
Abstract
Analysis of the structure of the morphological diversity of the Altai osmans of the genus Oreoleuciscus in the Urt River (water system of the Tes River) indicates that this is a polymorphic population, consisting of three forms, clearly differentiated by a set of characters. The ratio of differences between these forms and forms previously identified in the Zavkhan (water system of the Great Lakes Hollow) and Tuin (water system of the Lake Valley) rivers suggests that the characters considered in the study are adaptively plastic, and the revealed structure of the morphological diversity of the genus is mostly determined by local patterns of environmental conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
Investigations of populations and intraspecific forms of different groups of organisms are important not only in respect to the microevolution processes, but also towards an adaptation to local environmental conditions. Altai osmans (genus Oreoleuciscus) is a peculiar group of cyprinids (Cyprinidae) that inhabits drainless lakes and lake-river systems of Altai and northwestern Mongolia. This group has been relatively little studied. The native ichthyofauna of such water bodies, located at altitudes from 700 to 2200 m above sea level, is characterized by a relatively low species diversity, thus in some reservoirs osmans are the only representatives of fish (Dgebuadze, 1982; Baasanzhav et al., 1983; Dgebuadze et al., 2012).
The genus Oreoleuciscus was described more than a hundred years ago (Warpachowski, 1889); however, the Altai osmans are so much phenotypically diverse that researchers still do not have a common view on the number of species, subspecies, and forms with an uncertain taxonomic status that make up this genus. This is reflected in a number of review publications (Vasil’eva, 1985; Golubtsov et al., 1999; Bogutskaya, 2001; Kottelat, 2006; Dgebuadze et al., 2012; Kartavtsev et al., 2016). The authors of this study adhere to the classification of the genus Oreoleuciscus presented by Dgebuadze et al. (2012).
According to genetic studies, Oreoleuciscus populations are clearly divided into groups corresponding to three geographic regions (Slynko and Dgebuadze, 2009; Slynko and Borovikova, 2012). One group of populations, corresponding to the previously described species O. potanini (Kessler, 1879), inhabits the water systems of the Great Lakes Hollow, which belongs to the Central Asian Closed Basin. The second group corresponds to another previously described species, O. humilis Warpachowski, 1889, and inhabits the water bodies of the Lake Valley of the same basin. The level of genetic differences between these two groups of populations predictably corresponds to the species level. Populations of the third group identified by the above authors inhabit the Arctic Ocean Basin, in particular, the water systems of the Selenga, Orkhon, and Tes rivers and Lake Uvs. The level of genetic differences of the Arctic Basin populations from the Central Asian ones also corresponds to the interspecies, and in the article by Slynko and Borovikova (2012), the Arctic Basin osmans are considered as an independent species O. manchini. Unfortunately, due to the lack of the species description in accordance with the International Code of Zoological Nomenclature (2004), this name is nomen nudum, therefore, in this work the population of the Urt River (one of the objects of this study, belonging to the Arctic Ocean Basin) is designated as Oreoleuciscus sp.
In lakes, Altai osmans often form ecological forms that differ in their feeding and morphology (Dgebuadze, 1982; Baasanzhav et al., 1983; Borisovets et al., 1984, 1985; Mironovsky et al., 2018, 2019). Riverine populations of Altai osmans were considered monomorphic. Only recently, morphologically differentiable forms have been identified in O. potanini from the Zavkhan River (Great Lakes Hollow) (Dgebuadze et al., 2017) and in O. humilis from the Tuin River (Lake Valley) (Mironovsky et al., 2019). A similar situation should be noted in African barbs of the genus Labeobarbus. For a long time, it was believed that sympatric speciation in this fish group occurs only under lacustrine conditions, however recently, sympatric forms of the African barbs have been found in faunistically depauperated (similar to the Mongolian) fish communities in some rivers of the Ethiopian Highlands (Levin et al., 2020; Golubtsov et al., 2021). In the same region in the Blue Nile water system, sympatric forms of the representatives of the genus Garra have been found (Levin et al., 2021).
The present work, which continues the series devoted to the study of osman variability in different localities, represents a comparative analysis of the structures of phenetic diversity of the above-mentioned population of Oreoleuciscus sp. in the Urt (= Urtyn) River, which flows into the lake Sangiyn-Dalai of the Tes River water system, and populations of O. potanini in the Zavkhan River and O. humilis in the Tuin River.
