

Abstract
Extreme warming events can profoundly alter the transmission dynamics of mosquito-borne diseases by affecting mosquito life-history traits (e.g. survival, growth and reproduction). At local scales, temperatures are determined largely by vegetation structure and can be dramatically altered by drivers of land-use change (e.g. forest conversion). Disturbance activities can also hinder the buffering capacity of natural habitats, making them more susceptible to seasonal climate variation and extreme weather events (e.g. droughts). In experiments spanning three years, we investigated the interactive effects of tropical forest conversion and climate on fine-scale temperature, and the consequences for mosquito larval development. This study was conducted in the northern Malaysian Bornean state of Sabah using local Aedes albopictus mosquitoes; important vectors of dengue, chikungunya and Zika viruses. We demonstrate that variation in temperatures due to forest conversion dramatically increases development rates in Ae. albopictus mosquitoes. However, this effect was mediated by an El Niño Southern Oscillation (ENSO) drought event. In normal years, mean temperatures did not differ between land-use types, however mosquitoes reared in oil palm plantations typically emerged 2–3 days faster than in logged forests. During an ENSO drought, mean temperatures did differ between land-use types, but surprisingly this did not result in different mosquito development rates. Driving this idiosyncratic response may be the differences in daily temperature fluctuations between the land-use types that either push mosquito larvae towards optimal development, or over the thermal optimum, thereby reducing fitness. This work highlights the importance of considering the synergistic effects of land-use and seasonal climate variations for predicting the thermal response of a key mosquito life-history trait driving disease transmission dynamics.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Extreme warming events can influence the distribution and dynamics of vector-borne disease transmission (Messina et al 2015, Vincenti-Gonzalez et al 2018). Inter-annual climate variations, such as the El Niño southern oscillation (ENSO), have been linked to outbreaks of dengue (Vincenti-Gonzalez et al 2018) and malaria (Hashizume et al 2009, Chaves et al 2012) across the globe. Dramatic changes to local climates driven by land conversion (e.g. deforestation) are also associated with altered transmission dynamics of mosquito-borne diseases (Conn et al 2002, Fornace et al 2016). However, at both scales the relationships between extrinsic drivers and patterns of disease remains unclear (e.g. Johansson et al 2009), highlighting the need for better understanding of the interactions between the environment, mosquitoes, and pathogen transmission.
Composite metrics of mosquito-borne disease transmission are typically derived using mosquito functional or life-history traits (henceforth used synonymously). For example, vectorial capacity, which estimates the number of potentially infectious bites arising from all mosquitoes biting a single human on a single day, incorporates parameters relating to mosquito survival, adult feeding behaviour and vector competence. Small changes in these key parameters can profoundly alter transmission dynamics (Garrett-Jones 1964). As mosquitoes are small-bodied ectotherms, these life-history traits are all sensitive to ambient temperature (Lyimo et al 1992, Delatte et al 2009). The thermal sensitivity of a life-history trait is often summarised as a nonlinear asymmetric curve, comprising a thermal optimum (Topt) at which the performance rate is maximised, and critical minimum (CTmin) and maximum (CTmax) temperatures, at which performance is zero (Huey and Kingsolver 1989, Angilletta and Angilletta 2009). Typically, the thermal sensitivity of mosquito life history traits is assessed experimentally, under constant temperature conditions (Alto and Juliano 2001, Bayoh and Lindsay 2003). In contrast, wild mosquitoes encounter temperature fluctuations as great as 20 °C in many normal transmission settings (Paaijmans et al 2009).
Drivers of extreme warming events, such as ENSO and land-use change increase daily temperature fluctuations even further (Luskin and Potts 2011). Importantly, regional climate and local weather interact nonlinearly to determine the temperatures to which mosquitoes are exposed (Stenseth et al 2003). At local scales, vegetation cover modifies solar radiation, air and soil temperature, rainfall, air humidity and wind, to create a microclimate (Aussenac 2000). In forests, dense canopies absorb relatively high amounts of solar radiation producing cool, less variable microclimates (Hardwick et al 2015, Kovács et al 2017). Disturbance activities that reduce canopy cover (e.g. selective logging or forest conversion) profoundly alter microclimates, increasing mean and maximum temperatures and decreasing humidity (Luskin and Potts 2011, Meijide et al 2018). Importantly, the effects of vegetation cover on microclimate are mediated largely by the general climate (Aussenac 2000) and different land-use types vary in their buffering capacity against extreme climate events (e.g. ENSO).
