppat.1000382
Brown L., Wolf J. M., Prados-Rosales R., Casadevall A. (2015). Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 13, 620–630. doi: 10.1038/nrmicro3480
Cai Q., Qiao L., Wang M., He B., Lin F. M., Palmquist J., et al. (2018). Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Sci. 360, 1–8. doi: 10.1126/science.aar4142
Cañas M. A., Fábrega M. J., Giménez R., Badia J., Baldomà L. (2018). Outer membrane vesicles from probiotic and commensal Escherichia coli activate NOD1-mediated immune responses in intestinal epithelial cells. Front. Microbiol. 9:498. doi: 10.3389/fmicb.2018.00498
Chatterjee S., Mondal A., Mitra S., Basu S. (2017). Acinetobacter baumannii transfers the bla NDM-1 gene via outer membrane vesicles. J. Antimicrob. Chemother. 72, 2201–2207. doi: 10.1093/jac/dkx131
Chen R., Jiang N., Jiang Q., Sun X., Wang Y., Zhang H., et al. (2014). Exploring microRNA-like small RNAs in the filamentous fungus Fusarium oxysporum. PloS One 9, e104956. doi: 10.1371/journal.pone.0104956
Choi J. W., Kim S. C., Hong S. H., Lee H. J. (2017). Secretable Small RNAs via Outer Membrane Vesicles in Periodontal Pathogens. J. Dent. Res. 96, 458–466. doi: 10.1177/0022034516685071
Colombo M., Moita C., Van Niel G., Kowal J., Vigneron J., Benaroch P., et al. (2013). Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 126, 5553–5565. doi: 10.1242/jcs.128868
Cui C., Wang Y., Liu J., Zhao J., Sun P., Wang S. (2019). A fungal pathogen deploys a small silencing RNA that attenuates mosquito immunity and facilitates infection. Nat. Commun. 10, 1–10. doi: 10.1038/s41467-019-12323-1
Dagnelie M. A., Corvec S., Khammari A., Dréno B. (2019). Bacterial extracellular vesicles: A new way to decipher host-microbiota communications in inflammatory dermatoses. Exp. Dermatol. 29, 22–28. doi: 10.1111/exd.14050
Dauros-Singorenko P., Blenkiron C., Phillips A., Swift S. (2018). The functional RNA cargo of bacterial membrane vesicles. FEMS Microbiol. Lett. 365, 1–9. doi: 10.1093/femsle/fny023
Domingues S., Nielsen K. M. (2017). Membrane vesicles and horizontal gene transfer in prokaryotes. Curr. Opin. Microbiol. 38, 16–21. doi: 10.1016/j.mib.2017.03.012
Dong G., Filho A. L., Olivier M. (2019). Modulation of Host-Pathogen Communication by Extracellular Vesicles (EVs) of the Protozoan Parasite Leishmania. Front. Cell. Infect. Microbiol. 9:100. doi: 10.3389/fcimb.2019.00100
Eichenberger R. M., Stephanie R., Jones L., Buitrago G., Polster R., de Oca M. M., et al. (2018). Hookworm secreted extracellular vesicles interact with host cells and prevent inducible colitis in mice. Front. Immunol. 9, 1–14. doi: 10.3389/fimmu.2018.00850
Elhenawy W., Debelyy M. O., Feldman M. F. (2014). Preferential packing of acidic glycosidases and proteases into Bacteroides outer membrane vesicles. MBio 5, 1–12. doi: 10.1128/mBio.00909-14
Frases S., Eisenman H. C., Rodrigues M. L., Casadevall A., Nicola A. M. (2009). Vesicle-associated melanization in Cryptococcus neoformans. Microbiology 155, 3860–3867. doi: 10.1099/mic.0.032854-0
Ganiko L., Silva L. S., Puccia R., Freymüller-Haapalainen E., Matsuo A. L., Almeida I. C., et al. (2011). The Pathogenic Fungus Paracoccidioides brasiliensis Exports Extracellular Vesicles Containing Highly Immunogenic α-Galactosyl Epitopes. Eukaryot. Cell 10, 343–351. doi: 10.1128/ec.00227-10
Ghosal A., Upadhyaya B. B., Fritz J. V., Heintz-Buschart A., Desai M. S., Yusuf D., et al. (2015). The extracellular RNA complement of Escherichia coli. Microbiologyopen 4, 252–266. doi: 10.1002/mbo3.235
Gibbings D. J., Ciaudo C., Erhardt M., Voinnet O. (2009). Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity. Nat. Cell Biol. 11, 1143–1149. doi: 10.1038/ncb1929
Gil-Bona A., Llama-Palacios A., Parra C. M., Vivanco F., Nombela C., Monteoliva L., et al. (2015). Proteomics unravels extracellular vesicles as carriers of classical cytoplasmic proteins in Candida albicans. J. Proteome Res. 14, 142–153. doi: 10.1021/pr5007944
Gill S., Catchpole R., Forterre P. (2019). Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 43, 273–303. doi: 10.1093/femsre/fuy042
Han E.-C., Choi S.-Y., Lee Y., Park J.-W., Hong S.-H., Lee H.-J. (2019). Extracellular RNAs in periodontopathogenic outer membrane vesicles promote TNF-α production in human macrophages and cross the blood–brain barrier in mice. FASEB J. 33, 13412–13422. doi: 10.1096/fj.201901575r. fj.201901575R.