MATERIALS AND METHODS
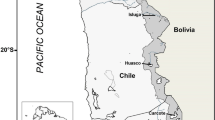
The study was carried out on the material collected on June 2, 2006 in the lower reaches of the Urt River, 49°17′ N, 99°00′ E (Fig. 1). Fish were caught with gillnets, mesh size of 12–60 mm; 30 specimens standard length (SL) of 95–144 mm were caught; most of the individuals were caught in nets with a mesh size of 12–18 mm. As in the samples from the Zavkhan and Tuin rivers (Dgebuadze et al., 2017; Mironovsky et al., 2019), we found no morphologically distinct groups of individuals during a visual assessment of the habitus of the caught fish, i.e. the sample seemed to be homogeneous.

Schematic map of the study area. 1, Zavkhan River; 2, Urt River; 3, Tuin River.
For the laboratory processing, fish heads along with the cleithrum (a pectoral girdle bone) were fixed with sodium chloride. After preparation of dry osteological preparations, 13 characters that were previously used in the study of the variability of Altai osmans and other groups of cyprinids (Vasil’eva and Ustarbekov, 1991; Mina et al., 1996; Dgebuadze et al., 2008; Mironovsky et al., 2018) have been measured (Fig. 2).

Scheme of osteological measurements: BL, skull basal length; B1, B2, and B3, distances between the outer edges of frontalia, pterotica, and sphenotica, respectively; B4, neurocranium width at the junction of frontale and pteroticum; HS1 and HS2, neurocranium height at the level of the parasphenoideum curve and the posterior edge of the parasphenoideum, respectively, Hm, hyomandibulare height; dff, distance between the external points of the branches of the pharyngeal tooth; Pop, diagonal length of praeoperculum; Op, operculum height; De, dentale length; Cltr, diagonal length of the pectoral girdle bone, cleithrum.
The measurement data were processed using the algorithms of the applied biostatistics package NTSYS 2.02k (cluster analysis and principal component analysis (PCA)) and the StatSoft Statistica 6 package (discriminant analysis). In the calculations, we used indices, i.e., the ratio of the absolute values of the measurement to the basal length of the skull (BL). Thus, all the characters mentioned in this work, are the indices but not the measurements itself. The degree of separation of objects in the multidimensional character space was assessed by calculating the taxonomic distance (Rohlf, 1998). Cluster analysis of similarity matrices was carried out by unweighted pair-group method; its results are presented by a dendrogram. In PCA, eigenvectors were calculated from the correlation matrix. The length of the vector was set to one. The minimum spanning tree (MST) was superimposed on the distribution of the objects in the space of the three first principal components (PC).
In addition to the traditional PCA algorithm, a method based on the construction of multidimensional ontogenetic channels was applied. This approach, which has proven itself well in the study of the phenetic diversity of a number of cyprinids, enables to assess the ratio of intergroup differences with differences in the size of individuals that make up the compared groups (Mina et al., 1996; Mina, 2001; Dgebuadze et al., 2017; Mironovsky et al., 2019).
In addition to Oreoleuciscus sp. from the Urt River, we used samples that served as the basis for our previous studies of the phenetic diversity of the river populations of the Altai osmans: 37 specimens of O. potanini (SL 133–188 mm) from the Zavkhan River (Dgebuadze et al., 2017) and 38 specimens of O. humilis (SL 78–136) from the Tuin River (Mironovsky et al., 2019).
RESULTS AND DISCUSSION
Figure 3 presents the results of a discriminant analysis of the set of considered osteological parameters for the osman samples from the three studied rivers. In the space of the first and second discriminant functions, the three distributions are clearly separated. This being said, the separation level of representatives of Oreoleuciscus sp. from O. potanini and O. humilis is similar to that between the latter two. It is important to note that in this scatterplot, the samples of different species differ with a pronounced hiatus, while the size range (SL) of the individuals of the analyzed samples overlap in any combination. So, the differences in the analyzed parameters cannot result only from changes in the skull proportions during the fish grow. Thus, according to the set of characters considered, the individuals of the three analyzed populations differ at the “each from each” level, which testify in favor of the based on genetic data idea of the Urt River population as of a previously undescribed species of the genus Oreoleuciscus (Slynko and Borovikova, 2012).

