

Abstract
Marine debris is a growing threat to hundreds of marine animal species. To understand the consequences of marine debris to wildlife populations, studies must go beyond reporting the incidence of wildlife and debris interactions and aim to quantify the harm resulting from these interactions. Tubenosed seabirds are globally threatened, with a near universal risk of debris ingestion and an unquantified risk of mortality due to eating plastics. In this paper, we explore the mortality risk narrative due to the acute effects of debris ingestion, and quantify behavioural and ecological factors including age, diet and foraging method. We examined ingested debris loads, types and mortality of 972 adult and immature seabirds across 17 albatross, shearwater and prion species in a global seabird biodiversity hotspot. Though age and foraging method interact to influence the incidence and number of items ingested, age and diet were the most important factors influencing mortality. Mortality is influenced by debris load and type of debris ingested and there is selectivity for items that visually resemble a seabird's prey. Immature birds that forage on cephalopods are more likely to ingest and die from eating debris than are adults. Conversely, the risk of death to seabirds that forage on crustaceans is linked to the number of plastic items ingested and is higher in adults. Debris ingestion is an under-recognised cause of tubenose mortality and is likely negatively affecting rare and threatened species.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Pollution of the marine environment by plastics and other anthropogenic 'marine debris' is a global problem for marine life, with more than 1400 species known to interact with marine debris [1]. There is particular concern for the potential impact of marine debris on seabirds due to the number of seabird species that are threatened or in decline [2], and their increasingly high frequency of reported debris ingestion [3, 4]. The conversation about marine debris ingestion is evolving from reporting species-specific debris ingestion rates to analysis of risk of harm and mortality in order to quantify population effects and the ecological consequences of debris ingestion [5, 6].
Tubenosed seabirds (Order Procellariiformes) have a global distribution with 144 extant species [7]. Nearly two-thirds (62.5%) of tubenoses are classified as near threatened or threatened with extinction, and they are affected by a number of threats including fisheries interactions, feral animals and climate change [2, 7]. Tubenoses commonly ingest marine debris; 63.2% of species are currently known to ingest plastic [4] and 99% of species are predicted to be affected by plastic ingestion by 2050 [3]. The ingestion of plastic debris is recognised as a likely threat to some seabird populations [6], but because direct mortality is infrequently recorded and population effects from ingesting debris are poorly understood [2, 6], 'garbage and solid waste' is listed as a threat for only 9% of species [7]. Disentangling which species' populations are threatened by debris ingestion begins with identifying the behavioural and ecological factors that may put some species at higher risk than others. Does more debris ingestion directly translate to more death, or do behavioural and ecological factors also influence the risk of mortality?
Age and maturity are factors commonly identified as affecting the rate of seabird debris ingestion [8–12]. Some studies show no difference in the frequency of occurrence of plastic ingestion between age groups in seabirds [13], while other found that juveniles are more likely to ingest marine debris, and ingest more items [8]. Spear and colleagues [12] reported that across seabirds as a whole, plastic ingestion increased with age, but some species had higher frequencies of plastic ingestion in subadults (sooty shearwater, Ardenna grisea, and white-winged petrel, Pterodroma leucoptera). Others found no significant differences between age classes (e.g. Leach's storm-petrel, Oceanodroma leucorhoa, Stejneger's petrel, Perodroma longirostris, and wedge-tailed shearwater, Ardenna pacifica) [12]. Even within an individual species, such as the northern fulmar, Fulmarus glacialis, some studies report that there are no differences in the frequency of debris ingestion between age classes [13], while other studies report that juveniles ingest more debris [8–10].
Juvenile and immature seabirds have higher mortality rates than adults, which has been linked to poor foraging proficiency [14]. Foraging in seabirds is partially an innate behaviour and partially a learned behaviour, as individuals hone their foraging skills with experience [15]. The first months of a seabirds life are critical for its survival [15], which begs the question: are immature birds consequently at greater risk of debris ingestion mortality?
Ecological factors including diet and foraging behaviour may also influence the relationship between debris ingestion and mortality. Though little research currently exists that addresses the relationship between ecological factors and debris mortality directly, diet and foraging behaviour have been shown to drive tubenose debris ingestion [3, 16–19], with increased debris ingestion linked to an increase in the risk of death [20]. But is the link between taxa, debris, and death a simple relationship? Different species selectively ingest different debris items [19, 21], some debris items are more deadly than others [20] and there is mixed evidence about the relationship between age and debris incidence in seabirds.
We aimed to disentangle the relationship between age, diet and foraging behaviour among tubenosed seabird taxa and understand how these factors shape the incidence and composition of debris ingestion, and ultimately influence mortality. Specifically, we modelled the relationship between age, diet, foraging method and taxonomic grouping on debris ingestion incidence, total debris intake, debris-related mortality and ingested debris item composition.
2. Methods
2.1. Carcass collection and determining cause of death
We examined the cause of death (COD) of 972 tubenosed seabirds (Order Procellariiformes) collected across Australia and New Zealand between 2013 and 2017 [16]. We compared the load of ingested marine debris and COD with respect to debris ingestion (table 1). The birds collected represented a combination of beach-washed seabirds (n = 754) and seabirds by-caught in fisheries (n = 218). Beach-washed birds comprised a combination of live birds received and assessed by wildlife veterinarians that were euthanised for veterinary purposes or died in care (usually within 48 h), and those that were collected dead. Of the beach-washed birds, 71.5% (n = 539) were associated with seabird mass mortality 'wreck' events while 28.5% (n = 215) were not wreck-associated.
