Target fragment insertion into the P. yezoensis genome
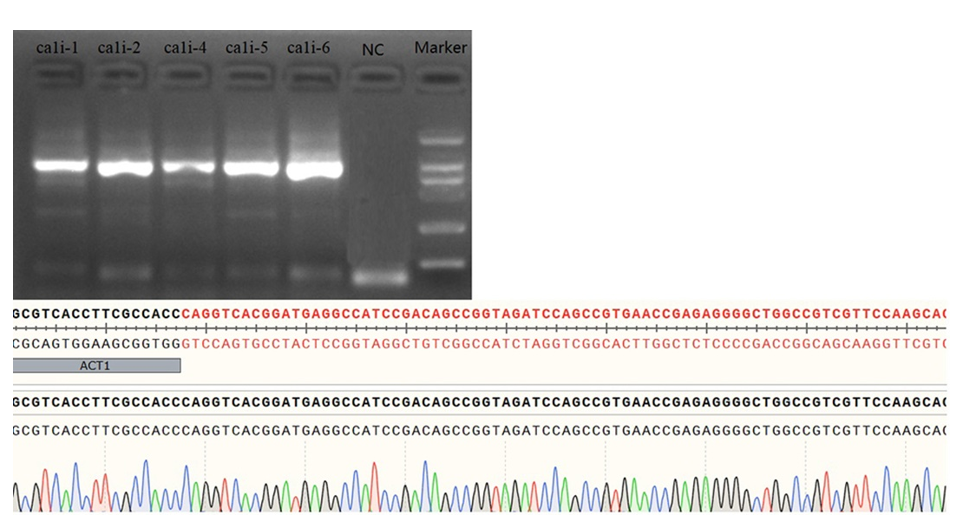
After being subjected to hygromycin B stress for 8 weeks, five hygromycin B-resistant strains of P. yezoensis were isolated and named PyβCA1i-1, PyβCA1i-2, PyβCA1i-4 to 6. Genomic PCR was conducted to verify the presence of the Pycali expression cassette in the P. yezoensis genome, and sequencing was performed to confirm the presence of the expression plasmid with the targeting gene sequence but not in the WT (Fig. 1). To assess the stability of the expression cassettes in the P. yezoensis genome across generations, the mutants were re-examined after 6 months of culturing through monospores germination (Supplemental Fig. 1). The results indicated that the target fragment was stably inserted into the P. yezoensis genome and could be transferred through reproduction.
Changes in the RNA and protein levels of PyβCA1i mutants
The RNA and protein levels of the mutants and WT were detected by RT‒qPCR and Western blotting, respectively. The RNA level of mutants and WT showed a significant difference, with the βCA1 gene expression in Pyβca1i lines decreasing approximately 5-fold compared to WT (Fig. 2a). Immunoblot analysis of total protein extracts using an anti-CA1 monoclonal antibody and a RbcL polyclonal antibody revealed a positive band at approximately 28 KD and 54 KD, respectively, which matched the predicted molecular weight of the PyβCA1 and RbcL protein. RbcL was used as a standard protein. The protein level of βCA1 was lower in the mutants than in the WT (Fig. 2b). On the other hand, immunoblot results also indicated that βCA1 was abundant in the 20 µg loading total protein.
Morphological characteristics and biomass of PyβCA1i strains
Under NC conditions (2 mM NaHCO3), the general morphological and developmental phenotypes of ca1i-1, ca1i-2 and ca1i-6 mutants were observed. The mutants, especially ca1i-1 and ca1i-2, were visibly smaller in length and width than the WT after 20 days of growth under the same conditions. The phenotype of ca1i-6 was similar to that of the WT (Fig. 2c). Based on the observed differences in RNA and protein levels, as well as morphological characteristics of the mutants, ca1i-1, ca1i-2 and ca1i-6 were selected for further investigation.
To investigate the physiological responses of the βca1i mutants to HC (8 mM NaHCO3), these three strains were cultured under NC and HC conditions, with WT as a control.