Distribution of the osman Oreoleuciscus potanini individuals from the Zavkhan River (△), O. humilis from the Tuin River (◻), and Oreoleuciscus sp. from the Urt River (◇) in the coordinates of the first (df 1) and second (df 2) discriminant functions.
The discriminant analysis algorithm used above is designed to test the hypothesis about the differences between obviously different groups of individuals, in our case, allopatric species. However, the presence of different groups of individuals in the Urt River was not previously shown, although the grounds for assuming their existence seem obvious by analogy with the morphological forms of the osmans of the Zavkhan and Tuin rivers. This assumption can be verified using cluster analysis algorithms and PCA (Aivazyan et al., 1989).
It is well seen on the Figure 4, that in according to considered characters, the dots corresponding to the individuals of the sample from the Urt River assemble distinctly separated groups (indicated by the numbers 1, 2, and 3). The dendrogram clusters (Fig. 4a) completely coincides with the composition of the three polygons on the plane of the first two PCs in Fig. 4b. The corresponding ontogenetic channels in the Fig. 4c are separated, and the ratio of differences in the skull length (abscissa axis) and in the PC1 of the set of analyzed characters (ordinate axis) does not provide opportunities to consider the differences in the groups nos. 1, 2, and 3 as differences in size groups of a monomorphic population, reflecting changes in proportions as individuals grow. It is more likely that, like in the cases of the populations of the Zavkhan and Tuin rivers (Dgebuadze et al., 2017; Mironovsky et al., 2019), the population of the Urt River is polymorphic and consists of three sympatric forms.

Results of multivariate analysis of morphological variability of the Altai osman Oreoleuciscus sp. populations in the Urt River: (a) dendrogram of the similarity of individuals: D, multidimensional taxonomic distance (according to: Rohlf, 1998); (b) distribution of individuals on the plane of the first two principal components (PC1 and PC2) of the variability of the studied characters; (c) distribution of individuals on the plane of ontogenetic channels: BL, skull base length; 1–3, intrapopulation morphological forms.
It follows from the Table 1 data that in the PCA of the variability of individuals in the Urt River, the eigenvalues (λ) of just PC1 and PC2 are >1, that is, according to the Kaiser’s rule (Kaiser, 1960), they are the only ones that comprise the constructive variability, the “signal”. The PC3, PC4, PC5, PC6, etc., involve mainly the “noise”, that is, stochastic, random variability. So, this suggests that the distribution of individuals in the PC1 and PC2 coordinates in the Figure 4b reflects the phenetic relationships of the individuals of the analyzed array with sufficient completeness and accuracy. The variance explained in PC1 is nearly four times greater than the variance in PC2, and only PC1 reflects the differences between the forms, while in PC2 the distributions of any pair of forms overlap almost completely. Thus, the differences between individuals of different forms determine the first (main) vector of dispersion (variability) in the given locality.
Figure 5 illustrates the relations of the three forms of the Urt River population, two forms of the Zavkhan River population, and two forms of the Tuin River population according to the average statistical evaluations of the characters of each form. The sum of the variance explained by PC1, PC2, and PC3 is more than 98% (Table 1). The eigenvalues of only PC1, PC2, and PC3 are >1, which suggests that the three-dimensional ordination in Fig. 5 quite fully characterizes inter- and intra-group variability. As in the Figure 4b, the forms within each population differ from each other mainly in PC1 (more than 60% of the total variance; Table). The forms of the Zavkhan river differ from the forms of the Tuin river primarily in PC2, while the contributions of PC2 and PC3 in the separation of the forms of the Urt River from the forms of the Zavkhan and Tuin rivers are approximately equal. More than that, the values of the explained variance in PC2 and PC3 (19.9 and 18.2%, respectively) are almost equal to each other and noticeably lower than the variance in PC1 (approximately three times each). Thus, in the diversification of morphological forms of three different species in three different water basins, the main vector of variability (PC1) determines the differences between the forms within each species, and not the differences between the species. At the same time, the morphological distances between the most distant (in the space of characters) forms of the same water basin are in some cases greater than the morphological distances between the closest forms of different water basins.