Table 1. COD determination methodology. Gross necropsy was conducted on all carcasses to assign a COD category with respect to their marine debris ingestion. We recorded acute signs of death only in 'known' categories and did not assess potential chronic causes of body condition.
| Category | Description | Example |
|---|---|---|
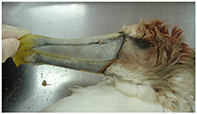
| Known not debris (KND) | Cause of death is identified by gross necropsy (examples include acute physical trauma, severe haemorrhage, necrotic enteritis, severe necrosis of tissue not linked to decomposition, and fresh fracture). Also includes carcasses where the COD is known when carcass is received (fishery by-catch, vehicle/boat strike, received from veterinary clinic where animal history is known). This category includes birds that have died with no ingested debris present irrespective of whether COD is able to be determined. |

|
| Photo: (top) Flesh-footed shearwater, KND, fisheries provided by-catch, drowned, fresh. |

| |
| (mid) Buller's albatross, KND, beach-washed, trauma from fisheries interaction. Fresh. | ||
| (bottom) Shy albatross, KND, beach-washed, severe head trauma. Mild decomposition. |
| |
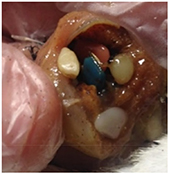
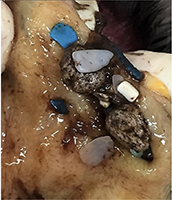
| Indeterminate (Ind) | Plastic ingestion and one or more pathologies where COD cannot be determined with respect to their plastic ingestion. The bird is typically emaciated (includes died during seabird wreck) and sometimes accompanied by other pathology. There are no accompanying signs that ingested debris is acutely linked to death such as obstruction of the gastro-intestinal tract, local inflammation or infection. As gross necropsy only is conducted, it is not known whether ingested debris is chronically linked to death (i.e. dietary dilution). Includes carcasses in old, mummified or heavily decomposed condition with plastic present. This category accounted for most cases of marine debris ingestion. |
|
| Photo: (top) Fairy prion, Ind, beach-washed, emaciated, plastic not associated with gastric obstruction. Fresh. |
| |
| (bottom) Short-tailed shearwater, Ind, beach-washed, emaciated, plastic not associated with gastric obstruction. Moderate decomposition. | ||
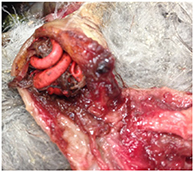
| Known Debris (KD) | Debris is acutely linked to death through either gastro-intestinal obstruction or gastric perforation in fresh and mildly decomposed carcasses. In addition to the obstruction/perforation, the bird is emaciated, there may be accompanied localized inflammation/infection and necrosis. For gastric obstruction, there may be undigested food in the gut (sometimes large amounts), the passage of which is blocked by the obstructing item. The obstructing item may require gentle force to remove with forceps. Subcategory 'Probable debris': Gastric obstruction apparent, but KD cannot be confidently assigned due to at least one of: partial rather than full obstruction, carcass decomposition, obstruction (full or partial) and emaciation but lacking clear gross evidence of local inflammation/infection (this is sometimes linked to carcass decomposition). |
|
| Photo: (top) Light-mantled sooty albatross, KD, beach-washed, fresh. Emaciated, complete gastric obstruction from balloon. Fresh. |

| |
| (bottom) Fairy prion, KD, beach-washed, emaciated, complete gastric obstruction from hard plastic, petri dish contains ingested food that was prevented from passing due to complete gastric obstruction. Mild decomposition. |
To assign COD with respect to ingested debris, we conducted gross necropsies on all birds. Categories were: 'known debris death' (KD): death resulting from debris ingestion, 'indeterminate death' (Ind): bird which had ingested debris, but it was unknown if debris was related to death or 'known non-debris death' (KND): death due to a non-debris cause such as fisheries by-catch, trauma, or starvation with no debris present. A modification to this method included a sub-category of KD to account for probable debris death cases. In these cases, birds appeared likely to have died due to debris ingestion but could not be confidently assigned KD. In this case, they were assigned the COD category 'Probable'. As only gross necropsy was performed, the COD categories were assigned based on acute causes of death only, with potential chronic conditions (plastic and non-plastic related) assigned as 'Indeterminate' (table 1).
2.2. Ingested debris count and categories
We recorded the number of debris items ingested by each bird and recorded the type of marine debris, visually sorting items into seven types; 'hard plastic', 'soft plastic packaging', 'balloon', 'rope', 'fishing debris' (fishing line and/or tackle), 'expanded rubber and foams' and 'other'.