Identifying the effects of microclimate on mosquito population dynamics and pathogen transmission is important. A growing body of work has found that fluctuating temperatures differ considerably in their effects on mosquito life-history traits when compared to equivalent constant mean temperatures (Lambrechts et al 2011, Carrington et al 2013a, Carrington et al 2013b). In general, fluctuations at lower mean temperatures act to speed up rate processes, whereas the opposite occurs at high mean temperatures (Paaijmans et al 2010). As a consequence, models that use averaged temperatures and fluctuations collected over coarse spatiotemporal scales may fail to accurately predict the effects on mosquito life-history, and consequently on the dynamics of mosquito-borne disease transmission. This has been demonstrated for larval development under laboratory conditions, where large fluctuations around a low mean temperature reduced development time by approximately five days compared to constant temperature at the same mean (Carrington et al 2013a). Given that mosquito population dynamics are determined in part by the rate at which new adult mosquitoes are produced (Garrett-Jones 1964), and that larval development time is linked to a suite of adult traits relevant to vectorial capacity (Alto et al 2008, Araújo and Gil 2012, Zirbel et al 2018), understanding how this life-history trait responds to ecologically realistic temperature fluctuations is critical to effective vector and disease control.
In a series of experiments conducted across a tropical agro-forest landscape, we investigated the effects of temperature variation on mosquito life-history. We used local Asian tiger mosquitoes, Aedes albopictus, a species native to Southeast Asia but that has spread rapidly throughout the globe, and is a vector for over 22 known arboviruses, including dengue, chikungunya and Zika viruses. Specifically, we ask: (1) how does the conversion of tropical forest to cropland alter temperatures to which mosquito larvae are exposed; (2) how does an ENSO drought affect these temperatures, and (3) how do land-use change and climate interact to affect mosquito larval development ?
2. Methods
2.1. Sampling sites
Studies were carried out annually at the Kalabakan Forest Reserve (4°33'N, 117°16'E) in Malaysian Borneo, between January and April 2016–2018. The site forms part of the Stability of Altered Forest Ecosystems (SAFE) project, a large-scale deforestation and forest fragmentation experiment comprising dipterocarp tropical forest and oil palm plantations (Ewers et al 2011). Forested sites are grouped into 14 sampling blocks, ranging from undisturbed primary forest to salvage logged forests, the latter of which are in the process of being converted into a fragmented agricultural landscape. The salvage logged forest sites have undergone multiple rounds of selective logging where medium to large commercial timber trees were removed once in the 1970s and one to three times from 1990–2008 (mean aboveground biomass; ABG, of trees with DBH ≥ 10 cm across all forest plots = 95.4 ± 72.8 t ha−1; ± standard deviation, SD; Pfeifer et al 2016; Riutta et al 2018. Differences in available timber and topography have resulted in a high degree of fine-scale spatial variation in logging intensity, and canopy cover ranges from 69.9% to 75% in salvage logged sites. Average tree height in logged forests is 8.9 m ± 2.31 (SD), and tree density is 336 ± 55 trees ha−1 (Ewers 2018). Oil palm plantations were established as monocultures in 2006 (Ewers et al 2011), and have considerably lower biomass than the forest sites (ABG = 38.1 ± 25.2 t ha−1 (±SD); Pfeifer et al 2016). The plantations are characterised by sparse understory vegetation (123 ± 8.32 trees ha−1) and low, open canopies (average tree height = 1.36 ± 1.26 m, SD; Ewers 2018). Forest and plantation sites are separated by approximately 15 km, but have been selected to minimise variation in altitude (mean = 460 m; median = 460 m; interquartile range = 72 m). We randomly selected study sites from established SAFE project second-order sampling points in an oil palm plantation (OP2) and salvage logged forest (Fragment B). Sampling points are comprised of three points arranged 178 m apart in a fractal pattern. For further details on the SAFE project sampling design, see Ewers et al (2011). Climate in the region is typically aseasonal (Walsh and Newbery 1999) with occasional droughts that, are less frequent but more severe in Eastern Borneo than in other regions of the island, and are sometimes but not always associated with the positive phase of ENSO events (Walsh 1996). However, the 2015–2016 El Niño event was amongst the strongest in historical records (Null 2016, L'Heureux et al 2017) and resulted in a severe drought in Borneo (Meijide et al 2018).