The pH changes were similar between the ca1i mutants and WT under NC conditions, but the highest pH reached during 14 days of cultivation was different. The pH increased from 8.2 ± 0.02 to 8.9 ± 0.02 for the ca1i mutants and to 9.1 ± 0.02 for WT. However, the pH changes in HC medium were notably faster than in NC, increasing to 9.2 ± 0.02 for ca1i-1 and ca1i-2 on the 14th day, whereas it increased to 9.4 ± 0.02 on the 12th day and then decreased to 9.2 ± 0.02 for WT on the 14th day, and the change in pH in ca1i-6 was similar to that in WT (Fig. 3a).
The POE rate of mutants was lower than that of WT under both DIC conditions. At pH 8.2 ± 0.02, the POE rates were 1.4 and 2.2 µmol ml-1·gFW-1·min-1 for the mutants and WT, respectively, under NC conditions. Under HC conditions, the POE rate increased to 2.0 and 3.0 µmol ml-1·gFW-1·min-1 for mutants and WT, respectively (Fig. 3b).
The respiration rates of these three mutant lines were all higher than that of the WT under both NC and HC conditions, with rates of 0.6 µmol ml-1·gFW-1·min-1 for the mutant lines and 0.34 for the WT (Fig. 3b).
Decreased intracellular carbonic anhydrase activity (CAint) and extracellular CA activity (CAext) compared to WT under NC and HC conditions
The basic function of CA is to catalyze the reversible interconversion of CO2 and HCO3-. Compared to WT, the CAint and CAext were decreased in ca1i mutants when cultured under NC and HC conditions (Fig. 3c). Specifically, the CAint of WT was 2.1- and 2.4-fold higher than that of ca1i-1 and ca1i-2, respectively, while being 1.5-fold higher than that of ca1i-6. However, with increasing DIC concentration, the CAint of both mutants and WT decreased, with mutants showing a reduction of up to 40% compared to the NC condition. WT decreased only approximately 16% compared to NC conditions.
Under HC conditions, the CAext of both mutants and WT increased, with the CAext of WT increasing by 48% and the CAext of ca1i-1 and ca1i-2 increasing by approximately 70% compared to NC conditions. However, the CAext of mutants remained lower than that of WT, and the CAext of WT was 1.7-1.5-fold higher than that of ca1i-1 and ca1i-2 under NC and HC conditions, respectively (Fig. 3d). The CAext and CAint activity of transgenic line ca1i-6 was similar to that of WT, with no apparent differences between them under the two DIC conditions (Fig. 3d).
Interference of chloroplast βCA1 impaired Ci affinity and resulted in relatively higher residual [HCO3-] in the mutant medium
The half-saturation constant for DIC (K1/2[DIC]) represents the subject’s DIC affinity, with a higher value indicating lower affinity for DIC. The K1/2[DIC] in artificial seawater-grown βCA1i lines was approximately 4.27-fold higher than that in WT lines (Fig. 3e).
On the 7th day, the bicarbonate concentration in the medium varied differently between the mutants and WT (Table 1). The residual bicarbonate concentration in the mutant medium was higher than that in the WT medium, especially under HC conditions, where the divergence was more evident. After 6 days of cultivation, there was approximately 3 mM HCO3- in the mutant medium, while it was approximately 1.4 mM HCO3- for the WT.
Biomass of Pyca1i mutants cultured under NC and HC conditions
Regarding biomass, the growth of ca1i-1 and ca1i-2 mutants was retarded relative to WT, resulting in a 31–38% lower biomass than WT under NC, while cali-6 lowered biomass by approximately 9%. Under HC conditions, ca1i-1 and ca1i-2 mutants showed 46–56% lower biomass than WT, while cali-6 lowered biomass by only 11% (Fig. 3f).
The total soluble protein, fatty acid (FA) and starch content in mutants and WT under different DIC
P. yezoensis is a macroalga widely known for its high nutritional value, particularly its rich protein, amino acid and unsaturated fatty acid (FA) contents. We compared the total soluble protein, FA and starch content of Pyca1i mutants and WT under different DIC conditions after 14 days of cultivation (Fig. 4).
Under NC conditions, the total soluble protein was similar for both mutants and WT at approximately 125 mg/g DW. However, under HC conditions, the total soluble protein content increased for mutants to 146 mg/g DW, while WT still had a higher content of 161.97 mg/g DW (Fig. 4a).