Heijnen H. F. G., Schiel A. E., Fijnheer R., Geuze H. J., Sixma J. J. (2015). Activated Platelets Release Two Types of Membrane Vesicles: Microvesicles by Surface Shedding and Exosomes Derived From Exocytosis of Multivesicular Bodies and Granules. Blood 94, 3791–3799. doi: 10.1182/blood.V94.11.3791
Hessvik N. P., Llorente A. (2018). Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 75, 193–208. doi: 10.1007/s00018-017-2595-9
Hurwitz S. N., Conlon M. M., Rider M. A., Brownstein N. C., Meckes D. G. (2016). Nanoparticle analysis sheds budding insights into genetic drivers of extracellular vesicle biogenesis. J. Extracell. Vesicles 5:31295. doi: 10.3402/jev.v5.31295
Hurwitz S. N., Nkosi D., Conlon M. M., York S. B., Liu X., Tremblay D. C., et al. (2017). CD63 Regulates Epstein-Barr Virus LMP1 Exosomal Packaging, Enhancement of Vesicle Production, and Noncanonical NF-κB Signaling. J. Virol. 91, e02251-16. doi: 10.1128/jvi.02251-16
Ikeda M. A. K., de Almeida J. R. F., Jannuzzi G. P., Cronemberger-Andrade A., Torrecilhas A. C. T., Moretti N. S., et al. (2018). Extracellular Vesicles From Sporothrix brasiliensis Are an Important Virulence Factor That Induce an Increase in Fungal Burden in Experimental Sporotrichosis. Front. Microbiol. 9:2286:2286. doi: 10.3389/fmicb.2018.02286
Im H., Lee H., Castro C. M., Weissleder R., Shao H., Breakefield X. (2018). New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 118 (4), 1917–1950. doi: 10.1021/acs.chemrev.7b00534
Johansson H. J., Vallhov H., Holm T., Gehrmann U., Andersson A., Johansson C., et al. (2018). Extracellular nanovesicles released from the commensal yeast Malassezia sympodialis are enriched in allergens and interact with cells in human skin. Nat. Sci. Rep. 8, 1–11. doi: 10.1038/s41598-018-27451-9
Joncas F.-H., Lucien F., Rouleau M., Morin F., Leong H. S., Pouliot F., et al. (2019). Plasma extracellular vesicles as phenotypic biomarkers in prostate cancer patients. Prostate 79, 1–10. doi: 10.1002/pros.23901
Kalluri R., LeBleu V. S. (2020). The biology, function, and biomedical applications of exosomes. Science 367, eaau6977. doi: 10.1126/science.aau6977
Kang S. M., Choi J. W., Lee Y., Hong S. H., Lee H. J. (2013). Identification of microRNA-Size, Small RNAs in Escherichia coli. Curr. Microbiol. 67, 609–613. doi: 10.1007/s00284-013-0411-9
Kim S. W., Park S., Im S. P., Lee J. S., Jung J. W., Gong T. W., et al. (2018). Outer membrane vesicles from β-lactam-resistant Escherichia coli enable the survival of β-lactam-susceptible E. coli in the presence of β-lactam antibiotics. Sci. Rep. 8, 5402. doi: 10.1038/s41598-018-23656-0
Koeppen K., Hampton T. H., Jarek M., Scharfe M., Gerber S. A., Mielcarz D. W., et al. (2016). A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PloS Pathog. 12, e1005672. doi: 10.1371/journal.ppat.1005672
Koliha N., Wiencek Y., Heider U., Jü Ngst C., Kladt N., Krauthä User S., et al. (2016). A novel multiplex bead-based platform highlights the diversity of extracellular vesicles Cologne Excellence Cluster on Cellular Stress Responses in Aging-Associated Diseases. J. Extracell. Vesicles 1, 1–15. doi: 10.3402/jev.v5.29975
Konečná K., Klimentová J., Benada O., Němečková I., Janďourek O., Jílek P., et al. (2019). A comparative analysis of protein virulence factors released via extracellular vesicles in two Candida albicans strains cultivated in a nutrient-limited medium. Microb. Pathog. 136, 103666. doi: 10.1016/j.micpath.2019.103666