2.3. Seabird taxonomic, diet and foraging grouping
To understand ecological factors, we grouped individuals by taxonomic group, age, diet and foraging method. Birds were separated into three groups by taxonomy; albatrosses (genera Diomedea, Thalassarche and Phoebetria), shearwaters (genera Ardenna and Puffinus) and prions (genus Pachyptila). Birds were aged by gonad development [22], and separated into two age categories: 'immature' (fledged juvenile and immature birds combined) and 'adult'. Only species that were represented by both adult and immature individuals are included in this study. Individual birds that could not be confidently separated into immature or adult were not included in the analysis. We allocated each species to a diet group and foraging behaviour group based on cluster analyses of diets and foraging methods listed in Marchant and Higgins (1990), following the grouping methodology in Roman et al (2019a). Diet groupings (DGs) included: 'cephalopod dominant' (DG1), 'fish dominant' (DG2), 'crustacean dominant' (DG3), 'cephalopod and crustacean' (DG4), 'fish and crustacean' (DG5) and 'fish and cephalopod' (DG6). Foraging groups included: 'diving', 'filtering', 'pursuit', 'surface seizing and plunging' and 'mixed surface foraging'.
2.4. Statistical analysis and model selection
Statistical analyses were performed using R (Version 3.5.3) [23]. We compared the influence of independent variables: age, diet, foraging method and taxonomic grouping against four dependent variables: (a) incidence of debris, (b) total debris intake (ingested debris count), (c) debris-related mortality and (d) debris item composition. For each model tested, we varied combinations of the independent variables (age, diet and foraging method and taxonomic grouping), including testing for interactions between variables. We selected the best model with the lowest Akaike Information Criterion (AIC) value [24]. In the cases where there was more than one model within 2 AIC of the lowest AIC, we chose the simplest model among them (least variables or those without interactions).
Influence of age, taxonomic grouping, diet and foraging behaviour on the incidence of debris ingestion was analysed by applying logistic regression using the R package 'MASS' [25]. The influence of seabird age, diet and foraging behaviour on ingested debris intake were analysed using general additive models, using the R package 'mgcv' [26]. The influence of seabird age, diet and foraging behaviour on chance of death were analysed by ordinal logistic regression using the R package 'MASS'. The order of COD categories used was: debris COD > probable debris COD > indeterminate COD > non-debris COD. Using the best model, we predicted the probability of debris death in well-represented diet groups.
To determine whether the composition of debris types ingested by seabirds differed between groups, we conducted a permutational multivariate analysis of variance using the R package 'vegan' [27]. We determined the Bray–Curtis dissimilarity between types of debris ingested between each adult and immature birds, diet groups and foraging groups, then applied a linear model to the dissimilarity value matrices, using a permutation test with pseudo F-ratios.
3. Results
We examined marine debris ingestion in 972 seabirds to explore the differences among three tubenose groups, albatrosses (N = 243; ad = 222, imm = 21), shearwaters (N = 391; ad = 327, imm = 64) and prions (N = 338; ad = 106, imm = 232) (tables 2 and S1 (available online at stacks.iop.org/ERL/00/0000/mmedia)). DGs included cephalopod dominant (N = 93; ad = 91, imm = 2), fish dominant (N = 8; ad = 3, imm = 5), crustacean dominant (n = 95; ad = 24, imm = 71), cephalopod and crustacean (N = 334; ad = 279, imm = 55), crustaceans and fish (3 sp, 292, ad = 125, imm = 167) and fish and cephalopod (N = 142; ad = 128, imm = 14) (table 2).
Table 2. Assigned tubenose taxonomic and behaviour groups for analysis. This table shows number of individuals within each group, number of adults, immature and species.
| Group | N individuals | Sp. | |
|---|---|---|---|
| Taxonomic | Albatrosses | 243 (ad = 222, imm = 21) | 8 |
| Shearwaters | 391 (ad = 106, imm = 64) | 4 | |
| Prions | 338 (ad = 106, imm = 232) | 5 | |
| Diet | Cephalopod dominant | 93 (ad = 91, imm = 2) | 2 |
| Fish dominant | 8 (ad = 3, imm = 5) | 1 | |
| Crustacean dominant | 95 (ad = 24, imm = 71) | 3 | |
| Cephalopod and crustacean | 334 (ad = 279, imm = 55) | 2 | |
| Crustacean and fish | 292 (ad = 125, imm = 167) | 3 | |
| Fish and cephalopod | 142 (ad = 128, imm = 14) | 5 | |
| Foraging | Diving | 321 (ad = 273, imm = 48) | 1 |
| Filtering | 41 (ad = 16, imm = 25) | 3 | |
| Pursuit | 62 (ad = 49, imm = 13) | 3 | |
| Surface seizing and plunging | 243 (ad = 222, imm = 21) | 8 | |
| Mixed surface foraging | 297 (ad = 90, imm = 207) | 2 |
Foraging groups included diving (N = 321; ad = 273, imm = 48), filtering (N = 41; ad = 16, imm = 25), pursuit (N = 62; ad = 49, imm = 13), surface seizing and plunging (N = 243; ad = 222, imm = 21) and mixed surface foraging (N = 297; ad = 90, imm = 207) (table 2). Diet and foraging grouping were not assigned for one species (N = 8) where diet and foraging behaviour are poorly understood. Bird group, foraging group, diet group, incidence of debris ingestion and total debris intake for each species is detailed in table S1. Details of cases of known and probable death are available in table S2.
3.1. Incidence of debris ingestion
The best model for incidence of debris ingestion included the interaction of age and foraging method in addition to diet (tables S2a and S2b). Adding taxonomic group to this model did not improve explanatory power (table S2a).