Distribution of forms of the osman Oreoleuciscus sp. of the Urt River (⚪, ⚫,  ), O. potanini of the Zavkhan River (
), O. potanini of the Zavkhan River ( ), and O. humilis of the Tuin River (
), and O. humilis of the Tuin River ( ) in the space of the first three principal components (PC) with the imposition of the minimum spanning tree. The numbers indicate the values of the multidimensional taxonomic distance (according to: Rohlf, 1998).
) in the space of the first three principal components (PC) with the imposition of the minimum spanning tree. The numbers indicate the values of the multidimensional taxonomic distance (according to: Rohlf, 1998).
The situation may seem unexpected; however, to explain it we can suggest quite a probable working hypothesis. If we refer the works on the assessment of the genetic parameters of the three considered species of osmans, we’ll see that the ratio of differences between them in the sequence of the Cyt b gene of mitochondrial (mt) DNA and in the analysis of isoenzymes do not coincide (Slynko and Dgebuadze, 2009; Slynko and Borovikova, 2012). According to the mtDNA Cyt b gene, O. potanini and the putative Oreoleuciscus sp. are closer to each other, while according to the isoenzymes analysis, O. potanini is closer to O. humilis. The authors believe that such a difference in the results of isoenzyme analysis and analysis of polymorphism of the nucleotide sequences of the Cyt b gene results from the “selective significance” of isoenzymes, the intersample differences of which are largely determined by the patterns of the living (environmental) conditions of the regions under consideration (Slynko and Borovikova, 2012, p. 732). This assumption is not unfounded, although in this context, in our opinion, it is more appropriate to point adaptive plasticity rather than selective significance. The minimum spanning tree in Figure 5 shows that O. humilis and O. potanini are nearest neighbors in the space of morphological parameters, similar to their proximity in isoenzymes in the cited work.
All the foregoing suggests that the variability of the considered morphological characters, like the variability of isoenzymes, largely results from the local adaptations. Based on this assumption, a high level of differences between some forms of the same species compared with not so big differences between some forms belonging to different species (Fig. 5), can be explained by the more pronounced differences in the habitats in the same river (the presence of high contrasting habitats) than the differences in the most similar biotopes of different rivers. This determines the need for further research, which should be aimed at studying the ecological living conditions of the identified forms in the rivers under consideration, as well as increasing the number of considered characters with the shown high adaptive plasticity in changing habitat conditions (Baranov and Vasil’ev, 2022).
CONCLUSIONS
1. Analysis of the variability of the complex of morphological characters shows that the population of the Altai osmans in the Urt River (the Tes River system) is not monomorphic and, similar to the osman populations of the Zavkhan (Great Lakes Hollow) and Tuin (Lake Valley) rivers, it is subdivided into morphologically differentiated forms.
2. The ratio of differences between the morphological forms of the osmans from the Zavkhan, Tuin and Urt rivers suggests that the considered characters are adaptively plastic, and the revealed structure of the diversity of the studied populations of O. potanini, O. humilis, and putative Oreoleuciscus sp. is largely determined by the local characteristics of habitat conditions.
REFERENCES
Aivazyan, S.A., Bukhshtaber, V.M., Enyukov, I.S., and Meshalkin, L.D., Prikladnaya statistika. Klassifikatsiya i snizhenie razmernosti (Applied Statistics. Classification and Dimensionality Reduction), Moscow: Finansy i Statistika, 1989.
Baasanzhav, G., Dgebuadze, Yu.Yu., Demin, A.N., et al., Review of species of the ichthyofauna of the Mongolian People’s Republic, in Ryby Mongol’skoi Narodnoi Respubliki (Fishes of the Mongolian People’s Republic), Moscow: Nauka, 1983, pp. 102–224.
Baranov, V.Yu. and Vasil’ev, A.G., Phenotypic plasticity of the Amur sleeper (Perccottus glenii) invasive populations during the colonization of water bodies, Russ. J. Biol. Invasions, 2022, vol. 13, no. 4, pp. 412–427. https://doi.org/10.1134/S2075111722040038
Bogutskaya, N.G., A revision of Altai osmans of the genus Oreoleuciscus (Leuciscinae, Cyprinidae) with a description of a new species, O. angusticephalus, from river Kobdo (Hovd) system, West Mongolia, Proc. Zool. Inst., 2001, vol. 287, pp. 5–43.