Kulp A., Kuehn M. J. (2010). Functions and Biogenesis of OMVs. Annu. Rev. Microbiol. 64, 163–184. doi: 10.1146/annurev.micro.091208.073413.Biological
Kwon S., Tosserant C., Tulinski M., Weiberg A., Feldbrügge M. (2019). Inside-out: from endosomes to extracellular vesicles in fungal RNA transport. Fungal Biol. Rev. 34, 89–99. doi: 10.1016/j.fbr.2020.01.001
Langlete P., Krabberød A. K., Winther-larsen H. C. (2019). Vesicles From Vibrio cholerae Contain AT-Rich DNA and Shorter mRNAs That Do Not Correlate With Their Protein Products. Front. Microbiol. 10:2708. doi: 10.3389/fmicb.2019.02708
Lee H. J., Hong S. H. (2012). Analysis of microRNA-size, small RNAs in Streptococcus mutans by deep sequencing. FEMS Microbiol. Lett. 326, 131–136. doi: 10.1111/j.1574-6968.2011.02441.x
Lee H. J. (2019). Microbe-host communication by small RNAs in extracellular vesicles: Vehicles for transkingdom RNA transportation. Int. J. Mol. Sci. 20, 1487. doi: 10.3390/ijms20061487
Liu S., Da Cunha A. P., Rezende R. M., Cialic R., Wei Z., Bry L., et al. (2016). The Host Shapes the Gut Microbiota via Fecal MicroRNA. Cell Host Microbe 19, 32–43. doi: 10.1016/j.chom.2015.12.005
Liu Y., Defourny K. A. Y., Smid E. J., Abee T. (2018). Gram-positive bacterial extracellular vesicles and their impact on health and disease. Front. Microbiol. 9:1502. doi: 10.3389/fmicb.2018.01502
Malabirade A., Habier J., Heintz-Buschart A., May P., Godet J., Halder R., et al. (2018). The RNA complement of outer membrane vesicles from Salmonella enterica Serovar Typhimurium under distinct culture conditions. Front. Microbiol. 9:2015. doi: 10.3389/fmicb.2018.02015
Mardahl M., Borup A., Nejsum P. (2019). A new level of complexity in parasite-host interaction: The role of extracellular vesicles. Adv. Parasitol. 1st ed (London, United Kingdom: Elsevier Ltd). doi: 10.1016/bs.apar.2019.02.003
Mashburn L. M., Whiteley M. (2005). Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Membr. vesicles traffic signals Facil. Gr. Act. Prokaryote 437, 422–425. doi: 10.1038/nature03925
Mashburn-Warren L., Howe J., Garidel P., Richter W., Steiniger F., Roessle M., et al. (2008). Interaction of quorum signals with outer membrane lipids: insights into prokaryotic membrane vesicle formation. Mol. Microbiol. 69, 491–502. doi: 10.1111/j.1365-2958.2008.06302.x
Mateescu B., Kowal E. J. K., van Balkom B. W. M., Bartel S., Bhattacharyya S. N., Buzás E. II, et al. (2017). Obstacles and opportunities in the functional analysis of extracellular vesicle RNA - An ISEV position paper. J. Extracell. Vesicles 6, 1286095. doi: 10.1080/20013078.2017.1286095
McKenzie A. J., Hoshino D., Hong N. H., Cha D. J., Franklin J. L., Coffey R. J., et al. (2016). KRAS-MEK Signaling Controls Ago2 Sorting into Exosomes. Cell Rep. 15, 978–987. doi: 10.1016/j.celrep.2016.03.085
McMahon H. T., Boucrot E. (2015). Membrane curvature at a glance. J. Cell Sci. 128, 1065–1070. doi: 10.1242/jcs.114454
Mitchell K. F., Azadi P., Jaromin A., Sanchez H., Dominguez E., Andes D. R., et al. (2018). Candida albicans biofilm–induced vesicles confer drug resistance through matrix biogenesis. PloS Biol. 16, e2006872. doi: 10.1371/journal.pbio.2006872