There were significant relationships between the incidence of debris ingestion and foraging behaviour. Behaviours associated with a lower incidence of debris ingestion included foraging by filtering (Est = −3.6, Std. Err = 0.8, P < 0.001), by pursuit (Est = −6.8, Std. Err = 1.3, P < 0.001), surface seizing and plunging (Est = −7.1, Std. Err = 1.2, P < 0.001) and mixed surface foraging (Est = −4.2, Std. Err = 0.9, P < 0.001). A crustacean dominant diet was associated with a higher incidence of debris ingestion (Est = 1.5, Std. Err = 0.3, P < 0.001).
Pursuit foraging by immature age individuals (Est = 3.6, Std. Err = 1.2, P < 0.01) and surface seizing/plunging foraging by immature age individuals (Est = 3.4, Std. Err = 1.2, P < 0.01) resulted in a significantly higher incidence of debris ingestion.
3.2. Total debris intake
The best model for the total debris intake included the interaction of age and foraging method, in addition to diet (Adjusted R2 = 0.31, tables S1, S3a and S3b). Adding taxonomic group to this model did not improve explanatory power (table S3a).
Age alone was not an important variable influencing debris intake (P > 0.5). Some foraging strategy groups were associated with lower debris intake, including pursuit foraging (Est = −5.8, Std. Err = 1.2, P < 0.001), filtering (Est = −1.5, Std. Err = 0.5, P < 0.001), surface-seizing and plunging (Est = −5.8, Std. Err = 0.0, P < 0.001) and mixed surface foraging (Est = −2.3, Std. Err = 0.6, P < 0.001).
For interactions between age and foraging method, immature birds that foraged by filtering ingested lower debris loads (Est = −0.9, Std. Err = 0.4, P < 0.05). Immature birds that foraged by pursuit (Est = 3.4, Std. Err = 1.2, P < 0.01), and surface seizing/plunging (Est = 3.4, Std. Err = 0.9, P < 0.001) ingested more debris.
3.3. Cause of death (COD)
Of 972 necropsies conducted, the COD of eleven (1.14%), three adults and eight immatures, were assigned as 'known debris' (KD). The COD for six birds (0.61%), one adult and five immatures, was probable debris ingestion. Most birds, 561 (57.72%) died for reasons not related to debris ingestion (KND, 'known not debris'), 381 adults and 180 immatures. An indeterminant COD was assigned to 386 birds (39.71%); 265 adults and 121 immatures (table S4). Samples were not evenly spread across causes of death or parameters examined, as the 754 carcasses that were sourced from beach-washed individuals were largely prions (n = 337) and shearwaters (n = 386), while the 218 received as by-catch (table 3) were largely albatrosses (n = 212). Only one of eleven debris deaths was associated with large mortality events (seabird wrecks).
Table 3. Sources of carcasses examined and COD assigned with respect to debris ingestion. Gross necropsy was conducted on all birds. Many birds were in an emaciated condition, though the cause of emaciation was not known. Note that some individuals had multiple pathologies.
| COD category assigned with respect to | Origin of carcass | Associated with seabird wreck? | Likely COD | Debris in gut? | Number of carcasses |
|---|---|---|---|---|---|
| Known not debris | Beach-washed | No | Emaciation | No | 87 |
| (KND) | Fisheries interaction | Yes | 1 | ||
| Trauma or disease | No | 5 | |||
| Trauma or disease | Yes | 5 | |||
| Unknown | No | 1 | |||
| NA | No | 8 | |||
| Yes | Emaciation | No | 234 | ||
| Emaciation | Yes | 1 | |||
| Fisheries interaction | Yes | 2 | |||
| Trauma or disease | No | 2 | |||
| Unknown | No | 1 | |||
| By-catch | NA | Fisheries interaction | No | 214 | |
| Fisheries interaction | Yes | 4 | |||
| Known not debris (KND) total: 565 carcasses | |||||
| Indeterminate (Ind) | Beach-washed | No | Emaciation | Yes | 90 |
| Trauma or disease | Yes | 3 | |||
| Unknown | Yes | 2 | |||
| NA | Yes | 1 | |||
| Yes | Emaciation | Yes | 291 | ||
| Trauma or disease | Yes | 3 | |||
| Indeterminate (Ind) total: 390 carcasses | |||||
| Known and Probable debris death (KD) | Beach- | No | Debris ingestion | Yes | 12 |
| Yes | Debris ingestion | Yes | 5 | ||
| Known and probable (KD) total: 17 carcasses | |||||
| — | — | — | — | Total: | 972 |
Species with a cephalopod-dominant diet had 1 KD while 92 were not linked to debris ingestion (KND). Those with fish dominant diets had zero KD deaths, one Ind death and seven non-debris deaths. Those with crustacean dominant diets had two known and three probable debris deaths (57 Ind, 33 KND). Cephalopod and crustacean diets had three known and three probable deaths, 265 indeterminant deaths and 63 non-debris deaths. Among taxa that predominantly eat fish and crustaceans, we observed five KD deaths (226 KND, 61 Ind). Fish and cephalopod diets had no known or probable debris deaths (140 KND, 2 Ind) (figure 1). Birds were reported to have died due to debris ingestion across all three taxonomic groups (figure 2).
