 Théo Veaudor
Théo Veaudor Corinne Cassier-Chauvat
Corinne Cassier-Chauvat Franck Chauvat
Franck Chauvat- Institute for Integrative Biology of the Cell (I2BC), CEA, CNRS, Univ Paris-Sud, Université Paris-Saclay, Gif-sur-Yvette, France
Cyanobacteria are widely-diverse prokaryotes that colonize our planet. They use solar energy to assimilate huge amounts of atmospheric CO2 and produce a large part of the biomass and oxygen that sustain most life forms. Cyanobacteria are therefore increasingly studied for basic research objectives, as well as for the photosynthetic production of chemicals with industrial interests. One potential approach to reduce the cost of future bioproduction processes is to couple them with wastewater treatment, often polluted with urea, which in any case is cheaper than nitrate. As of yet, however, research has mostly focused on a very small number of model cyanobacteria growing on nitrate. Thus, the genetic inventory of the cyanobacterial phylum is still insufficiently employed to meaningfully select the right host for the right purpose. This review reports what is known about urea transport and catabolism in cyanobacteria, and what can be inferred from the comparative analysis of the publicly available genome sequence of the 308 cyanobacteria. We found that most cyanobacteria mostly harbor the genes encoding the urea catabolytic enzymes urease (ureABCDEFG), but not systematically, together with the urea transport (urtABCDE). These findings are consistent with the capacity of the few tested cyanobacteria that grow on urea as the sole nitrogen source. They also indicate that urease is important for the detoxification of internally generated urea (re-cycling its carbon and nitrogen). In contrast, several cyanobacteria have urtABCDE but not ureABCDEFG, suggesting that urtABCDE could operate in the transport of not only urea but also of other nutrients. Only four cyanobacteria appeared to have the genes encoding the urea carboxylase (uc) and allophanate hydrolase (ah) enzymes that sequentially catabolize urea. Three of these cyanobacteria belongs to the genera Gloeobacter and Gloeomargarita that have likely diverged early from other cyanobacteria, suggesting that the urea carboxylase and allophanate hydrolase enzymes appeared in cyanobacteria before urease.
Introduction
Cyanobacteria are ancient Gram-negative prokaryotes that perform the plant-like oxygen-evolving photosynthesis (here we consider the cyanobacterial phylum as consisting only of oxygenic phototrophs; Soo et al., 2017), which are regarded as the producers of our oxygenic atmosphere (Schopf, 2011), and the ancestor of plant chloroplasts (Archibald, 2009). In colonizing most waters (fresh, brackish and marine) soils and rocks of our planet, cyanobacteria have evolved as morphologically and metabolically widely-diverse microorganisms that are of high interest for basic and applied research, for a review see Cassier-Chauvat and Chauvat (2018). Attesting to their wide diversity, their genomes are widely diverse in size (and GC content, ranging from 30 to 60%), probably as the result of gain-and-loss of genes transferred by plasmids, insertion sequences and/or phages. Most cyanobacteria possess a single circular chromosome ranging from about 1.4 Mbp to about 9.0 Mbp in size and one to several plasmids (a few Kbp to several hundreds of Kbp in size). By contrast, a few marine cyanobacteria (Prochlorococcus and Synechococcus) have no plasmids, whereas Cyanothece ATCC 51142 possesses two chromosomes (one circular, 4.9 Mbp; and one linear, 0.4 Mbp) and four plasmids (ranging from 10 to 39 Kbp) (Shih et al., 2013). However, thus far research has mostly focused on a few model strains with well-established genetics and cyanobacterial diversity which has been insufficiently explored.
Collectively, cyanobacteria fix a huge amount of inorganic carbon (CO2) and nitrogen (N2, NH4, NO2, NO3 and urea) into an enormous biomass that supports a large part of the food chain. Hence, Arthrospira platensis strains have been used for centuries as food for animals and/or humans, and they are currently being tested as a way to replenish O2, recycle wastes (CO2 and urea) and provide food during long-term space missions (Verseux et al., 2016).
Cyanobacteria are also regarded as promising microbial factories for the ecological production of chemicals from solar energy, waters and atmospheric CO2 (for reviews see Knoot et al., 2018; Sun et al., 2018). To decrease the operating costs of such future cyanobacterial bio-refineries, it would be useful to develop a waste-to-biorefinery system that couples chemical production with waste-water treatment, i.e., feeding the cyanobacterial producer with organic pollutants, such as urea (Ramsundar et al., 2017) which is a cheaper nitrogen source than nitrate or ammonium and is often present in natural or waste waters.
Urea, the first organic molecule synthesized in the laboratory, is ubiquitous in nature (Carlini and Ligabue-Braun, 2016). In mammals, urea is the primary waste product of amino acid catabolism. It is distributed throughout the body and it is evacuated in urine (Rutherford, 2014). Thus, research is ongoing to recover urea (and phosphorus) from human urine (Zhang et al., 2014; Vasconcelos Fernandes et al., 2015). Urea is also present in natural waters due to its production by macro-fauna and benthic heterotrophic microorganisms, as well as its release by sediments from urea-fertilized agricultural soils (Glibert et al., 2006; Belisle et al., 2016) since about half of the nitrogen used for fertilization is applied as urea (Witte, 2011). In oceanic-estuarine waters, urea ranging from 1 nM to 50 μM can contribute to 50% or more of the total nitrogen used by cyanobacteria-rich phytoplanktonic communities (Solomon et al., 2010; Su et al., 2013). The diverse metabolic pathways of urea transport and decomposition may contribute to differences in the role that urea plays in the physiology and ecology of cyanobacteria, and in the role that each species plays in the biogeochemistry of urea.
Urea [(NH2)2CO] can be catabolized by several enzymes into NH3 (ammonia) and CO2 [(NH2)2CO + H2O ↔ CO2 + 2NH3], which can be re-injected into the cell metabolism. These ureolytic enzymes are, namely: urease (EC 3.5.1.5) and urea aminolyase (EC 6.3.4.6). They can be distinguished by their cofactor requirement (nickel for urease; ATP for urea amidolyase) as well as their inhibitor (hydroxyurea or acetohydroxamic acid for urease; avidin for amidolyase). In cyanobacteria, these inhibitors were used to show that Anabaena cylindrica ATCC 27899 possess an active urease but no urea amidolyase activity (Mackerras and Smith, 1986).
Urease (EC 3.5.1.5), also called urea aminohydrolase, is an important enzyme that plays a crucial role in various biological processes. It was the first enzyme to be crystallized and the first protein shown to contain nickel (for a review see Carlini and Ligabue-Braun, 2016). Urease is widespread in bacteria, fungi (except hemiascomyces), diatoms, and plants (Witte, 2011), but it is not present in green algae and animals. It catalyzes the ATP- and NAD(P)H-independent hydrolysis of urea into ammonia and carbamic acid (H2N–COOH), which is spontaneously hydrolyzed in carbonic acid (H2CO3) and a second ammonia molecule. These reactions are represented by the following equations (Carlini and Ligabue-Braun, 2016):
Under physiological conditions the proton of carbonic acid dissociates, and the ammonia molecules become protonated to form ammonium (NH4+) that increases the local pH. This can enable microorganisms to cope with acid challenges (Carlini and Ligabue-Braun, 2016). Hence, the pathogenic bacterium Helicobacter pylori, produces large amounts of urease (about 10% of its total proteins) to produce ammonia and to neutralize the acidic medium of the stomach to colonize it (this ammonia is toxic to host epithelial cells which significantly increases the risk of gastric ulcers and cancer). To persist in the gastric mucosa, H. pylori must also combat the host-produced reactive oxygen species. For this purpose, H. pylori uses the numerous methionine residues of its urease to quench the host oxidants; the resulting oxidized methionine residues being re-reduced by its methionine sulfoxide reductase (Schmalstig et al., 2018). Similarly, a urease-dependent alkalization of urine by Proteus mirabilis can lead to the formation of infection stones (ammonium magnesium phosphate or carbonate apatite) that contribute to the pyelonephritis (Rutherford, 2014).
Microbial ureases can also have beneficial roles for their hosts. The ureolytic bacteria thriving in the forestomach of ruminants cleaves animal-generated urea and releases ammonia that serves as the nitrogen source for the rumen microbiota, which plays a crucial role in the feeding of these animals (Carlini and Ligabue-Braun, 2016).
Plant ureases can have insecticidal effects (Witte, 2011). Upon plant ingestion by insects, the urease proteolysis catalyzed by the insect digestive enzyme releases peptides that can affect the contraction of insect muscles (Lopes et al., 2015).
In calcium-rich natural environments the increased pH and carbonate concentration caused by ureolytic microbes favors the formation and precipitation of calcium carbonate (CaCO3). This biomineralization process can be applied to the removal of calcium, heavy metals, and radionucleotides from water, as well as to the strengthening of soil, sand, stone, and cementitious materials. Many cyanobacteria of various genera have been reported to precipitate calcium carbonate as their metabolic product, thereby contributing to the formation of reservoir rocks like stromatolites and dolomites (Sarayu et al., 2014). The undeniable multifunctionality of ureases allows their inclusion in the moonlighting protein group (Carlini and Ligabue-Braun, 2016).
While most organisms catabolizing urea use a urease (EC 3.5.1.5) to break it down into ammonia and carbon dioxide, chlorophytes and some yeasts use the ATP- and biotin-dependent urea amidolyase enzyme (EC 3.5.1.4.5). This enzyme possesses two activities that can be exhibited by two different proteins (in prokaryotes and green algae): urea carboxylase (EC 6.3.4.6) and allophanate hydrolase (EC 3.5.1.54) (Kanamori et al., 2005). The urea carboxylase possesses two separate catalytic domains: the biotin carboxylase domain, where a tethered biotin cofactor is carboxylated by bicarbonate with concomitant ATP cleavage, and the carboxyl-transferase domain, where a carboxyl group is transferred from carboxybiotin to urea, forming allophanate. Subsequently, allophanate is hydrolyzed to ammonia and CO2 by allophanate hydrolase (Strope et al., 2011; Lin et al., 2016), as shown by the following equations:
(1) urea + ATP + HCO3 → urea-1-carboxylate (also called allophanate) + ADP + Pi
(2) allophanate + H2O → 2NH3 + 2CO2
It has been suggested that urea carboxylase and allophanate hydrolase co-evolved in bacteria and, following horizontal gene transfer, subsequently fused into a single urea amidolyase protein in fungi (Strope et al., 2011), not necessarily more active than the two enzymes urea carboxylase and allophanate hydrolase (Lin et al., 2016). Similar to bacterial urease, the yeast urea amidolyase can contribute to virulence and kidney disease pathogenesis of Candida albicans (Navarathna et al., 2012).
This review summarizes what is known about urea transport and catabolism in cyanobacteria, and what can be inferred from the comparative analysis of the publicly available genome sequence of 308 cyanobacteria.
