

Abstract
In seasonal plant communities, it is recognized that topography-related variation in water availability and solar radiation determine vegetation structure and community composition; however, the effects on functional structure, particularly through changes in resource use strategies of plants are still poorly understood. This study examines the effects of slope aspect and topographic position on functional trait dominance in a tropical dry forest landscape and explores whether strategies for coping with drought (avoidance vs. tolerance) segregate spatially along the water stress gradient created by the interaction of these two topographic factors. The study was conducted in the Balsas river basin in south-central Mexico. Functional traits were evaluated in 63 species of trees (≥ 2.5 cm diameter at breast height) dominant in plots located at three topographic positions (low, medium and high) and on two slope aspects (north and south). Eight leaf and four stem functional traits, relating to the plants' ability to avoid or tolerate water and temperature stress, were measured. Community-level functional traits were strongly affected by topographic position while only a weak signal was detected by the slope aspect. Contrary to our expectations, attributes associated with drought tolerance predominated on the lower topographic positions of the slopes, (moister and warmer sites), while on the upper parts with drier soil, but cooler air, attributes associated with water stress avoidance dominated. In addition, variation in the dominance of leaf pulvini and trichomes along the topographic gradient suggests environmental filtering by elevated air temperatures and water stress, respectively. Overall, our results suggest that the upper topographic positions that generate a shorter and more fluctuating water-availability window, favor readily-deciduous plants with high levels of water storage in their tissues, traits allowing for a rapid avoid of water stress, whereas on the lower topographic positions, where the soil remains moist for longer periods of time, tardily-deciduous species thrive with dense, low-water content tissues, attributes that are associated with physiological tolerance to drought.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence.
Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
In plant communities that develop in highly seasonal climates, topography-related spatial changes in water availability and solar radiation determine important vegetation properties (Gallardo-Cruz et al 2009, Méndez-Toribio et al 2016). In such ecosystems, slope aspect and topographic position (the relative height position along the hills side slope), interact to form a gradient of thermal and water stress for plants across the landscape (Galicia et al 1999, Gallardo-Cruz et al 2009). In particular, in the Northern Hemisphere, the lower parts of the slopes facing north constitute cooler and more humid habitats with lower solar radiation and evapotranspiration rates, and deep soils capable of accumulating runoff water, variables translating into habitats of low risk of drought and thermal damage (Holland and Steyn 1975, Daws et al 2002). In contrast, towards the upper parts of the slopes, where soils are shallow with little capacity for moisture retention and where water table can be more distant, the risk of drought and overheating is high, especially where the slope is oriented towards the south, since exposure to high levels of solar radiation translates into high rates of evapotranspiration (Galicia et al 1999, Wang et al 2011, Ebel 2013). Previous studies have detected the interactive effects of topographic position and slope aspect on the composition of woody vegetation, suggesting environmental filtering processes (Gallardo-Cruz et al 2009, Méndez-Toribio et al 2016). However, the effects of these two factors on community attributes that potentially alter ecosystem functioning, such as functional dominance and functional diversity of species (Violle et al 2007), remain to be explored.
The effects of slope aspect and topographic position on the vegetation have mainly been studied in alpine or sub-alpine, temperate, Mediterranean or semiarid regions (Wales 1972, Armesto and Martínez 1978, Badano et al 2005, Li et al 2011, Elliott and Cowell 2015, Mären et al 2015). While the interactive effect of these drivers on the environment and on plant communities is expected to diminish towards the equator, as the difference in insolation between north- and south-facing slopes decreases, the fact is that there are very few studies at low latitudes, which limits our understanding of the importance of these topographic drivers in tropical landscapes with marked seasonality (Gallardo-Cruz et al 2009, Méndez-Toribio et al 2016).
Plant species of seasonally dry tropical forests face the marked seasonality of rain, high levels of solar radiation and water loss, with morphological, physiological or phenological adaptations allowing them to resist drought and exploit the water when it is available (Borchert 1994, Markesteijn and Poorter 2009, Alvarez-Añorve et al 2012, Méndez-Alonzo et al 2013, Hulshof et al 2014, Wolfe and Kursar 2015). In this community, a trade-off between tolerance and avoidance has been proposed as a major axis of species differentiation (Sobrado 1993, Engelbrecht and Kursar 2003, Tyree et al 2003, Poorter and Markesteijn 2008, Méndez-Alonzo et al 2012). Tolerant species generally present dense tissues in their leaves, stems and roots, and their leaf blades are small and long lasting, frequently composite. They present a xylem that is resistant to hydraulic failure, but with a restricted water conduction efficiency, limiting the capacity of the species to exploit water when it is available, and to realize rapid growth (Pineda-García et al 2015). On the other hand, drought-avoiding species have soft stems with a high water storage capacity, and their leaves are large and short-lived (Méndez-Alonzo et al 2012). Strategies to withstand drought and prevent overheating are key to the process of niche differentiation and the coexistence and dominance of these species is expected to be influenced by topography (Borchert 1994, Pockman and Sperry 2000, Engelbrecht et al 2007, Markesteijn and Poorter 2009, Wolfe and Kursar 2015).