Borisovets, E.E., Dgebuadze, Yu.Yu., and Ermokhin, V.Ya., Experience in studying the morphological variability of fish of the genus Oreoleuciscus (Pisces, Cyprinidae) using multivariate statistics methods, Zool. Zh., 1984, vol. 63, no. 4, pp. 563–572.
Borisovets, E.E., Dgebuadze, Yu.Yu., and Ermokhin, V.Ya., Morphometric analysis of Altai osmans (Oreoleuciscus; Pisces, Cyprinidae) in the water bodies of the Mongolian People’s Republic: A multivariate approach, Ibid., 1985, vol. 64, no. 8, pp. 1199–1212.
Dgebuadze, Yu.Yu., Mechanisms of morphogenesis and taxonomy of fish of the genus Oreoleuciscus (Cyprinidae, Pisces), in Zoologicheskie issledovaniya v MNR (Zoological research in the Mongolian People’s Republic), Moscow: Nauka, 1982, pp. 81–92.
Dgebuadze, Yu.Yu., Mina, M.V., and Mironovsky, A.N., On estimation of phenetic relationships of Altai Osmans (Oreoleuciscus, Cyprinidae) from three lakes in Mongolia by skull characters, J. Ichthyol., 2008, vol. 48, no. 4, pp. 292–300. https://doi.org/10.1134/S0032945208040024
Dgebuadze, Yu.Yu., Mendsaikhan, B., and Dulmaa, A., Diversity and distribution of Mongolian fish: Recent state, trends and studies, Erforsh. Biol. Ress. Mongolei (Halle/Saale), 2012, vol. 12, pp. 219–230.
Dgebuadze, Yu.Yu., Mironovskii, A.N., Mendsaikhan, B., and Slyn’ko, Yu.V., The first case of morphological differentiation of Altai osman Oreoleuciscus potanini (Cyprinidae, Actinopterigii) in a river, Dokl. Biol. Sci., 2017, vol. 473, no. 1, pp. 57–60. https://doi.org/10.1134/S0012496617020053
Golubtsov, A.S., Berendzen, P.B., and Cynthya, A., Morphological variation and taxonomic status of Altai osmans, Oreoleuciscus, from the upper reaches of the Ob River system, J. Fish. Biol., 1999, vol. 54, no. 4, pp. 878–899. https://doi.org/10.1111/j.1095-8649.1999.tb02039.x
Golubtsov, A.S., Korostelev, N.B., and Levin, B.A., Monsters with a shortened vertebral column: A population phenomenon in radiating fish Labeobarbus (Cyprinidae), PLoS ONE, 2021, vol. 16, no. 1, Article e0239639. https://doi.org/10.1371/journal.pone.0239639
Kaiser, H.F., The application of electronic computers to factor analysis, Educ. Psychol. Meas., 1960, vol. 20, no. 1, pp. 141–151. https://doi.org/10.1177/001316446002000116
Kartavtsev, Yu.Ph., Batischeva, N.M., Bogutskaya, N.G., et al., Molecular systematics and DNA barcoding of Altai osmans, Oreoleuciscus (Pisces, Cyprinidae, and Leuciscinae), and their nearest relatives, inferred from sequences of cytochrome b (cyt-b), cytochrome oxidase c (co-1), and complete mitochondrial genome, Mitochondrial DNA pt. A, 2016, vol. 28, no. 4, pp. 502–517. https://doi.org/10.3109/24701394.2016.1149822
Kottelat, M., Fishes of Mongolia. A Check-List of the Fishes Known to Occur in Mongolia with Comments on Systematics and Nomenclature, Washington: The World Bank, 2006.
Levin, B.A., Simonov, E., Dgebuadze, Yu.Yu., et al., In the rivers: Multiple adaptive radiations of cyprinid fishes (Labeobarbus) in Ethiopian Highlands, Sci. Rep., 2020, vol. 10, no. 1, Article 7192. https://doi.org/10.1038/s41598-020-64350-4
Levin, B., Simonov, E., Franchini, P., et al., Rapid adaptive radiation in a hillstream cyprinid fish in the East African White Nile River basin, Mol. Ecol., 2021, vol. 30, no. 21, pp. 5530–5550. https://doi.org/10.1111/mec.16130
Mezhdunarodnyi kodeks zoologicheskoi nomenklatury (International Code of Zoological Nomenclature), Moscow: KMK, 2004.