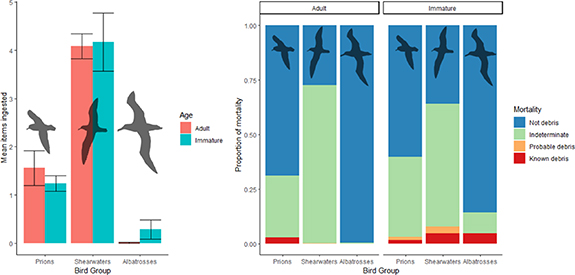
Figure 1. Mean number (±standard error) of marine debris items ingested and mortality in adult and immature tubenosed seabirds across six diet groups following Roman et al (2019a). DGs are cephalopod dominant (DG1, ad = 91, imm = 2), fish dominant (with some cephalopod and crustaceans) (DG2, ad = 3, imm = 5), crustacean dominant (DG3, ad = 24, imm = 71), cephalopod and crustacean (DG4, ad = 279, imm = 55), fish and crustacean (DG5 ad = 125, imm = 167) and fish and cephalopod (DG6, ad = 128, imm = 14). The bar-plot on the left (a) shows the mean of all items ingested, including birds that ingested zero marine debris items. The stacked bar-plot on the right (b) shows the causes of death recorded in each bird group across both adults and immature birds. DG was not assigned for eight individuals where diet is poorly known.
Download figure:
Standard image High-resolution image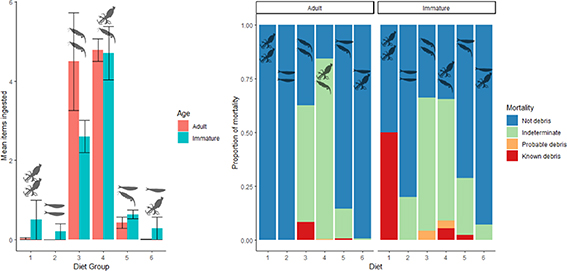
Figure 2. Mean number (±standard error) of marine debris items ingested and mortality in adult and immature tubenosed seabirds across three taxonomic groups; prions (five species, ad = 106, imm = 232), albatrosses (8 species, ad = 222, imm = 21) and shearwaters (four species, ad = 327, imm = 64). The bar-plot on the left (a) shows the mean of all items ingested, including birds that ingested zero marine debris items. The stacked bar-plot on the right (b) shows the causes of death recorded in each bird group across both adults and immature birds.
Download figure:
Standard image High-resolution imageThe best model for COD included the interaction of age, diet and number of items ingested (Best model: table S5a, Model output: table S5b). There was a significant positive effect for the odds of a debris COD with a crustacean dominant diet (Est = 15.7, Std. Err = 9.5, P < 0.05) and a cephalopod and crustacean diet (Est = 17.1, Std. Err = 9.5, P < 0.05). Debris intake had a significant positive interaction with each immature age (Est = 42.7, Std. Err = 9.4, P < 0.001) and fish and cephalopod diet (Est = 14.1, Std. Err = 8.4, P < 0.05), increasing the odds of debris COD.
Odds of debris COD was negatively influenced by the interactions between immature age, debris intake and each crustacean dominant diet (Est = −42.6, Std. Err = 9.4, P < 0.001), cephalopod and crustacean diet (Est = −42.4, Std. Err = 9.4, P < 0.001), fish and crustacean diet (Est = −42.2, Std. Err = 9.4, P < 0.001) and fish and cephalopod diet (Est = −55.8, Std. Err = 9.7, P < 0.001).
Probability of death was predicted for crustacean feeding species, where more representative data occurred (figure 3). For birds with a crustacean dominant diet, the greatest probability of an indeterminate COD occurred at ten (adult) and nine (immature) items, after which the probability of probable or KD death steeply increased (figure 3). After an intake of 26 (adult) and 25 (immature) items there was a 95% probability of KD death. For birds with a cephalopod and crustacean diet, the maximum probability of indeterminate COD occurred at eight (adult) and ten (immature) items. After an intake of 25 (adult) and 26 (immature) items among cephalopod and crustacean feeders, there was a 95% probability of KD death (figure 3). For fish and crustacean feeders, the odds of an indeterminate COD peaks at 13 (adults) and 11 (immature) items, with a 95% probability of KD death at 30 (adult) and 27 (immature) items (figure 3).
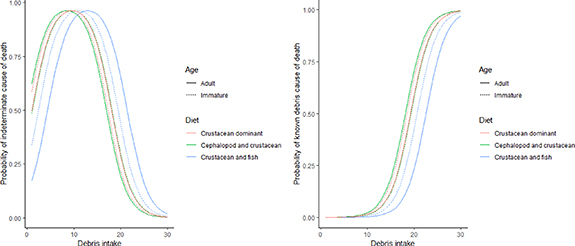
Figure 3. Probability of indeterminate cause of death (left) and KD cause of death (right) for adult and immature tubenose seabirds of crustacean-feeding diet groups according to their debris intake (total number of ingested debris items). Model terms used in this predictive model are displayed in table S5b.
Download figure:
Standard image High-resolution image3.4. Composition of ingested debris
We examined whether there was a difference in the composition of items between the ecological factors and behaviours appearing in the best model for cause of death, age and diet. There were significant differences in the composition of ingested debris types between ages (SumsOfSqs = 2.1, MeanSqs = 2.1, F.Model = 16.4, R2 =0.03, P < 0.001) and diet group (SumsOfSqs = 4.4, MeanSqs = 0.88, F.Model = 6.9, R2 =0.06, P < 0.001) (figure 4).