The main objective of this study was to evaluate the effect of slope aspect and topographic position on the functional dominance in a seasonally dry tropical forest landscape. The following specific questions were asked: (i) Is the functional dominance of the community affected by slope aspect and topographic position? (ii) Are drought-coping strategies (avoidance vs. tolerance) spatially segregated along the water and heat stress gradient created by the interaction of these two factors?
It is generally accepted that, in areas of low soil water availability, plant communities will be dominated by desiccation-tolerant species, while in relatively more humid environments, drought-avoiding species of elevated water-use efficiency are expected to thrive (Grime 1977, Chapin et al 1993, Cavender-Bares and Wilczek 2003, Lohbeck et al 2013, Buzzard et al 2015). However, an alternative hypothesis may be more appropriate when considering the dynamics of water availability as dictated by topography. In this case, upper hill shallow soils, with large running off and low water storage capacity on the upper parts of the slopes (particularly the south-facing slopes), may impose a shorter and more variable growth time-window thus favoring opportunistic species that rapidly evade drought by shedding their leaves or maximizing the use of water stored in their tissues (Pineda-García et al 2011, Méndez-Alonzo et al 2012, Méndez-Alonzo et al 2013). A response exacerbated if also distance to water table increases towards the top-hills, as suggested by different hydraulic studies (see Blazkova et al 2002 and references there in) Another hypothesis explored in this study is that in environments where plants face a high risk of overheating (for example, on the lower parts of the south-facing slopes), leaves with attributes that decrease the incidence of solar radiation on their surfaces will dominate.
2. Materials and methods
2.1. Study area and vegetation sampling
Located towards the lower part of the Balsas river basin in south-central Mexico (19° 01' 30.1'' N, 101° 58' 58.1'' W), the study area presents a landscape covered by tropical dry forest (see site description details in Méndez-Toribio et al 2016). The forest grows on low hills with altitudes ranging from 300 to 800 m a.s.l. where lithosols are the most abundant soil type (Ferrusquía-Villafranca 1993, Nava et al 1998). The climate is dry warm and highly seasonal, with an annual average temperature of 27.1 °C (30.2 °C monthly maximum during the dry season) and an annual average rainfall of 755.4 mm, of which 80% falls in the period June to September. A marked dry season occurs from October to May (data from the Tziritzícuaro meteorological station, 1951–2010). The structure and diversity of the area are similar to those documented for seasonal tropical forests of Mexico and the Neotropics (Méndez-Toribio et al 2014).
Vegetation sampling plots were located on the slopes of three mountains. To classify slope aspect, an ArcGIS 9.3 (ESRI 2009) geographic information system was used. North-facing slopes (315° − 45°) had an average steepness value of 40% (range 15%−63%) while south-facing slopes (135° − 225°) was 45% (range 14%−47%). Four 100 m2 (10 × 10 m) plots, two facing north and two facing south, were established on three relative positions along the slope: the lower (380–500 m), middle (480−540 m) and upper (560–780 m) portions of each mountain slope, giving a total of 36 plots (0.36 ha; Méndez-Toribio et al 2016). Average steepness values at the lower, middle and upper portions of the slopes were 25% (14–47), 53% (40–68) and 48% (39–58), respectively. At each one of the 36 plots, rooted trees with a diameter at breast height (DBH) ≥ 2.5 cm were recorded. Plots were established in areas with no apparent evidence of disturbance, although the extraction of firewood and cattle breeding are common in the study area.
2.2. Characterization of environmental gradients
Environmental gradients generated by the interaction of the two topographic factors were characterized during two critical periods for plant growth and survival in the tropical dry forest regions: end of the dry season (March–April) and middle of the rainy season (June–July). To do so, we obtained values per plot of mean daily air temperature (°C), total incident radiation (MJ m−2) and potential evapotranspiration (mm) for the two mentioned periods. At each plot, temperature was registered hourly 2 m above the soil surface using one temperature sensor and data-logger (HoBo proV2, Onset, EUA). Incident solar radiation values per plot were derived by using ArcGis 9.3 ESRI (2009), based on a 20 m digital elevation model obtained for the studied landscape (see details in Méndez-Toribio et al 2016). Monthly potential evapotranspiration (a critical indicator of water demand, according to van der Maarel 2005) was then calculated based on air temperature and radiation, applying:
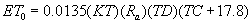
where TD = Tmax − Tmin (°C); TC = average air daily temperature (°C); KT is an empirical coefficient and Ra = solar radiation × 0.408 (Hargreaves 1994, Samani 2000). Finally, values of topographic wetness index (TWI; given a hydrological model, a steady state indicator of topographic induced soil water flux and content, Beven and Kirkby 1979), for each plot were taken from Méndez-Toribio et al (2016). Thus for each plot we obtained mean daily temperatures (T), integrated incident radiation (RAD) and potential evapotranspiration (ET0) for the two mentioned periods, and one single value of TWI, as an indicator of the ranking of each plot in terms of soil water content after rain events. In the studied landscape it is common to observe portions of the surface with rock exposed or covered by a thin layer of soil, particularly on the middle slopes and near the top hills. For this reason, we characterized variation in soil depth by measuring the maximum depth of mineral soil (MSD; dm) and % of coarse fragments on superficial horizon by excavating 1 m2 trenches, following Siebe et al (2006). Additionally, maximum main rooting depth was taken at each trench (MRD, Siebe et al 2006). Due to restrictions on permission to do excavations in the area, MSD, CF and MRD were measured only for nine plots; one low, one middle and one upper hill positions, located on the north-facing slope of the three studied hills.
