Solanum Fruits: Phytochemicals, Bioaccessibility and Bioavailability, and Their Relationship With Their Health-Promoting Effects
 Cristina Alicia Elizalde-Romero1 Luis Aurelio Montoya-Inzunza1
Cristina Alicia Elizalde-Romero1 Luis Aurelio Montoya-Inzunza1  Laura Aracely Contreras-Angulo1
Laura Aracely Contreras-Angulo1  J. Basilio Heredia1
J. Basilio Heredia1  Erick Paul Gutiérrez-Grijalva2*
Erick Paul Gutiérrez-Grijalva2*- 1Centro de Investigación en Alimentación y Desarrollo, Culiacán, Mexico
- 2Cátedras CONACYT-Centro de Investigación en Alimentación y Desarrollo, Culiacán, Mexico
The Solanum genus is the largest in the Solanaceae family containing around 2,000 species. There is a great number of edibles obtained from this genus, and globally, the most common are tomato (S. lycopersicum), potato (S. tuberosum), and eggplant (S. melongena). Other fruits are common in specific regions and countries, for instance, S. nigrum, S. torvum, S. betaceum, and S. stramonifolium. Various reports have shown that flavonoids, phenolic acids, alkaloids, saponins, and other molecules can be found in these plants. These molecules are associated with various health-promoting properties against many non-communicable diseases, the main causes of death globally. Nonetheless, the transformations of the structure of antioxidants caused by cooking methods and gastrointestinal digestion impact their potential benefits and must be considered. This review provides information about antioxidant compounds, their bioaccessibility and bioavailability, and their health-promoting effects. Bioaccessibility and bioavailability studies must be considered when evaluating the bioactive properties of health-promoting molecules like those from the Solanum genus.
Introduction
The numerous species of the Solanum genus are distributed mainly in tropical and subtropical areas around the globe; these are used in folk medicine or food crops. The positive effects on the human health of these plants are linked to their content of phenols, alkaloids, saponins, terpenes, flavonoids, coumarins, and carotenoids (1, 2). Some have been reported with anticancer, antioxidant, antidepressant, antihypertensive, anti-inflammatory, hypolipidemic, hypoglycemic, hepatoprotective, anti-obesogenic, and antidiabetic properties (1–4).
In addition, there are reports regarding the bioaccessibility and bioavailability of these molecules. Bioactive compounds are subjected to modifications during processing and gastrointestinal digestion. Moreover, those that permeate the intestinal barrier are metabolized, and then most are distributed for excretion. Thus, the bioavailability of bioactive molecules is often low (5). The low bioavailability of bioactive compounds can hinder their potential bioactive effects on human health. Thus, they are important factors to consider during in vitro and in vivo evaluations. The present document focuses on consumables matrices and those with potential bioactive effects. A summary of the compounds identified and isolated from Solanum species and their potential bioactive properties can be found in Table 1.
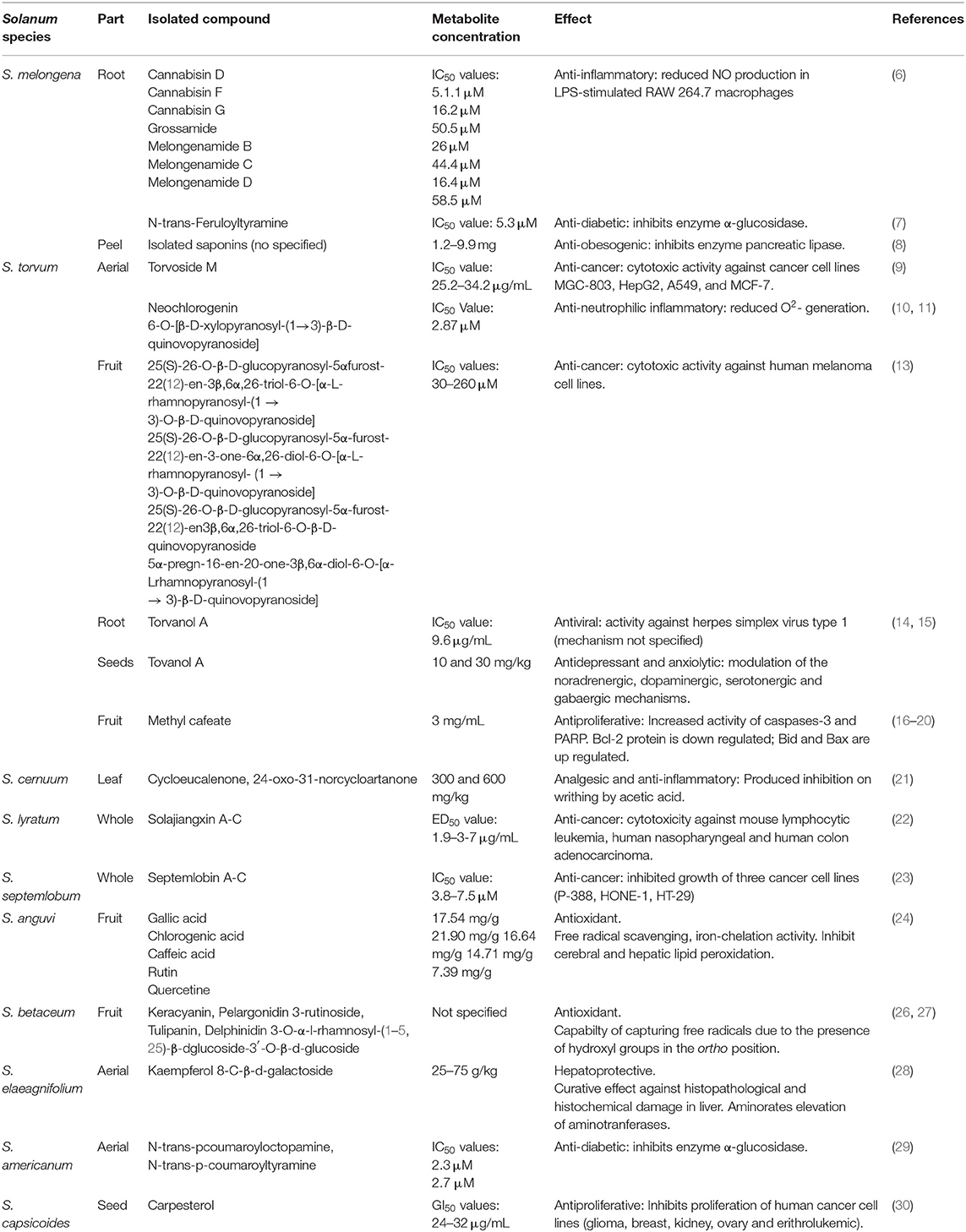
Table 1. Phytochemicals of Solanum species and their biological effects.
Phenolic Compounds
Phenolics have at least one aromatic ring with one hydroxyl group in their structure. Phenolics can be classified into flavonoids and non-flavonoids (25). One of the most common species worldwide is S. lycopersicum, which is reported with large concentrations of phenolic like chlorogenic acid, resveratrol, quercetin, and myricetin (31–33). Moreover, S. tuberosum, has been reported with high concentrations of phenolics in the peels, mainly phenolic acids, especially chlorogenic acid (34). Furthermore, S. melongena, where hydroxycinnamic acid derivatives are reported as major phenolics (35). Also, S. nigrum had as major compounds myricetin, 3,4-dicaffeoylquinic acid, 3-caffeoylquinic acid, 5-caffeoylquinic acid, and 4,5-dicaffeoylquinic (36). S. betaceum had chlorogenic acid and 3-O-caffeoylquinic acid as major compounds (37). The S. stramonifolium plant reports high concentrations of phenolics, and the root extract presents anticancer effects (38). Less common species such as S. scabrum and S. burbankii have the presence of petunidin, delphinidin, and malvidin (39). Some health-related effects of the main phenolics present in this genus are antioxidant, anti-inflammatory, antidiabetic, cardioprotective, and anti-obesity. Phenolic compounds from diverse sources are highly unstable during the gastrointestinal digestion and have low bioavailability (3, 40).