Mittelbrunn M., Gutiérrez-Vázquez C., Villarroya-Beltri C., González S., Sánchez-Cabo F., González M. Á., et al. (2011). Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2, 1–10. doi: 10.1038/ncomms1285
Morales D. K., Hogan D. A. (2010). Candida albicans interactions with bacteria in the context of human health and disease. PloS Pathog. 6, 1–4. doi: 10.1371/journal.ppat.1000886
Morrison E. N., Donaldson M. E., Saville B. J. (2012). Identification and analysis of genes expressed in the Ustilago maydis dikaryon: Uncovering a novel class of pathogenesis genes. Can. J. Plant Pathol. 34, 417–435. doi: 10.1080/07060661.2012.697077
Nevermann J., Silva A., Otero C., Oyarzún D. P., Barrera B., Gil F., et al. (2019). Identification of genes involved in biogenesis of Outer Membrane Vesicles (OMVs) in Salmonella enterica Serovar Typhi. Front. Microbiol. 10:104. doi: 10.3389/fmicb.2019.00104
O’Brien K., Breyne K., Ughetto S., Laurent L. C., Breakefield X. O. (2020). RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol 2, 585–606. doi: 10.1038/s41580-020-0251-y
Panepinto J., Komperda K., Frases S., Park Y. D., Djordjevic J. T., Casadevall A., et al. (2009). Sec6-dependent sorting of fungal extracellular exosomes and laccase of Cryptococcus neoformans. Mol. Microbiol. 71 (5), 1165–1176. doi: 10.1111/j.1365-2958.2008.06588.x
Park K. S., Lee J., Lee C., Park H. T., Kim J. W., Kim O. Y., et al. (2018). Sepsis-like systemic inflammation induced by nano-sized extracellular vesicles from feces. Front. Microbiol. 9:1735. doi: 10.3389/fmicb.2018.01735
Park Y. D., Chen S. H., Camacho E., Casadevall A., Williamson P. R. (2020). Role of the ESCRT Pathway in Laccase Trafficking and Virulence of Cryptococcus neoformans. Infect. Immun. 88 (7), e00954-19. doi: 10.1128/IAI.00954-19
Pathirana R. D., Kaparakis-Liaskos M. (2016). Bacterial membrane vesicles Biogenesis, immune regulation and pathogenesis. Cell. Microbiol. 18, 1518–1524. doi: 10.1111/cmi.12658
Peres da Silva R., Puccia R., Rodrigues M. L., Oliveira D. L., Joffe L. S., César G. V., et al. (2015). Extracellular vesicle-mediated export of fungal RNA. Sci. Rep. 5:7763. doi: 10.1038/srep07763
Peres da Silva R., Longo L. G. V., Cunha J. P. C., da, Sobreira T. J. P., Rodrigues M. L., Faoro H., et al. (2019). Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii. Cells 8:765. doi: 10.3390/cells8070765
Pérez-Cruz C., Carrión O., Delgado L., Martinez G., López-Iglesias C., Mercade E. (2013). New type of outer membrane vesicle produced by the gram-negative bacterium Shewanella vesiculosa M7T: Implications for DNA content. Appl. Environ. Microbiol. 79, 1874–1881. doi: 10.1128/AEM.03657-12
Pérez-Cruz C., Delgado L., López-Iglesias C., Mercade E. (2015). Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PloS One 10, 1–18. doi: 10.1371/journal.pone.0116896
Raeven P., Zipperle J., Drechsler S. (2018). Extracellular vesicles as markers and mediators in sepsis. Theranostics 8, 3348–3365. doi: 10.7150/thno.23453
Ragusa M., Barbagallo C., Cirnigliaro M., Battaglia R., Brex D., Caponnetto A., et al. (2017). Asymmetric RNA distribution among cells and their secreted exosomes: Biomedical meaning and considerations on diagnostic applications. Front. Mol. Biosci. 4:66. doi: 10.3389/fmolb.2017.00066
