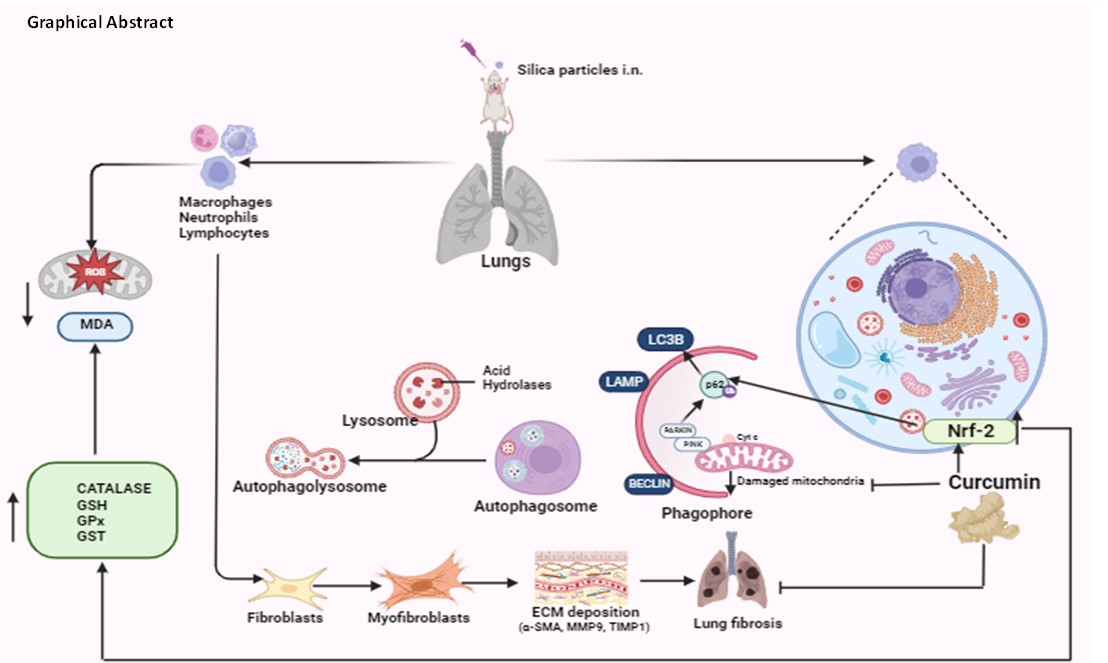
As an occupational illness, silicosis is brought on by extended exposure to dust that contains an excessive amount of respirable silica particles. It is the most prevalent and dangerous form of occupational disease in the world (3), but as of yet, no particular remedial medication is being used in clinical settings. The body's innate immune cells, lung macrophages play active phagocytic, immunological, and secretory functions. To investigate the role of curcumin on autophagy, including mitophagy in silica induced oxidative stress and lung damage, silicosis model was established using swiss mice by administering sonicated sterile silica suspension every alternate day for 35 days. In this study, curcumin and dexamethasone are effective in ameliorating silicosis. Superoxide (O2−) radical is catalyzed by SOD to produce H2O2 or regular molecular oxygen (O2). CAT and GSH-PX then catalyze the produced H2O2 to generate H2O [41]. Lipid peroxidation produces MDA, a byproduct which is measured to assess lipid peroxidation. Here, we showed that curcumin might increase CAT, GSH, GST and GSH-PX activities to enhance cells' ability to remove ROS. It has been shown to have two functions: it scavenges ROS and functions as an antioxidant both directly and indirectly (42). Lipid peroxidation and ROS levels were found to be significantly elevated in silicosis group whereas the curcumin and dexamethasone groups showed significant decrease in the level of MDA (Fig. 1a-f).
The H&E staining results demonstrated marked inflammatory cell infiltration around both the alveolar septa and bronchioles in the model group. Additionally, disruptions in bronchiolar structure, thickening of alveolar walls, and the formation of glomeruli were observed. In contrast, curcumin and dexamethasone groups exhibited patterns closer to those observed in the control group (Fig. 2). Dexamethasone, being a corticosteroid, is given clinically to asthma patients but its role in silicosis is not known. Alveolar macrophages and inflammatory cells play an integral role in the pathogenesis of silicosis [43].
Silica particles, when inhaled, deposit in the respiratory tract, subsequently penetrating and causing damage to the alveolar structure. Prolonged ingestion of silica particle hinders the macrophages' capacity to eliminate them and induces mitochondria-dependent apoptosis in alveolar macrophages [44]. In cytospin preparations, differential cell counts revealed morphology of BALF cells. The bronchoalveolar lavage fluid (BALF) in the silicosis group exhibited higher macrophage numbers, whereas these numbers were significantly diminished in the curcumin and dexamethasone groups resembling control group (Fig. 3). This finding was further confirmed by immunostaining of BALF cells with macrophages and neutrophils specific markers i.e., CD 11b and Gr-1 and the results were very close to cytospin images where curcumin groups showed significant reduction in macrophages as well as neutrophils (Fig. 5).