Alkaloids
Alkaloids are compounds with at least one nitrogen atom in their structure. They can be classified according to their origin, chemical characteristics, or depending on the biosynthetic pathway from which they were derived. However, we can mostly find them classified as true alkaloids, protoalkaloids, and pseudoalkaloids (41). The genus Solanum is within the Solanaceae family and is recognized for its alkaloid content and anti-proliferative effects. Other health-promoting effects attributed to alkaloids are antioxidant, antidiabetic; they are considered potential drugs for treating neurodegenerative disorders such as Huntington disease, Parkinson's disease, epilepsy, schizophrenia, and Alzheimer's disease (2, 42, 43). In S. tuberosum, dehydrochaconine, chaconine isomers, α-chaconine, solanidadienol chacotriose, solanidadiene solatriose, solanidenol chacotriose, α-solanine, leptine II, α- solanine, α-chaconine, dehydrocommersonine, commersonine, demissine, sisunine, are distributed in different parts of the plant (12, 44–48). Likewise, in S. lycopersicum, the compounds α-tomatine and dehydrotomatine are synthesized mainly in the fruit and can also be found in aglycone form as tomatidienol and tomatidine. Esculeoside A and B, dehydrotomatoside also are isolated from the fruit (44, 49–51). In S. melongena, α-solamargine, α-solanine, solasonine, and solasodine have been reported in fruit, root, and peel (1, 44, 49, 50). In addition, Lelario, De Maria (52) reported presence of solanidenetriol chacotriose, solanidenedio chacotriose, dehydrosolamargine, solanandaine isomer I, II and II, solanandaine, robenoside B, spirosolenol chacotriose, malonyl-solanandaine, solanidatetraenol chacotriose, arudonine, and malonyl-solamargine. Other species of therapeutic interest, such as S. torvum have shown the content of solasodine, solasonine, and solamargine (53–56). Likewise, S. nigrum presents solanine and other steroidal glycoalkaloids, such as solasodine, α and β-solamargine, β2-solasonine, solasonine, solamargine, and 12β, 27-dihydroxy solasodine (57–59).
Saponins
Saponins are glycosidic compounds consisting of an aglycone (sapogenin) linked to one or more o oligosaccharide moieties. Saponins are classified according to their structure into steroidal or triterpenoid (60, 61). Most saponins are poorly absorbed in the intestine, have foaming properties in aqueous solutions, exert a hemolytic effect, and cause a bitter taste and astringency (60). Nonetheless, they have been proved to have potential health benefits with anti-insulin resistance and anti-obesogenic effects (62). Saponins obtained from Solanum species have shown antitumor, anti-inflammatory, antiviral, antimycotic, antioxidant, hypoglycemic, and hypolipidemic activities (63). In S. melongena, the presence of saponins has been reported especially in the seeds; cholestane-type steroidal melongosides-N, O, P, R, S (64), as well as furostanol-type steroidal saponins, melongoside T-V (63). Saponins isolated from S. melongena peels showed inhibition of the enzyme lipase, which was more effective than the drug used as control (orlistat) (8). Diosgenin is present in dietary Solanum species and has been isolated to study its health-promoting effects: modulating oxidative stress, improving lipid profile, and regulating mitochondrial dysfunction pathway (65). In S. surattense at least 11 different saponins have been isolated and shown in vitro cytotoxic activities against cancer lines (66). The fruits from S. torvum have various steroidal saponins, proven to have anticancer effects against breast, liver, gastric, and lung cancer lines (9, 67).
Carotenoids
Carotenoids are lipophilic isoprenoid compounds, classified as cyclic or acyclic according to the presence or absence of an end ring on their structure (68–71). The most common carotenoids found in blood plasma are lycopene, β-carotene, and lutein (72). Several health benefits have been attributed to carotenoids like immunomodulators (73, 74), improving eye (75, 76), heart health (77), protecting skin from UV damage, improving brain functions during childhood (78), cancer prevention (79), among others (69). In the Solanum genus, S. lycopersicum has been an outstanding source of carotenoids, lycopene being the most common (2, 70). Thus, the tomato fruit and its products are the principal sources of lycopene for humans (71). Other types of carotenoids are found in different tomato cultivars are β-carotene, phytoene, and phytofluene (80–82). S. phureja has zeaxanthin, violaxanthin, antheraxanthin, lutein, and β-carotene (83). A worldwide important crop S. tuberosum is a great source of β-carotene, astaxanthin, and zeaxanthin (71). Less common sources of carotenoids have been found, for example, S. betaceum (yellow tamarillo) and S. sessiliflorum (cocona). The yellow tamarillo is known for its carotenoid content, being β-cryptoxanthin, β-carotene, zeaxanthin, and lutein, the most abundant (84). Cocona is rich in lycopene and presents higher levels of β-carotene than tomato (85). Carotenoids are pro-vitamin A compounds, a key vitamin for growing, seeing, protecting against diseases, and for reproduction (2).
Bioaccessibility of Antioxidants in Solanum Edibles
It is important to consider the structural changes antioxidants suffer after ingestion. Processing is a starting point in this journey, from cutting or mashing to boiling, baking, or even freezing, that can help release antioxidant compounds, thus increasing their bioaccessibility. Cooking in water can reduce the content of hydrosoluble compounds but cooking with oil can cause synergy and increase the bioaccessibility of lipophilic antioxidants. High temperatures break down cell walls and could increase phytochemicals bioaccessibility, but thermolabile compounds are highly degraded (86). Bioaccessibility is studied using different models to simulate gastrointestinal digestion, allowing researchers to calculate the portion of phytochemicals available for absorption. Phytochemicals can be released from the food matrix due to the simulated conditions based on the physiological data of digestion: electrolytes, digestive enzymes, dilution, pH, and time of digestion (87). Therefore, gastrointestinal digestion can degrade and transform antioxidant compounds, especially those that are highly sensitive to pH changes, such as anthocyanins. Digestive enzymes can also breakdown phytochemicals; like phenolic compounds attached to sugars. Enzymes like the lactase phloridzin hydrolase can hydrolysate sugar moieties from glycosylated bioactive molecules (5, 88).
Eggplant total phenolic content (TPC) was reported before and after four different cooking methods: baking, boiling, frying, and grilling. Antioxidant activity was determined using the ABTS assay, and reducing capacity was quantified using the FRAP (ferric reducing/antioxidant power) assay. Results indicated that TPC was improved >300% by frying, 67% by baking, and 42% by boiling. Grilling eggplant decreased TPC by 34.5%. Raw, boiled, and baked eggplant samples subjected to in vitro digestion showed bioaccessibility of TPC of 112.5, 93.4, and 101.8%, respectively; this suggests that phenolic compounds were released after in vitro digestion. Fried undigested samples showed the highest amount of TPC, but once the simulated digestion was performed, the bioaccessibility was 67%, the lowest compared to the other three samples. Grilling and in vitro digesting eggplant showed that bioaccessibility of TPC went up to 217.4%. The ABTS and FRAP results were consistent with the TPC in all digested and undigested samples. There were two exceptions: the digested boiled samples in the ABTS assay and the raw samples in the FRAP assay. The boiled digested samples increased by 336.4 %; this could be because the phenolic compounds reactive to the ABTS radical were released only after the simulated digestion. Moreover, in the second case, a bioaccessibility of 24% was attributed to a failure in extracting the reducing compounds during digestion, assuming the solvent used with the raw samples was more efficient than in vitro digestion (35). Thus, it is suggested that cooking can break down cell walls releasing antioxidants and can increase their bioaccessibility; however, they can also be more susceptible to degradation.