2.3. Functional traits
A total of 63 species of trees with DBH ≥ 2.5 cm were recorded at the 36 sample plots (Méndez-Toribio et al 2016). For each species, 12 functional traits of the leaves and branches were measured (table 1). These traits are informative of the strategies of water use, gas exchange and resistance to drought, which are fundamental aspects for the persistence of trees in dry forests (Tyree et al 2003).
Table 1. List and description of the 12 functional traits measured in 63 species of trees in a tropical deciduous forest in south-central Mexico.
| Trait | Abbreviation (units) | Functional role |
|---|---|---|
| Leaf | ||
| Leaf areaa | LA (cm2) | Leaf cooling, light capture efficiency (self shading) |
| Leaf compoundnessa,b | LC (O) | Foliar cooling efficiency, light capture economics (structural investment) |
| Leaf dry matter contentc,d | LDMC (g g−1) | Drought tolerance, leaf longevity |
| Leaf mass per areae,f | LMA (g cm−2) | Light capture efficiency, maximum photosynthetic rate, leaf carbon economy |
| Leaf pubescencea,b | LPb (B) | Leaf cooling, water balance |
| Leaf pulvinationa,b | LPl (O) | Irradiance control |
| Leaf retention timeg | LRT (days) | Drought avoidance |
| Petiole lengthh | PL (mm) | Light capture efficiency |
| Branches | ||
| Bark thicknessi,j | BT (mm) | Desiccation resistance (to avoid water loss) |
| Bark water contentk | Bwc (%) | Delaying water-stress, resistance to desiccation |
| Wood densityl,m | WD (g cm−3) | Drought tolerance (xylem resistance to cavitation) |
| Xylem water contentk | Xwc (%) | Lagging water-stress, resistance to desiccation |
aSchulze et al (2002). bLambers et al (2008). cNiinemets (1999). dMarkesteijn et al (2011). eWestoby et al (2002). fWright et al (2004). gMéndez-Alonzo et al (2012). hTakenaka (1994). iScholz et al (2007). jPaine et al (2011). kRoger (2001). lHacke and Sperry (2001). mHolbrook and Zwieniecki (2005).
During the rainy season, five individuals with a DBH ≥ 2.5 cm were selected from each species, with five leaves collected from each individual. The selected leaves were exposed to solar radiation, with no apparent damage by herbivores, and located at an equivalent height among the individuals. The leaves were stored in darkness inside a cooler in an airtight humid environment until subsequent processing. Fresh leaves were weighed and scanned, and petiole length measured with a digital caliper to 0.01 mm. Leaf area was calculated from digital images by using the ImageJ program (Abràmoff et al 2004). Finally, leaves were oven-dried for 48 h at 60 °C and leaf mass per area was calculated by dividing leaf dry mass by leaf area, while leaf dry matter content was obtained from the ratio of dry leaf mass: fresh leaf mass. In the case of composite leaves, the leaflets were used to obtain these attributes. Leaf compoundness was assigned the value of 1 for simple leaves, 2 for compound leaves and 3 for double-compound leaves. Leaf pubescence was a binary variable to which, the values of 0 and 1 were assigned, respectively, in the absence and presence of trichomes on the upper or lower leaf surfaces. Leaves without pulvini were assigned a value of 0, while those with pulvini on the first order leaflets had a value of 1 and those with pulvini on the second order leaflets had a value of 2. To calculate the leaf retention time for each species, a leaf area loss curve was generated following the procedures described in Méndez-Alonzo et al (2012), and the time taken by plants to lose 50% of their leaves was calculated.
Stem attributes (wood density, wood water content, bark thickness and bark water content) were measured on second order branches (1.5 to 3 cm in diameter) fully exposed to sunlight, taken from five trees at a similar height (2 m). On each branch, bark thickness was measured with a digital caliper to 0.01 mm, while wood density was calculated by obtaining the volume of a branch section without bark using the water displacement method, and drying the tissue in a conventional oven at 90 °C for 72 h, following Chave et al (2006). Maximum water content in the xylem (Xwc) or bark (Bwc) was estimated as ([fresh weight − dry weight] / dry weight) × 100. The fresh weight of the branches was obtained after hydration for 24 h in distilled water.
2.2. Data analysis
Variation of physical variables (T, RAD, ET0), in relation to slope aspect and topographic position were examined by using two-way ANOVA, one for each season. In order to reduce any potential effect of spatial autocorrelation between the sampling units, the ANOVA was constructed using a re-sampling algorithm with 9999 permutations (Anderson and Legendre 1999). When one of the main factors (slope aspect or topographic position) was significant, the different factor levels were compared with a post-hoc test. When the slope aspect x slope position was significant, mean values among the six resulting groups were compared. Adjustment of P values were carried out following Holm (1979). Similar ANOVA was applied on values of TWI, while a one-way ANOVA was used for the case of MSD, % CF and MRD, to test only for topographic position on the N-facing slope.
