

Abstract
Linking annual tree growth with remotely-sensed terrestrial vegetation indices provides a basis for using tree rings as proxies for ecosystem primary productivity over large spatial and long temporal scales. In contrast with most previous tree ring/remote sensing studies that have focused on temperature-limited boreal and taiga environments, here we compare the normalized difference vegetation index (NDVI) with a network of Pinus brutia tree ring width chronologies collected along ecological gradients in semiarid Cyprus, where both radial tree growth and broader vegetation activity are controlled by drought. We find that the interaction between precipitation, elevation, and land-cover type generate a relationship between radial tree growth and NDVI. While tree ring chronologies at higher-elevation forested sites do not exhibit climate-driven linkages with NDVI, chronologies at lower-elevation dry sites are strongly correlated with NDVI during the winter precipitation season. At lower-elevation sites, land cover is dominated by grasslands and shrublands and tree ring widths operate as a proxy for ecosystem-scale vegetation activity. Tree rings can therefore be used to reconstruct productivity in water-limited grasslands and shrublands, where future drought stress is expected to alter the global carbon cycle, biodiversity, and ecosystem functioning in the 21st century.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence.
Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
There is extensive scientific debate regarding the direction and magnitude of recent changes in ecosystem primary productivity across global biomes (Guay et al 2014, Poulter et al 2014), but knowledge of long-term, multidecadal to centennial scale vegetation activity fluctuations and their causes is limited (Tielbörger et al 2014). Global semiarid shrublands and grasslands are major drivers of interannual variability in the global carbon cycle, and experience critical loss of biodiversity and ecosystem functioning, plant community restructuring, and water cycle changes following drought-induced vegetation die-off events (Ahlström et al 2015, Breshears et al 2005, Kunstler et al 2006, Lloret et al 2016, Poulter et al 2014, Royer et al 2011). Understanding the long-term response and resilience of these ecosystems to drought and aridity is particularly important in the Mediterranean region, a transitional climate zone that is especially vulnerable to precipitation declines, increasing temperatures, and a greater frequency of droughts and heat waves, as a result of anthropogenic climate change (Hoerling et al 2012, IPCC 2013).
An emerging body of research attempts to relate remotely-sensed vegetation activity estimates to tree ring variables (including ring width, density, and stable isotopes), for the purpose of using tree rings as both in-situ indicators and long-term paleoenvironmental proxies of primary productivity (Vicente-Serrano et al 2016). The latter application would allow for examination of long-term vegetation activity and climate interactions and provide a fuller context for evaluating modern ecosystem, productivity, and carbon cycle shifts. In northern high latitude forests, radial tree growth and forest canopy reflectance have been linked based on their common sensitivity to seasonal temperature, yet it remains unclear if and how canopy status and xylogenesis are physiologically connected (Beck et al 2011, 2013, Berner et al 2011, Bunn et al 2013, D'Arrigo et al 2000). In semiarid systems, both annual radial tree growth and broader vegetation activity are mainly controlled by moisture availability (Vicente-Serrano 2007, Vicente-Serrano et al 2013) and there is evidence of linkages between tree rings and vegetation indices in these settings (Leavitt et al 2008, Pasho and Alla, 2015, Poulter et al 2013, Vicente-Serrano et al 2013). A longer-term perspective on the response of productivity to drought could allow for a better understanding of ecosystem resistance, resilience, and vulnerability to mean state changes and extreme drought events (Vicente-Serrano et al 2013). The goal of this study was to identify topoclimatic, land cover, and phenological characteristics that promote a relationship between radial tree growth and ecosystem productivity, as measured by the normalized difference vegetation index (NDVI), in a semiarid environment. We analyze a network of Pinus brutia (Ten.) trees growing across an elevational gradient in Cyprus, where the species is distributed across a large range of environments and elevations and where its growth is often moisture-limited (Griggs et al 2014, Touchan et al 2014a, 2014b, 2016).