2.2. Larval development experiments
Mosquito eggs were collected every year from a single logged forest site (4°33'N, 117°16'E) and stored on paper towels until a sufficient number had been amassed. All eggs were less than two months old at the time of hatching. Eggs were hatched at the field camp by submerging egg papers in water for 24 h, after which any unhatched eggs were discarded. The 1st instar larvae were distributed in rearing tanks (N = 3 tanks per site with 50 larvae per tank) to sites in oil palm plantation (N = 9) and in logged forest (N = 9) land-use types. Rearing tanks were comprised of 330 ml plastic cups (80 mm × 103 mm) covered with mesh cloth, and were placed in shaded areas at each sampling point. To minimise their risk of being knocked over by wildlife, tanks were buried ∼3 cm into the ground. Rearing tanks were visited every 1–2 d and larvae were provisioned with approximately five Tetra Cichlid Colour™ fish pellets every 2 d. Pupae were removed and hatched at the field camp, and the number of development days (taken as days from egg hatching to emergence) and the sex of emerging adults were recorded. Experiments were repeated once per year between 2016 and 2018.
2.3. Temperature data
Air temperature was recorded every 30 min for the duration of each experiment, using a Data Logger EL-USB-2 (LASCAR electronics, Salisbury). Loggers were secured to small wooden stakes at a height of 5 cm, and placed with the rearing tanks at each sampling point. The loggers were shaded from direct sunlight and rain with plastic plates suspended approximately 30 cm above each post.
3. Analysis
3.1. Land-use mediated temperature
Temperature data were filtered to remove outliers due to instrument failure. Outliers were identified as points falling outside 1.5× the interquartile range of daily temperatures for each site (see supplementary material (S1) for summary of outliers). All analyses were carried out in R Version 3.5 (R Core Team 2014, http://R-project.org). Generalised linear models with Gaussian errors (package 'lme4'; link = log; Bates et al 2012) were used to investigate the effects of land-use type and ENSO drought event on mean temperature and daily temperature range, and Gamma errors were used for minimum and maximum temperature. Both land-use type and ENSO drought were treated as categorical variables. Models selection was carried out using stepwise selection and Chi-squared statistic. Post hoc significance tests using the glht function (package = 'multcomp'; Hothorn et al 2008) were used to compare treatment effects and their interactions.
3.2. Probability of mosquito emergence
Mean mosquito development times were compared between the different land-use types for an ENSO and non-ENSO years using Wilcoxon–Mann–Whitney tests. To investigate the effects of temperature and land-use type on mosquito larvae development, time-to-event models (Cox proportional hazards; package 'survival'; Terry 2012) were used. Proportional hazards models are a semi-parametric regression method that analyse the effect of explanatory variables on a hazard rate, defined as the instantaneous risk of an event occurring, given that it has not occurred up until that time. Here, event was defined as day of adult mosquito emergence, taken as the number of days from hatching of eggs. For this study, higher hazard rates thus denote earlier mosquito emergence times.
Due to high correlation between ENSO and the temperature variables, we retained ENSO alone in the model along with the uncorrelated land-use type, and mosquito sex. Model selection was achieved using Akaike Information Criterion and Chi-squared tests were used to ensure that hazard functions were proportional over time for each treatment.
