The use of ASOs for the treatment of important genetic disorders has emerged as a promising approach (11,12,14,33,34). More recently, this approach has also shown promissory results in the control of C. albicans virulence determinants (35). However, ASO penetration into Candida cells can be limited by the cell wall envelope which is composed of several layers surrounding the cytosol (36). Therefore, delivery strategies are needed to ensure that ASO crosses Candida’s cell wall and that it also confers protection against degradation by serum proteases and nucleases in the host (37). At the present time, most delivery approaches for gene delivery employ either viral or non-viral vectors (17). Viral vectors show high gene silencing activity, however they present associated risks, including the activation of inflammatory and immune responses in the host (17,38). Some of these problems can be circumvented by employing non-viral vectors, such as cationic lipoplexes (16,18,39,40), which are assemblies that result from the electrostatic complexation between cationic liposomes and negatively charged ASO (31). Lipoplexes tend to have well-ordered hexagonal, cubic bicontinuous or multilamellar structures with the ASO sandwiched between the bilayers (18). Thus, the main goal of this work was to study the formation of cationic lipid- anti-EFG1 2’OMe oligomer complexes along with their capability to control C. albicans filamentation in vitro and in vivo way using the G. mellonella model. Three different neutral lipids (DOPC, DOPE and MO) combined with DOTAP (cationic lipid) were used, in a composition range where multilamellar structures are expected to be formed.
Lipoplex characterization
To note, cationic and neutral lipids are usually used to form liposomes for gene delivery due to their favourable interactions with negatively charged RNA or DNA and with cell membranes (31). Moreover, the neutral lipids are usually included as ‘helper’ lipids to enhance the transfection activity and nanoparticle stability (17,18,40). Lipoplexes can adopt different structures (e.g. lamellar) depending on the type of lipids used and number of cationic and neutral charges in its composition (Table 1). The association between n ASO and m cationic liposomes (CLs) to form a mCL.nASO complex can be considered according to the following equilibrium:
|
mCL + nASO ⇌ mCL.nASO
|
(3)
|
For simplicity we will refer to mCL.nASO complexes simply as CL-ASO complexes or lipoplexes. If ASO and CLs are labelled with two different fluorescent dyes (in this case, ASO with Tye-563 – red – and CLs with Atto-488 – green), this association can be followed with Dual-color Fluorescence Cross-Correlation Spectroscopy (FCCS) (41–43).
For such scenario, if one neglects cross-talk between the two fluorescent channels, the amplitudes of the autocorrelation curves of the individual green and red channels (GG and GR, respectively) and the amplitude of the cross-correlation curve between them (GX) are given by Eqs 4 (42,43):

Here qCL and qASO are factors accounting for quenching (qi <1) or FRET (qi >1) of the molecular brightness of complexed liposomes and ASO in the green and red detectors, respectively. VG and VR are the effective volumes in the green and red channels and VX is the volume overlap between them. CCL, CASO and CCL-ASO are the concentrations of CLs, ASO and CL-ASO in those effective volumes, respectively. Importantly, while GG and GR are concerned with the dynamics and relative concentrations of all the species that are labelled green or red, respectively, GX is concerned with the dynamics and relative concentrations of species that carry both dyes simultaneously (i.e. CL-ASO complexes). Hence, FCCS is an excellent probe to monitor the association between ASO and liposomes.
While, experimentally, we try to minimize cross-talk between the two channels, the equations are still very complicated to solve since there are many experimental unknowns difficult to determine. This is in contrast with more conventional lipid-DNA lipoplexes, where the large DNA length and charge ensure a practically complete DNA encapsulation for ρchg >1, and complete cationic liposome consumption for ρchg<1, allowing a simplification of some of the terms in eqs. 4 (43). For ASO these assumptions cannot be entirely met. However, even though Eqs. 4 cannot be used to determine m and n and overall encapsulation efficiency, FCCS can still provide important information, especially if the experimental results from the cationic liposome formulations are compared with a control system where complexation is not expected to occur. Figure 1 shows the auto- and cross-correlation curves for the different cationic liposome-ASO formulations at ρchg=3 and high cationic lipid content (80/20). In addition, pure DOPC liposomes incubated with ASO used as a control system is also shown (Figure 1 D). Since DOPC liposomes are net-neutral, no significant interactions with ASO leading to the formation of lipoplexes are expected. The concentration of DOPC used was equivalent to the concentration of DOTAP used for ρchg=3.