2. Study area
Cyprus experiences a typical Mediterranean climate characterized by hot dry summers and mild wet winters. During summer, subsidence corresponds with a northward shift of the subtropical high and Persian trough, and thunderstorms deliver less than 5% of the total annual rainfall (Price et al 1999). Small, unstable low pressure systems resulting from the continental anticyclone of Eurasia and persistent low-pressure belt over north Africa dominate during winter, delivering about 60% of the total annual precipitation between December and February. Spring and autumn are short with highly variable temperature and precipitation (Price et al 1999). The island's topography is dominated in the west by the Troodos massif (maximum elevation 1951 m asl) and in the northeast by the Kyrenia range (maximum elevation 1000 m asl) (Price et al 1999). The higher western ranges receive greater precipitation, and may experience up to 1.5 m of snow persisting for several weeks during winter (Pashiardis and Michaelides 2008). The tree ring sample sites are located at varying elevations within the two mountainous regions, from 181 to 1489 m asl near the species upper distributional limit (table 1). At higher-elevation sites Pinus brutia dominates closed- to open-canopy forests, treeline, and parkland, while at lower-elevation sites it is relatively sparsely distributed within savannas, grasslands, and shrublands.
3. Materials and methods
3.1. Tree ring data
We examined eight residual tree ring width chronologies from Cyprus, developed from tree core samples collected from Pinus brutia trees over the period 2003–2013. Three of the chronologies are used and described by Touchan et al (2014a), where sampling technique, sample preparation, dating, and chronology development procedures are reported. The same methods were used to develop five additional chronologies used in this study (table 1). Briefly, two increment cores per tree were collected at breast height from 20−30 mature Pinus brutia trees, and were surfaced and crossdated using standard dendrochronological techniques (Stokes and Smiley, 1968). Each measurement series was fit with a cubic smoothing spline with a 50% frequency response cutoff equal to 67% of the series length to remove trends unrelated to climate, and a low-order autoregressive model to remove additional persistence (Cook and Kairiūkštis, 1990). Tree ring chronologies were developed using a biweight robust estimate of the mean, and their lengths were truncated where an expressed population signal value of 0.85 indicated a sample size inadequate for approximating the true population tree ring signal at the sample site (Cook and Kairiūkštis, 1990, Wigley et al 1984).
Table 1. Tree ring sample site and chronology information.
| AMB | 34.94 | 32.92 | 1489 | 1552–2010 | 36/72 |
| STP | 35.02 | 32.63 | 1059 | 1741–2002 | 20/38 |
| AMF | 35.00 | 32.63 | 859 | 1718–2010 | 21/42 |
| HIK | 35.30 | 33.26 | 795 | 1791–2012 | 22/44 |
| ALK | 35.28 | 33.55 | 489 | 1898–2012 | 22/41 |
| DEG | 35.26 | 33.44 | 243 | 1947–2012 | 20/39 |
| YEK | 35.49 | 34.19 | 231 | 1915–2012 | 20/38 |
| HAO | 35.28 | 33.02 | 181 | 1924–2012 | 20/39 |
3.2. NDVI data
The NDVI is an estimate of the fraction of photosynthetically active radiation absorbed by Earth's vegetation (Sellers 1985) that has been extensively used to monitor global productivity over the past three decades (Running et al 2000). The satellite-derived Global Inventory Modeling and Mapping Studies (GIMMS) NDVI product generated from the Advanced Very High Resolution Radiometer provides the longest continuous NDVI record and is typically used for long-term vegetation studies (Tucker et al 2005). Despite its relatively coarse spatial resolution (approximately 8 km grid cells), GIMMS generally agrees with vegetation patterns estimated by more modern and better spatially-resolved NDVI datasets (Guay et al 2014) and in-situ measurements (Vicente-Serrano et al 2016). However, datasets from the first GIMMS version (GIMMSg data period 1981–2006) (Tucker et al 2005) and its successor (GIMMS3g data period 1981–2015) (Pinzon and Tucker 2014) are known to differ significantly in some instances, particularly in arid and mountainous regions, shrublands, and grasslands (Fensholt and Proud 2012, Guay et al 2014, Scheftic et al 2014). The GIMMSg data were also developed using lower-latitude calibration targets that may be more relevant to our study than the high-latitude calibration targets largely used for GIMMS3g (Xu et al 2013). We compared both GIMMS datasets at each study site with 1 km Moderate Resolution Imaging Spectroradiometer (MODIS) (DAAC ORNL 2008) NDVI estimates, considered the best-calibrated and highest quality record currently available (Guay et al 2014). Over the six common years of data (2000–2006), and consistent with the findings of Guay et al (2014), the closest agreement is between the means and standard deviations of the GIMMSg and MODIS data and the GIMMSg record was therefore used in our analysis.