The response of the functional dominance of vegetation to slope aspect and topographic position was evaluated for the 12 individual attributes and functional strategies, using multivariate and univariate approaches. The dominance of each functional attribute in each plot was obtained by calculating a community weighted mean (CWM), following the equation:
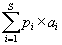
where S is the total number of species, pi is the relative abundance of species (i) in the community, and a is the attribute value of the ith species. This index indicates the average functional value of a given plant in the community, is very sensitive to environmental variation and has been correlated with certain ecosystem properties in various regions (Diaz et al 2004, Díaz et al 2007, Vandewalle et al 2010, Roscher et al 2012, Zhang et al 2014). CWM values of each attribute were calculated with the 'functcomp' function available in the 'FD' library of the R-CRAN (R Development Core Team 2016). The effects of the slope aspect and topographic position on the CWM of the individual attributes were assessed with a two-way analysis of variance. In order to reduce any potential effect of spatial autocorrelation between the sampling units, the ANOVA was constructed using a re-sampling algorithm, as the one previously described for the physical variables.
Exploration of co-variation among the attributes in the community, and determination of functional strategy continuums were both performed with principal component analysis (PCA), using the CWM values for each attribute in each plot. The effect of topographic factors on the dominance of functional strategies was assessed by analyzing the change in the average value of PCA scores along the main axes 1 and 2 using a two-way ANOVA. All data analyses were carried out with the open source program R 3.0.2 (R Development Core Team 2016); 'ade4' was used to perform the PCA with the function 'dudi.pca' (Dray and Dufour 2007), and the two-way analysis of variance with permutations was performed.
3. Results
3.1. Environmental gradients
Physical variables were affected by slope-aspect and topographic position but less clearly by their interaction (table 2). During the dry season south-facing slopes presented higher values of RAD, T and ET0 than N-facing slopes, and these results reversed during the rainy season (table 3). In addition, values of T and ET0 decreased from the lower positions to the top hills, independently of the aspect and season (tables 2 and 3). During both seasons, values of RAD were lower in the middle slope position, mostly on the north-face slope (tables 2 and 3). The topographic wetness index exhibited similar values between slope aspects, with values increasing from top hills to valleys, while % of coarse fragments and main root depth exhibited similar, but marginal trends along topographic position, together indicating increasing of soil depth towards the valleys (tables 2 and 3).
Table 2. Summary of two-way ANOVA with permutations to test the effect of slope aspect and topographic position on environmental variables in a tropical deciduous forest located at the Balsas river basin in south-central Mexico. Statistically significant values are shown in bold text.
| Variable | Aspect | Topographic position (Tp) | Aspect × Tp |
|---|---|---|---|
| F1,30 (P) | F2,30 (P) | F2,30 (P) | |
| Dry season end | |||
| T | 6.05 (0.019) | 7.86 (0.001) | 0.25 (0.793) |
| RAD | 83.88 (<0.001) | 10.07 (0.001) | 3.37 (0.052) |
| ET0 | 26.81 (<0.001) | 14.74 (<0.001) | 1.57 (0.225) |
| Rainy season | |||
| T | 0.57 (0.471) | 6.01 (0.002) | 1.17 (0.343) |
| RAD | 143.12 (<0.001) | 22.46 (<0.001) | 8.56 (0.002) |
| ET0 | 48.79 (<0.001) | 15.48 (<0.001) | 2.59 (0.099) |
| Soil variables | |||
| TWI | 0.64 (0.429) | 5.14 (0.008) | 0.98 (0.391) |
| MSD | − | 3.79 (0.081) | − |
| CF | − | 4.35 (0.085) | − |
| MRD | − | 0.57 (0.593) | − |
T, mean daily air temperature (°C); RAD, total incident radiation; ET0, potential evapotranspiration; TWI, topographic wetness index; MSD, maximum mineral soil depth; CF, coarse fragments (%/100) on superficial horizon (Ah) and MRD, main rooting depth (dm). MSD, CF and MRD were evaluated only for topographic position taking nine plots on the north-facing slope (see section 2).
Table 3. Environmental and soil (mean ± SE) variables as affected by topographic factors. Different letters indicate significant differences at P ≤ 0.05 among factor levels using two-way ANOVA with permutations and posteriori pair-wise comparisons.