Results
Cyanobacteria Have the Capability to Grow on Various Nitrogen Sources
Attesting their metabolic diversity, cyanobacteria exhibit different abilities to grow on various nitrogen sources (N2, NH4, NO2, NO3, and urea). The highly-abundant marine cyanobacteria of the genus Prochlorococcus, which employs a chlorophyll a/b light harvesting antenna instead of phycobilisomes of other cyanobacteria, cannot grow on NO3. The Prochlorococcus species can be classified into two major groups depending on their growth requirements. The low-chlorophyll b/a-containing Prochlorococcus ecotypes, which are adapted to high light and predominate in nutrient-depleted surface-waters of the open ocean, such as the strains MED4, MIT9215, MIT9312, MIT9401, and AS9601, grow well on recycled N sources NH4 and urea, but not on NO2 (Moore et al., 2002). In contrast, high-chlorophyll b/a (low-light adapted) ecotypes MIT9303, MIT9313, NATL1A, and NATL2A, which thrive in the deep euphotic zone, can grow on NH4, urea and NO2 which is often abundant at these depths (Moore et al., 2002). Thus, high-b/a and low-b/a Prochlorococcus partition the water column with respect to depth because of differences in not only their light utilization capabilities, but also their N utilization capabilities (ability to grow on NO2).
The other widely-abundant marine cyanobacteria of the genus Synechococcus that thrive in surface waters (such as strains PCC7002, WH7805, WH8102, WH8103) are able to grow on four N sources: NH4 (preferred substrate), urea (slight growth decrease), NO3, and NO2, except for Synechococcus MIT S9220 which cannot grow on NO3 (Sakamoto et al., 1998; Collier et al., 1999; Moore et al., 2002). These findings indicate that N, in addition to light, plays a critical role in determining the dynamics between the ecotypes of closely related marine Prochlorococcus and Synechococcus genera, and contributes to their stability in the world’s oceans (Moore et al., 2002).
Many urease-endowed cyanobacteria can grow on urea as the sole nitrogen source (Collier et al., 1999) but a high-concentration of urea and/or prolonged cultivation on urea (≥10 mM) can be toxic to cyanobacteria. This finding was shown with Arthrospira PCC 8005 (edible cyanobacterium, Deschoenmaeker et al., 2017), Microcystis aeruginosa (fresh water cyanobacterium, Wu et al., 2015), Synechococcus PCC 7002 (costal cyanobacterium, Sakamoto et al., 1998) and Synechocystis PCC 6803 (euryhaline cyanobacterium, Veaudor et al., 2018). The cell death and color change (from blue-green to yellowish) triggered by the prolonged growth on urea could be due to lipid peroxidation, a phenomenon that increases in parallel with cell death and pigment oxidation (Sakamoto et al., 1998). By contrast, urease defective mutants of Synechococcus PCC7002 and Synechocystis PCC 6803 (inactivation of the ureC gene, see below) were not killed by prolonged incubation in the presence of a high urea concentration, demonstrating that urea-consumption driven by urease can become toxic (Sakamoto et al., 1998; Veaudor et al., 2018). Furthermore, the (marine) Synechococcus WH7803 strain and the (freshwater) Synechococcus PCC7942 strain cannot grow on urea and neither have urease activity (Collier et al., 1999). This toxicity is not likely due to a urea-catabolism elicited modification of the pH, as these studies were carried out in the presence of pH buffers. Similarly, in plants, nitrogen nutrition based only on urea leads to a reduction in growth (Witte, 2011).
Urease activity appeared to be constitutive in the phylogenetically-distant cyanobacteria Synechococcus PCC 7002 cells (Ludwig and Bryant, 2012), Synechocystis PCC 6803 (Veaudor et al., 2018), and Anabaena PCC 7120 (Valladares et al., 2002). In contrast, the marine cyanobacteria Synechococcus WH7805 and Synechococcus WH8112 have a much higher urease activity when grown on NO3– than on urea. In addition, Synechococcus WH7805 exhibits a twofold lower urease activity when grown on NH4+ compared to urea, whereas Synechococcus WH8112 has similar urease activities on NH4+ or urea. Urease expression increased in response to N deprivation in both of the Prochlorococcus strains MED4 and MIT9313 (Tolonen et al., 2006a). In cyanobacteria as diverse as Anabaena and Prochlorococcus, the urea transport genes are regulated by N availability via the global N transcription regulator NtcA (Valladares et al., 2002; Tolonen et al., 2006a), which also regulates urease activity in some cyanobacteria (Solomon et al., 2010).
The Urease (ureABCDEFG) and Urea Transport (urtABCDE) Genes Are Widely Distributed in Cyanobacteria
In bacteria, urea penetrates in cells by passive diffusion or ATP-requiring uptake systems. Three types of urea transport systems have been described: the Yut protein in Yersinia, the UreI protein in Helicobacter and the UrtABCDE proteins in cyanobacteria (Valladares et al., 2002; Sachs et al., 2006). UrtA is the lipid-anchored urea binding protein; UrtB and UrtC are integral membrane proteins and UrtD and UrtE are ATP-binding proteins.
Once inside the cells, urea can be catabolized by urease. Most bacterial urease is a trimer (UreABC)3 of two small (UreA and UreB) and one large (UreC, catalytic) subunits, while in plants the UreABC subunits are fused in a single protein (Witte, 2011). The urease enzyme complex is assembled by up to three (accessory) chaperone proteins (UreD, UreE, and UreF), and an intrinsically disordered (Palombo et al., 2017) GTPase (UreG) that transfers, likely with UreD (Farrugia et al., 2015), two nickel atoms into the urease metallocenter active site located in UreC (Figure 1). Mutation in either ureD, ureE, ureF, or ureG nearly abolish the activity of urease (Carter et al., 2009) The amino-acid sequences of the UreA, UreB, UreC, and to a slightly lesser extent of UreG, subunits are highly conserved, whereas UreD, UreE, and UreF sequences are more variable (Carter et al., 2009; Farrugia et al., 2015).
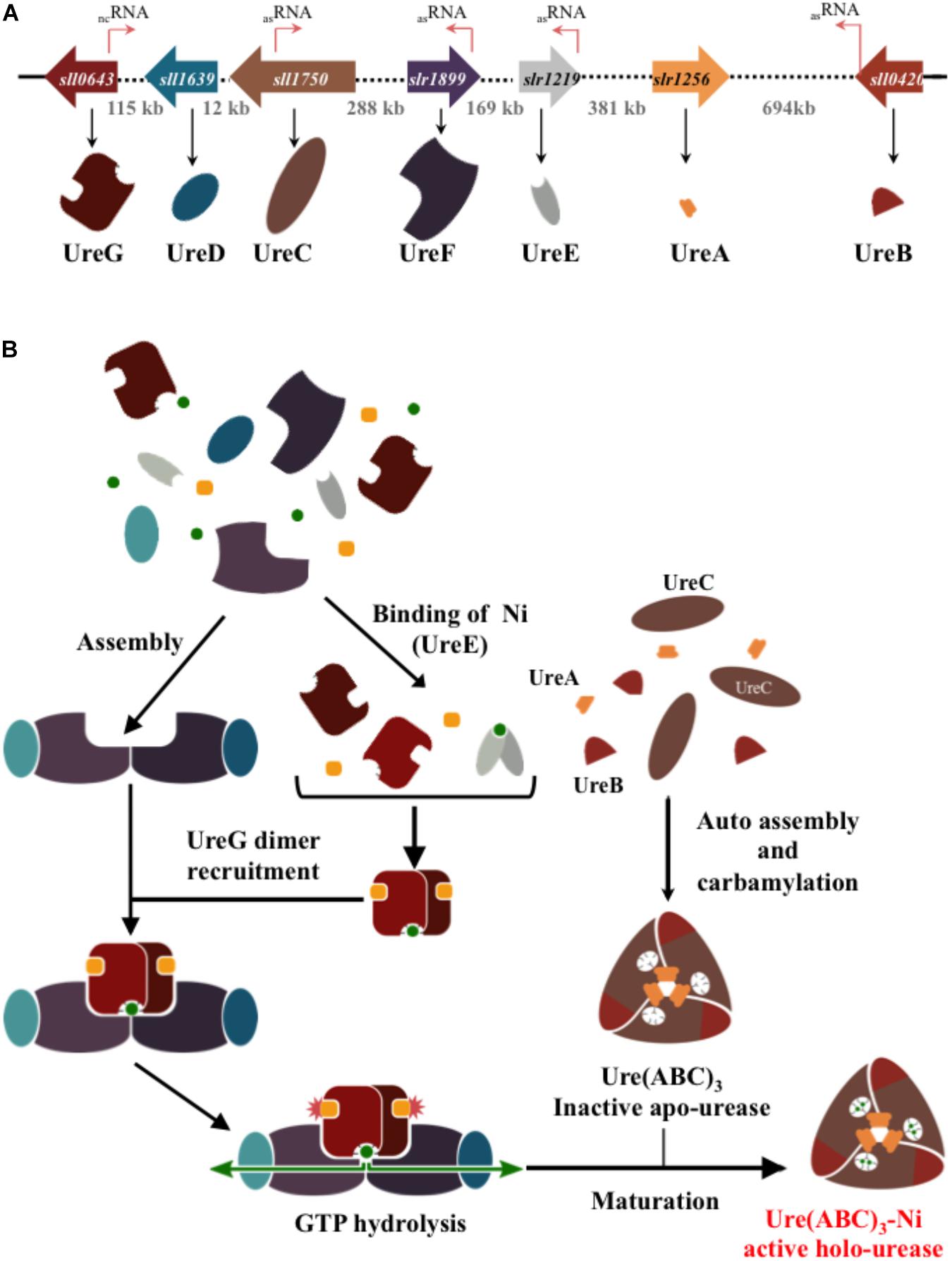
Figure 1. Schematic representation of the genes and proteins involved in the formation of the urease in the best-studied cyanobacterium Synechocystis PCC6803. (A) The genes encoding the urease catalytic subunits (UreABC) and the chaperones (UreDEFG) involved in its assembly are represented by large colored arrows indicating the direction of their transcription. The dotted lines and the numbers indicate the length of DNA (in Kb) separating the urease genes. The thin orange arrows stand for the small RNA (see cyanobase) of as yet unknow function. (B) Steps of the urease assembly and maturation process. The nickel ion and the guanoside-5′-triphosphate (GTP) are represented by green dots and orange squares, respectively.
Cyanobacteria were shown to possess urease activity many years ago (Berns et al., 1966). Like other urease, the cyanobacterial enzyme depends on nickel (Mackerras and Smith, 1986; Collier et al., 1999) and can be inactivated by a mutation in ureC (Sakamoto et al., 1998; Collier et al., 1999; Wu et al., 2015; Deschoenmaeker et al., 2017; Veaudor et al., 2018). Furthermore, in Anabaena sp. PCC 7120 and Synechocystis PCC 6803 inactivation of urtA, urtB, and/or urtE strongly decreases (97–98%) urea uptake down to a level that likely corresponds to passive urea uptake (Valladares et al., 2002; Sachs et al., 2006).
Using a comparative genomic approach, we have analyzed the publicly available genome sequence of 308 cyanobacteria. We found that urea transport and utilization genes (Table 1) are unevenly distributed in cyanobacteria (Supplementary Table S1), in agreement with their wide genome diversity (Shih et al., 2013; Cassier-Chauvat et al., 2016).