3.3. NDVI, tree ring, and climate relationships
Exploratory Pearson's product moment correlation tests were used to compare each tree ring chronology with the nearest grid point of monthly and seasonal (averaged) mean NDVI (Tucker et al 2005). Correlations were tested over periods ending in each month of the 14 month period beginning in August of the previous year and ending in September of the current year. A seasonal NDVI record was developed to represent mean vegetation activity during the month or season when it is most strongly correlated with tree ring width records at the greatest number of sites. Monthly NDVI data were standardized as z-scores, and these z-scores were averaged across the target season.
Radial tree growth and seasonal NDVI records were then compared against fluctuations in monthly precipitation, temperature, and drought. First, tree ring chronologies were compared with monthly gridded NDVI, climate, and drought index records estimated on each study site location, in every month from August of the previous year to September of the current year, using the program SEASCORR (Meko et al 2011). For the climate data, correlations with Global Precipitation Climatology Centre v6 0.5° precipitation (Becker et al 2013, Schneider et al 2014) were calculated (data period 1901–2013), and partial correlations were then used to identify any additional influence of seasonal mean Climatic Research Unit TS3.1 0.5° temperature (Mitchell and Jones 2005) on radial tree growth independent of the precipitation influence (data period 1901–2012). For the drought data, correlations with SPEIbase v2.4 0.5° gridded values of the standardized precipitation evapotranspiration index (Vicente-Serrano et al 2010, SPEI) were calculated (data period 1901–2014). The SPEI is a metric of drought that accounts for precipitation as well as changes in evaporative demand caused by temperature variations. The common data period of the climate and SPEI data with tree ring chronology records ranges from 89–112 years. Second, to determine if linkages between tree rings and NDVI are driven by climate, the seasonal NDVI record was also compared with climate and SPEI data in the same manner as above. SEASCORR (Meko et al 2011) computes confidence intervals for correlations and partial correlations by a Monte Carlo method (Dietrich and Newsam 1997), and tests temporal stability of relationships by a difference of correlations test (Snedecor and Cochran 1989) for which the sample size is adjusted downward as needed to account for persistence in the time series. Throughout this study, correlations and partial correlations were calculated over the maximum common data period. MODIS Land Product Subsets classifications (2.25 km2 resolution) (DAAC ORNL 2008) were used to determine land cover composition at each sample site. The most common two classes from the 25 classified pixels per grid cell are reported.
4. Results
All of the lower-elevation Cyprus chronologies (<800 m asl) except HIK are significantly (p < 0.05) positively correlated with NDVI during the winter precipitation season, with the highest seasonal correlation occurring during February through May (FMAM; figure 1; table 2; figure 2). The dominant, favorable influence of December precipitation on both FMAM NDVI and radial tree growth at lower-elevation sites is evident in the monthly SEASCORR analysis (figures 3(a) and (c)). These chronologies are also negatively correlated with NDVI at some time during the autumn prior to growth (October–December; figure 2). FMAM NDVI and tree ring chronologies at lower-elevation sites generally exhibit additional significant negative correlations with winter temperature that are independent from the influence of precipitation (figures 3(b) and (d)). Monthly correlations with SPEI reaffirm that, overall, FMAM NDVI and radial tree growth at lower elevation sites are negatively influenced by drought during December (figure S1, available at stacks.iop.org/ERL/12/084008/mmedia). FMAM NDVI and tree rings are also negatively correlated with SPEI in the late spring/summer (figure S1). The lower-elevation sites are mainly characterized by shrublands (shrub canopy cover between 10%–100%) and grasslands(<10% tree or shrub canopy cover; table 2), conditions are dry and warm, and the period of maximum annual NDVI occurs over winter/spring (figure 4).
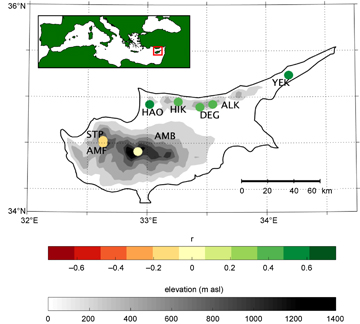
Figure 1 Map of correlations between Cyprus tree ring chronologies and nearest gridded February through May NDVI. An inset map shows the location of Cyprus within the broader Mediterranean region.