| Variables | Slope Aspect | Topographic Position | ||||
|---|---|---|---|---|---|---|
| North | South | Lower | Middle | Upper | ||
| Dry season end | ||||||
| T | 28.0a(0.2) | 28.6b (0.2) | 28.8b (0.2) | 28.4b (0.2) | 27.7a (0.2) | |
| RAD | 506.6a(0.3) | 576.1b (0.2) | 555.1b (9.6) | 517.4a (14.6) | 551.6b (13) | |
| ET0 | 84.8a(0.5) | 96.4b (0.5) | 99.2b (2.5) | 85.3a (2.6) | 87.4a (2.7) | |
| Rainy season | ||||||
| T | 27.2a(0.4) | 26.9a (0.2) | 27.6b (0.2) | 27.4b (0.4) | 26.1a (0.3) | |
| RAD | 643.4b(4.4) | 572.6a (8.7) | 625.6b (7.2) | 580.3a (15.4) | 618a,b (12.9) | |
| ET0 | 87.0b(1.2) | 76.3a (1.8) | 87.5b (1.6) | 77.4a (2.4) | 80.1a (2.2) | |
| Soil variables | ||||||
| TWI | 5.7a(0.7) | 5.1a (0.4) | 6.9b (0.7) | 5.1ab (0.4) | 4.1a (0.8) | |
| MSD | – | – | 7.8 (1.7) | 6.6 (0.53) | 5.1 (1.2) | |
| CF | – | – | 0.09 (0.04) | 0.45 (0.22) | 0.48 (0.47) | |
| MRD | – | – | 6.4 (1.6) | 5.3 (2.0) | 5.1 (1.2) | |
T mean daily air temperature (°C); RAD, total incident radiation (MJ m−2); ET0, potential evapotranspiration (mm); TWI, topographic wetness index; MSD, maximum mineral soil depth (dm); CF, % of coarse fragments on superficial horizon, and MRD, main rooting depth (dm). Mean and standard errors were calculated from 36 sites, except for MSD, CF and MRD which included only nine sites on the north-facing slope (see section 2). TWI values were taken with permission from Méndez-Toribio et al (2016).
3.2. Functional dominance
The statistical tests applied to the CWM values of the individual functional traits, indicated that the response was more strongly affected by topographic position (eight attributes) than by slope aspect (two attributes) or the interaction of these two factors (two attributes; table 4). Bark water content, bark thickness, petiole length and leaf pubescence increased significantly towards the upper parts of the slopes, while wood density, leaf pulvini, leaf dry matter content and leaf retention time were higher on the lower parts of the slopes (figure 1).
Table 4. Summary of two-way ANOVA with permutations to test the effect of slope aspect and topographic position on CWM of individual functional traits in a tropical deciduous forest located at the Balsas river basin in south-central Mexico. Statistically significant values are shown in bold text.
| Traits | Aspect F1,30 (P) | Topographic position (Tp) F2,30 (P) | Aspect × Tp F2,30 (P) |
|---|---|---|---|
| Leaf | |||
| Leaf area | 1.02 (0.37) | 1.02 (0.41) | 0.32 (0.81) |
| Leaf compoundness | 1.78 (0.20) | 1.35 (0.28) | 2.12 (0.14) |
| Leaf dry matter content | 3.51 (0.07) | 10.84 (< 0.001) | 2.39 (0.11) |
| Leaf mass per area | 0.00 (0.98) | 0.05 (0.95) | 0.36 (0.70) |
| Leaf pubescence | 2.67 (0.09) | 8.17 (0.002) | 1.17 (0.32) |
| Leaf pulvination | 2.86 (0.09) | 2.72 (0.05) | 2.11 (0.13) |
| Leaf retention time | 7.83 (0.009) | 9.78 (< 0.001) | 3.96 (0.03) |
| Petiole length | 0.49 (0.57) | 3.11 (0.03) | 0.89 (0.47) |
| Branches | |||
| Bark thickness | 1.84 (0.19) | 7.40 (0.003) | 3.82 (0.03) |
| Bark water content | 1.07 (0.31) | 9.50 (0.001) | 0.20 (0.82) |
| Wood density | 17.19 (< 0.001) | 18.56 (< 0.001) | 0.33 (0.74) |
| Xylem water content | 0.17 (0.69) | 1.95 (0.16) | 2.39 (0.11) |
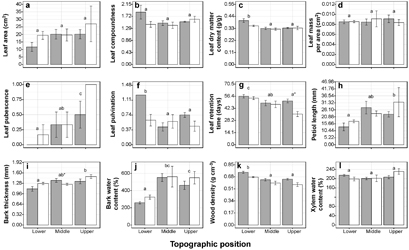
Figure 1 Average values (± 1 standard error) of 12 functional traits of trees evaluated at community level (CWM) in three topographic positions facing north (gray bars) and south (white bars) in a tropical deciduous forest, located at the Balsas river basin in south-central Mexico. Different letters above the bars indicate significant differences between topographic positions, while asterisks indicate differences between slope aspects at each topographic position (P ≤ 0.05). See table 4 for effects of slope aspect.
Download figure:
Standard image High-resolution imageCWM for wood density and leaf retention time were significantly higher on north-facing slopes compared with those south-facing slopes (table 4, figure 1). Bark thickness and leaf retention time were the only attributes affected by the interaction of both topographic factors (table 4). In particular, thick barks were more dominant in the south than in the north-facing aspect at both the lower and upper parts of the slopes, while in the middle portions the opposite trend was presented (figure 1). Similarly, the dominance of readily-deciduous species (species with low leaf retention time) was higher on the southern aspect, although only on the upper parts of the slopes, while aspect did not affect dominance in the other two positions (figure 1).