Figure 1 shows high GX amplitudes, comparable to both GG, and GR, for the cationic liposome formulations. In contrast, the DOPC sample shows a much reduced amplitude of GX, whose value is ca. 0.4 (this value is not zero most likely because of cross-talk of the green fluorescence of the liposomes to the red channel). Through eq. 4c it is possible to see that the larger GX amplitudes (beyond cross-talk) observed for the cationic formulations is indicative of association between ASO and cationic liposomes, and hence, the formation of lipoplexes. Another strong indication for the formation of lipoplexes can be extracted from the GR amplitudes, which are related with the ASO. In the DOPC formulations the amplitude of GR is practically zero, indicating a very large number of independent ASO molecules diffusing in solution. This is what would be expected if no lipoplex formation occurs. In contrast, in the cationic liposome formulations the amplitude of GR is finite, comparable to the amplitudes of GG and GX, thus indicating a much smaller number of independent ASO species in solution. This is what would be expected if the ASO are complexed with cationic liposomes. Details on the differences in amplitudes of GR, GG and GX for the different cationic liposome formulations are difficult to interpret due to the large number of unknowns in eqs 4, but overall, even without the possibility to determine m, n and encapsulation efficiency, the formation of lipoplexes is confirmed. These results are also in agreement with what is found for other systems of small oligonucleotides with cationic liposomes and cationic surfactants, in which lipoplex formation is observed for different ρchg (25,44,45).
The amplitudes of GX are very high in the case of cationic liposome formulations, but very low for DOPC, indicating the formation of CL-ASO complexes in the former but not on the latter. The very low amplitude of GR for DOPC also suggests that a very high amount of ASO is diffusing independently (thus not complexed), whereas the high amplitudes of GR for the cationic liposome formulations suggests the opposite.
In addition to the quantitative information regarding the association between ASO and cationic liposomes, fitting of the cross-correlation curves also provides the size of the lipoplexes. FCCS can be expected to provide more accurate size estimates for a polydisperse solution than, for example, Dynamic Light Scattering (DLS) (43). For DOTAP/DOPC formulations, one observed a radius ranging from 25-35 nm, with lipoplexes prepared at ρchg=3 slightly smaller to those prepared at ρchg=10 (Table S1). For DOTAP/DOPE formulations, a similar trend was observed with particles ranging from 25-40 nm, and with a slight smaller size observed for formulations prepared are ρchg=3 being slightly smaller (Table S1). Finally, DOTAP/MO formulations appeared to produce the smallest particles with sizes ranging from 18-25 nm (Table S1), were those containing more cationic lipid were shown to be slightly larger, independent to the ρchg.
In vitro effects of lipoplexes on C. albicans filamentation
The anti-EFG1 2’OMe oligomer cationic lipid-based formulations efficacy was evaluated in terms of its ability for controlling C. albicans filamentation (Figure 2). Noteworthy, all formulations were able to control C. albicans filamentation, although to different extents. It is also important to highlight that in general the reduction on C. albicans filamentation increases over time for all lipoplexes, with a similar trend observed for free ASO.
It should be noted that the encapsulation of anti-EFG1 2’OMe oligomer with DOTAP/DOPC liposomes (Figure 2 A) shows a slight improvement in the C. albicans filamentation reduction after 48 h and 72 h when compared to free-ASO (P-value>0.05). Figure 2 B presents the data related to treatment of C. albicans cells with DOTAP/DOPE lipoplexes. All the DOTAP/DOPE formulations feature a similar or superior efficacy to that of free-ASO. All DOTAP/MO lipoplexes tested improved the efficacy in comparison to free-ASO, especially at 48 h (P-value<0.05). The DOTAP/MO 80/20 ρchg=10 lipoplexes, typically with a lamellar structure, showed the highest ability to control C. albicans filamentation. This could be related with the fusogenicity of MO which may facilitate the fusion of the lipoplex membranes with endosomal membranes and consequently facilitate endosomal release (46,47). Both effects are thought to facilitate the fusion of the lipoplex membranes with endosomal membranes and consequently facilitate endosomal release (47,48). Overall, these results are in agreement with the findings for an analogous system composed by DOTAP/MO but encapsulating siRNA (46).