Drying methods have been assessed to preserve the phenolic content and the antioxidant capacity of S. melongena. Freezing, drying tunnel, and drying oven methods were evaluated, combined with slicing and mincing the eggplant. Results indicate that sliced eggplant dried at 45–50°C in a drying oven was the best option to obtain a flour rich in bioactive compounds. In the same study, freezing the material had a negative effect on this objective (89). A study was conducted using tomato products to determine the bioaccessibility of lycopene. The tomato pulp was processed by high-pressure homogenization and microwave heating into different end-products in the presence of three different oils: coconut, olive, and fish. High-pressure homogenization, followed by heating at 90°C, increased the bioaccessibility of lycopene. It hypothesized that these processes damage the cellular barriers allowing lycopene to be more bioaccessible (90).
The bioaccessibility of β-carotene in grape tomatoes ranged between 14 and 31%, considering they used two different digestion methods (91). Common home processes like paste processing and drying significantly increased total lycopene, phenolic, and flavonoid content, as well as the total antioxidant capacity. It is suggested that thermal processing disrupts cell membranes and cell walls, releasing lycopene from the insoluble portion of the matrix (92). Synergic interactions have been identified to benefit bioaccessibility. For instance, red cabbage was co-digested with different vegetables, enhancing bioaccessibility of total anthocyanins by 10–15% when samples were carotenoid-rich, like tomatoes. In contrast, the carotenoid bioaccessibility was decreased by 42–56%. This example of phytochemical interaction shows that some combinations exert synergy and others antagonism in bioaccessibility (93). Anthocyanins are highly sensitive to pH changes that naturally occur in the digestive tract, which has led to developing techniques to improve bioaccessibility and bioactivity, such as microencapsulation (94).
After boiling, the bioaccessibility of polyphenols in white and purple potatoes was evaluated. All polyphenols in the samples increased during the gastric phase of the in vitro digestion but decreased during the intestinal phase. Nonetheless, the boiled undigested samples had lower content of polyphenols. It is discussed that common chemical extractions underestimate the polyphenol content that could be released in the intestine. p-Coumaric acid in the purple potato was not detectable in the gastric phase, but it was detected in the intestinal phase, and it was 16x higher than in the boiled undigested samples. Also, caffeic acid in white potato was the only phenolic that increased its bioaccessibility in the intestinal phase. It is hypothesized that soluble polyphenols accumulate in cell vacuoles, which are released by pepsin action during gastric digestion. Chlorogenic acid interacts with starch, increasing its bioaccessibility and delaying absorption because it is only released once digested (95).
Two purple potatoes (Amachi and Leona) were subjected to simulated digestion, showing that the total anthocyanin concentration was over 30-fold higher in Amachi compared to Leona digests. In descending colon digesta, concentrations in Leona were 7-fold higher than in Amachi. This data was relevant because of the interest in testing anthocyanins against tumorigenic colon cells (Caco-2). Amachi digesta caused cytotoxicity in non-tumorigenic cells, while Leona's only caused cytotoxicity in tumorigenic cells. Also, it is suggested that microbial metabolism can decrease anthocyanin levels (96).
The bioaccessibility of phenolic compounds in S. nigrum leaves to evaluate the effect of heating in the release of phenolic compounds has been studied. The phenolic compounds myricetin, quercetin-3-O-robinoside, 3,4-dicaffeoylquinic acid, 3-caffeoylquinic acid, and rutin were the most abundant. The boiling process improved the phenolic content but decreased after the in vitro digestion (36). These results differ from other species of the genus where phenolics were also put through simulated digestion, but the process increases the content. The discrepancies are attributed to the interaction of certain phenolics like chlorogenic acid with starch, which was not abundant in the samples (95). Despite the decreased content of phenolic compounds in S. nigrum samples after digestion, they still had bioactivity against oxidative stress and prevented DNA oxidative damage (36).
The fruits of S. betaceum are known as a functional ingredient; their phytochemicals have been linked to metabolic syndrome prevention. In this subject, simulated digestion was carried out using the fruit's seeds, pulp, and skin, and bioactive effects such as enzyme inhibition and antioxidant activity were analyzed. Phenolic acids, anthocyanins, condensed tannins, carotenoids (only in pulp) were quantified and showed enzyme inhibitory properties. The extracts were able to inhibit α-glucosidase, α-amylase, and lipase before and after digestion, and the seeds extract inhibition power was improved after digestion. The seed extracts showed to be the most bioactive, and this was linked to the presence of condensed tannins that was higher than in skin and pulp samples (97).
Implications of the Bioavailability of Solanum Antioxidants and their Health-Promoting Properties
The bioavailability of most compounds is crucial to the bioactive effect, and is affected by factors like the individual factors and characteristics of the consumer, interaction with other compounds, delivery matrix, preparation processes, bioactive type/category, chemical structure, and others (98, 99). The pharmacological concept of bioavailability considers liberation, absorption, distribution, metabolism, and excretion, generally known as LADME (100). For all of these factors and the low absorption in the gastrointestinal tract, phytochemicals have low bioavailability (5). Information about the bioavailability of specific phytochemicals is still limited, but we can predict the bioavailability of phytochemicals by using the Lipinski's rule (101); which predicts the drug-likeness of the passive absorption of a molecule considering five chemical characteristics: a molecular weight ≤500, partition coefficient (LogP) <5, hydrogen bond donors <5, and a maximum of 10 hydrogen receptors (102) (See Supplementary Table 1).
The evaluation of the bioavailability of phenolic compounds has led to determine that their glycosylation directs the route of their absorption; glycones are transported through active absorption and aglycones through passive diffusion. After their absorption in the intestine, most phenolics are extensively metabolized by xenobiotic enzymes abundant in enterocytes and liver for their excretion through bile, feces, and urine (5). Even though most phenolics are easily degraded during their passage through the gastrointestinal tract, some are highly absorbable. Caffeic acid is 95% absorbed in the small intestine and stomach. Also, chlorogenic acid, the caffeic acid ester form, is absorbed in the colon after the microbiota transforms it into several metabolites (103, 104). The fact that chlorogenic acid usually reaches the colon has been considered beneficial for gut health because it promotes the growth of Bifidobacterium species (105). Therefore, chlorogenic acid is considered highly bioavailable for humans; it is estimated that 33% is absorbed intact in the stomach and 7% in the small intestine after hydrolysis (106).