4. Results
4.1. Effect of ENSO drought and land-use type on temperature
Land-use type and ENSO drought both significantly affected the temperature to which mosquito larvae were exposed (figure 1). In non-ENSO periods, average daily temperatures in logged forest and oil palm plantation sites did not differ significantly (GLM, F = 12.37, p = 0.57; figure 2(A)). While the average daily temperature did not differ during this period, the daily temperature ranges at oil palm sites were greater (figure 2(A)) due to higher temperature fluctuations (GLM, F = 13.17, p = 0.003; figure 1). Maximum temperatures were higher in oil palm plantation than in logged forest sites during the ENSO period (GLM, F = 6.41, p = 0.02), however minimum temperatures did not differ significantly between land-use types (GLM, F = 1.11, p = 0.38) in either period.
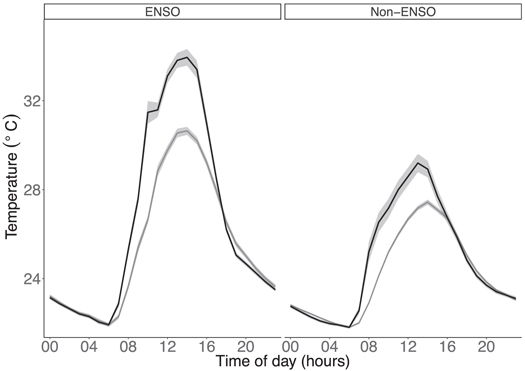
Figure 1. Daily diurnal temperature cycles across the two land-use types. Mean hourly temperature in logged forest (grey line) and oil palm plantations (black line) during a non-ENSO and ENSO period. Shaded areas denote standard errors.
Download figure:
Standard image High-resolution image
{kind=link}
Figure 2. Summary of land-use effects on microclimate and on mosquito larval development. (A) Mean temperatures in logged forest and oil palm plantations; (B) Mean development time of Ae. albopictus larvae (measured as number of days from eggs hatching to adults emerging), vertical boxplot lines represent values within 1.5× the interquartile range of the upper and lower quartiles, horizontal boxplot lines indicate the quartile values, large points denote means for logged forest (grey circles) and oil palm (black diamonds), small points are outliers; (C) Conceptual thermal performance curve demonstrating how temperature fluctuations in logged forest (grey circles) and oil palm plantations (black diamonds) may affect larval development rate (CTmin = critical minimum, CTmax = critical maximum and Topt = thermal optimum).
Download figure:
Standard image High-resolution image{kind=link}
During the ENSO drought, average daily temperatures increased significantly across all land-use types (GLM, F = 59.36, p < 0.001). However, temperatures in oil palm plantations were higher and increased by more than twice as much than in logged forest, resulting in a higher average daily temperature (figure 2(A)). Again, daily temperature range was significantly greater in oil palm plantations than in logged forests (GLM, F = 13.17, p = 0.003). Average maximum temperatures were significantly greater in oil palm plantation than in logged forest, with oil palm plantations experiencing peak temperatures of 7 °C warmer on average than in logged forest (GLM, F = 5.73, p = 0.03). Daily minimum temperatures did not differ by land-use type during the ENSO drought (GLM, F = 0.83, p = 0.38). In both land-use types, average maximum temperatures were significantly greater during the ENSO drought than during non-drought periods (GLM, F = 44.81, p < 0.001).
4.2. Effect of temperature on development time of Aedes albopictus mosquito larvae
Mosquito larval development was significantly affected by local temperature and by ENSO (figure 2(B); table 1). During non-drought periods, the duration of larval development was longer in logged forests (Mann-Whitney U, W = 119 450, p < 0.001). In logged forests development was 16.8 ± 0.29 (SE) days for females and 14.2 ± 0.25 (SE) days for males, while in oil palm average development was 14.6 ± 0.25 (SE) days for females and 11.4 ± 0.22 (SE) days for males. The regional climatic shift associated with the ENSO drought decreased the average emergence time for mosquitoes reared in both land use types. During the ENSO drought there was no difference in the development time of larvae reared in different land use types (Mann-Whitney U, W = 7282, p = 0.15). In logged forests females took 10.9 ± 0.21 (SE) days to emerge and males took 9.29 ± 0.20 (SE) days. In oil palm plantations females took 10.3 ± 0.25 (SE) days and males took 9.09 ± 0.25 (SE) days to emerge.