Figure 3 (A and B) shows that all lipoplexes tested were not only able to decrease the number of C. albicans cells as filaments but also to reduce their filament’s length up to 40-70 % (P-value>0.05).
In vivo effects of lipoplexes on G. mellonella survival
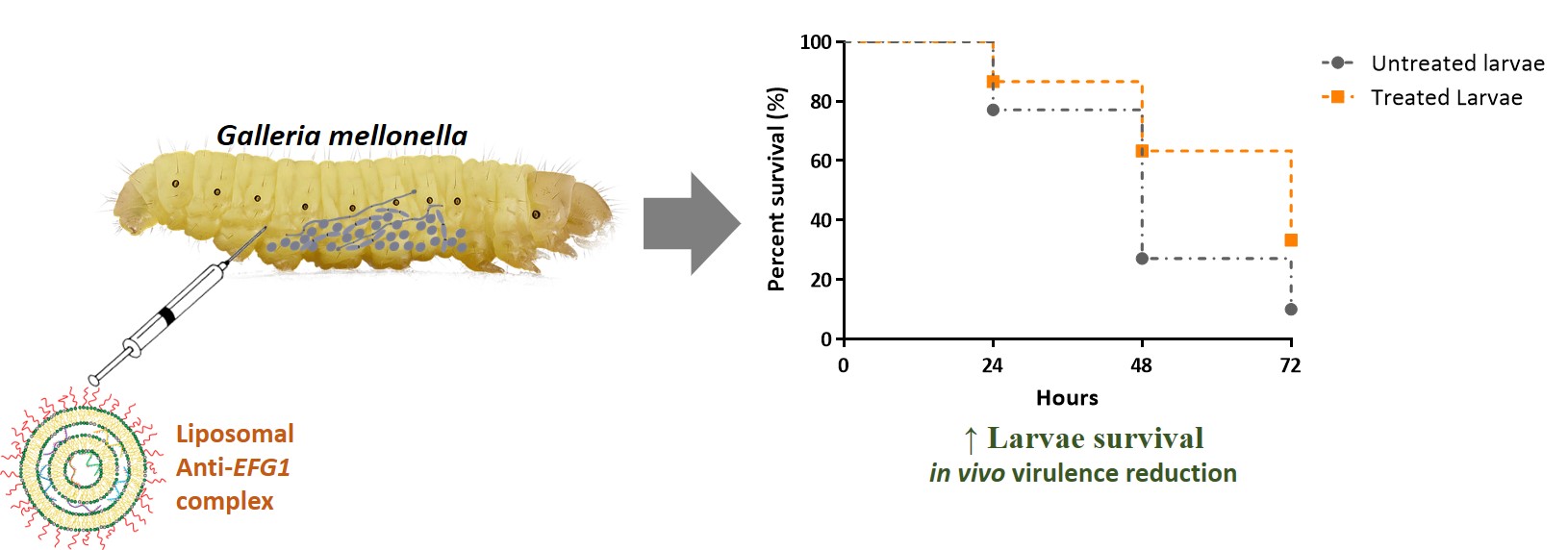
The invertebrate model of G. mellonella was used to test in vivo the efficacy of lipoplexes carrying anti-EFG1 2’OMe ASO. The lipoplexes tested in vivo were DOTAP/DOPC 80/20 ρchg=3, DOTAP/DOPC 80/20 ρchg=10, DOTAP/DOPE 80/20 ρchg=10 and DOTAP/MO 80/20 ρchg=10 formulations. Two of which were selected based on the in vitro data: DOTAP/MO 80/20 ρchg=10 with the best and the DOTAP/DOPC 80/20 ρchg=3 due to its lower efficacy in vitro. Additionally, were also chose the formulaltions DOTAP/DOPC 80/20 ρchg=10 and DOTAP/DOPE 80/20 ρchg=10.