Alkaloids found in Solanum have low bioavailability. Therefore, some studies have been focused to improve their absorption, mainly delivery via liposomes, nanoparticles, gels, and emulsions. Transdermal delivery has also been another option, seeking to obtain effective products and avoiding side effects (2, 107). Furthermore, most saponins are hydrosoluble due to their glycosidic groups. They have an amphiphilic nature and, therefore, the ability for self-micellization in the gastrointestinal environment and have shown to be stable to pH variations during digestion. Some saponins can be chemically hydrolyzed by acid or alkali, forming sapogenins, prosapogenins, sugar residues, or monosaccharides. When gastric digestion is simulated, some saponins show deglycosylation, dehydration, hydration, and oxygenation, leading to the presence of different structures connected to saponins' anticarcinogenic activities. Nevertheless, the bioavailability of saponins has not been widely studied (61, 104, 108). Moreover, carotenoids found in cultivars of tomato and their products have been widely studied, and there are valuable results in this context (See Table 2). Pro-vitamin A (β-carotene) has also been evaluated in eggplant. Although it is hypothesized that bioavailability can be reduced when these compounds interact with some vitamins, aspirin, and sulphonamides (69, 113), because these groups of phytochemicals may compete for absorption; for example, co-consumption of lutein has a negative effect on the absorption of β-carotene and vice versa (99, 115) (Table 2).
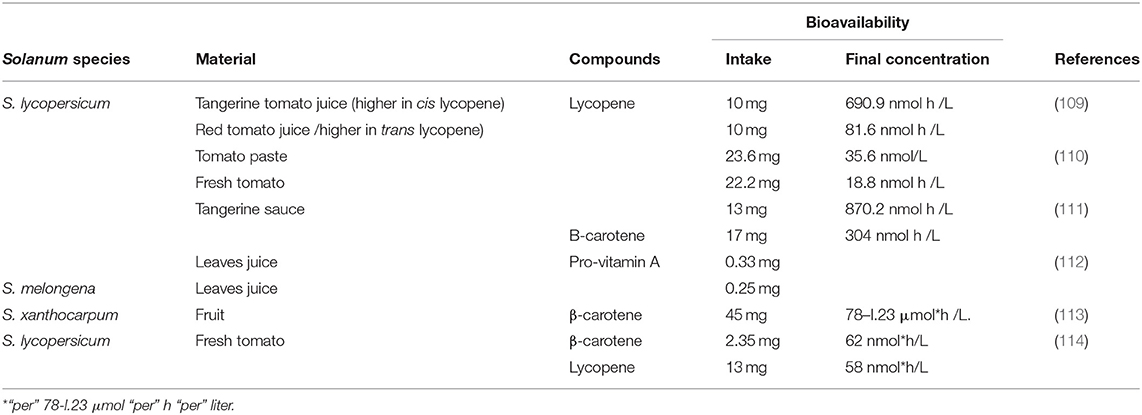
Table 2. Bioavailability of compounds in Solanum species.
Conclusions
Plants of the Solanum genus contain bioactive compounds that are antioxidant agents and have different mechanisms of action to prevent or lessen diseases and their complications. The bioactive potential of diverse materials has been proven, but there is constant interest in evaluating the transformations during their digestion and absorption. The amount of compound or mixture of compounds needed to achieve the desired effect is also a matter of research to formulate effective and safe phytopharmaceuticals. Fruits, roots, and aerial parts of plants among the Solanum genus can benefit human beings by improving their health when consumed as part of the daily diet, as a nutraceutical, or biopharmaceutical.
Author Contributions
EG-G and JH contributed to the conception of the manuscript. CE-R, LC-A, and LM-I contributed to the writing. EG-G and JH edited the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
CE-R thanks CONACYT for the doctoral scholarship. EG-G would like to thank Cátedras CONACYT for the project 397.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2021.790582/full#supplementary-material
References
1. Kaunda JS, Zhang Y-J. The genus Solanum: an ethnopharmacological, phytochemical and biological properties review. Nat Prod Bioprospect. (2019) 9:77–137. doi: 10.1007/s13659-019-0201-6
2. Contreras-Angulo L, Emus-Medina A, Gutierrez-Grijalva E, Ambriz-Pérez D, Elizalde-Romero C, Heredia JB. Pharmacological Potential of Solanum Genus. In: Souza AV, editor. Solanum: an Overview. Plant Science Research and Practices. 1. New York, NY: Nova Science (2020). p. 199.
3. Gürbüz N, Uluişik S, Frary A, Frary A, Doganlar S. Health benefits and bioactive compounds of eggplant. Food Chem. (2018) 268:602–10. doi: 10.1016/j.foodchem.2018.06.093
4. Scorsatto M, Rosa G, Luiz R, Mulder A, Teodoro A, De Oliveira JM. Effect of eggplant flour (Solanum melongena L.) associated with hypoenergetic diet on antioxidant status in overweight women - a randomised clinical trial. Int J Food Sci. (2019) 54:2182–189. doi: 10.1111/ijfs.14125
5. Velderrain-Rodríguez GR, Palafox-Carlos H, Wall-Medrano A, Ayala-Zavala JF, Chen CY, Robles-Sánchez M, et al. Phenolic compounds: their journey after intake. Food Funct. (2014) 5:189–97. doi: 10.1039/C3FO60361J
6. Sun J, Gu Y-F, Su X-Q, Li M-M, Huo H-X, Zhang J, et al. Anti-inflammatory lignanamides from the roots of Solanum melongena L. Fitoterapia. (2014) 98:110–6. doi: 10.1016/j.fitote.2014.07.012
7. Liu X, Luo J, Kong L. Phenylethyl cinnamides as potential α-glucosidase inhibitors from the roots of Solanum Melongena. Nat Prod Commun. (2011) 6:1934578X1100600623. doi: 10.1177/1934578X1100600623
8. Subandi S, Zakiyaturrodliyah L, Brotosudarmo T. Saponin from purple eggplant (Solanum melongena L.) and their activity as pancreatic lipase inhibitor. IOP Conf Ser Mater Sci Eng. (2019) 509:012139. doi: 10.1088/1757-899X/509/1/012139
9. Lu Y, Luo J, Huang X, Kong L. Four new steroidal glycosides from Solanum torvum and their cytotoxic activities. Steroids. (2009) 74:95–101. doi: 10.1016/j.steroids.2008.09.011
10. Lee C-L, Hwang T-L, He W-J, Tsai Y-H, Yen C-T, Yen H-F, et al. Anti-neutrophilic inflammatory steroidal glycosides from Solanum torvum. Phytochemistry. (2013) 95:315–21. doi: 10.1016/j.phytochem.2013.06.015
11. Ma T, Zhang Y, Zhang C, Luo J-G, Kong L-Y. Downregulation of TIGAR sensitizes the antitumor effect of physapubenolide through increasing intracellular ROS levels to trigger apoptosis and autophagosome formation in human breast carcinoma cells. Biochem Pharmacol. (2017) 143:90–106. doi: 10.1016/j.bcp.2017.07.018
12. Shakya R, Navarre DA. LC-MS analysis of solanidane glycoalkaloid diversity among tubers of four wild potato species and three cultivars (Solanum tuberosum). J Agric Food Chem. (2008) 56:6949–58. doi: 10.1021/jf8006618
13. Li J, Zhang L, Huang C, Guo F, Li Y. Five new cyotoxic steroidal glycosides from the fruits of Solanum torvum. Fitoterapia. (2014) 93:209–15. doi: 10.1016/j.fitote.2014.01.009
14. Arthan D, Svasti J, Kittakoop P, Pittayakhachonwut D, Tanticharoen M, Thebtaranonth Y. Antiviral isoflavonoid sulfate and steroidal glycosides from the fruits of Solanum torvum. Phytochemistry. (2002) 59:459–63. doi: 10.1016/S0031-9422(01)00417-4
15. Mohan M, Attarde D, Momin R, Kasture S. Antidepressant, anxiolytic and adaptogenic activity of torvanol A: an isoflavonoid from seeds of Solanum torvum. Nat Prod Res. (2013) 27:2140–3. doi: 10.1080/14786419.2013.778853
16. Balachandran C, Emi N, Arun Y, Yamamoto Y, Ahilan B, Sangeetha B, et al. In vitro anticancer activity of methyl caffeate isolated from Solanum torvum Swartz. Fruit Chem Biol Interact. (2015) 242:81–90. doi: 10.1016/j.cbi.2015.09.023