The PCA showed that axes 1 and 2 explained 45.7% and 15.4% of trait co-variation, respectively (figure 2; table 5). The first axis of the PCA was negatively correlated with bark thickness, petiole length, bark water content, leaf area and leaf pubescence, and positively with wood density, leaf retention time, leaf dry matter content and leaf pulvini, partly suggesting a trade-off between two drought-coping strategies: (i) avoidance via the use of stored water with a rapid reduction in leaf area, and (ii) tolerance, via dense tissues and small leaves. Attributes negatively associated with axis 2 were leaf compoundness and leaf pubescence (two traits related to mechanisms of overheating avoidance) and xylem water content. This latter attribute showed relatively low variation among species and was independent of those traits related to tissue density (wood density and leaf dry matter content) and bark water content. In addition, leaf retention time was related to both axes, i.e. it was positively associated with dense tissues and negatively related to xylem water content and leaf compoundness. Leaf mass per area was the only attribute not correlated with either of the two axes.
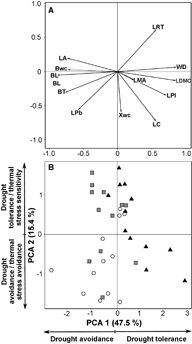
Figure 2 Principal component analysis (PCA) of the CWM of 12 functional traits of trees (stems and leaves) evaluated in a tropical dry forest in south-central Mexico. (a) Eigenvector plot, and (b) species score plots along the PC1 and PC2 axes. Sites on lower slope position (triangles) middle slope (squares) and upper slope. Functional trait names are given in table 1.
Download figure:
Standard image High-resolution imageTable 5. Pearson correlations between PCA scores and community weighted means of traits.
| Traits | Axis-1 | Axis-2 |
|---|---|---|
| Leaf | ||
| Leaf area | −0.73⁎⁎⁎ | 0.20n.s. |
| Leaf compoundness | 0.56⁎⁎⁎ | −0.72⁎⁎⁎ |
| Leaf dry matter content | 0.90⁎⁎⁎ | −0.13n.s. |
| Leaf mass per area | 0.23n.s. | −0.12n.s. |
| Leaf pubescence | −0.57⁎⁎⁎ | −0.56⁎⁎⁎ |
| Leaf pulvination | 0.71⁎⁎⁎ | −0.34⁎ |
| Leaf retention time | 0.56⁎⁎⁎ | 0.61⁎⁎⁎ |
| Petiole length | −0.88⁎⁎⁎ | −0.05n.s. |
| Branches | ||
| Bark thickness | −0.76⁎⁎⁎ | −0.30n.s. |
| Bark water content | −0.73⁎⁎⁎ | 0.03n.s. |
| Wood density | 0.86⁎⁎⁎ | 0.07n.s. |
| Xylem water content | 0.05n.s. | −0.59⁎⁎⁎ |
⁎P < 0.05. ⁎⁎P < 0.01 y. ⁎⁎⁎P < 0.001. n.s.= not significant.
The ANOVA applied to the scores of axis 1 of the PCA indicated that these values decreased from the lower to the upper parts of the slopes (figure 3(a); F2,30 = 12.73, P ≤ 0.001) and were significantly higher on north-facing slopes than on those south-facing (F1,30 = 6.27, P ≤ 0.01). The average values of axis 2 were more negative towards the upper parts of the slopes (figure 3(b); F2,30 = 5.62, P ≤ 0.01), particularly on the southern slopes (F2,30 = 4.65, P ≤ 0.05, interaction term).

{kind=link}
{kind=link}
Figure 3 Dominance of functional strategies of trees in relation to topographic position and slope aspect (north-facing: grey bars, south-facing: white bars). Values of Y-axis correspond to average values (± 1 standard error) of the PCA scores of sites on axis 1 (a) and axis 2 (b). Different letters above the bars indicate significant differences among topographic positions, while asterisks indicate significant difference between slope aspects at each topographic position (P ≤ 0.05). See the main text for effects of the slope aspect.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
4.1. Topographic position and slope-aspect generate gradients of drought risk, both above and below ground
In our study site, slope-aspect strongly affected physical variables (RAD, T and ET0), but such effects varied in opposing directions between the wet and the dry season. During the rainy season, growth may be favored on the north-facing slope, as this receives higher radiation, while during the dry season, a higher radiation load, higher temperature and potential evapotranspiration may compromise plant growth and survival (see Méndez-Toribio et al 2016, for an analysis integrating physical variables across the year in the same study site). The finding of similar results for another tropical dry forest landscape in Mexico (Gallardo-Cruz et al 2009), indicates that this pattern of topographic variation in physical environment is not unique to our study site, raising the hypothesis that in tropical dry forest regions slope-aspect does not impose a clear filter on plant water use strategies.
It has been proposed that in undulating landscapes, superficial water movements impulsed by gravity from the top hill to the lower slope positions, typically with reduction in slope steepness, is commonly accompanied by an increase in both soil depth and soil water content, generating a gradient of soil drought risk for plants (Holland and Steyn 1975, Daws et al 2002, Tesfa et al 2009, Liu et al 2012, for empirical and modelling evidence, Tromp-van Meerveld and McDonnell 2006, for discussions on oversimplification issues). In our study landscape, the detected increase in topographic wetness index, a trend for increasing soil depth and reducing rock fragment content, together comply with the view, that the risk of soil drought increases from the lower slope positions to the top hills. Remarkably, observations from local farmers point to the same view5, however our conclusions need to be corroborated by a more extensive characterization of soil depth across the landscape, because other factors as micro-topography may play a role.