Download figure:
Standard image High-resolution imageTable 2. Correlations of tree ring chronologies with FMAM NDVI, and vegetation cover information.
| AMB | −0.06 (0.78) | woody savanna | 84 | closed shrubland | 8 |
| STP | −0.17 (0.46) | woody savanna | 92 | evergreen needleleaf forest | 8 |
| AMF | −0.10 (0.62) | woody savanna | 100 | − | − |
| HIK | 0.35 (0.09) | grassland | 44 | closed shrubland | 28 |
| ALK | 0.42 (0.04) | open shrubland | 68 | woody savanna | 20 |
| DEG | 0.39 (0.05) | grassland | 80 | closed shrubland | 12 |
| YEK | 0.58 (0.00) | woody savanna | 48 | grassland | 44 |
| HAO | 0.54 (0.01) | closed shrubland | 49 | open shrubland | 28 |
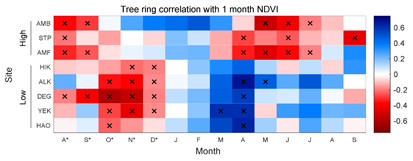
Figure 2 Monthly correlations of tree ring chronologies with NDVI. Tree ring sample sites are ordered from high (top) to low (bottom) elevation, and the NDVI data come from the grid cell nearest to each sample site. Significance (p<0.05) indicated by x.
Download figure:
Standard image High-resolution image
Figure 3 Correlations and partial correlations of FMAM NDVI and tree ring chronologies with monthly climate variables. NDVI correlations with (a) precipitation and (b) temperature are shown at left and tree ring correlations with (c) precipitation and (d) temperature at right. Tree ring sample sites are ordered from high (top) to low (bottom) elevation, and the NDVI and climate data come from the grid cell nearest to each sample site. Significance (p < 0.05) indicated by x.
Download figure:
Standard image High-resolution image
{kind=link}
{kind=link}
{kind=link}
Figure 4 Vegetation activity and climate characteristics of the study sites over the period 1981–2006. (a) and (b) Study site climographs. On each box, the central mark indicates the median, and the bottom and top edges of the box indicate the 25th and 75th percentiles, respectively. The whiskers extend to the most extreme data points not considered outliers, and the outliers are plotted individually using 'o' symbols. (c) Time plots of mean monthly NDVI. (d) Time plots of monthly NDVI variance.
Download figure:
Standard image High-resolution image{kind=link}
The higher-elevation chronologies (>800 m asl) exhibit different relationships with NDVI and climate than the lower-elevation chronologies. Correlations with NDVI are negative, occur during a different seasons (prior August–September and May–June), and are weaker, than at lower-elevations (table 2; figure 2). Only AMB is significantly correlated with NDVI, and in a different season (August through October, r = −0.67, p < 0.01; figure 1). FMAM NDVI at the higher-elevation sites is significantly negatively correlated with precipitation during May (figure 3(a)) and, independently, significantly negatively correlated with temperature during April (figure 3(b)), however radial tree growth is not significantly influenced by precipitation or temperature in a consistent way among these sites (figures 3(c) and (d)). Both FMAM NDVI and radial tree growth at higher-elevation sites are significantly negatively correlated with SPEI in summer and winter, respectively (figure S1). The higher-elevation sites are substantially tree covered and mainly characterized by woody savannas (40%–60% tree cover), with wetter and cooler conditions and higher but much less variable annual NDVI than lower-elevation sites (figure 4). All significant SEASCORR correlations reported in this study are stable over comparison periods.
5. Discussion
Our results point to the potential of tree rings as a proxy for grassland and shrubland primary productivity in low-elevation and/or moisture-controlled regions, and possibly more broadly within the global Mediterranean biome. In these environments both radial tree growth and grassland and shrubland vegetation activity are strongly controlled by winter drought, but at different times. Winter precipitation governs coincident green-up in grass- and shrub- dominated ecosystems, and also controls soil moisture quantities and cambial activity in Pinus brutia later in spring. These findings are consistent with prior studies that document the importance of winter water availability in determining shrubland and grassland phenology (Papanastasis 1997), and spring water availability in determining tree ring width and the properties of tracheids in Mediterranean pines (Campelo et al 2013, DeLuis et al 2013, Touchan et al 2014a, Vieira et al 2010, 2014) and other conifers (Lebourgeois et al 2012).