17. Balachandran C, Duraipandiyan V, Al-Dhabi NA, Balakrishna K, Kalia NP, Rajput VS, et al. Antimicrobial and antimycobacterial activities of methyl caffeate isolated from Solanum torvum Swartz. Fruit Indian J Microbiol. (2012) 52:676–81. doi: 10.1007/s12088-012-0313-8
18. Gandhi GR, Ignacimuthu S, Paulraj MG, Sasikumar P. Antihyperglycemic activity and antidiabetic effect of methyl caffeate isolated from Solanum torvum Swartz. Fruit in streptozotocin induced diabetic rats. Eur J Pharmacol. (2011) 670:623–31. doi: 10.1016/j.ejphar.2011.09.159
19. Gandhi GR, Ignacimuthu S, Paulraj MG. Solanum torvum Swartz. Fruit containing phenolic compounds shows antidiabetic and antioxidant effects in streptozotocin induced diabetic rats. Food Chem Toxicol. (2011) 49:2725–33. doi: 10.1016/j.fct.2011.08.005
20. Takahashi K, Yoshioka Y, Kato E, Katsuki S, Iida O, Hosokawa K, et al. Methyl caffeate as an alpha-glucosidase inhibitor from solanum torvum fruits and the activity of related compounds. Biosci Biotechnol Biochem. (2010) 74:741–5. doi: 10.1271/bbb.90789
21. Lopes LC, de Carvalho JE, Kakimore M, Vendramini-Costa DB, Medeiros MA, Spindola HM, et al. Pharmacological characterization of Solanum cernuum Vell.: 31-norcycloartanones with analgesic and anti-inflammatory properties. Inflammopharmacology. (2014) 22:179–85. doi: 10.1007/s10787-013-0182-8
22. Yao F, Song Q-L, Zhang L, Li G-S, Dai S-J. Solajiangxins A–C, three new cytotoxic sesquiterpenoids from Solanum lyratum. Fitoterapia. (2013) 89:200–4. doi: 10.1016/j.fitote.2013.05.020
23. Zhang L, Li G-S, Yao F, Yue X-D, Dai S-J. Three new sesquiterpenoids from Solanum septemlobum with cytotoxic activities. Phytochem Lett. (2015) 11:173–6. doi: 10.1016/j.phytol.2015.03.016
24. Elekofehinti OO, Kamdem JP, Bolingon AA, Athayde ML, Lopes SR, Waczuk EP, et al. African eggplant (Solanum anguivi Lam.) fruit with bioactive polyphenolic compounds exerts in vitro antioxidant properties and inhibits Ca2+-induced mitochondrial swelling. Asian Pac J Trop Biomed. (2013) 3:757–66. doi: 10.1016/S2221-1691(13)60152-5
25. Laura A, Moreno-Escamilla JO, Rodrigo-García J, Alvarez-Parrilla E. Phenolic Compounds. Postharvest Physiology and Biochemistry of Fruits and Vegetables. Quéretaro: Elsevier (2019). p. 253–71.
26. Hurtado NH, Morales AL, González-Miret ML, Escudero-Gilete ML, Heredia FJ. Colour, pH stability and antioxidant activity of anthocyanin rutinosides isolated from tamarillo fruit (Solanum betaceum Cav.). Food Chem. (2009) 117:88–93. doi: 10.1016/j.foodchem.2009.03.081
27. Osorio C, Hurtado N, Dawid C, Hofmann T, Heredia-Mira FJ, Morales AL. Chemical characterisation of anthocyanins in tamarillo (Solanum betaceum Cav.) and andes berry (Rubus glaucus Benth.) fruits. Food Chem. (2012) 132:1915–21. doi: 10.1016/j.foodchem.2011.12.026
28. Hawas UW, Soliman GM, El-Kassem LTA, Farrag ARH, Mahmoud K, León F. A new flavonoid C-Glycoside from Solanum elaeagnifolium with hepatoprotective and curative activities against paracetamol- induced liver injury in mice. Z Naturforsch C J Biosci. (2013) 68:19–28. doi: 10.1515/znc-2013-1-204
29. Silva EL, Almeida-Lafetá RC, Borges RM, Staerk D. Dual high-resolution inhibition profiling and HPLC-HRMS-SPE-NMR analysis for identification of α-glucosidase and radical scavenging inhibitors in Solanum americanum Mill. Fitoterapia. (2017) 118:42–8. doi: 10.1016/j.fitote.2017.02.002
30. Petreanu M, Guimarães ÁAA, Broering MF, Ferreira EK, Machado ID, Gois ALT, et al. Antiproliferative and toxicological properties of methanolic extract obtained from Solanum capsicoides all. Seeds and carpesterol. Naunyn Schmied Arch Pharmacol. (2016) 389:1123–31. doi: 10.1007/s00210-016-1275-x
31. Alhaithloul HAS, Galal FH, Seufi AM. Effect of extreme temperature changes on phenolic, flavonoid contents and antioxidant activity of tomato seedlings (Solanum lycopersicum L.). PeerJ. (2021) 9:e11193. doi: 10.7717/peerj.11193
32. Pinela J, Montoya C, Carvalho AM, Martins V, Rocha F, Barata AM, et al. Phenolic composition and antioxidant properties of ex-situ conserved tomato (Solanum lycopersicum L.) germplasm. Food Res Int. (2019) 125:108545. doi: 10.1016/j.foodres.2019.108545
33. Staszek P, Krasuska U, Bederska-Blaszczyk M, Gniazdowska A. Canavanine increases the content of phenolic compounds in tomato (Solanum lycopersicum L.) roots. Plants. (2020) 9:1595. doi: 10.3390/plants9111595
34. Lima MD, Andreou R, Charalampopoulos D, Chatzifragkou A. Supercritical carbon dioxide extraction of phenolic compounds from potato (Solanum tuberosum) peels. Appl Sci. (2021) 11:3410. doi: 10.3390/app11083410
35. Martini S, Conte A, Cattivelli A, Tagliazucchi D. Domestic cooking methods affect the stability and bioaccessibility of dark purple eggplant (Solanum melongena) phenolic compounds. Food Chem. (2021) 341:128298. doi: 10.1016/j.foodchem.2020.128298
36. Moyo SM, Serem JC, Bester MJ, Mavumengwana V, Kayitesi E. The impact of boiling and in vitro human digestion of Solanum nigrum complex (Black nightshade) on phenolic compounds bioactivity and bioaccessibility. Food Res Int. (2020) 137:109720. doi: 10.1016/j.foodres.2020.109720
37. Diep T, Pook C, Yoo M. Phenolic and anthocyanin compounds and antioxidant activity of tamarillo (Solanum betaceum Cav.). Antioxidants. (2020) 9:169. doi: 10.3390/antiox9020169
38. Svobodova B, Barros L, Sopik T, Calhelha RC, Heleno S, Alves MJ, et al. Non-edible parts of Solanum stramoniifolium Jacq. - a new potent source of bioactive extracts rich in phenolic compounds for functional foods. Food Funct. (2017) 8:2013–21. doi: 10.1039/C7FO00297A
39. Oszmianski J, Kolniak-Ostek J, Wojdylo A. Characterization of phenolic compounds and antioxidant activity of Solanum scabrum and Solanum burbankii berries. J Agric Food Chem. (2014) 62:1512–9. doi: 10.1021/jf4045233
40. Plazas M, Andujar I, Vilanova S, Hurtado M, Gramazio P, Herraiz FJ, et al. Breeding for chlorogenic acid content in eggplant: interest and prospects. Not Bot Horti Agrobo. (2013) 41:26–35. doi: 10.15835/nbha4119036
41. Gutiérrez-Grijalva EP, López-Martínez LX, Contreras-Angulo LA, Elizalde-Romero CA, Heredia JB. Plant alkaloids: structures and bioactive properties. In: Swamy MK, editor. Plant-derived Bioactives: Chemistry and Mode of Action. Singapore: Springer Singapore (2020). p. 85–117.