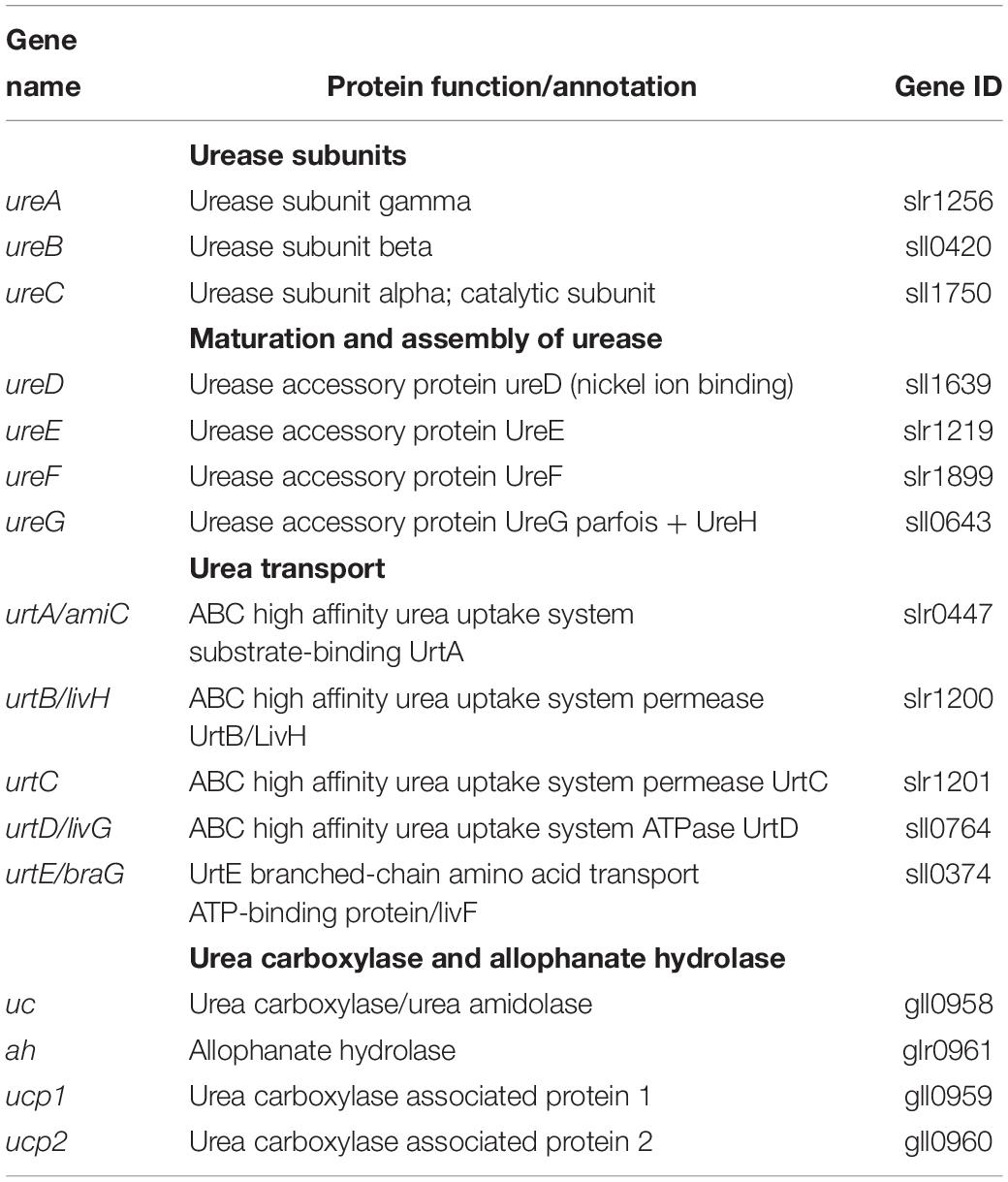
Table 1. Reference of the Synechocystis PCC 6803 (sll or slr) or Gloeobacter violaceus PCC 7421 (gll or glr) genes (from CyanoBase https://www.ncbi.nlm.nih.gov/protein) used to identify genes operating in urea acquisition and catabolism in other cyanobacteria.
The vast majority of the studied genomes (264 out of 308, i.e., 85%) possess all three genes encoding urease catalytic proteins (ureABC, Supplementary Table S2), mostly but not systematically accompanied with all five genes ureDEFG encoding urease accessory proteins (255 genomes, Supplementary Table S3). These findings show that urease is an abundant enzyme in cyanobacteria. The simultaneous presence (255 strains, Supplementary Table S3) or absence (40 strains, Supplementary Table S4) of all ureABCDEFG genes is consistent with the fact that both the urease catalytic proteins and the urease assembly proteins are essential to urease activity, as previously shown through a mutation in ureC or ureG (Sakamoto et al., 1998; Collier et al., 1999; Valladares et al., 2002; Veaudor et al., 2018). Thirteen cyanobacteria harbor an incomplete set of ureABCDEFG while ureDEFG, the urease chaperone genes, are more frequently absent than ureABC, the urease activity genes (Supplementary Table S5). This finding suggests that these cyanobacteria have either no active urease or they assemble it through an unknown process independent of some of the UreD, UreE, UreF, and UreG urease chaperones. The latter hypothesis is appealing in both Aphanocapsa montana BDHKU210001 and Lyngbya confervoides BDU141951 that possess a complete set of both ureABC (these genes are even duplicated in Lyngbya confervoides BDU141951) and urtABCDE (urea transport), in particular (Supplementary Table S5). In some cases, the absence of a gene might result from the fact that not all 308 cyanobacterial genomes are closed genomes. However, the lack of one or several of the ureABCDEFG genes has been observed in other (pathogenic) bacteria. Helicobacter pylori has only two urease-subunit genes, ureA and ureB (this ureA gene is a fusion of the ureAB genes occurring in other bacterial enzymes). These H. pylori ureAB genes are clustered with the following five downstream genes ureI (encoding a urea channel not encountered in other bacteria), ureE, ureF, ureG, and ureH (UreH is a nickel permease homologous to the UreD subunit of other bacteria). Thus, H. pylori has a UreH/UreF/UreG complex, not a UreD/UreF/UreG complex (Fong et al., 2013). Y. pestis harbors a complete urease locus (ureABC) and four accessory (ureEFGD) genes, but it has no urease activity because the nickel-incorporation gene ureD is disrupted (Carlini and Ligabue-Braun, 2016). Similarly, Bacillus subtilis has no urease accessory genes, suggesting that accessory proteins are not always required for in vivo urease activation or that genetically distinct cellular maturation factors are utilized in some cases (Carter et al., 2009).
Two lines of evidence showed that urease transport genes are also widely distributed in cyanobacteria. First, 235 strains harbor all urtABCDE genes, which are duplicated in the two strains of the genus Acaryochloris (Supplementary Table S6). Second, among the 255 cyanobacteria that possess all urease activity genes ureABCDEFG (Supplementary Table S3) 225 strains also have all urea transport genes urtABCDE (Supplementary Table S7). For example, among the 45 Prochlorococcus strains presently studied, 34 have ureABCDEFG and urtABCDE, including the species MED4, MIT9215, MIT9312, MIT9401, MIT9303, MIT9313, NATL1A, and NATL2A, which grow well on urea (Moore et al., 2002). The other 11 Prochlorococcus strains lack both ureABCDEFG and urtABCDE (Supplementary Table S4), as mentioned earlier for MIT9211, MIT8515, and CCMP1375, the growth of which was not tested on urea (Solomon et al., 2010). Furthermore, 235 cyanobacteria have all urtABCDE genes.
A minority of the cyanobacteria endowed with a complete set of urease genes ureABCDEFG possess no urea transport genes urtABCDE (23 strains, Supplementary Table S8). This observation suggests that the urease of these cyanobacteria likely operates in the detoxification of internally generated urea (recycling of nitrogen lost during the urea-generating catabolism of arginine). It is also possible that urea diffuses through aquaporins as observed in other organisms (Li and Wang, 2014). Conversely, several cyanobacteria harbor urtABCDE but an incomplete set of ureABCDEFG (10 strains, Supplementary Table S5), suggesting that urtABCDE could operate in the transport of not only urea but also other nutrients.
Occurrence of Duplication of Either Urea Transport (urtABCDE) or Urease Activity (ureABCDEFG) Genes but Not Both
Twenty-nine cyanobacteria harbor two copies of one or several genes ureABC (urease activity), ureDEFG (urease assembly) and/or urtABCDE (urea transport). In 13 cases, the duplication concerned one or two, but not all three sets of ureABC, ureDEFG and urtABCDE (Supplementary Table S9). Seven cyanobacteria possess two copies of ureABC, three strains have two copies of ureDEFG, while six strains harbor two copies of urtABCDE. Two cyanobacteria possess two copies of all urease assembly and activity genes ureABCDEFG (Chamaesiphon minutus PCC 6605 and Mastigocoleus testarum BC00), while one strain has two copies of both ureABC and urtABCDE (Xenococcus sp. PCC 7305). By contrast, we found no cyanobacterium with a duplication of ureDEFG and urtABCDE (Supplementary Table S9). In the future, it will be interesting to study whether the products encoded by these duplicated genes have distinct biochemical properties or are differently regulated.
The occurrence of multiple copies of urease genes is not unprecedented in prokaryotes. For instance, the beta-proteobacterium Nitrosospira strain NpAV possesses two copies of ureC (Koper et al., 2004; Carter et al., 2009). Furthermore, three Helicobacter species (i.e., H. mustelae, H. acinonychis, and H. felis) harbor two sets of urease genes (Fong et al., 2013). The first set contains the complete urease gene cluster (ureA1B1EFGH) while the second set ureA2B2 (50% identical to ureA1 and ureB1, respectively) encodes the UreA2B2 enzyme that does not require urease accessory proteins to be active. The ureA1B1EFGH cluster encoding the Ni-containing urease is induced by nickel ions while the ureA2B2 cluster encoding an iron-containing enzyme is up regulated by Fe (it is downregulated by nickel). These findings are consistent with the observation that these Helicobacter species are associated with carnivores that eat an iron-rich food depleted in nickel (Carter et al., 2009).
We found no duplication of all three gene-sets ureABC, ureDEFG, and urtABCDE (Supplementary Table S9), suggesting that a very high transport and catabolism of urea would be toxic. This hypothesis is consistent with the findings that phylogenetically-distant cyanobacteria were killed by a prolonged growth on high urea concentration, namely Arthrospira PCC 8005 (Deschoenmaeker et al., 2017), Microcystis aeruginosa (Wu et al., 2015), Synechococcus PCC 7002 (Sakamoto et al., 1998), and Synechocystis PCC 6803 (Veaudor et al., 2018).
Distribution of the Genes Encoding the Other Urea Catabolytic Enzymes: Urea Carboxylase and Allophanate Hydrolase Genes Are Less Frequent Than the Urease Genes
Several lines of evidence suggest that the urea carboxylase and/or allophanate hydrolase enzymes have little importance for the photoautotrophic metabolism of cyanobacteria.