Mina, M.V., Morphological diversification of fish as a consequence of the divergence of ontogenetic trajectories, Russ. J. Dev. Biol., 2001, vol. 32, no. 6, pp. 397–402.
Mina, M.V., Mironovsky, A.N., and Dgebuadze, Yu.Yu., Lake Tana large barbs: Phenetics, growth and diversification, J. Fish. Biol., 1996, vol. 48, no. 3, pp. 383–404. https://doi.org/10.1111/j.1095-8649.1996.tb01435.x
Mironovsky, A.N., Dgebuadze, Yu.Yu., Mendsaikhan, B., and Slyn’ko, Yu.V., Morphological Differentiation of Altai Osman Oreoleuciscus humilis (Cyprinidae) in the Tuin-Gol River, Lake Valley, Mongolia, J. Ichthyol., 2019, vol. 59, no. 1, pp. 115–119. https://doi.org/10.1134/S0032945219010077
Mironovsky, A.N., Slyn’ko, Yu.V., Kasyanov, A.N., and Goroshkova, T.V., Comparative analysis of variation in ecological forms of Altai osman, Oreoleuciscus potanini (Cyprinidae), in lakes Ayrag, Khyargas, and Nogon (Western Mongolia), Russ. J. Ecol., 2018, vol. 49, pp. 160–165. https://doi.org/10.1134/S1067413618020091
Rohlf, F.J., NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System (Version 2.02k), New York: Exeter softw. Publ., 1998.
Slynko, Yu.V. and Borovikova, E.A., Phylogeography of Altai osmans fishes (Oreoleuciscus sp., Cyprinidae, Pisces) inferred from nucleotide variation of the mitochondrial DNA cytochrome b gene, Russ. J. Genet., 2012, vol. 48, no. 6, pp. 618–627. https://doi.org/10.1134/S1022795412060154
Slyn’ko, Yu.V. and Dgebuadze, Yu.Yu., Population-genetical analysis of Altai osmans (Oreoleuciscus, Cyprinidae) from water bodies of Mongolia, J. Ichthyol., 2009, vol. 49, no. 8, pp. 614–626. https://doi.org/10.1134/S0032945209080050
Varpakhovskii, N.A., Monografiya novogo roda karpovykh ryb (Oreoleuciscus) (Monograph of a New Genus of Cyprinids (Oreoleuciscus)), St. Petersburg: Tip. Imperat. Akad. Nauk, 1889.
Vasil’eva, E.D., On the taxonomic status of some forms of the genus Oreoleuciscus (Cyprinidae), Vopr. Ikhtiol., 1985, vol. 25, no. 2, pp. 196–211.
Vasil’eva, E.D. and Ustarbekov, A.K., Skull morphology of the shemaya Chalcalburnus chalcoides (Cyprinidae), Ibid., 1991, vol. 31, no. 4, pp. 556–564.
ACKNOWLEDGMENTS
The authors are sincerely grateful to the leadership of the Joint Russian-Mongolian Complex Biological Expedition of the Russian Academy of Sciences and the Academy of Sciences of Mongolia for assistance in organizing work in Mongolia, as well as to Yu.Yu. Dgebuadze (Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences) and B.A. Levin (Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences), for their valuable critical comments for the first versions of the manuscript.
Funding
The work was carried out within the framework of the State Orders of the Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences (nos. 0109-2018-0076, AAAA-A18-118042490059-5 and FFER-2021-0006) and Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences (nos. 121051100109-1 and 121051100104-6).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflicts of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by T. Kuznetsova
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mironovsky, A.N., Slynko, E.E. Comparative Analysis of the Morphological Diversity Structure of Altai Osmans of the Genus Oreoleuciscus (Cyprinidae) in the Riverine Populations of Three Mongolian Water Systems. J. Ichthyol. 63, 417–424 (2023). https://doi.org/10.1134/S0032945223030098
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0032945223030098