Table 1. Coefficients of Cox proportional hazards survival analysis estimating effects of ENSO, sex and land-use type on the timing of adult mosquito emergence. Regression coefficients and hazard ratios (exponentiated coefficients) are shown. Positive coefficients imply higher risk; negative coefficients imply the opposite. P values indicate the significance of the coefficients using a Wald z statistic.
| Factors | β | e(β) | se(β) | z | p |
|---|---|---|---|---|---|
| ENSO | 1.96 | 7.08 | 0.12 | 16.42 | <0.001** |
| Land-use type (oil palm) | 0.69 | 1.99 | 0.08 | 8.79 | <0.001** |
| Sex (male) | 0.71 | 2.02 | 0.07 | 10.42 | <0.001** |
| ENSO * Land-use type (oil palm) | −0.59 | 0.55 | 0.15 | −3.85 | 0.0001** |
Results of the Cox proportional hazards model supported the interacting effects of land-use type and ENSO drought on mosquito emergence times (table 1). Mosquito larvae developing during an ENSO drought, that survive to adulthood, experienced a 7 fold increase in the hazard rate, leading to earlier emergence times compared to those in a non-drought period (Cox Regression, z = 16.42, p < 0.001). Developing in an oil palm plantation also significantly increased the hazard rate of mosquito emergence, with mosquitoes 1.99 ± 0.08 times more likely to emerge per day than in logged forest (HR = 1.99; Cox Regression, z = 8.79, p < 0.001). Emergence times were also strongly influenced by sex, with male mosquitoes emerging earlier with a daily emergence probability of 2.0 ± 0.07 times that of females (Cox Regression, z = 10.42, p < 0.001). There was also a significant interaction between ENSO drought and land-use, such that the daily probability of emergence of mosquitoes in oil palm plantations was 0.6 ± 0.15 times less during the drought (Cox Regression, z = −3.85, p = 0.0001).
5. Discussion
5.1. Logged forests buffer the effects of ENSO drought on local temperature conditions
The results of this study indicate that oil palm plantations experience more severe changes in local temperature than logged forests during an ENSO drought event. The greater buffering capacity of logged forests has been identified in a number of other studies (Frey et al 2016, Meijide et al 2018), and is related to greater canopy closure (Renaud et al 2011) and higher leaf area index (Hardwick et al 2015). However, in contrast with a previous study, mean daily temperature in the logged forest and oil palm plantation sites did not differ significantly in non-drought years (Meijide et al 2018). Where average midday temperatures are compared, the differences in mean temperature between forest and oil palm can be even larger (Luskin and Potts 2011, Sabajo et al 2017). The similarity in temperature observed in this study is likely due to the maturity of the oil palm plantations sampled, as older plantations have more closed canopies and more complex understory vegetation structure (Luskin and Potts 2011), and to the history of selective logging which has reduced canopy cover in the forest sites (Ewers et al 2011).
5.2. Land-use and ENSO synergistically affect mosquito development
The effects of land-use type on mosquito emergence times are surprising, given the difference in their temperatures. In normal years, logged forests and oil palm plantations did not differ significantly in mean temperature, however mosquitoes had considerably higher hazard rates in oil palm plantations compared to logged forest. During an ENSO drought, when mean temperatures and daily temperature ranges did differ significantly, mosquito hazard rates were almost identical between the sites. It is worth reiterating that for this study, the hazard rate refers only to adult emergence times and not to mosquito survival. It is possible that other microclimate factors may influence mosquito larval development. For example, relative humidity is expected to be greater in logged forest than in oil palm (Hardwick et al 2015), which could decrease the surface tension of water in the experimental pots (Pérez-Díaz et al 2012) and thereby impact mosquito pupation success (Murdock et al 2017).