First, among the 308 cyanobacteria presently studied 237 have a complete set of urease genes ureABCDEFG whereas they lack one or both the uc and ah genes encoding the urea carboxylase and allophanate hydrolase (Supplementary Table S10). Second, a few cyanobacteria have uc (19 strains) and/or ah (13 strains), while only nine cyanobacteria possess both uc and ah (Supplementary Table S11), which together, could allow a urease-independent urea catabolysis, as shown in other organisms (Strope et al., 2011; Lin et al., 2016). Third, four of these nine cyanobacteria endowed with uc and ah are actually devoid of the ureABCDEFG urease genes, namely Calothrix sp. PCC 7507, Gloeomargarita lithophora D10, Gloeobacter kilaueensis JS1, and Gloeobacter violaceus PCC 7421 (Supplementary Table S11). It will be interesting in the future to test if these cyanobacteria are able to grow on urea as the sole nitrogen source to verify whether their urea carboxylase and allophanate hydrolase are truly active. Because, Gloeobacter and Gloeomargarita are regarded to have diverged early from other cyanobacteria (de Vries and Archibald, 2017; Ponce-Toledo et al., 2017), it is possible the urea carboxylase and allophanate hydrolase enzymes appeared in cyanobacteria before urease. Furthermore, interestingly, Calothrix sp. PCC 7507, Gloeomargarita lithophora D10, Gloeobacter kilaueensis JS1, and Gloeobacter violaceus PCC 7421 not only lack the genes encoding the Ni-containing urease but also the genes encoding the Ni-Fe hydrogenase. This finding suggests that these cyanobacteria may live in nickel poor environments.
Few Cyanobacteria Possess the Complete Panoply of Urea Transport (urtABCDE) and Catabolism (ureABCDEFG, uc, and ah) Genes
Only five cyanobacterial strains possess the complete panoply of urea transport (urtABCDE) and catabolism (ureABCDEFG, uc, and ah) genes (Supplementary Table S12), namely Cyanothece PCC7425, Microcoleus vaginatus FGP-2, Microcoleus vaginatus PCC 9802, and Synechococcus PCC 7502. This finding suggests that these five cyanobacteria growing in their natural environment frequently uses urea as a nitrogen source. We have verified that Cyanothece PCC 7425 can grow not only on nitrate and ammonium, but also on urea as the sole nitrogen source.
Several Cyanobacterial Genera Display a Highly Heterogeneous Panel of Urea Acquisition and Catabolism Genes
Among the four Calothrix strains, three possess the ureABCDEFG genes but neither urtABCDE, nor uc and ah, whereas Calothrix PCC 7507 lacks ureABCDEFG and urtABCDE but possess both uc and ah (Supplementary Table S13). The eight studied Cyanothece strains also have different gene panoplies. Cyanothece PCC 7425 phylogenetically distant for the other Cyanothece has all urea transport and catabolism genes (ureABCDEFG, urtABCDE, uc, and ah, see Figure 2), whereas the two Cyanothece strains PCC 7424 and PCC 7822 (Supplementary Table S1 and Supplementary Figure S1) possess ureABCDEFG and urtABCDE, but neither uc nor ah, and the five other strains ATCC 51472, ATCC 51142, CCY 0110, PCC 8801, and PCC 8802 lack all ureABCDEFG, urtABCDE, uc and ah genes (Supplementary Tables S1, S4). These observations are consistent with the findings that Cyanothece ATCC51142, PCC 8801, and PCC 8802, which fix atmospheric N2 in aerobiosis, are less dependent of an organic nitrogen source than Cyanothece PCC 7425, which fix N2 only in anaerobiosis (Bandyopadhyay et al., 2011).
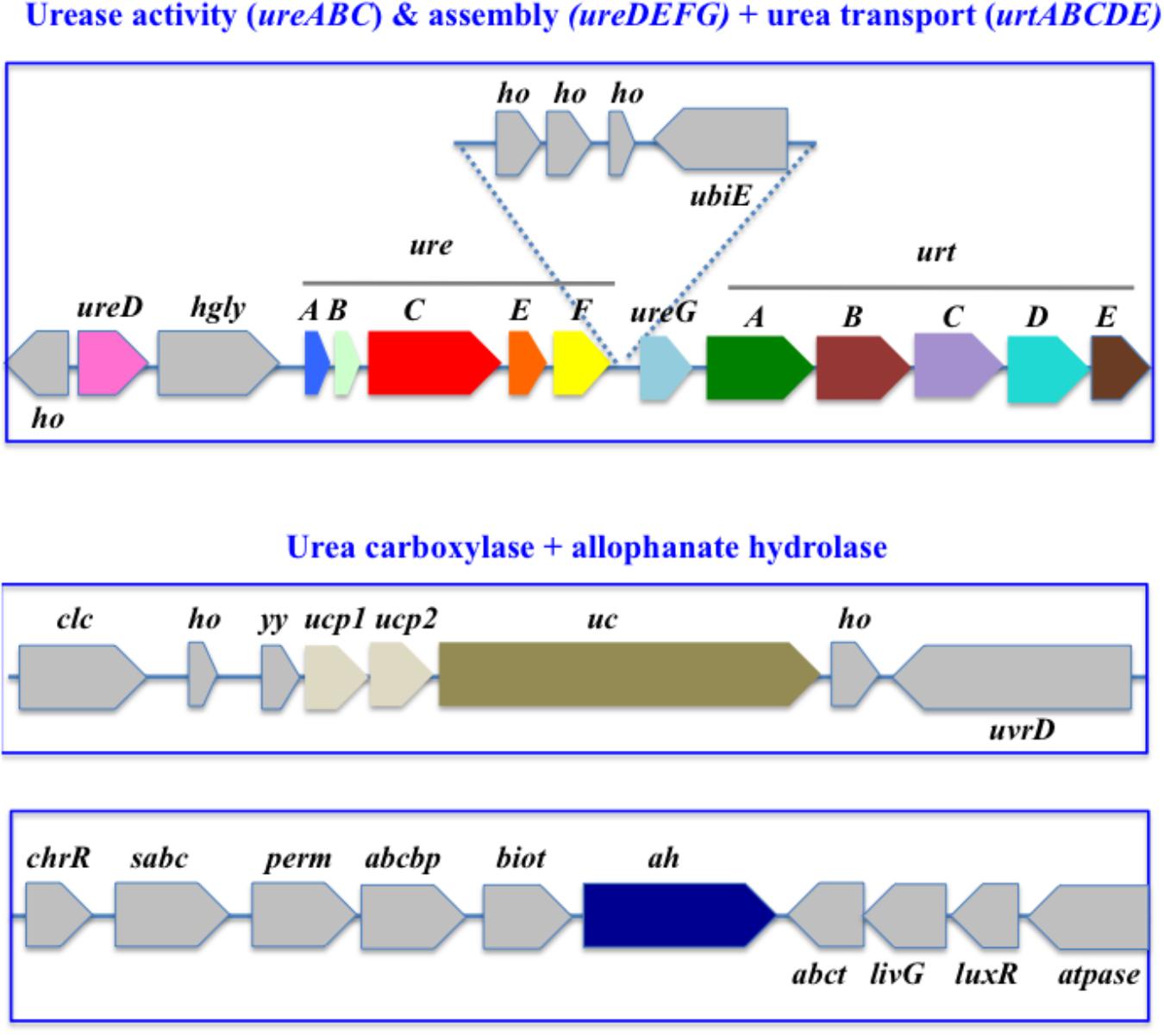
Figure 2. Organization of the urea acquisition and catabolism genes in Cyanothece PCC 7425. The genes are shown as colored boxes pointing in the direction of their transcription. Gray boxes correspond to genes not known to be involved in urea metabolism or transport. abcbp, ABC transporter ATP-binding protein; biot, biotin-[acetyl-CoA-carboxylase] synthetase; chrR, anti-ECF regulator; clc, chloride channel core; ubiE, ubiquinone biosynthesis protein; ho, hypothetical protein; sabc, sulfonate binding ABC transporter protein; ucp1, urea carboxylase associated protein 1; ucp2, urea carboxylase associated protein 2; uvrD, DNA helicase; yy, YKKC-YXXD leader in MBGD; perm, ABC transporter permease; abct, amino acid ABC transporter ATPase; livG, leucine/isoleucine/valine transporter ATP-binding subunit; luxR, LuxR family transcriptional regulator; atpase, ATPase.
The 12 Leptolyngbya strains display a different panel of the urea acquisition and catabolism, in agreement with phylogenetic analysis showing that Leptolyngbya strains are found in different clusters. Eight strains have all the ureABCDEFG and urtABCDE genes, but one of them has the uc gene but not ah (Leptolyngbya boryana PCC 6306) whereas two of them have ah, but not uc (Leptolyngbya sp. PCC 7375 and Leptolyngbya sp. Heron Island J). Furthermore, two of these eight strains harboring the ureABCDEFG and urtABCDE genes have a duplication of all the urtABCDE genes and posess uc (Leptolyngbya sp. NIES-2104 and Leptolyngbya sp. NIES-3755), whereas one strain has two copies of urtAB genes but possess neither uc nor ah (Leptolyngbya sp. PCC 6406). Leptolyngbya valderiana BDU 20041 has neither ureFG nor urtABCDE genes. Furthermore, among the 45 Prochlorococcus species analyzed, 34 strains have ureABCDEFG and urtABCDE, whereas 11 strains lack both ureABCDEFG and urtABCDE (Supplementary Table S14). In contrast the Microcystis genera is homogenous since all 14 Microcystis aeruginosa strains possess all ureABCDEFG and urtABCDE genes, but neither uc nor ah.
Several Cyanobacteria Lack the Whole Panoply of Urea Transport and Catabolism Genes
Thirty-six cyanobacteria lack the complete set of urea transport (urtABCDE) and catabolism (ureABCDEFG, uc, and ah) genes (Supplementary Table S14). This is the case of all four strains of the genus Crocosphaera watsonii (marine unicellular diazotrophic cyanobacteria) and all four strains of the genus Gastranaerophilaceae (belonging to the new phylum of Melainabacteria that are not able to perform photosynthesis.
The same is true for the symbiotic (marine) cyanobacterium UCYN-A, in agreement with the fact that it possesses the smallest genome (1.44 Mb), and for the two strains of Richelia intracellularis. Similarly, 11 strains of the Prochlorococcus genus, which is known to possess a small-genome, are totally devoid of the urea transport (urtABCDE) and catabolism (ureABCDEFG, uc, and ah) genes. By contrast, the 34 other Prochlorococcus strains have ureABCDEFG and urtABCDE (but neither uc nor ah). Also similarly, five Synechococcus strains lacks all urea transport and catabolism genes, including the two closely-related models Synechococcus elongatus PCC7942 and Synechococcus elongatus PCC6301 and the fast-growing strain Synechococcus elongatus UTEX 2973 (doubling time as short as 1.5–1.9 h) that are closely related (there are only 55 single nucleotide differences separating the two strains (Ungerer et al., 2018; Supplementary Table S15 model strains).
Genomic Context of Urea Transport and Catabolism Genes in Model Cyanobacteria: The Urea Transport Genes (urtABCDE) Are Clustered Whereas the Urease Genes (ureABCDEFG) Are Often Scattered
In bacteria, the three urease structural genes ureABC are often clustered with those genes encoding urease associated proteins ureCDEFG, but their number and order differ among species (Carter et al., 2009). In K. aerogenes, ureABC are flanked by ureDEFG in a ureDABCEFG gene cluster. The same gene order, ureDABCEFG, occurs in the Beta-proteobacterium Nitrosospira NpAV and the Gamma-proteobacterium Nitrosococcus oceani (Koper et al., 2004). By contrast, many bacteria position ureD after ureG, like that occurring in the thermophilic Bacillus sp. TB-90 (Carter et al., 2009).