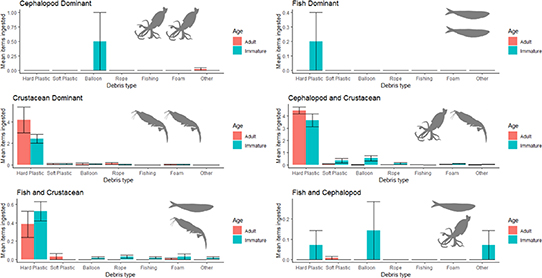
Figure 4. Mean numbers (±standard error) of differing types of marine debris items ingested by immature and adult tubenosed seabirds across six diet groups. DGs are: cephalopod dominant (DG1, ad = 91, imm = 2), fish dominant (with some cephalopod and crustaceans) (DG2, ad = 3, imm = 5), crustacean dominant (DG3, ad = 24, imm = 71), cephalopod and crustacean dominant (DG4, ad = 279, imm = 55), fish and crustacean dominant (DG5, ad = 125, imm = 167) and fish and cephalopod (with scavenging) (DG6, ad = 128, imm = 14). This graph shows the means across all birds examined, including those with no debris in the gut.
Download figure:
Standard image High-resolution imageWhen the composition of debris was assessed with an interaction between age and diet, there was a further significant interaction between ages within diet groups (SumsOfSqs = 1.3, MeanSqs = 0.3, F.Model = 2.5, R2 =0.02, P < 0.05), as shown in figure 4. We found that crustacean diets were associated with a greater diversity of ingested debris across all items than non-crustacean diets. Hard plastic most frequently appeared in diets containing crustaceans as a major component, balloons most frequently appeared in diets containing cephalopods as a major component, and fish dominant diets had the lowest debris incidence overall. Adults of species with crustacean dominant, and crustacean and cephalopod diets ingested more debris, predominantly hard plastic, than did immatures. Among species with fish and crustacean dominant diets, immatures had higher average (mean) hard plastic ingestion, though there was overlap with adult ingestion rates. Among the three diet groups for which crustaceans were not a major component (fish and cephalopod dominant diets), debris ingestion was rare in adults, though immatures ingested a mix of items. Immature birds that had cephalopods as a major component of their diet ingested balloons, though balloon ingestion was not observed in adults of these groups (figure 3).
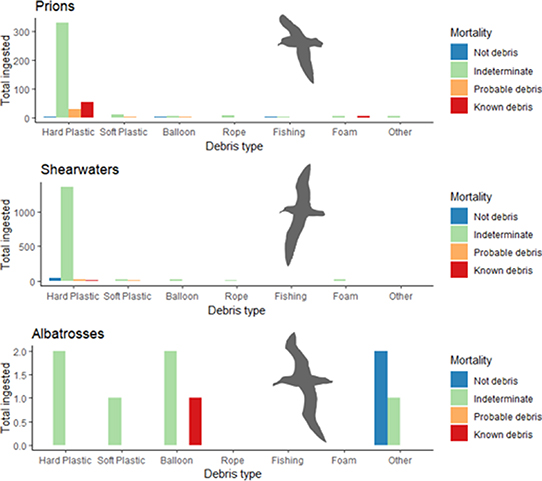
The taxonomic groups investigated in this study contained a mix of diet groups within each. We further assessed differences in debris composition taxonomic groups, finding significant differences in debris composition between taxonomic groups (SumsOfSqs = 2.1, MeanSqs = 0.36, F.Model = 2.8, R2=0.03, P < 0.001) and between adults and immatures within taxonomic groups (SumsOfSqs = 1.2, MeanSqs = 0.3, F.Model = 2.3, R2=0.02, P < 0.05). We found that prions ingested a mix of debris items, but mostly ingest hard plastic, with adults ingesting more hard plastic than immatures (figure 4). When composition of ingested debris and mortality were assessed, the count and type of ingested items occurred inconsistently between mortality categories (figure 5, table S7).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 5. Histogram of counts of items ingested by seabirds across cause of death categories Among albatrosses, there were 239 KND, 3 Ind and 1 KD, among shearwaters, 113 KND, 272 Ind, 3 Probable and 3 KD, and among prions 213 KND, 115 Ind, 3 Probable and 7 KD.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
The risk of debris death among tubenosed seabirds differs according to diet (presence and dominance of cephalopods, fish and crustaceans) and age (adult/immature). The risk of death associated with ingesting different items also differed between seabird groups.
4.1. Disentangling interactions that explain risk of debris death
Tubenose age, foraging method and diet influenced the incidence of ingested debris, supporting observations in previous studies [12, 17, 19]. We found that the highest incidence of ingested debris occurred in birds that foraged at the sea surface, and whose diets included crustaceans and/or cephalopods. Importantly, age and diet influenced the risk of debris death. There was a large improvement in explanatory power between an age/diet COD model with a debris intake addition (AIC 782.8), and a model with a debris intake interaction (AIC 636.5). This demonstrates that the odds of debris death differentially interact with debris intake between diet groups.