42. Jayakumar K, Murugan K. Solanum alkaloids and their pharmaceutical roles: a review. J Anal Pharm Res. (2016) 3:00075. doi: 10.15406/japlr.2016.03.00075
43. Hussain G, Rasul A, Anwar H, Aziz N, Razzaq A, Wei W, et al. Role of plant derived alkaloids and their mechanism in neurodegenerative disorders. Int J Biol Sci. (2018) 14:341–57. doi: 10.7150/ijbs.23247
44. Milner SE, Brunton NP, Jones PW, O'Brien NM, Collins SG, Maguire AR. Bioactivities of glycoalkaloids and their aglycones from Solanum species. J Agric Food Chem. (2011) 59:3454–84. doi: 10.1021/jf200439q
45. Sánchez Maldonado AF, Mudge E, Gänzle MG, Schieber A. Extraction and fractionation of phenolic acids and glycoalkaloids from potato peels using acidified water/ethanol-based solvents. Food Res Int. (2014) 65:27–34. doi: 10.1016/j.foodres.2014.06.018
46. Lee K-G, Lee S-G, Lee H-H, Lee HJ, Shin J-S, Kim N-J, et al. α-Chaconine isolated from a Solanum tuberosum L. cv Jayoung suppresses lipopolysaccharide-induced pro-inflammatory mediators via AP-1 inactivation in RAW 264.7 macrophages and protects mice from endotoxin shock. Chem Biol Interact. (2015) 235:85–94. doi: 10.1016/j.cbi.2015.04.015
47. Caprioli G, Cahill MG, Vittori S, James KJ. Liquid chromatography–hybrid linear ion trap–high-resolution mass spectrometry (LTQ-Orbitrap) method for the determination of glycoalkaloids and their aglycons in potato samples. Food Anal Methods. (2014) 7:1367–72. doi: 10.1007/s12161-013-9758-6
48. Deng Y, He M, Feng F, Feng X, Zhang Y, Zhang F. The distribution and changes of glycoalkaloids in potato tubers under different storage time based on MALDI-TOF mass spectrometry imaging. Talanta. (2021) 221:121453. doi: 10.1016/j.talanta.2020.121453
49. Chowański S, Adamski Z, Marciniak P, Rosiński G, Büyükgüzel E, Büyükgüzel K, et al. A review of bioinsecticidal activity of solanaceae alkaloids. Toxins. (2016) 8:60. doi: 10.3390/toxins8030060
50. Friedman M, Levin CE, Lee S-U, Kim H-J, Lee I-S, Byun J-O, et al. Tomatine-containing green tomato extracts inhibit growth of human breast, colon, liver, and stomach cancer cells. J Agric Food Chem. (2009) 57:5727–33. doi: 10.1021/jf900364j
51. Hövelmann Y, Hahn M, Hübner F, Humpf H-U. Detection of novel cytotoxic imidazole alkaloids in tomato products by LC-MS/MS. J Am Chem Soc. (2019) 67:3670–8. doi: 10.1021/acs.jafc.9b00461
52. Lelario F, De Maria S, Rivelli AR, Russo D, Milella L, Bufo SA, et al. A complete survey of glycoalkaloids using LC-FTICR-MS and IRMPD in a commercial variety and a local landrace of eggplant (Solanum melongena L.) and their anticholinesterase and antioxidant activities. Toxins. (2019) 11:230. doi: 10.3390/toxins11040230
53. Nurit-Silva K, Costa-Silva R, Coelho VP, Agra MdF. A pharmacobotanical study of vegetative organs of Solanum torvum. Rev Bras Farmacogn. (2011) 21:568–74. doi: 10.1590/S0102-695X2011005000101
54. Panigrahi S, Muthuraman MS, Natesan R, Pemiah B. Anticancer activity of ethanolic extract of Solanum torvum sw. Int J Pharm Pharm Sci. (2014) 6:93–8.
55. Yousaf Z, Wang Y, Baydoun E. Phytochemistry and pharmacological studies on Solanum torvum Swartz. J Appl Pharm Sci. (2013) 3:152–60. doi: 10.7324/japs.2013.3428
56. Pérez-Amador M, Muñoz Ocotero V, García Castañeda J, González Esquinca A. Alkaloids in Solanum torvum Sw (Solanaceae). Phyton Int J Exp Bot. (2007) 76:39–45. doi: 10.32604/phyton.2007.76.039
57. Atanu F, Ebiloma U, Ajayi E. A review of the pharmacological aspects of Solanum nigrum Linn. Biotechnol Mol Biol Rev. (2011) 6:1–7. doi: 10.5897/bmbr2011.0001
58. Gu X-Y, Shen X-F, Wang L, Wu Z-W, Li F, Chen B, et al. Bioactive steroidal alkaloids from the fruits of Solanum nigrum. Phytochemistry. (2018) 147:125–31. doi: 10.1016/j.phytochem.2017.12.020
59. Rani YS, Reddy VJ, Basha SJ, Koshma M, Hanumanthu G, Swaroopa P. A review on Solanum nigrum. World J Pharm Res. (2017) 6:293–303. doi: 10.20959/wjpps201712-10538
60. Mohan VR, Tresina PS, Daffodil ED. Antinutritional factors in legume seeds: characteristics and determination. In: Caballero B, Finglas PM, Toldrá F, editors. Encyclopedia of Food and Health. Oxford: Academic Press (2016). p. 211–20.