Raposo G., Stahl P. D. (2019). Extracellular vesicles: a new communication paradigm? Nat. Rev. Mol. Cell Biol. 20, 509–510. doi: 10.1038/s41580-019-0158-7
Raposo G., Stoorvogel W. (2013). Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 200, 373–383. doi: 10.1083/jcb.201211138
Rayner S., Bruhn S., Vallhov H., Andersson A., Billmyre R. B., Scheynius A. (2017). Identification of small RNAs in extracellular vesicles from the commensal yeast Malassezia sympodialis. Sci. Rep. 7, 39742. doi: 10.1038/srep39742
Reithmair M., Buschmann D., Märte M., Kirchner B., Hagl D., Kaufmann I., et al. (2017). Cellular and extracellular miRNAs are blood-compartment-specific diagnostic targets in sepsis. J. Cell. Mol. Med. 21, 2403–2411. doi: 10.1111/jcmm.13162
Roderburg C., Luedde M., Vargas Cardenas D., Vucur M., Scholten D., Frey N., et al. (2013). Circulating microRNA-150 serum levels predict survival in patients with critical illness and sepsis. PloS One 8, e54612. doi: 10.1371/journal.pone.0054612
Rodrigues M. L., Nimrichter L., Oliveira D. L., Frases S., Miranda K., Zaragoza O., et al. (2007). Vesicular Polysaccharide Export in Cryptococcus neoformans Is a Eukaryotic Solution to the Problem of Fungal Trans-Cell Wall Transport. Eukaryot. Cell 6, 48–59. doi: 10.1128/EC.00318-06
Roier S., Zingl F. G., Cakar F., Durakovic S., Kohl P., Eichmann T. O., et al. (2016). A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 7, 257–259. doi: 10.1038/ncomms10515
Schwechheimer C., Kuehn M. J. (2015). Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 13, 605–619. doi: 10.1038/nrmicro3525
Shah R., Patel T., Freedman J. E. (2018). Circulating extracellular vesicles in human disease. N. Engl. J. Med. 379, 958–966. doi: 10.1056/NEJMra1704286
Shen Y., Letizia M., Torchia G., Lawson G. W., Karp C. L., Ashwell J. D., et al. (2012). Outer Membrane Vesicles of a Human Commensal Mediate Immune Regulation and Disease Protection. Cell Host Microbe 12, 509–520. doi: 10.1016/j.chom.2012.08.004.Outer
Shopova I. A., Belyaev I., Dasari P., Jahreis S., Stroe M. C., Cseresnyés Z., et al. (2020). Human neutrophils produce antifungal extracellular vesicles against Aspergillus fumigatus. bioRxiv 11, e00596-20. doi: 10.1128/mBio.00596-20
Silva B. M. A., Prados-Rosales R., Espadas-Moreno J., Wolf J. M., Luque-Garcia J. L., Gonçalves T., et al. (2014). Characterization of Alternaria infectoria extracellular vesicles. Med. Mycol 52, 202–210. doi: 10.1093/mmy/myt003
Simon C., Greening D. W., Bolumar D., Balaguer N., Salamonsen L. A., Vilella F. (2018). Extracellular vesicles in human reproduction in health and disease. Endocr. Rev. 39, 292–332. doi: 10.1210/er.2017-00229
Sjöström A. E., Sandblad L., Uhlin B. E., Wai S. N. (2015). Membrane vesicle-mediated release of bacterial RNA. Sci. Rep. 5, 1–10. doi: 10.1038/srep15329
Souza J. A. M., Baltazar L., de M., Carregal V. M., Gouveia-Eufrasio L., de Oliveira A. G., et al. (2019). Characterization of Aspergillus fumigatus Extracellular Vesicles and Their Effects on Macrophages and Neutrophils Functions. Front. Microbiol. 10, 1–13. doi: 10.3389/fmicb.2019.02008
Tatischeff I. (2019). Dictyostelium: A Model for Studying the Extracellular Vesicle Messengers Involved in Human Health and Disease. Cells 8:225. doi: 10.3390/cells8030225