Immature age interacted strongly with ingested debris load, suggesting that the risk of debris death increased sharply with debris load among the immature individuals of some species. However, the negative coefficients for interactions (roughly equivalent to the prior positive immature/debris intake interaction) showed that for four diet groups (the three crustacean-feeding groups and fish/cephalopod feeder), the odds of debris cause of death rose consistently with debris intake. For fish and cephalopod feeders, this negative coefficient was greater than the age/sum debris positive coefficient, suggesting that there were increased odds of debris death among immature fish and cephalopod feeders (mostly albatrosses in this study), even when few items of debris were ingested.
The lack of significant interactions among fish dominant and cephalopod dominant diets may result from a lack of debris-ingesting birds sampled in these groups. For this reason, we cannot predict whether the absence of this interaction is real, or due to low sample size. Low sample size affects several categories/sub-categories investigated in this study. Results derived from low sample sizes; for example, fish diet, immatures of the cephalopod diet and debris type preference patterns in albatrosses, should be interpreted with caution.
An important influence on the odds of debris death was the type of debris ingested (figures 4 and 5). Previous studies show that seabirds and other marine wildlife select debris that resembles their prey [21, 28]. This study adds to these observations by demonstrating an influence of both age and diet on selection of debris item types. Furthermore, our results show that the type of debris items selected affects the chance that a bird will consequently die from ingesting debris. Some ingested items, such as balloons, soft plastic, rope and foam, that were found to be ingested by birds with 'probable' and 'known' debris deaths, seldom occurred in birds known to have died from incidental and non-debris causes of death. We propose that there is a lower probability of survival following the ingestion of balloons, soft plastic, rope and foam compared to the other items examined. Conversely, hard plastic commonly occurred among items ingested by birds that did not die from debris, showing that the survivability of eating hard plastic is high, and more so for albatrosses and shearwaters. However, hard plastic occurred more often among birds with an indeterminate cause of death compared to those with a non-debris causes of death (figure 5), which may indicate that hard plastic is linked to mortality. If so, the mechanism is likely chronic rather than acute. Chronic causes of death are not readily observable as a gross finding on necropsy and therefore not identified in this study.
Immature cephalopod-feeding species are more likely to mistake debris for food and subsequently die than adults. These age-related variations likely reflect inexperience and naïve foraging [15], but may also be influenced by differences in the foraging range and/or prey species compared to adults. Conversely, we found higher debris loads and subsequent mortality in adults among species that forage on crustaceans. This study presents evidence of debris selectivity due to (likely visual) prey resemblance among diet groups. The interaction of diet with age resulted in varying patterns of ingested debris item type. We posit that this may reflect learning and potentially a selective pressure on birds reaching adulthood, varying by the type of debris items ingested.
4.2. Diet, debris and death
Specific patterns linked diet and the type and quantity of ingested debris, reflecting seabirds' dietary preferences and influencing the risk of mortality. These patterns included the low frequency of plastic ingestion in piscivores, balloon ingestion in cephalopod feeders and hard plastic ingestion/the large plastic loads ingested by crustacean feeders. The lowest frequency of debris ingestion and subsequent death occurred in species that predominantly ate fish (figure 1), perhaps because no debris type resembles fish in either appearance or behaviour. All species that ate cephalopods as a dominant part of their diet had balloon ingestion recorded among immatures but not adults (figure 3). Inexperienced foraging may influence cephalopod feeders' predisposition to eat (and potentially die from) balloons. Species with crustacean diets had the highest incidence and loads of total debris and hard plastic ingestion (figure 4), occurring in both adults and immatures, potentially due to dietary resemblance. We propose that for crustacean feeders, the risk of death is directly linked to the quantity of plastic ingested. This is because the chance of ingesting an item of a size/shape that causes a gastric obstruction increases as more plastic is ingested. Hard plastic mortalities occurred more frequently in small prions (figure 2 and table S2) than larger seabirds (7 of 338 prions, 3 of 391 shearwaters, 0 of 243 albatrosses). The small 2–10 mm hard plastic available in the environment may pose an obstruction risk in prions while passing through the digestive tracts of the larger shearwaters and albatrosses [29, 30]. Hard plastic deaths may occur more frequently in small crustacean-feeding birds due to the physiology of their proventriculus and gizzard, which is thinner and less muscular than the species that feed on larger prey (Roman, pers. obs.), disproportionately affecting mortality risk in smaller, crustacean-feeding species. For additional discussion, please see supplementary information: debris and diet.
4.3. Ecological relevance and significance for seabird conservation
Tubenosed seabirds face many threats, of which 'garbage and solid waste' is not considered for 91% of species, despite near universal predicted debris ingestion [3, 7]. While debris is not listed as an important threat, solid waste management is likely to be overlooked in conservation and management plans for these species [6]. Many Procellariiformes, including under-studied, rare and threatened species, feed on crustaceans and cephalopods as a major part of their diet [31]. We suggest that debris ingestion is an under-recognised threat to these species. In this study, the cause of death of 4.7% (10 of 215) of all beach-washed seabirds (those not associated with seabird wrecks) was acutely associated with debris ingestion (table S8). The rate of background mortality due to debris ingestion may not have been previously realised because of infrequent post mortem examination of beach-washed seabirds or the common occurrence of plastic in incidentally killed seabirds leading to the perception of low mortality risk. While shot and incidentally dead seabirds (i.e. by-catch, boat strike, car strike) are the most appropriate carcasses to sample rates of debris ingestion in healthy seabird populations [32], marine debris mortality rates can only be appraised among background deaths, for example beach-washed or ocean-floating carcasses, to avoid survivorship bias in cause of death sampling (SI survivorship bias: additional discussion).