Intriguingly RAD observed slightly lower values in the mid slope position, but those changes did not reflect into parallel changes in temperature or evapotranspiration. Interestingly, the detected increase of air temperature and potential evapotranspiration, from the top hill to the lower slope position suggest that surface air gets warmer from the top hills to the lower slope positions, potentially increasing risk of drought above ground (particularly during the rainy season, when values became more affected by topographic positions). We are unable to size at what extent surface conditions counteract the soil drought gradient, however a stronger change of wetness index across topographic positions (40%), compared to a slight change of RAD, T or ET0, (1%, 5%, 8%, respectively), suggests that lower positions are warmer, but subject to lower risk of drought, at least during the growing season while rain water is flowing across the terrain surface. Soil movement downhill may also result in changes of soil nutrient content towards the valleys, affecting plant traits (Silver et al 1994). In this study we miss this factor as no data on soil nutrients were recorded.
4.2. Topographic position is a main driver of functional composition
This study demonstrates that topographic position is an overriding factor structuring functional dominance in tropical dry forests. The lack of response of the functional traits to slope aspect suggests that the difference in total annual radiation between north- and south- facing slopes is a less important filtering factor, probably due to the fact that at inter-tropical latitudes, the difference in annual radiation load between slopes is relatively small compared to that of higher latitudes (Holland and Steyn 1975, Gallardo-Cruz et al 2009, Méndez-Toribio et al 2016). The fact that slope aspect in these landscapes does structure species composition of the community (Méndez-Toribio et al 2016), but not its functional composition, suggests a convergence towards similar functional traits among different plant lineages. An answer to this interesting question would require a phylogenetic analysis to be conducted in the studied forest.
4.3. Evasive species dominate the upper parts of slopes, while drought-tolerant species dominate the lower parts
This study proposed the hypothesis that, in tropical dry forest landscapes, water-scarce environments (e.g. the upper parts of the slopes, particularly those facing south) tend to be covered by vegetation communities with attributes such as hard wood and dense tissues, that allow plants a greater tolerance to desiccation, compared to those of less dry areas (e.g. lower parts of the slopes, particularly those facing north). However, our results indicated the opposite trend, supporting the notion that opportunistic species, which quickly evade drought by shedding their leaves or by using water stored in their tissues, have an advantage over other plants on the upper parts of slopes, especially those facing south, where the growth period is shorter, more variable and exposed to high levels of insolation (Pineda-García et al 2011).
Our study suggests that the marked dominance of avoidance strategies on the upper portions of the slopes may be the result of filtering processes related to a soil depth gradient, from shallow soils on the high parts, to deep soils on the lower sections of the slopes. This affects the water dynamics, both spatially and temporally. Shallow soils on the high portions of slopes have a low water retention capacity and dry quickly after rain events. This translates into a high risk of death for plants over prolonged periods of drought during both the growing season and the beginning of the dry season (Tromp-van Meerdel and McDonnell 2006, Pineda-García et al 2011, Méndez-Alonzo et al 2013). Under this scenario, it is likely that the rapid loss of foliage during drought may drastically reduce the risk of cavitation, especially in species with wide vessels, typically associated with low-density wood (see Hacke and Sperry 2001 for wood density-vessel anatomy relationships; Méndez-Alonzo et al 2012, for TDF species). Moreover, it is likely that trees with low-density wood and large water deposits in the parenchyma and/or bark are able to use this stored water to rehydrate the xylem and temporarily dissociate the plant water balance from the soil water potential, as previous studies suggest (Goldstein et al 1989, Daws et al 2002, Scholz et al 2007, Meinzer et al 2008, Pineda-García et al 2012, Rosell et al 2014). Other studies have advanced the idea that species with water stored in their stems (xylem or bark) tend to be highly efficient in terms of water conduction and photosynthetic activity, and may be able to respond to water pulses by rapidly developing leaves and roots (Sobrado 1993, Holbrook et al 1995, Holbrook and Zwieniecki 2005, Pineda-García et al 2015). Such attributes contribute to maximize total carbon assimilation in environments with short and unpredictable growth periods (Eamus 1999, Santiago et al 2004, Choat et al 2005, Franco et al 2005, Pineda-García et al 2011). Taken together, the results of our study combine to support the notion that water-stress delaying strategies and the capacity to exploit water pulses may both represent an adaptive advantage in temporarily heterogeneous environments with long episodes of drought and short growth periods.
The dominance of readily-deciduous and water-storing species on the upper portions of the slopes, as well as the dominance of species with dense tissue tolerant to water stress on the lower portions of slopes, may also be reinforced by selective pressures of soil depth on rooting depth, and the occurrence of a tight coordination of rooting habit with stem and leaf traits. It is therefore expected that shallow soils with high runoff and poor vertical percolation prevent the development of deep roots, favoring species with water-storing strategies, while deeper soils, with greater influx and percolation of water, favor species with deep roots that are capable of capturing water from deeper and longer-lasting deposits. This is a particularly interesting possibility, given the fact that a previous study done with young plants in a dry tropical forest (Paz et al 2015), reported evidence of a potential trade-off between the two water-stress delay mechanisms; storing water in the tissues or rooting more deeply. This hypothesis seems plausible and its further exploration calls for a detailed study of above and below ground traits along topographic gradients, coupled with extensive measurements of soil depth. The expected increase of distance to water table from the valleys to the top-hills may exaggerate the detected drought risk for plants and selection on rooting depth, however this important factor still awaits to be measured, as water table depth can vary with local specific stratigraphy (Moore and Thompson 1996).