We performed a gene neighborhood survey of the genes operating in urea acquisition and assimilation in cyanobacteria emphasizing on phylogenetically distant model cyanobacteria (Supplementary Table S15). We focused our attention on several phylogenetically-distant cyanobacteria (Supplementary Table S16) because they are presently well studied thanks to their powerful genetics (for example Synechocystis PCC 6803, Synechococcus PCC 7002, and Synechococcus PCC 7942). Furthermore, they should also be increasingly investigated in the near future because of their interesting natural properties (Supplementary Table S16) and the likely possibility that they could be manipulated with a broad-host-range of RSF1010 plasmids that have been shown to replicate in various cyanobacteria since first being reported (Marraccini et al., 1993; Mühlenhoff and Chauvat, 1996; Tolonen et al., 2006a, b; Araki et al., 2013; Taton et al., 2014).
Our gene neighborhood survey revealed that these selected cyanobacteria harbor diverse panoplies of urea acquisition and utilization genes that define five groups. The first group comprises 13 cyanobacteria, including the extensively-studied species Synechocystis PCC6803 (Figure 3) and Synechococcus PCC 7002 (Figure 4), which possess a single copy of the ureABCDEFG and urtABCDE, but neither uc nor ah (Supplementary Table S15 and Supplementary Figure S1). The second group of cyanobacteria (five species including the well-studied strain Synechococcus PCC 7942) has no urea acquisition and catabolic genes. The third, fourth and fifth groups are each defined by a single model cyanobacterium, as follows. Cyanothece PCC 7425 (Figure 2) possesses the complete panoply of the studied genes (Supplementary Table S12). Gloeobacter violaceus PCC 7421 has only uc + ah, the importance of which can be studied in this host or in Cyanothece PCC 7425. Acaryochloris marina MBIC11017 possesses ureABCDEFG + a duplicated copy of urtABCDE (Supplementary Figure S1).
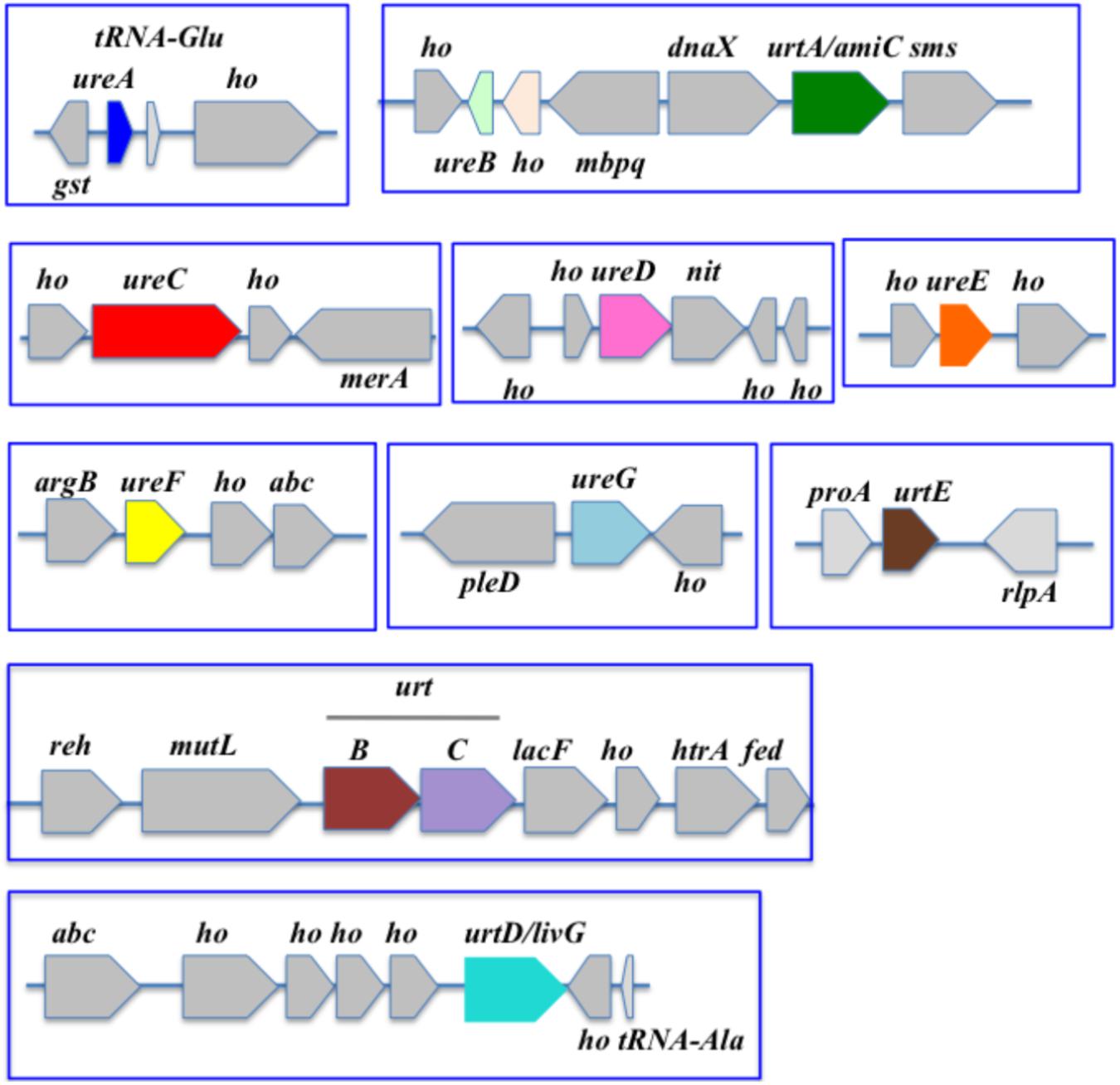
Figure 3. Organization of the urea acquisition and catabolism genes in Synechocystis PCC 6803. The genes are shown as colored boxes pointing in the direction of their transcription. Gray boxes correspond to genes not known to be involved in urea metabolism or transport. abc, ABC transporter; argB, acetylglutamate kinase; abc, ABC transporter; dnaX, DNA polymerase III subunit delta; fed, ferredoxin; gst, glutathione-S-transferase; ho, hypothetical protein; htrA, serine protease; lacF, lactose ABC transporter permease; mbpq, methyl-6-phytyl 1,4-hydroquinone methyltransferase; merA, mercuric reductase; mutL, DNA mismatch repair; nit, nitrilase; pleD, pleD protein; proA, gamma-glutamyl phosphate reductase; radA homolog; reh, rehydrin; rlpA, rare lipoprotein A; sms, DNA repair protein.
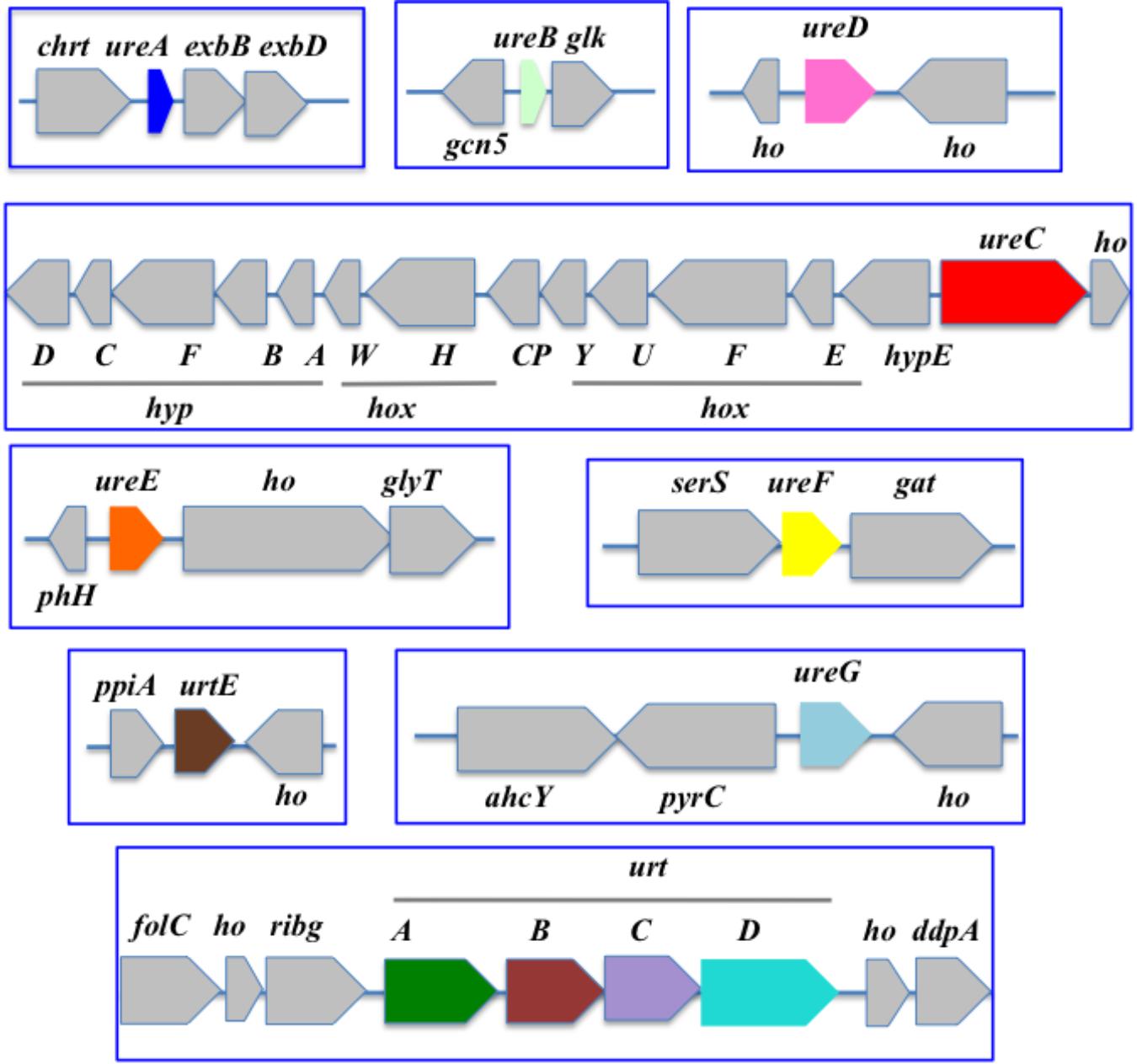
Figure 4. Organization of the urea acquisition and catabolism genes in Synechococcus PCC 7002. The genes are shown as colored boxes pointing in the direction of their transcription. Gray boxes correspond to genes not known to be involved in urea metabolism or transport. ahcY, adenosyl homocysteinase; chrt, chromate transport; ddpA, peptide ABC transporter substrate binding protein; exbB and exbD, biopolymer transport; folC, folypolyglytamte; gat, glutamine amidotransferase. gcn5, N-acetyltransferase GCN5; glk, glucokinase; glyT, glycosyl transferase; ho, hypothetical protein; hoxEFGHU, Ni-Fe hydrogenase subunits; hoxW, HoxH-specific protease; hypABCDEF, Ni-Fe hydrogenase chaperone subunits; phh, phosphohydrolase; ppia, peptidylprolyl isomerase A; pyrC, dihydroorotase; ribg, ADP-ribosylglycohydrolase; serS, serine-tRNA ligase synthase.