In this study, we did not assess chronic causes of debris death, which are difficult (or not possible) to determine by gross post mortem examination. The 'indeterminant' category of death in this study may contain a number of chronic cases. Quantifying deaths due to chronic effects of plastic ingestion is recommended for future studies seeking to better understand plastic ingestion and seabird mortality. Previous work finds ingested plastic affects satiety and nutrition in sea turtles with potentially lethal consequences [33, 34]. Chronic lethal effects of ingested plastic is challenging to study in seabirds, though it is likely that plastic in the gut can influence seabird nutritional status [35, 36]. Lower tissue concentrations of dietary minerals, including manganese, cobalt and iron, has been associated with ingested plastic in storm-wrecked prions, demonstrating a potential link between plastic retained in the gut and seabird nutrition [37]. In extreme or chronic cases, a nutritional disadvantage may be a chronic contributor to death, especially in situations where challenging environmental circumstances push sub-lethally malnourished birds over an energetic knife's edge [37]. For this reason, the ingestion of plastic may be an additive or interacting pressure with broader challenges to seabird survival, such as competition with fisheries for declining fish stocks and the increased frequency of severe weather in a changing climate.
The threat of mortality from gastric obstruction is higher for some items such as balloons, rope and foam, which occur commonly among beach-washed seabirds, but are rarely observed in by-caught seabirds. Identifying which items, when ingested, are associated with low(er) seabird survivability provides policy makers with direct targets to manage the loss to the environment of these items with policy decisions to reduce seabird mortality. To gauge the rate of debris mortality more accurately in seabirds, we recommend that beach-washed wildlife beach patrol programs submit fresh specimens to trained persons for necropsy/post mortem examination to determine the cause of death. We also recommend that seabirds among wildlife hospital casualties are examined to determine the cause of death to better gauge the frequency of debris death.
5. Conclusion
Age and diet are important factors for determining the risk of mortality resulting from marine debris ingestion to seabirds, but the influence of age on risk is not consistent between species. Immature birds are at greater risk of mortality due to debris ingestion among species that eat cephalopods, such as albatross and shearwaters, and these species are more likely to die from high-risk soft debris items, such as balloons. Conversely, risk of death among species that feed on crustaceans, which ingest hard plastic, is linked to the number of items ingested, and is higher in adults. Ultimately, debris ingestion was the acute cause of death of 4.7% of beach-washed Australian and New Zealand albatrosses, shearwaters and prions examined in this study (excluding wrecks). The rate of death due to chronic debris ingestion was unknown. Given this mortality rate, we posit that debris ingestion is an underrecognised source of mortality to tubenosed seabirds, particularly for cephalopod and crustacean feeding species.
Acknowledgments
The authors would like to acknowledge the following Australian and New Zealand organizations who have supported this seabird sampling for this project, provided facilities and storage, and assisted with the collection of, veterinary examination of and care of seabirds used in this study: Auckland War Memorial Museum, Australia Zoo Wildlife Hospital, Australian Museum, Australian Seabird Rescue Inc, BirdLife Australia, BirdLife Tasmania, Bonorong Wildlife Sanctuary, Currumbin Sanctuary Wildlife Hospital, Department of Conservation (New Zealand), Department of Environment and Science (QLD Australia), Department of Primary Industries, Parks, Water and Environment (TAS Australia), Department of Parks and Wildlife (WA Australia), Friends of Shorebirds SA, Holistic Vets Tauranga, Melbourne Museum, Moreton Bay Research Station, Ornithological Society of New Zealand, Pelican and Seabird Rescue, Queensland Museum, RSPCA QLD Wildlife Hospital, South Australian Museum, Tasmanian Museum and Art Gallery, Te Papa Museum, University of Queensland, University of Western Australia and Wildlife Management International. The authors would also like to thank the following people who have assisted in seabird collection, necropsy, veterinary care and examination: B. Bauer, L. Behrendorff, B. Bell, B. Bird, T. Bishop, S. Borelli, B. Burford, K. Carlyon, M. Clarkson, P. Collins, B. Cumming, L. Esler, R. Ferris, L. Feasey, K. Fleischfresser, H. Forrest, N. Forrest, A. Gennings, A. Gillet, I. Godbert, P. Harris, G. Heathcote, M. Hines, P. Horton, G. Irons, H. Janetzki, C. Lacasse, J. Lesel, K. Matthews, C. Mischela, H. Patterson, L. Pulo, M. Pyne, M. Rayner, P. Rhodes, H. Rogers, S. Shaw, J. Sladek, L. Short, I. Southey, K. Southwell, D. Stewart, L. Schneider, P. Swift, P. Taylor, A. Tennyson, K. Townsend, L. Tsang and E. Woehler. Funding for the seabird sampling was provided by the BirdLife Australia's Australian Bird Environment Foundation, the Australian Wildlife Society, and the Ecological Society of Australia's Holsworth Wildlife Research Endowment.
Data availability statement
All data that support the findings of this study are included within the article and supplementary information files.
Supplementary data (PDF)
Supplementary data (XLSX)