4.4. Heat and water stress-coping mechanisms in the leaves appear to vary with slope position
In the dry forest, tree foliage is subject to strong thermal stress, especially during periods of low soil water availability. It is therefore common to find leaf structures, such as the pulvini and trichomes, that help decrease the heating load and the risk of overheating or water loss. Species with leaf pulvination optimize the functioning of photosynthetic structures in extremely hot environments by changing the position of leaves or leaflets, this in turn affects transpiration and thus the water relations of the plant (Comstock and Mahall 1985, Schulze et al 2002). In our study, the dominance of species with leaf pulvination on the lower portions of slopes suggests a filtering process through this mechanism, as was in these landscape units where higher air temperatures were recorded (Méndez-Toribio et al 2016). The fact that this attribute is strongly linked to certain lineages of the Fabaceae family is remarkable. This family dominates the lower parts of the slopes of the dry tropical forests under study (Méndez-Toribio et al 2016), which suggests the existence of a significant phylogenetic component in the functional structure of the forest (Pennington et al 2009). We are not aware that other attributes analyzed here have a phylogenetic component as extreme as leaf pulvination, although both the functional structure and phylogenetic relationships among species remain to be clarified.
It has been proposed that trichomes act as mechanical filters, reducing the incidence of ultraviolet radiation and potential damage to the leaf tissue (Levizou et al 2004) and that they attenuate the drying effect of wind due to the increased thickness of the boundary layer (Meinzer et al 1985, Brewer and Smith 1994), reducing water loss through transpiration (Brewer and Smith 1994) as well as increasing water retention via a decoupling from stressful environmental conditions (Konrad et al 2015). In this context, the dominance of trichomes towards the upper parts of slopes could result from the exposure of the leaves to environments with drier soils but relatively cooler temperatures, where higher exposure to wind and ultraviolet radiation are to be expected. Previous studies in semi-arid areas of North America also have identified dominance of plant species with trichomes, suggesting that this phenomenon is not unique to our study area (Sandquist and Ehleringer 1997).
4.5. Leaf mass per area: irrelevant as an indicator of drought tolerance
It is interesting to note that leaf mass per area, which is important in the context of carbon foliar economy (Wright et al 2005), proved irrelevant to the characterization of functional strategies, since it showed no response to the topographical factors analyzed, nor was it included on the main axes of morpho-functional variation detected for this forest. This would confirm the proposition that, while this attribute is strongly related to the competitive ability of species in communities with marked vertical light gradients (Westoby et al 2002), it does not seem to be an important component in the functional strategies of trees in dry tropical forests, where heterogeneity in water availability strongly selects for a diversity of water use strategies among species (Pineda-García et al 2011).
4.6. Water reserves in the bark vary along the topographic gradient
Another attribute that was expected to vary significantly along the water topographic gradient is xylem water content, a key attribute related to the delay of water stress (Roger 2001). However, this attribute did not respond individually to any of the topographic features analyzed. This may be due to our failure to capture the most important water storage sections during branch sampling, which in certain species are found in the main stem (Gartner 1995, León and Espinoza 1999). In contrast, the strong co-variation between the topographic gradient and the bark water content and thickness suggests that the storing of water in thick barks may play an important role in maintaining water xylem water status during drought conditions (Gartner 1995). This could be equally or more important than the role of xylem water storage in community processes of functional differentiation along a topographic gradient, an aspect that deserves to be investigated in more detail (Rosell et al 2014).
5. Conclusions
It has repeatedly been asserted that topography is a core geographic feature of landscape, acting as a selective primary force that determines the high diversity of tree species in the tropics (Wright 2002, Valencia et al 2004). The present study shows evidence of filtration processes of plant functional strategies to deal with drought and heat stress, operating in the dry forest mostly along topographic positions but not much along the gradient generated by slope aspect. The incorporation of a functional dimension to the study of the effects of topography may help predict spatial variation of key processes and services provided by ecosystems in a given landscape.
Acknowledgments
The first author (M M T) is grateful to the Posgrado en Ciencias Biológicas program of UNAM and the Consejo Nacional de Ciencia y Tecnología for the facilities and a scholarship awarded to conduct doctoral studies. Additional funding was provided by PAPIIT- UNAM, grant IN207512. We are indebted to the people of Tziritzícuaro community for assistance in the field work, and to Atzimba López Maldonado, Alberto Valencia García, Heberto Ferreira Medina and Guadalupe Cornejo Tennorio for technical and field assistance support. We also thank Biol. Melissa Zermeño Hernández and Biol. Stephany Manzo Melchor for their support during biological material processing.
Footnotes
- 5
Personal communication.