61. Navarro del Hierro J, Herrera T, Fornari T, Reglero G, Martin D. The gastrointestinal behavior of saponins and its significance for their bioavailability and bioactivities. J Funct Foods. (2018) 40:484–97. doi: 10.1016/j.jff.2017.11.032
62. Luo Z, Xu W, Zhang Y, Di L, Shan J. A review of saponin intervention in metabolic syndrome suggests further study on intestinal microbiota. Pharmacol Res. (2020) 160:105088. doi: 10.1016/j.phrs.2020.105088
63. Chen F-F, Zhou J, Zhang Y-W, Chen Y-P, Wang Y-R, Zhao Y-F, et al. Five new steroidal saponins from the seeds of Solanum melongena L. Phytochem Lett. (2021) 41:21–6. doi: 10.1016/j.phytol.2020.10.008
64. Jeepipalli SPK, Du B, Sabitaliyevich UY, Xu B. New insights into potential nutritional effects of dietary saponins in protecting against the development of obesity. Food Chem. (2020) 318:126474. doi: 10.1016/j.foodchem.2020.126474
65. Gong G, Qin Y, Huang W, Zhou S, Wu X, Yang X, et al. Protective effects of diosgenin in the hyperlipidemic rat model and in human vascular endothelial cells against hydrogen peroxide-induced apoptosis. Chem Biol Interact. (2010) 184:366–75. doi: 10.1016/j.cbi.2010.02.005
66. Lu Y, Luo J, Kong L. Steroidal alkaloid saponins and steroidal saponins from Solanum surattense. Phytochemistry. (2011) 72:668–73. doi: 10.1016/j.phytochem.2011.01.028
67. Pérez Colmenares A, Rojas LB, Mitaine-Offer A-C, Pouységu L, Quideau S, Miyamoto T, et al. Steroidal saponins from the fruits of Solanum torvum. Phytochemistry. (2013) 86:137–43. doi: 10.1016/j.phytochem.2012.10.010
68. Milani A, Basirnejad M, Shahbazi S, Bolhassani A. Carotenoids: biochemistry, pharmacology and treatment. Br J Pharmacol. (2017) 174:1290–324. doi: 10.1111/bph.13625
69. Eggersdorfer M, Wyss A. Carotenoids in human nutrition and health. Arch Biochem Biophys. (2018) 652:18–26. doi: 10.1016/j.abb.2018.06.001
70. Szabo K, Emoke Teleky B, Ranga F, Simon E, Lelia Pop O, Babalau-Fuss V, et al. Bioaccessibility of microencapsulated carotenoids, recovered from tomato processing industrial by-products, using in vitro digestion model. LWT. (2021) 152:112285. doi: 10.1016/j.lwt.2021.112285
71. Rodriguez-Concepcion M, Avalos J, Bonet ML, Boronat A, Gomez-Gomez L, Hornero-Mendez D, et al. A global perspective on carotenoids: metabolism, biotechnology, and benefits for nutrition and health. Prog Lipid Res. (2018) 70:62–93. doi: 10.1016/j.plipres.2018.04.004
72. Zimmer JP, Hammond Jr BR. Possible influences of lutein and zeaxanthin on the developing retina. Clin Ophthalmol. (2007) 1:25–35.
73. Jonasson L, Wikby A, Olsson A. Low serum β-carotene reflects immune activation in patients with coronary artery disease. Nutr Metab Cardiovasc Dis. (2003) 13:120–5. doi: 10.1016/S0939-4753(03)80170-9
74. Palmer AC, West KP, Dalmiya N, Schultink W. The use and interpretation of serum retinol distributions in evaluating the public health impact of vitamin A programmes. Public Health Nutr. (2012) 15:1201–15. doi: 10.1017/S1368980012000560
75. Group A-REDSR. Lutein+ zeaxanthin and omega-3 fatty acids for age-related macular degeneration: the age-related eye disease study 2 (AREDS2) randomized clinical trial. JAMA. (2013) 309:2005–15. doi: 10.1001/jama.2013.4997
76. Chew EY, Clemons TE, SanGiovanni JP, Danis RP, Ferris FL, Elman MJ, et al. Secondary analyses of the effects of lutein/zeaxanthin on age-related macular degeneration progression: AREDS2 report No. 3. JAMA Ophthalmol. (2014) 132:142–9. doi: 10.1001/jamaophthalmol.2013.7376
77. Cheng HM, Koutsidis G, Lodge JK, Ashor AW, Siervo M, Lara J. Lycopene and tomato and risk of cardiovascular diseases: a systematic review and meta-analysis of epidemiological evidence. Crit Rev Food Sci Nutr. (2019) 59:141–58. doi: 10.1080/10408398.2017.1362630
78. Walk AM, Khan NA, Barnett SM, Raine LB, Kramer AF, Cohen NJ, et al. From neuro-pigments to neural efficiency: the relationship between retinal carotenoids and behavioral and neuroelectric indices of cognitive control in childhood. Int J Psychophysiol. (2017) 118:1–8. doi: 10.1016/j.ijpsycho.2017.05.005
79. Wan L, Tan H-L, Thomas-Ahner JM, Pearl DK, Erdman JW, Moran NE, et al. Dietary tomato and lycopene impact androgen signaling-and carcinogenesis-related gene expression during early TRAMP prostate carcinogenesis. Cancer Prev Res. (2014) 7:1228–39. doi: 10.1158/1940-6207.CAPR-14-0182
80. Riahi A, Hdider C. Bioactive compounds and antioxidant activity of organically grown tomato (Solanum lycopersicum L.) cultivars as affected by fertilization. Sci Hortic. (2013) 151:90–6. doi: 10.1016/j.scienta.2012.12.009
81. Meléndez-Martínez AJ, Mapelli-Brahm P, Benítez-González A, Stinco CM. A comprehensive review on the colorless carotenoids phytoene and phytofluene. Arch Biochem Biophys. (2015) 572:188–200. doi: 10.1016/j.abb.2015.01.003
82. Melendez-Martinez AJ, Stinco CM, Liu C, Wang X-D. A simple HPLC method for the comprehensive analysis of cis/trans (Z/E) geometrical isomers of carotenoids for nutritional studies. Food Chem. (2013) 138:1341–50. doi: 10.1016/j.foodchem.2012.10.067
83. Bonierbale M, Grüneberg W, Amoros W, Burgos G, Salas E, Porras E, et al. Total and individual carotenoid profiles in Solanum phureja cultivated potatoes: II. Development and application of near-infrared reflectance spectroscopy (NIRS) calibrations for germplasm characterization. J Food Compos Anal. (2009) 22:509–16. doi: 10.1016/j.jfca.2008.08.009
84. García JM, Giuffrida D, Dugo P, Mondello L, Osorio C. Development and characterisation of carotenoid-rich microencapsulates from tropical fruit by-products and yellow tamarillo (Solanum betaceum Cav.). Powder Technol. (2018) 339:702–9. doi: 10.1016/j.powtec.2018.08.061
85. Sereno AB, Bampi M, dos Santos IE, Ferreira SMR, Bertin RL, Krüger CCH. Mineral profile, carotenoids and composition of cocona (Solanum sessiliflorum Dunal), a wild Brazilian fruit. J Food Compos Anal. (2018) 72:32–8. doi: 10.1016/j.jfca.2018.06.001
86. Navajas-Porras B, Perez-Burillo S, Valverde Moya Á, Hinojosa-Nogueira D, Pastoriza S, Rufián Henares J. Effect of cooking methods on the antioxidant capacity of plant foods submitted to in vitro digestion–fermentation. Antioxidants. (2020) 9:1312. doi: 10.3390/antiox9121312
87. Brodkorb A, Egger L, Alminger M, Alvito P, Assunção R, Ballance S, et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat Protoc. (2019) 14:991–1014. doi: 10.1038/s41596-018-0119-1
88. Vacek J, Ulrichová J, Klejdus B, Šimánek V. Analytical methods and strategies in the study of plant polyphenolics in clinical samples. Anal Methods. (2010) 2:604–13. doi: 10.1039/c0ay00042f
89. Rodriguez-Jimenez JR, Amaya-Guerra CA, Baez-Gonzalez JG, Aguilera-Gonzalez C, Urias-Orona V, Nino-Medina G. Physicochemical, functional, and nutraceutical properties of eggplant flours obtained by different drying methods. Molecules. (2018) 23:3210. doi: 10.3390/molecules23123210