Our gene neighborhood survey revealed that ureA, ureB, and ureC were often contiguous, as were ureE, ureF, and ureG. Furthermore urease genes are often clustered with those genes encoding the ATP-dependent uptake system (urtABCDE) (Valladares et al., 2002; Wu et al., 2015).
A closer look at the gene organization of these cyanobacteria shows the following findings. The chlorophyll d-containing symbiotic cyanobacterium Acaryochloris marina MBIC11017 has the particularity of having two spatially distant urtABCDE clusters (Supplementary Figure S1). One of them is located downstream of the ureFG cluster. The ureE gene is located far away, including from the locus encompassing the ureDA upstream of ureB and ureC in that order.
In the edible (Deschoenmaeker et al., 2017) cyanobacterium Arthrospira PCC 8005 (Supplementary Figure S1), urtABCDE and ureDABC are displayed, in that order, in two non-neighboring clusters, away from ureEF and ureG.
In Nostoc (Anabaena) PCC7120 (Figure 5), the urea transport genes (urtABCDE) are clustered in that order, whereas the urease (ureABCDEFG) is displayed in two loci. One locus comprises the cluster ureDAB and ureC separated by an unknown gene, while the other locus contains ureEFG in that order, as previously observed (Valladares et al., 2002).
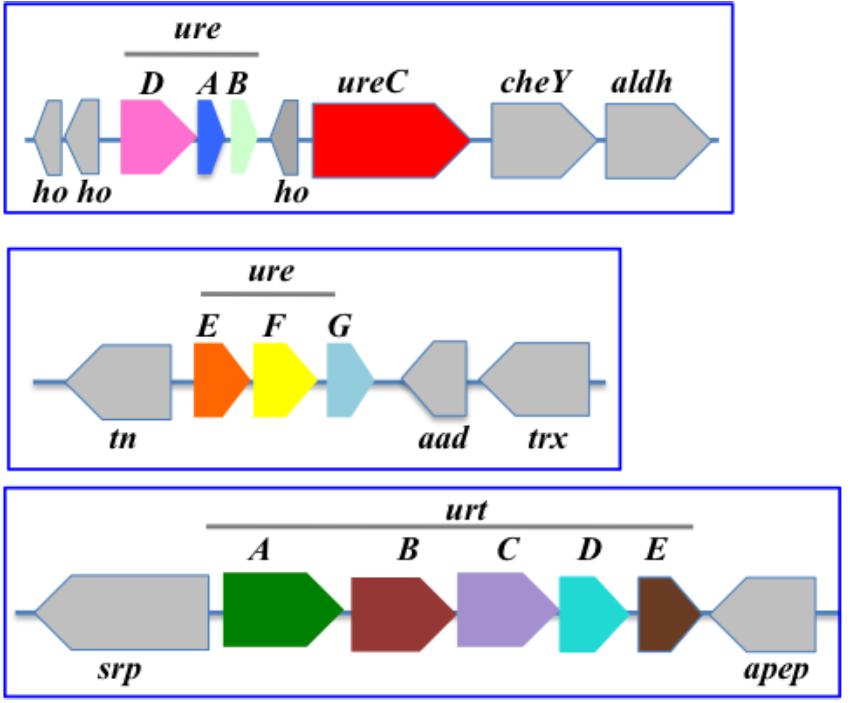
Figure 5. Organization of the urea acquisition and catabolism genes in Nostoc PCC 7120. The genes are shown as colored boxes pointing in the direction of their transcription. Gray boxes correspond to genes not known to be involved in urea metabolism or transport. Each blue box corresponds to a genomic region. aad, acetoacetate decarboxylase; aldH, aldehyde dehydrogenase; apep, aminopeptidase; ho, hypothetical protein; cheY, chemotaxis protein CheY; tn, tranposase; srp, signal recognition particle; trx, thioredoxin.
In Cyanothece PCC 7425, which has the complete panoply of urea transport and catabolism genes, the urease (ureABCDEFG) and urea transport (urtABCDE) genes are located in the same chromosomal region (Figure 2) where ureD precedes the two clusters ureABCDEF and ureG-urtABCDE. In contrast, the uc and ah genes are not close to each other. Interestingly, the uc genes is clustered with two genes, ucp1 and ucp2, which presumably code for two urea carboxylase associated proteins. A similar situation occurs in several other cyanobacteria such as Chamaesiphon minutus PCC 6605, Gloeocapsa PCC 7428, Oscillatoria PCC 6505, Oscillatoria nigro viridis PCC 7112, Microcoleus vaginatus FGP-2, Pseudanabaena biceps PCC7429, and Synechococcus PCC7502.
In Cyanothece PCC 7822, the urease (ureABCDEFG) and urea transport (urtABCDE) genes define two non-neighboring clusters, ureABGDEFC and urt ABCDE (Supplementary Figure S1).
In Gloeobacter violaceus PCC 7421, which has neither ureABCDEFG nor urtABCDE, the ah and uc genes are located in the same region but in opposite directions (Supplementary Figure S1).
In the well-studied heterocyst-forming filamentous strain Anabaena PCC 7120, ureDAB are clustered upstream of ureC, far away from both the ureEFG and the urtABCDE clusters (Figure 5).
In the marine Prochloroccus strains MED4 and MIT9313, the ure and urt genes are located in the same locus comprising two opposite clusters urtEDCBAGFE and ureDABC (Supplementary Figure S1).
In the marine strain Synechococcus WH8102 (Supplementary Figure S1), the ure and urt genes are located mostly in one locus comprising two opposite clusters urtEDCBAureGFE and ureDABC (as observed in the Prochloroccus strains MED4 and MIT9313) far away from urtA2 the second copy of urtA (Supplementary Figure S1).
In the costal strain Synechococcus PCC7002 (Figure 4), the urt genes are displayed in two non-neighboring regions harboring the urtABCD cluster and urtE, respectively. In contrast, the ure genes are scattered in different loci. One of them harbors ureC in an opposite direction from the clustered genes hox and hyp encoding hydrogenase, the activity of which requires Ni like urease. It is possible that the proximity of the genes encoding Ni-dependent enzymes facilitates their Ni-dependent expression.
In Synechocystis PCC 6803, the ureABCDEFG and urtABCDE genes all are completely scattered (Figure 3).
Finally, in Thermosynechococcus elongatus BP1, the urea transport genes (urtABCDE) are clustered in that order, whereas the urease (ureABCDEFG) are scattered (Supplementary Figure S1).
Discussion
Cyanobacteria are widely diverse photosynthetic microorganisms that play a crucial role for the biosphere (production of biomass and oxygen for the biosphere) and have great biotechnological interests (photoproduction of chemicals). As of yet, however, research has mostly focused on few model strains with well-established genetics, while cyanobacterial diversity is insufficiently explored. Furthermore, most studies on cyanobacteria are carried out with cells growing on nitrate (NO3) while we show in this review that most cyanobacteria have the potential to grow on urea [(NH2)2CO] as the nitrogen source. This finding is important because urea is cheaper than nitrate or ammonium, and it is often present in natural waters because it is produced by heterotrophic organisms and it is released by the sediments of urea-fertilized agricultural soils (Glibert et al., 2006; Belisle et al., 2016). Thus, in the future it will be interesting to use cyanobacteria to couple the photosynthetic production of chemicals and the removal of the urea pollutant, in economically viable waste-to-biorefinery industrial systems.
In this study, we report what is known about urea transport and catabolism in cyanobacteria, and what can be inferred from the comparative analysis of the publicly available genome sequence of 308 cyanobacteria.
We show that most cyanobacteria possess the genes encoding one or several enzymes, urease, urea carboxylase and allophanate hydrolase, which catabolize urea [(NH2)2CO] into NH3 (ammonia) and CO2 that can be re-injected into the cell metabolism.
Urease is a very frequent enzyme in cyanobacteria. The vast majority of the 308 studied genomes (264 out of 308, i.e., 85%) possess all three genes encoding urease catalytic proteins (ureABC, Supplementary Table S2), mostly accompanied with all four genes ureDEFG encoding urease accessory proteins (255 genomes, Supplementary Table S3). These findings are consistent with the fact that both the urease catalytic and assembly proteins are essential to urease activity, as previously shown through mutations in ureC or ureG (Sakamoto et al., 1998; Collier et al., 1999; Valladares et al., 2002; Veaudor et al., 2018). A large number of these 255 cyanobacteria possessing ureABCDEFG (Supplementary Table S3) also have all urea transport genes urtABCDE (225 strains, Supplementary Table S7), suggesting that they grow in urea-containing environments. Interestingly, we found that the urtABCDE genes are frequently clustered, whereas ureABCDEFG are often scattered (Figures 2–5 and Supplementary Figure S1). A minority of cyanobacteria possessing all urease genes (ureABCDEFG) lack all urea transport genes urtABCDE (23 strains, Supplementary Table S8) thereby suggesting that their urease operates in the detoxification of internally-generated urea (for example by the catabolism of arginine). Conversely, several cyanobacteria harboring urtABCDE lack one or several ureABCDEFG genes (10 strains, Supplementary Table S5), suggesting that urtABCDE could operate in the transport of not only urea but also of other nutrients.
Two lines of evidence suggest that the urea carboxylase and/or allophanate hydrolase enzymes, encoded by the uc and ah genes, have less importance than urease for the metabolism of cyanobacteria. First, only five of the 308 cyanobacteria presently studied have the complete panoply of urea transport (urtABCDE) and catabolism (ureABCDEFG, uc and ah) genes. Second, only four cyanobacteria that possess both the uc and ah genes (Supplementary Table S11) presumably involved in urea catabolysis are actually lacking the ureABCDEFG urease genes. These cyanobacteria are Calothrix PCC 7507, Gloeomargarita lithophora D10, Gloeobacter kilaueensis JS1, Gloeobacter violaceus PCC 7421 (Supplementary Table S11). It will be interesting to assay if these cyanobacteria can really grow on urea as the sole nitrogen source, to test whether the uc and ah genes truly operates in urea catabolism. Interestingly, these four cyanobacteria that lack the genes encoding the Ni-requiring urease enzyme also lack the genes encoding the Ni-Fe hydrogenase, suggesting that they may live in nickel-poor environments. Because the cyanobacteria of the genera Gloeobacter and Gloeomargarita are regarded as having diverged early from other cyanobacteria (de Vries and Archibald, 2017; Ponce-Toledo et al., 2017) it is possible that the urea carboxylase and allophanate hydrolase enzymes appeared in cyanobacteria before urease, and were subsequently lost by various cyanobacteria, most of which possess urease genes, to catabolize urea by the ATP-independent urease instead of the ATP- and HCO3-consuming enzymes urea carboxylase and allophanate hydrolase. This assumption is supported by the phylogenetic analysis of the distribution of the genes encoding the urease (ureC subunit, panel A), allophanate hydrolase (B) and urea carboxylase (C) proteins in various organisms (Supplementary Figure S2). Among the 264 cyanobacterial UreC amino-acids sequences considered in this study, 39 were selected so as to maximize the coverage and representation of all five sub-sections of the cyanobacterial phylum proposed by other workers (Shih et al., 2013). All these genes appeared to be distributed in the cyanobacterial clade, likely ruling out horizontal gene transfer events, which if occurs, would blur their distribution. Similarly, all cyanobacterial uc (19) and ah (13) genes were found to be distributed within the cyanobacterial clade, likely in absence of horizontal-gene-transfer introduction of foreign uc and/or ah genes into some cyanobacteria. However, one of the two archaeal ah genes included in this analysis (Haloterrigena daqingensis) appeared to cluster with the ah genes of Synechococcus PCC7502, Calothrix PCC7507 and Pseudanabaena PCC7367 thereby suggesting that a relatively ancient horizontal gene transfer event of ah may have occurred from cyanobacteria to Archaea (Supplementary Figure S2). Also, interestingly, the ah gene of Ferrovibrio sp. and Magnetospirillium marisnigri representative of the Rhodospirillaceae family of non-sulfur purple bacteria, appeared to cluster with the deep-rooting cyanobacteria. This finding suggests that the allophanate hydrolase enzyme was present in an “ancient” photosynthetic ancestor, and it was then frequently lost in cyanobacteria. Thirty-six cyanobacteria lack the complete set of urea transport (urtABCDE) and catabolism (ureABCDEFG, uc, and ah) genes (Supplementary Table S14), in agreement with the fact that they possess a small genome.