Teng X., Chen L., Chen W., Yang J., Yang Z., Shen Z. (2015). Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation. Cell. Physiol. Biochem. 37, 2415–2424. doi: 10.1159/000438594
Théry C., Ostrowski M., Segura E. (2009). Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 9, 581–593. doi: 10.1038/nri2567
Théry C., Witwer K. W., Aikawa E., Alcaraz M. J., Anderson J. D., Andriantsitohaina R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 7, 3078. doi: 10.1080/20013078.2018.1535750
Toyofuku M., Nomura N., Eberl L. (2019). Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 17, 13–24. doi: 10.1038/s41579-018-0112-2
Turchinovich A., Drapkina O., Tonevitsky A. (2019). Transcriptome of extracellular vesicles: State-of-the-art. Front. Immunol. 10:202. doi: 10.3389/fimmu.2019.00202
Vallejo M. C., Nakayasu E. S., Longo L. V. G., Ganiko L., Lopes F. G., Matsuo A. L., et al. (2012). Lipidomic analysis of extracellular vesicles from the pathogenic phase of Paracoccidioides brasiliensis. PloS One 7, 1–10. doi: 10.1371/journal.pone.0039463
Van Niel G., D’Angelo G., Raposo G. (2018). Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 19, 213–228. doi: 10.1038/nrm.2017.125
Vargas G., Rocha J. D. B., Oliveira D. L., Albuquerque P. C., Frases S., Santos S. S., et al. (2015). Compositional and immunobiological analyses of extracellular vesicles released by Candida albicans. Cell. Microbiol. 17, 389–407. doi: 10.1111/cmi.12374
Wang Y. F., Fu J. (2019). Secretory and circulating bacterial small RNAs: a mini-review of the literature. ExRNA 1, 1–5. doi: 10.1186/s41544-019-0015-z
Wang M., Weiberg A., Lin F.-M., Thomma B. P. H. J., Huang H.-D., Jin H. (2016). Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2, 16151. doi: 10.1038/nplants.2016.151
Weiberg A., Wang M., Lin F.-M., Zhao H., Zhang Z., Kaloshian I., et al. (2013). Fungal Small RNAs Suppress Plant Immunity by Hijacking Host. Science 342, 118–123. doi: 10.1126/science.1239705.Fungal
Weiberg A., Bellinger M., Jin H. (2015). Conversations between kingdoms: small RNAs. Curr. Opin. Biotechnol. 32, 207–215. doi: 10.1016/j.copbio.2014.12.025
Yáñez-Mó M., Siljander P. R. M., Andreu Z., Zavec A. B., Borràs F. E., Buzas E. II, et al. (2015). Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 4, 1–60. doi: 10.3402/jev.v4.27066
Zhang H., Zhang Y., Song Z., Li R., Ruan H., Liu Q., et al. (2020). sncRNAs packaged by Helicobacter pylori outer membrane vesicles attenuate IL-8 secretion in human cells. Int. J. Med. Microbiol. 310:151356. doi: 10.1016/j.ijmm.2019.151356
Keywords: extracellular vesicles, host–pathogen interaction, bacteria, fungi, infection
Citation: Munhoz da Rocha IF, Amatuzzi RF, Lucena ACR, Faoro H and Alves LR (2020) Cross-Kingdom Extracellular Vesicles EV-RNA Communication as a Mechanism for Host–Pathogen Interaction. Front. Cell. Infect. Microbiol. 10:593160. doi: 10.3389/fcimb.2020.593160
Received: 09 August 2020; Accepted: 13 October 2020;
Published: 18 November 2020.
Edited by:
Ana Claudia Torrecilhas, Federal University of São Paulo, BrazilReviewed by:
Arianna Tavanti, University of Pisa, ItalyAndre Cronemberger Andrade, Universidade Federal de São Paulo, Brazil
Copyright © 2020 Munhoz da Rocha, Amatuzzi, Lucena, Faoro and Alves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lysangela Ronalte Alves, lysangela.alves@fiocruz.br, lys.alves@gmail.com