90. Colle IJP, Lemmens L, Van Buggenhout S, Met K, Van Loey AM, Hendrickx ME. Processing tomato pulp in the presence of lipids: the impact on lycopene bioaccessibility. Food Res Int. (2013) 51:32–8. doi: 10.1016/j.foodres.2012.11.024
91. Rodrigues DB, Chitchumroonchokchai C, Mariutti LRB, Mercadante AZ, Failla ML. Comparison of two static in vitro digestion methods for screening the bioaccessibility of carotenoids in fruits, vegetables, and animal products. J Agric Food Chem. (2017) 65:11220–8. doi: 10.1021/acs.jafc.7b04854
92. Kamiloglu S, Demirci M, Selen S, Toydemir G, Boyacioglu D, Capanoglu E. Home processing of tomatoes (Solanum lycopersicum): effects on in vitro bioaccessibility of total lycopene, phenolics, flavonoids, and antioxidant capacity. J Sci Food Agric. (2014) 94:2225–33. doi: 10.1002/jsfa.6546
93. Phan MAT, Bucknall MP, Arcot J. Co-ingestion of red cabbage with cherry tomato enhances digestive bioaccessibility of anthocyanins but decreases carotenoid bioaccessibility after simulated in vitro gastro-intestinal digestion. Food Chem. (2019) 298:125040. doi: 10.1016/j.foodchem.2019.125040
94. Condurache NN, Aprodu I, Craciunescu O, Tatia R, Parfene G, Barbu V, et al. Probing the functionality of bioactives from eggplant peel extracts through extraction and microencapsulation in different polymers and whey protein hydrolysates. Food Bioprocess Technol. (2019) 12:1316–329. doi: 10.1007/s11947-019-02302-1
95. Miranda L, Deußer H, Evers D. The impact of in vitro digestion on bioaccessibility of polyphenols from potatoes and sweet potatoes and their influence on iron absorption by human intestinal cells. Food Funct. (2013) 4:1595–601. doi: 10.1039/c3fo60194c
96. Kubow S, Iskandar MM, Melgar-Bermudez E, Sleno L, Sabally K, Azadi B, et al. Effects of simulated human gastrointestinal digestion of two purple-fleshed potato cultivars on anthocyanin composition and cytotoxicity in colonic cancer and non-tumorigenic cells. Nutrients. (2017) 9:953. doi: 10.3390/nu9090953
97. Orqueda ME, Torres S, Zampini IC, Cattaneo F, Di Pardo AF, Valle EM, et al. Integral use of argentinean Solanum betaceum red fruits as functional food ingredient to prevent metabolic syndrome: effect of in vitro simulated gastroduodenal digestion. Heliyon. (2020) 6:e03387. doi: 10.1016/j.heliyon.2020.e03387
98. Ting Y, Jiang Y, Ho C-T, Huang Q. Common delivery systems for enhancing in vivo bioavailability and biological efficacy of nutraceuticals. J Funct Foods. (2014) 7:112–28. doi: 10.1016/j.jff.2013.12.010
99. van het Hof KH, West CE, Weststrate JA, Hautvast JGAJ. Dietary factors that affect the bioavailability of carotenoids. J Nutr. (2000) 130:503–6. doi: 10.1093/jn/130.3.503
100. Holst B, Williamson G. Nutrients and phytochemicals: from bioavailability to bioefficacy beyond antioxidants. Curr Opin Biotechnol. (2008) 19:73–82. doi: 10.1016/j.copbio.2008.03.003
101. Wise K, Selby-Pham SNB, Selby-Pham J, Gill H. Development of intestinal bioavailability prediction (IBP) and phytochemical relative antioxidant potential prediction (PRAPP) models for optimizing functional food value of Cannabis sativa (hemp). Int J Food Prop. (2020) 23:1287–95. doi: 10.1080/10942912.2020.1797783
102. Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv Drug Deliv Rev. (1997) 23:3–25. doi: 10.1016/S0169-409X(96)00423-1
103. Tomas-Barberan F, García-Villalba R, Quartieri A, Raimondi S, Amaretti A, Leonardi A, et al. In vitro transformation of chlorogenic acid by human gut microbiota. Mol Nutr Food Res. (2014) 58:1122–31. doi: 10.1002/mnfr.201300441
105. Mills CE, Tzounis X, Oruna-Concha M-J, Mottram DS, Gibson GR, Spencer JP. In vitro colonic metabolism of coffee and chlorogenic acid results in selective changes in human faecal microbiota growth. Br J Nutr. (2015) 113:1220–7. doi: 10.1017/S0007114514003948
106. Lu H, Tian Z, Cui Y, Liu Z, Ma X. Chlorogenic acid: a comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanisms of action, and future directions. Compr Rev Food Sci Food Saf. (2020) 19:3130–58. doi: 10.1111/1541-4337.12620
107. Zheng X, Wu F, Lin X, Shen L, Feng Y. Developments in drug delivery of bioactive alkaloids derived from traditional Chinese medicine. Drug Deliv. (2018) 25:398–416. doi: 10.1080/10717544.2018.1431980
108. Singh D, Chaudhuri PK. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr Med Res. (2018) 7:33–43. doi: 10.1016/j.imr.2018.01.003
109. Cooperstone JL, Ralston RA, Riedl KM, Haufe TC, Schweiggert RM, King SA, et al. Enhanced bioavailability of lycopene when consumed as cis-isomers from tangerine compared to red tomato juice, a randomized, cross-over clinical trial. Mol Nutr Food Res. (2015) 59:658–69. doi: 10.1002/mnfr.201400658
110. Gärtner C, Stahl W, Sies H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am J Clin Nutr. (1997) 66:116–22. doi: 10.1093/ajcn/66.1.116
111. Unlu NZ, Bohn T, Francis D, Clinton SK, Schwartz SJ. Carotenoid absorption in humans consuming tomato sauces obtained from tangerine or high-beta-carotene varieties of tomatoes. J Agric Food Chem. (2007) 55:1597–603. doi: 10.1021/jf062337b
112. Madukwe E, Nwabunze A, Onyibalu C. Bioavailability of vitamins C, E and pro-vitamin A in extracts of fluted pumpkin (Telfairia occidentalis), tomato (Lycopersicum esculentum) and eggplant (Solanum melongena). Int J Basic Appl. (2013) 2:244–50. doi: 10.14419/ijbas.v2i3.883
113. Dimitrov NV, Meyer C, Ullrey DE, Chenoweth W, Michelakis A, Malone W, et al. Bioavailability of beta-carotene in humans. Am J Clin Nutr. (1988) 48:298–304. doi: 10.1093/ajcn/48.2.298
114. Schweiggert RM, Kopec RE, Villalobos-Gutierrez MG, Högel J, Quesada S, Esquivel P, et al. Carotenoids are more bioavailable from papaya than from tomato and carrot in humans: a randomised cross-over study. Br J Nutr. (2014) 111:490–8. doi: 10.1017/S0007114513002596
Keywords: solanum, antioxidant, bioactive, bioaccessibility, bioavailability, eggplant
Citation: Elizalde-Romero CA, Montoya-Inzunza LA, Contreras-Angulo LA, Heredia JB and Gutiérrez-Grijalva EP (2021) Solanum Fruits: Phytochemicals, Bioaccessibility and Bioavailability, and Their Relationship With Their Health-Promoting Effects. Front. Nutr. 8:790582. doi: 10.3389/fnut.2021.790582
Received: 07 October 2021; Accepted: 31 October 2021;
Published: 25 November 2021.
Edited by:
Abraham Wall-Medrano, Universidad Autónoma de Ciudad Juárez, MexicoReviewed by:
Efigenia Montalvo-Gonzalez, Instituto Tecnológico de Tepic, MexicoMaría Janeth Rodríguez-Roque, Autonomous University of Chihuahua, Mexico
Victor Manuel Zamora Gasga, Instituto Tecnológico de Tepic, Mexico
Copyright © 2021 Elizalde-Romero, Montoya-Inzunza, Contreras-Angulo, Heredia and Gutiérrez-Grijalva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erick Paul Gutiérrez-Grijalva, erick.gutierrez@ciad.mx