The timing of the annual FMAM NDVI-tree ring moisture sensitivity is remarkably similar among lower-elevation sites (figure 3(a) and (c), figure S1). December, the month of greatest mean monthly precipitation, strongly significantly influences both NDVI and tree rings, and likely plays a major role in regulating annual and seasonal drought (figure 3(a)). At the grassland and shrubland sites, FMAM incorporates the period of maximum NDVI and is strongly correlated with mean annual NDVI (r = 0.68–0.92, p < 0.01) such that tree ring based reconstructions of vegetation activity during that season have the potential to inform on annual productivity patterns over time (figure 4(c)).
Both FMAM NDVI and annual tree growth are also negatively influenced by warm winter temperatures at the lower-elevation sites. The temperature and precipitation data at those sites are not significantly intercorrelated during February through March, indicating partial temperature correlations are not artifacts of covariance between the climate datasets. The influence of temperature on NDVI may result from warm temperatures exacerbating moisture deficit conditions and causing productivity declines in grasses and shrubs (Papanastasis 1997). Cambial processes of Mediterranean Pinus may be active during winter and could be negatively affected by warm temperatures through enhanced transpiration and internal water stress caused by warm and dry winds (Liphschitz et al 1984). The (negative) sign of the correlations with temperature indicates they do not drive the (positive) FMAM-NDVI connection.
Correlations with SPEI offer another perspective on precipitation and temperature interactions, and their influence on lower-elevation FMAM NDVI and radial tree growth. Similar to the precipitation and temperature results, FMAM NDVI and tree rings at these sites are significantly negatively influenced by drought during December, although relationships with SPEI are relatively weaker than those with climate data (figure S1, figure 3). Since this drought index integrates both precipitation and temperature information, weaker correlations with December SPEI might suggest that precipitation plays a disproportionately important role in controlling productivity and radial tree growth at lower-elevation sites. A similar phenomenon, where tree growth is observed to have a stronger relationship with precipitation alone than with the Palmer drought severity index, is seen throughout the eastern Mediterranean (Touchan et al 2014a). FMAM NDVI at all elevations and tree rings at lower-elevations are also significantly negatively correlated with SPEI in the late spring/summer, though no connection between NDVI and tree rings exists during that season and accounting for a positive influence of summer drought on the green-up of grasses and shrubs in antecedent spring is problematic.
Negative correlations of tree rings with monthly NDVI at both lower-and higher-elevation sites are also difficult to explain (figure 2). In higher-elevation forests, this relationship might arise from within-tree resource trade-offs between canopy and radial growth (Jarvis and Leverenz 1983). We have no explanation for why high(low) grass and shub productivity and moisture during October-December might precondition low(high) radial tree growth in the following growing season. The monthly NDVI data are not significantly autocorrelated at lags >1 at any site.
The type of vegetation driving NDVI measurements at each site is a critical element in the tree ring-NDVI connection. In the arid shrubland and grassland ecosystems, NDVI fluctuations are typically spatially and temporally cohesive, and are strongly determined by and susceptible to drought (figure 4(c)) (Vicente-Serrano 2007). NDVI at the higher-elevation sites is driven by forest canopy productivity, which exhibits smaller relative seasonal and annual changes (figure 4(d)), corresponds weakly with drought seasonality (figure 4(c)) (Vicente-Serrano 2007), and may have indirect, lagged, or absent physiological connections to xylogenesis (Berner et al 2011, Bunn et al 2013). Tree rings and NDVI may not be as strongly linked at higher elevations because those sites are less water-limited, receiving greater moisture from rain and periodically from snowmelt and experiencing more moderate temperatures during FMAM than lower-elevation sites (figures 4(a) and (b)). The mean sensitivity (Cook and Kairiūkštis, 1990) of tree ring chronologies at these sites is lower than at lower elevation sites. Given the very low variance of the higher-elevation NDVI data, however, it is also likely the weaker association between tree rings and NDVI at higher elevations stems from a relatively muted response of forest canopy vegetation (eg. leaf area) to environmental fluctuations compared with grasses and shrubs. Higher-elevation sites examined in this study are occasionally snow-covered, however values of the NDVI at those sites are greater than 0.1 and too large to suggest that variance is dampened by the presence of snow (Tucker et al 2005).