So far, little is known concerning the regulation of the urea transport and catabolism genes. Several studies performed with different cyanobacteria showed that the urease genes are not regulated by changes in nitrogen availabilities or exposure to various stress (see http://cyanoexpress.sysbiolab.eu/). In Synechocystis PCC6803, the urt genes appears to be strongly regulated (negatively) in response to H2O2, a high concentration of iron and cadmium, whereas the expression of ureC encoding the urease catalytic subunit is almost unchanged (Houot et al., 2007). In Synechococcus PCC 7002, the expression of the urease genes were not affected by changes in nitrogen source or nitrogen starvation (Ludwig and Bryant, 2012). In Microcystis aeruginosa exposed to nitrogen limitation, all urt genes were significantly upregulated, whereas ure transcript levels were not affected (Harke and Gobler, 2013). Furthermore, interestingly, the cyanobacterial PII signal transduction protein was recently shown to operate in the control of the uptake of ammonium, nitrate and urea. First, PII controls ammonium uptake by interacting with the Amt1 ammonium permease. Second, PII mediates the ammonium- and dark-induced inhibition of nitrate uptake by interacting with the NrtC and NrtD subunits of the nitrate/nitrite transporter NrtABCD. PII regulates urea uptake by interacting with the UrtE subunit. The deregulation of urea uptake in a PII deletion mutant causes ammonium excretion when urea is provided as nitrogen source.
Conclusion
Using a comparative genomic approach, we have analyzed the publicly available genome sequence of 308 cyanobacteria, the photosynthetic prokaryotes that are increasingly studied for basic and applied science. We found that most cyanobacteria harbor all genes encoding the urea transport (urtABCDE) and the urea catabolytic enzyme urease (ureABCDEFG), in agreement with the capacity of the few tested cyanobacteria to grow on urea as the sole nitrogen source. This finding has major implications for the future engineering of effective cyanobacterial factories for an economically viable production of chemicals coupled to the consumption of urea, which is cheaper than nitrate (the usual nitrogen source) and is frequently present in natural or waste waters. Other cyanobacteria have ureABCDEFG or urtABCDE, indicating that urease also operates in the detoxification (recycling of carbon and nitrogen) of internally generated urea and that urtABCDE could operate in the transport of not only urea but also of other nutrients. Three cyanobacteria of the genera Gloeobacter and Gloeomargarita, which likely diverged early from other cyanobacteria, have the genes encoding the urea carboxylase (uc) and allophanate hydrolase (ah) enzymes that sequentially catabolize urea. This finding indicates that the urea carboxylase and allophanate hydrolase enzymes may have appeared in cyanobacteria before urease. The diverse metabolic pathways of urea transport and decomposition of cyanobacteria may contribute to differences in their role in the biogeochemistry of urea, as well as in the role that urea plays in the physiology and ecology of cyanobacteria.
Author Contributions
FC conceived the project, wrote the manuscript, and agreed to serve as the author responsible for contact and ensures communication. TV and CC-C retrieved all genomic information, conceived the figures and the tables, and read out the manuscript. TV, CC-C, and FC analyzed the data.
Funding
TV received a Ph.D. fellowship from CEA (Irtelis).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Owing to space limitations, it was not possible to cite all research papers relevant to the presented subject. We sincerely apologize to those authors whose work we could not include.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02052/full#supplementary-material
FIGURE S1 | Organization of the urea acquisition and catabolism genes in various model cyanobacteria.
FIGURE S2 | Phylogenetic distribution of the genes encoding the urease (ureC subunit, A), allophanate hydrolase (B) and urea carboxylase (C) proteins in various organisms.
TABLE S1 | Distribution of genes involved urea transport and catabolism in 308 cyanobacterial species.
TABLE S2 | List of the 264 species of cyanobacteria harboring ureABC genes encoding the urease catalytic subunits.
TABLE S3 | List of the 255 species of cyanobacteria harboring ureABC genes encoding the urease catalytic subunits and the ureDEFG genes encoding the urease accessory proteins.
TABLE S4 | List of the 40 species of cyanobacteria that possess no ureABCDEFG genes in their genome.
TABLE S5 | List of the 13 species of cyanobacteria that possess an incomplete set of ure genes in their genome.
TABLE S6 | List of the 235 species of cyanobacteria harboring urtABCD genes involved in the urea transport.
TABLE S7 | List of the 225 species of cyanobacteria harboring simultaneously ureABCDEFG genes and urtABCD genes in their genome.
TABLE S8 | List of the 23 species of cyanobacteria harboring ureABCDEFG genes without urtABCDE genes in their genome.
TABLE S9 | List of the 237 species of cyanobacteria harboring a duplication of ureABCDEFG genes and/or urtABCDE genes in their genome.
TABLE S10 | List of the 237 species of cyanobacteria harboring ureABCDEFG genes but no urea carboxylase (uc) and allophanate hydrolase (ah) encoding genes in their genome.
TABLE S11 | List of the nine species of cyanobacteria harboring both urea carboxylase (uc) and allophanate hydrolase (ah) encoding genes in their genome.
TABLE S12 | List of the nine species of cyanobacteria harboring both urea carboxylase (uc) and allophanate hydrolase (ah) encoding genes in their genome.
TABLE S13 | Only five species of cyanobacteria harbor all ureABCDEFG, urtABCD uc and ah) encoding genes in their genome.
TABLE S14 | List of 36 species of cyanobacteria with no ure and urt genes nor any uc and ah genes in their genome.
TABLE S15 | Distribution of ureABCDEFG, urtABCDE, uc and ah genes in model cyanobacteria.
TABLE S16 | Properties of model cyanobacteria with a sequenced genome.
References
Araki, M., Shimada, Y., Mimuro, M., and Tsuchiya, T. (2013). Establishment of the reporter system for a thylakoid-lacking cyanobacterium, Gloeobacter violaceus PCC 7421. FEBS Open Bio. 3, 11–15. doi: 10.1016/j.fob.2012.11.003
Archibald, J. M. (2009). The puzzle of plastid evolution. Curr. Biol. 19, 81–88. doi: 10.1016/j.cub.2008.11.067
Bandyopadhyay, A., Elvitigala, T., Welsh, E., Stöckel, J., Liberton, M., Min, H., et al. (2011). Novel metabolic attributes of the genus cyanothece, comprising a group of unicellular nitrogen-fixing cyanobacteria. mBio 2:e00214-11. doi: 10.1128/mBio.00214-11
Belisle, B. S., Steffen, M. M., Pound, H. L., Watson, S. B., DeBruyn, J. M., Bourbonniere, R. A., et al. (2016). Urea in lake erie: organic nutrient sources as potentially important drivers of phytoplankton biomass. J. Great Lakes Res. 42, 599–607. doi: 10.1016/j.jglr.2016.03.002
Berns, D. S., Holohan, P., and Scott, E. (1966). Urease activity in blue-green algae. Science 152, 1077–1078. doi: 10.1126/science.152.3725.1077
Carlini, C. R., and Ligabue-Braun, R. (2016). Ureases as multifunctional toxic proteins: a review. Toxicon 110, 90–109. doi: 10.1016/j.toxicon.2015.11.020
Carter, E. L., Flugga, N., Boer, J. L., Mulrooney, S. B., and Hausinger, R. P. (2009). Interplay of metal ions and urease. Metallomics 1, 207–721. doi: 10.1039/b903311d
Cassier-Chauvat, C., and Chauvat, F. (2018). Cyanobacteria: Wonderful Microorganisms for Basic and Applied Research. Hoboken: Wiley.