Pulmonary fibrosis emerges as a key pathogenic feature in the later stages of silicosis. The inflammation triggered by silica exposure is recognized for instigating an overactive tissue repair response, leading to the deposition of collagen and the irregular restructuring of lung architecture. Despite the absence of a singular mechanism to comprehensively elucidate the fibrotic progression in silicosis, it is widely acknowledged that the condition involves these intricate processes [45]. Plasma levels of MMP9 is involved in the onset and development of silicosis, and it may be useful for early diagnosis of silicosis [46]. Degrading extracellular matrix, matrix metalloproteinases (MMPs) are linked to airway injury in chronic inflammatory lung illnesses such as cystic fibrosis. The presence of silica particles induces the generation of pro-inflammatory mediators, free radicals, and metalloproteases at the lesion site, attracting an increased number of inflammatory cells, including neutrophils. These cells play a role in the lung fibrosis process [47]. The silicosis group showed a notable increase in the measured MMP9 level through gelatin zymography, while the administration of curcumin significantly reduced this level (Fig. 5a). This was further validated by protein expressions of MMP9 and TIMP1 which were similar to their activity profiles studied in gelatin zymography. Increased expression of α-SMA, a primary contributor to pulmonary fibrosis was found in silica group. However, this upregulation was significantly suppressed in curcumin and dexamethasone administered group (Fig. 5b).
Nrf-2 has the potential to regulate cellular defense against oxidants by influencing the expression of various cytoprotective genes associated with xenobiotic metabolism, antioxidant and anti-inflammatory responses. A variety of antioxidant response, element-dependent genes are regulated by Nrf-2, which controls their baseline and stimulated expressions. This regulates the genes associated with oxidative stress and has a direct impact on the homeostasis of reactive oxygen and nitrogen species [48]. Under normal condition, Nrf2 is strongly bound to Keap1 and constantly ubiquitylated by the Cul3 E3 ubiquitin ligase and subsequently degraded by the proteasome. In response to stress, Keap1 is inactivated, resulting in Nrf2 stabilization. However, Keap1–Cul3–Rbx1 E3 ubiquitin ligase or β-TrCP–Skp1–Cul1–Rbx1 E3 ubiquitin ligase is responsible for the cellular degradation of Nrf-2 in response to stress, depending on whether the pathway is keap-1 dependent or independent [49]. The Nrf2 level was found significantly higher in the curcumin and dexamethasone group than that of the silica group. keap-1 expression in the curcumin group was similar to silica group supporting keap-1 independent regulation of Nrf-2 activation (Fig. 6). It is possible that other proteins have phosphorylated the serine/threonine residues in the Nrf-2 Neh6 domain, disrupting the Nrf-2 keap-1 connection and promoting keap-1 independent Nrf-2 activation. Keap1-independent activation of Nrf-2 has been linked to a number of protein kinase pathways, including phosphatidylinositol 3-kinase (PI3K), MAPKs, PKC, glycogen synthase kinase-3 (GSK-3), c-Jun, and N-terminal kinase (JNK), which requires further exploration [50–51].
Mitophagy, or mitochondrial selective autophagy, has been primarily linked to the putative protein kinase 1 (PINK1) and PARK2 pathway that is triggered by phosphatase and tensin homolog (PTEN). When stress causes the mitochondrial membrane to depolarize, PINK1 stabilizes on the outer membrane of the mitochondria, which attracts PARK2 to the mitochondria. To monitor the autophagy mechanism in silicosis group after pretreatment with curcumin and dexamethasone, we investigated the LC3BII/I ratio and p62. Intriguingly, curcumin apparently reduced PINK and PARK protein expression in 35 days model group. Research on the function of mitophagy in type II alveolar epithelial cells and fibroblasts from patients with IPF has produced contradictory results [12, 52, 53, 54]. In comparison to normal patients, it was discovered that the mitochondria isolated from IPF alveolar macrophages showed higher immunoreactive PINK1 and Parkin expression. The rise was correlated with an increase in LC3-II expression in alveolar macrophages, which suggests autophagosome production is a sign of mitophagy. In our study, it was found that the PINK and PARKIN levels were higher in silicosis group whereas the levels were significantly lower in curcumin and dexamethasone groups (Fig. 7). To examine mitophagy or the accumulation of damaged mitochondria visually electron microscopy was used. Aberrant mitochondria were found in silicosis group as compared to the control group.
The administration of curcumin led to a recovery in mitophagy levels, resulting in a reduction in the count of abnormal mitochondria and the manifestation of cytoplasmic vacuolization (Fig. 8). When the mitochondrial permeability transition pore (MPT) opens as a result of mitochondrial depolarization, Cyt c, which is positioned between the inner and outer membranes of the mitochondrial pores, enters the cytoplasm from the MPT and binds to receptors connected to the endoplasmic reticulum to further activate the mitochondrial apoptosis pathway [55]. Following treatment with curcumin, the Cyt c level is decreased as compared to the silicosis model group, exhibiting a positive correlation with PINK1 and PARKIN levels. This suggests that the mitigation of the mitochondrial apoptosis pathway triggered by reactive oxygen species (ROS) forms the basis of the reduction in Cyt c expression, aligned with similar observations in existing research [56].
{kind=link}