Knowledge of conifer cambial activity in drought-limited environments is incomplete, especially with respect to the influence of climate on the processes comprising xylogenesis. Vieira et al (2014) emphasize that the xylem anatomical response may fluctuate considerably between environments and/or species, for example based on different physiological adjustments for tolerating drought. We did not observe the drought-moderated relationships between tree ring features and late summer and autumn forest canopy productivity that are often documented in the Mediterranean region (Pasho and Alla, 2015), and even observed the opposite (negative) relationship during the same season at the STP site. Nevertheless, many studies describe the dominant role of winter precipitation and temperature in controlling the annual radial growth of Mediterranean Pinus species in dry, low-elevation areas (Cherubini et al 2003, DeLuis et al 2011, 2013, Klein et al 2013, Olivar et al 2012, Sarris et al 2011, Touchan et al 2014a, 2014b, Vieira et al 2010), and despite considerable land use change and disturbance, climate exerts an overwhelming influence on NDVI across the terrestrial Mediterranean (Diodato and Bellocchi 2008). Based on these observations and our results here, we anticipate that Pinus and other drought-sensitive conifers can be used to reconstruct winter vegetation activity in non-forest ecosystems in that region. Such an effort may require filtering or extracting NDVI data to target specific land-cover types. Since the tree ring climate response reflects spring moisture availability, the bimodal spring and autumn growing pattern that commonly produces false rings in Mediterranean water-limited conifers (Camarero et al 2010), as well as earlywood density and wood anatomical features, may offer further opportunities for distilling a tree ring-productivity linkage.
6. Conclusions
Increasing drought stress is expected to strongly influence interannual variability in the global carbon cycle, and profoundly affect the biodiversity, ecosystem functioning, and resilience of Mediterranean grasslands and shrublands under future climate warming (Breshears et al 2005, Kunstler et al 2006, Lloret et al 2016, Poulter et al 2014, Royer et al 2011). A multi-century view of productivity in these environments would provide context for the severity of recent ecosystem changes and carbon cycle shifts, quantifying the timescales at which semiarid ecosystem productivity is affected by drought, and anticipating ecosystem resistance, resilience, and vulnerability to extreme drought events (Vicente-Serrano et al 2013). This study is the first to investigate the relationship between NDVI and tree ring growth in the eastern Mediterranean, and demonstrates that tree ring width is a promising proxy for developing long records of productivity for grassland and shrubland ecosystems and similar water-limited environments across the global Mediterranean biome. At low elevations both FMAM grassland and shrubland productivity and the annual radial growth of Pinus brutia trees are controlled by winter season precipitation. This relationship decouples at higher-elevations where water is less limiting for productivity, and where forest vegetation activity drives NDVI estimates. Our study identifies the link between tree ring records and broad-scale non-forest vegetation activity during two separate seasons. Paleoreconstructions using this approach could be optimized by reducing confounding influences in the NDVI data through filtering based on land-cover type, or by capitalizing on summer cambial suppression and earlywood features to capture the spring moisture signal in low-elevation Mediterranean conifers.
Acknowledgments
We extend our thanks to Wim VanLeeuwen and Kyle Hartfield (Arizona Remote Sensing Center), Pieter Beck (European Commission), Scott Goetz (Northern Arizona University), and Georgia Clyde (University of Victoria, Hyperspectral—LiDAR Research Group) for their assistance accessing and analyzing NDVI data. We wish to thank Mr. Salih Göneş and Dr. Andreas Christou from the Forestry Departments in Cyprus for their support in making this study possible. We thank Alexis H Arizpe and Christopher Baisan for their valuable assistance in the field. Funding was provided by the US National Science Foundation, ATMGEO/ATM-Paleoclimate Program 0758486 and Grant No. 1103314 from AGS-Paleo Perspectives on Climate Change Program.