Cassier-Chauvat, C., Veaudor, T., and Chauvat, F. (2016). Comparative genomics of DNA recombination and repair in cyanobacteria: biotechnological implications. Front. Microbiol. 7:1809. doi: 10.3389/fmicb.2016.01809
Collier, J. L., Brahamsha, B., and Palenik, B. (1999). The marine Cyanobacterium Synechococcus sp. WH7805 requires urease (urea amidohydrolase, EC 3.5.1.5) to utilize urea as a nitrogen source: molecular-genetic and biochemical analysis of the enzyme. Microbiology 145, 447–459. doi: 10.1099/13500872-145-2-447
de Vries, J., and Archibald, J. M. (2017). Endosymbiosis: did plastids evolve from a freshwater Cyanobacterium? Curr. Biol. 27, R103–R122. doi: 10.1016/j.cub.2016.12.006
Deschoenmaeker, F., Bayon-Vicente, G., Sachdeva, N., Depraetere, O., Cabrera Pino, J. C., Leroy, B., et al. (2017). Impact of different nitrogen sources on the growth of Arthrospira sp. Bioresour. Technol. 237, 78–88. doi: 10.1016/j.biortech.2017.03.145
Farrugia, M. A., Wang, B., Feig, M., and Hausinger, R. P. (2015). Mutational and computational evidence that a nickel-transfer tunnel in UreD Is used for activation of Klebsiella aerogenes Urease. Biochemistry 54, 6392–6401. doi: 10.1021/acs.biochem.5b00942
Fong, Y. H., Wong, H. C., Yuen, M. H., Lau, P. H., Chen, Y. W., and Wong, K. B. (2013). Structure of UreG/UreF/UreH complex reveals how urease accessory proteins facilitate maturation of Helicobacter pylori urease. PLoS Biol. 11:e1001678. doi: 10.1371/journal.pbio.1001678
Glibert, P. M., Harrison, J., Heil, C., and Seitzinger, S. (2006). Escalating worldwide use of urea - A global change contributing to coastal eutrophication. Biogeochemistry 77, 441–463. doi: 10.1007/s10533-005-3070-3075
Harke, M. J., and Gobler, C. J. (2013). Global transcriptional responses of the toxic Cyanobacterium, Microcystis aeruginosa, to nitrogen stress, phosphorus stress, and growth on organic matter. PLoS One 8:e69834. doi: 10.1371/journal.pone.0069834
Houot, L., Floutier, M., Marteyn, B., Michaut, M., Picciocchi, A., Legrain, P., et al. (2007). Cadmium triggers an integrated reprogramming of the metabolism of Synechocystis PCC6803, under the control of the Slr1738 regulator. BMC Genomics 8:350. doi: 10.1186/1471-2164-8-350
Kanamori, T., Kanou, N., Kusakabe, S., Atomi, H., and Imanaka, T. (2005). Allophanate hydrolase of Oleomonas sagaranensis involved in an ATP-dependent degradation pathway specific to urea. FEMS Microbiol. Lett. 245, 61–65. doi: 10.1016/j.femsle.2005.02.023
Knoot, C. J., Ungerer, J., Wangikar, P. P., and Pakrasi, H. B. (2018). Cyanobacteria: promising biocatalysts for sustainable chemical production. J. Biol. Chem. 293, 5044–5052. doi: 10.1074/jbc.R117.815886
Koper, T. E., El-Sheikh, A. F., Norton, J. M., and Klotz, M. G. (2004). Urease-encoding genes in ammonia-oxidizing bacteria. Appl. Environ. Microbiol. 70, 2342–2348. doi: 10.1128/AEM.70.4.2342-2348.2004
Li, C., and Wang, W. (2014). Urea transport mediated by aquaporin water channel proteins. Sub Cell. Biochem. 73, 227–265. doi: 10.1007/978-94-017-9343-8_14
Lin, Y., Boese, C. J., and St. Maurice, M. (2016). The urea carboxylase and allophanate hydrolase activities of urea amidolyase are functionally independent. Protein Sci. 25, 1812–1824. doi: 10.1002/pro.2990
Lopes, F. C., Dobrovolska, O., Real-Guerra, R., Broll, V., Zambelli, B., Musiani, F., et al. (2015). Pliable natural biocide: jaburetox is an intrinsically disordered insecticidal and fungicidal polypeptide derived from jack bean urease. FEBS J. 282, 1043–1064. doi: 10.1111/febs.13201
Ludwig, M., and Bryant, D. A. (2012). Acclimation of the global transcriptome of the Cyanobacterium Synechococcus sp. strain PCC 7002 to nutrient limitations and different nitrogen sources. Front. Microbiol. 3:145. doi: 10.3389/fmicb.2012.00145
Mackerras, A. H., and Smith, G. D. (1986). Urease activity of the Cyanobacterium Anabaena cylindrica. J. Gen Microbiol. 132, 2749–2752. doi: 10.1099/00221287-132-10-2749
Marraccini, P., Bulteau, S., Cassier-Chauvat, C., Mermet-Bouvier, P., and Chauvat, F. (1993). A conjugative plasmid vector for promoter analysis in several cyanobacteria of the genera Synechococcus and Synechocystis. Plant Mol. Biol. 23, 905–909. doi: 10.1007/BF00021546
Moore, L. R., Post, A. F., Rocap, G., and Chisholm, S. W. (2002). Utilization of different nitrogen sources by the marine cyanobacteria Prochlorococcus and Synechococcus. Limnol. Oceanogr. 47, 989–996. doi: 10.4319/lo.2002.47.4.0989
Mühlenhoff, U., and Chauvat, F. (1996). Gene transfer and manipulation in the thermophilic Cyanobacterium Synechococcus elongatus. Mol. Gen. Genet. MGG 252, 93–100. doi: 10.1007/BF02173209
Navarathna, D. H., Lionakis, M. S., Lizak, M. J., Munasinghe, J., Nickerson, K. W., and Roberts, D. D. (2012). Urea amidolyase (DUR1,2) contributes to virulence and kidney pathogenesis of candida albicans. PLoS One 7:e48475. doi: 10.1371/journal.pone.0048475
Palombo, M., Bonucci, A., Etienne, E., Ciurli, S., Uversky, V. N., Guigliarelli, B., et al. (2017). The relationship between folding and activity in UreG, an intrinsically disordered enzyme. Sci. Rep. 7:5977. doi: 10.1038/s41598-017-06330-6339
Ponce-Toledo, R. I., Deschamps, P., López-García, P., Zivanovic, Y., Benzerara, K., and Moreira, D. (2017). An early-branching freshwater Cyanobacterium at the origin of plastids. Curr. Biol. 27, 386–391. doi: 10.1016/j.cub.2016.11.056
Ramsundar, P., Guldhe, A., Singh, P., and Bux, F. (2017). Assessment of municipal wastewaters at various stages of treatment process as potential growth media for Chlorella sorokiniana under different modes of cultivation. Bioresour. Technol. 227, 82–92. doi: 10.1016/j.biortech.2016.12.037
Rutherford, J. C. (2014). The Emerging Role of Urease as a General Microbial Virulence Factor. PLoS Pathogens 10:e1004062. doi: 10.1371/journal.ppat.1004062
Sachs, G., Kraut, J. A., Wen, Y., Feng, J., and Scott, D. R. (2006). Urea transport in bacteria: acid acclimation by gastric Helicobacter spp. J. Mem. Biol. 212, 71–82. doi: 10.1007/s00232-006-0867-867
Sakamoto, T., Delgaizo, V. B., and Bryant, D. A. (1998). Growth on urea can trigger death and peroxidation of the Cyanobacterium Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 64, 2361–2366.
Sarayu, K., Iyer, N. R., and Murthy, A. R. (2014). Exploration on the biotechnological aspect of the ureolytic bacteria for the production of the cementitious materials - A review. Appl. Biochem. Biotechnol. 172, 2308–2323. doi: 10.1007/s12010-013-0686-680
Schmalstig, A. A., Benoit, S. L., Misra, S. K., Sharp, J. S., and Maier, R. J. (2018). Noncatalytic antioxidant role for Helicobacter pylori urease. J. Bacteriol. 200:e00124-18. doi: 10.1128/JB.00124-118
Schopf, J. W. (2011). The paleobiological record of photosynthesis. Photosyn. Res. 107, 87–101. doi: 10.1007/s11120-010-9577-9571
Shih, P. M., Wu, D., Latifi, A., Axen, S. D., Fewer, D. P., Talla, E., et al. (2013). Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. U.S.A. 110, 1053–1058. doi: 10.1073/pnas.1217107110
Solomon, C. M., Collier, J. L., Berg, G. M., and Glibert, P. M. (2010). Role of urea in microbial metabolism in aquatic systems: a biochemical and molecular review. Aquati. Microb. Ecol. 59, 67–88. doi: 10.3354/ame01390
Soo, R. M., Hemp, J., Parks, D. H., Fischer, W. W., and Hugenholtz, P. (2017). On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 355, 1436–1440. doi: 10.1126/science.aal3794
Strope, P. K., Nickerson, K. W., Harris, S. D., and Moriyama, E. N. (2011). Molecular evolution of urea amidolyase and urea carboxylase in fungi. BMC Evol. Biol. 11:80. doi: 10.1186/1471-2148-11-80
Su, J., Jin, L., Jiang, Q., Sun, W., Zhang, F., and Li, Z. (2013). Phylogenetically diverse urec genes and their expression suggest the urea utilization by bacterial symbionts in marine sponge xestospongia testudinaria. PLoS One 8:e64848. doi: 10.1371/journal.pone.0064848
Sun, T., Li, S., Song, X., Diao, J., Chen, L., and Zhang, W. (2018). Toolboxes for cyanobacteria: recent advances and future direction. Biotechnol. Adv. 36, 1293–1307. doi: 10.1016/j.biotechadv.2018.04.007
Taton, A., Unglaub, F., Wright, N. E., Zeng, W. Y., Paz-Yepes, J., Brahamsha, B., et al. (2014). Broad-host-range vector system for synthetic biology and biotechnology in cyanobacteria. Nucleic Acids Res. 42:e136. doi: 10.1093/nar/gku673
Tolonen, A. C., Aach, J., Lindell, D., Johnson, Z. I., Rector, T., Steen, R., et al. (2006a). Global gene expression of Prochlorococcus ecotypes in response to changes in nitrogen availability. Mol. Syst. Biol. 2:53. doi: 10.1038/msb4100087
Tolonen, A. C., Liszt, G. B., and Hess, W. R. (2006b). Genetic manipulation of Prochlorococcus strain MIT9313: green fluorescent protein expression from an RSF1010 plasmid and Tn5 transposition. Appl. Environ. Microbiol. 72, 7607–7613. doi: 10.1128/AEM.02034-2036
Ungerer, J., Lin, P. C., Chen, H. Y., and Pakrasi, H. B. (2018). Adjustments to photosystem stoichiometry and electron transfer proteins are key to the remarkably fast growth of the Cyanobacterium Synechococcus elongatus UTEX 2973. mBio 9:e02327-17.
Valladares, A., Montesinos, M. L., Herrero, A., and Flores, E. (2002). An ABC-type, high-affinity urea permease identified in cyanobacteria. Mol. Microbiol. 43, 703–715. doi: 10.1046/j.1365-2958.2002.02778.x
Vasconcelos Fernandes, T., Shrestha, R., Sui, Y., Papini, G., Zeeman, G., Vet, L. E. M., et al. (2015). Closing domestic nutrient cycles using microalgae. Environ. Sci. Technol. 49, 12450–12456. doi: 10.1021/acs.est.5b02858
Veaudor, T., Cassier-Chauvat, C., and Chauvat, F. (2018). Overproduction of the cyanobacterial hydrogenase and selection of a mutant thriving on urea, as a possible step towards the future production of hydrogen coupled with water treatment. PLoS One 13:e0198836. doi: 10.1371/journal.pone.0198836
Verseux, C., Baqué, M., Lehto, K., De Vera, J. P. P., Rothschild, L. J., and Billi, D. (2016). Sustainable life support on Mars - The potential roles of cyanobacteria. Intern. J. Astrobiol. 15, 65–92. doi: 10.1017/S147355041500021X
Witte, C. P. (2011). Urea metabolism in plants. Plant Sci. 180, 431–438. doi: 10.1016/j.plantsci.2010.11.010
Wu, X., Yan, Y., Wang, P., Ni, L., Gao, J., and Dai, R. (2015). Effect of urea on growth and microcystins production of microcystis aeruginosa. Bioresou. Technol. 181, 72–77. doi: 10.1016/j.biortech.2015.01.035
Keywords: urea transport and catabolism, urease, urea carboxylase, allophanate hydrolase, cyanobacteria
Citation: Veaudor T, Cassier-Chauvat C and Chauvat F (2019) Genomics of Urea Transport and Catabolism in Cyanobacteria: Biotechnological Implications. Front. Microbiol. 10:2052. doi: 10.3389/fmicb.2019.02052
Received: 28 June 2019; Accepted: 20 August 2019;
Published: 04 September 2019.
Edited by:
Wendy Schluchter, University of New Orleans, United StatesReviewed by:
Peter Lindblad, Uppsala University, SwedenMartin Hagemann, University of Rostock, Germany
Copyright © 2019 Veaudor, Cassier-Chauvat and Chauvat. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Franck Chauvat, franck.chauvat@cea.fr