We also analyzed the FA composition by gas chromatography and found that mutants had lower levels of C16:0, the main saturated fatty acid in Pyropia, compared to WT, with levels of 21.8–22.3% and 24.1%, respectively. Additionally, mutants had lower levels of EPA, the main unsaturated fatty acid, than WT at 28.4–30% and 32.1%, respectively, under NC conditions (Fig. 4b).
The starch content for mutants and WT was 11.06–12.02 mg/g DW and 9.85 mg/g DW, respectively, under NC conditions. However, under HC conditions, the starch content increased to 12.98–13.73 mg/g DW for mutants and 10.73 mg/g DW for WT (Fig. 4c). This was further confirmed by transmission electron microscopy (TEM) images of starch accumulation in both Pyca1i mutants and WT under NC conditions (Fig. 4d).
Pyruvate content in mutants and WT under NC and HC conditions.
Under the two DIC conditions, pyruvate content was significantly lower in mutants compared to WT (P < 0.05) and did not increase much with higher [HCO3-] concentration in the medium (Fig. 4e).
Activity of enzymes involved in floridean starch synthesis, the PP pathway and photorespiration showed total divergence between ca1i mutants and WT
To account for changes in pH under two DIC conditions, with pH levels not exceeding 9.0 ± 0.1 on the 3rd day, all enzyme activities were detected during cultivation on the 3rd day.
The Rubisco carboxylation activity was similar for both mutants and WT under NC conditions but increased with higher [HCO3-] concentration in WT while showing a slight decrease in mutants (Fig. 5a). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a ubiquitous enzyme in plants, with two subtypes: one participates in glycolysis in the cytosol, and the other is involved in the Calvin cycle in plastids. In chloroplasts, GAPDH belongs to the NADPH-dependent subtype (Zaffagnini et al. 2013). NADPH-GAPDH catalyzes 1,3-bisphosphoglycerate into 3-phosphoglyceraldehyde in the Calvin cycle and plays a central role in CO2 utilization. The activity of NADPH-GAPDH in mutants was significantly higher than in WT under both DIC conditions, with activity ranging from 5.7–6.5 nmol/min/mg prot to 8.0-10.3 nmol/min/mg prot with HCO3- increase. In contrast, its activity in WT increased from 3.3 to 4.5 nmol/min/mg prot from NC to HC conditions. The activity of NADPH-GAPDH in mutants was approximately one-fold higher than that in WT (Fig. 5b).
The activities of glucose-6-phosphate dehydrogenase (G6PDH), which is involved in the pentose-phosphate (PP) pathway, were significantly higher in Pyca1i mutants than in WT under two DIC conditions, with activities of approximately 15 nmol/min/mg prot and 9.28 nmol/min/mg prot in mutants and WT, respectively, under NC conditions. With the increase in [HCO3-] in the medium, the enzyme activity increased as well (Fig. 5c).
Fructose-1,6-bisphosphatase (FBP) catalyzes fructose-1,6-bisphosphate conversion into fructose-6-P (Fru-6-P). The FBP activity in mutants and WT was approximately 1.5–1.6 and 1.0 nmol/min/mg prot, respectively, under NC conditions, and this enzyme activity in mutants and WT under HC conditions was similar to that under NC conditions (Fig. 5d).
UDPG pyrophosphorylase (UGP) and starch synthase (SSS) participate in the formation of carbonate storage, such as starch and floridoside, in Pyropia (Yu et al., 2021). The UGP enzyme activity in mutants and WT was approximately 6 and 3.55 nmol/min/mg prot under NC conditions, respectively, and increased to approximately 10 and 5 nmol/min/mg prot under HC conditions for mutants and WT, respectively (Fig. 5e). The change in SSS activity was similar to that of UGP activity between mutants and WT; furthermore, the SSS activity in Pyca1i-2 was approximately 1-fold higher than that of WT under these two DIC conditions (Fig. 5f).
2-Phosphoglycolate phosphatase (PGP) catalyzes the initial step of the photorespiratory pathway in various organisms, including yeast. The PGP content under NC conditions was measured to be 73.19 ng L-1 for WT and approximately 80 ng L-1 for the mutants. However, under HC conditions, the PGP content in the mutants increased to approximately 90 ng L-1 when the [HCO3-] in the medium was increased, while the PGP content in WT remained lower than that of the mutants, measuring 81.2 ng L-1 (Fig. 5g).
{kind=link}