An alternative explanation for the absence of a land-use effect on development during the ENSO drought is that the elevated temperatures in the oil palm plantation were high enough to cause stress and reduce the fitness of developing mosquitoes (Feder and Hofmann 1999). Thermal performance curves, which describe the effects of changes in body temperature on physiological sensitivity and fitness, are nonlinear, such that mean performance of a life-history trait under fluctuating conditions may differ to the performance of that life-history trait under mean temperature (figure 2(C); Martin and Huey 2008). Fluctuations that raise temperatures towards a mosquito's optimum temperature are expected to increase performance relative to constant temperature around the same mean. However, fluctuations that push temperatures beyond the thermal optimum may result in a decline in performance (Huey and Stevenson 1979). As the slope of the performance curve is steeper above the thermal optimum, increases in body temperature beyond this are associated with a relatively rapid reduction in fitness (Angilletta and Angilletta 2009).
This would explain the homogeneity of mosquito emergence rates during the ENSO drought despite temperatures differing significantly between the land-use types. Under drought conditions, mosquitoes in both logged forest and oil palm sites would have experienced temperatures closer to their thermal optimum, resulting in the observed faster development rates compared to normal years. However, temperature fluctuations in oil palm were 5 °C greater than in logged forest, potentially exposing mosquito larvae to deleterious temperatures for long enough to reduce growth rates (figure 2(C)). The temperature fluctuations could also explain the difference in larval development in non-ENSO years, when mean temperatures are similar in the land-use types, but greater daily temperature fluctuations in oil palm plantations result in faster larval development times. It is worth reiterating here that our study included only one ENSO event, and that the response of mosquito larvae may vary with the strength and frequency of different drought events. Additionally, whilst we controlled for larval habitat in this study, these sites may vary considerably under natural conditions in the two land-use types; other factors may then be more dominant drivers of larval development rates.
A considerable body of work has been dedicated to elucidating the effects of temperature on the life-histories of mosquitoes (Parker 1952, Lyimo et al 1992, Alto and Juliano 2001, Delatte et al 2009, Paaijmans et al 2011, Couret et al 2014), however key knowledge gaps remain in our understanding of the thermal environment experienced by these small-bodied ectotherms under natural conditions. This study identifies synergistic effects of land-use and climate on the development rates of mosquitoes, highlighting the need to characterise environmental conditions at finer scales than is typically done (e.g. using regional weather stations) to fully understand how environmental change may drive mosquito population dynamics. Importantly, because vectorial capacity is a composite measure of multiple life-history parameters, each with potentially different thermal sensitivities, the effect of temperature on disease transmission will be integrated across these parameters (Martin and Huey 2008). For example, mosquito larval development rate is negatively correlated with adult body size (Mohammed and Chadee 2011), which in turn can have downstream effects on adult survival and feeding behaviour (Nasci and Mitchell 1994, Farjana and Nobuko 2013). Thus, although mosquitoes developing during the ENSO drought emerged earlier than those in a non-ENSO year, they are likely to be smaller, have reduced adult survival (Nasci 1986), and therefore fewer opportunities for host contact over a lifetime. However, due to their limited teneral reserves smaller mosquitoes may take more frequent bloodmeals (Takken et al 1998), which would increase host contacts (Farjana and Nobuko 2013). Importantly, microclimate is one of a many linked predictors of land-use change. Understanding how key environmental drivers (e.g. temperature) mediate these trade-offs in transmission ecology will be critical to prediction of disease transmission and effective disease control.
Acknowledgments
This study was supported by an Imperial College London Grantham Institute for Climate Change Research Science and Solutions for a Changing Planet doctoral training program (SSCP DTP) studentship awarded to NG, and Sime Darby Foundation funding to the SAFE Project. Permission to conduct research was provided by the Sabah Biodiversity Council, Yayasan Sabah, Maliau Basin Management Committee, and the Southeast Asia Rainforest Research Partnership (SEARRP). Logistical support was provided by the SAFE project coordinator, Ryan Gray, and the fieldwork support by the SAFE project field staff. Two anonymous reviewers provided helpful comments on a previous version of the manuscript.
Authors contributions
LC, RE and NG designed the study. NG carried out data collection, conducted the analysis and wrote the manuscript with input from LC and RE. AC provided logistical support. All authors contributed to editing the manuscript. The authors declare no conflict of interest.