The toxicity effects of the anti-EFG1 2’OMe ASO cationic lipid-based formulations was evaluated. Notably, the results in Figure 4 A show the absence of toxicity for all lipoplexes tested.
To study the effect of anti-EFG1 2’OMe ASO lipid-based formulations in vivo, G. mellonella larvae infected with a lethal dose of C. albicans were treated with the anti-EFG1 2’OMe oligomer cationic lipid-based formulations. The treatment with a single-dose of each lipoplex enhanced the survival of larvae over 72 h (Figure 5). To note, that DOTAP/DOPC 80/20 ρchg=3 presented the best performance, enhancing the G. mellonella survival by 19 % (at 24 h and 48 h) and by 14 % (at 72 h) (P-value<0.05). In contrast, DOTAP/DOPC 80/20 ρchg=10 revealed to be the less efficient. Therefore, in contrast to the in vitro findings, in vivo the ρchg seems to influence the DOTAP/DOPC 80/20 lipoplexes efficacy, with the ρchg=3 formulation presenting a higher increase on larvae survival than ρchg=10.
It was also observed that DOTAP/DOPE 80/20 ρchg=10 (Figure 5 B) increased the G. mellonella survival over 72 h (P-value<0.05), however to a slight extension when compared to the performance of DOTAP/DOPC 80/20 ρchg=3. Although the G. mellonella survival increased after 24 h and 48 h of treatment with the DOTAP/MO 80/20 ρchg=10 lipoplex in comparison to the untreated larvae, no statistic differences were observed after 72 h (Figure 5 C) (P-value>0.05).
The invasiveness progression of C. albicans into the fatty body of larvae is represented in Figure 5D. Figure 5A confirms that C. albicans cells tend to locate mainly in the G. mellonella digestive system (49). Moreover, an extensive progression is evident over the 48 h of infection and is predominantly composed of the filamentous form, which are more invasive to tissues than yeast cells and provides a mechanism for evasion of the host’s defence mechanism (50). The treatment of G. mellonella with all the lipoplex formulations resulted in a lower quantity of C. albicans cells on the larvae digestive system with a significant reduction on the number of filaments (Figure 5 B-D).
To mimic a more real clinical application (51,52), the effect of double-dose administration of the DOTAP/DOPC 80/20 ρchg=3 formulation (the lipoplex with the best efficacy in the single-dose studies) was analysed. To note, the double-dose administration of DOTAP/DOPC 80/20 ρchg=3 enhances the G. mellonella survival by a factor of ca. 1.5 when comparing to the single-dose administration, increasing the larvae survival by 40 % after 48 h and by 25 % after 72 h of treatment (P-value<0.005) (Figure 6 A). It is important to point out that the double-dose administration also potentiates the effect of free-ASO by a factor of ca. 1 (a survival increase by 18 % after 24 h and 13 % after 72 h of infection (P-value<0.05)). Moreover, the fatty bodies histological analysis confirms the decrease on C. albicans cells filamentation when treated with the DOTAP/DOPC 80/20 ρchg=3 lipoplex (Figure 6 B).
Effectively, the survival rate of G. mellonella when treated with the anti-EFG1 2’OMe ASO cationic lipid-based formulations was significantly higher than when treated with free-ASO. This evidence confirms the importance of the encapsulation for protecting ASO against the degradation by serum proteases and nucleases.
Lipoplexes have been studied for the delivery of nucleic acids in vitro and in clinical trials (53) often with promising results (21). Here we show that lipoplexes also successfully encapsulate and deliver anti-EFG1 2’OMe oligomers into C. albicans cells, helping to control filamentation and improving the survival of infected larvae model.
This study demonstrated the successful delivery of anti-EFG1 2’OMe ASO encapsulated in cationic lipid-based formulations, providing valuable information for further assays. Although, DOTAP/DOPC 80/20 ρchg=3 had a lower efficacy to inhibit filamentation in vitro, it was revealed to be the best formulation in vivo to control C. albicans virulence. Therefore, the DOTAP/DOPC 80/20 ρchg=3 formulation has the potential to improve nanodrug administration for C. albicans species.


{kind=link}