
Abstract
In Thymus vulgaris L., sex determination involves both the nuclear and the cytoplasmic genomes: the cytoplasm is responsible for male sterility (the female phenotype) whereas specific nuclear genes may restore male fertility (the hermaphrodite phenotype). The evolutionary dynamics of cytoplasmic male-sterility genes and nuclear restorer genes represents a coevolutionary conflict. Here we draw a parallel between this conflict and the coevolutionary interaction between hosts and parasites. Local adaptation of parasites to their hosts may be predicted under some situations of relative evolutionary rates of the two partners in this conflict and is often observed in natural populations. We tested for local adaptation between nuclear restorer genes and cytoplasmic male-sterility factors using 12 female plants that were hand-pollinated with pollen from hermaphrodites from more or less distant populations, and measuring progeny sex ratios. No significant effects of the geographical distance between maternal and paternal parents could be detected, although highly significant differences among individual females for the restorability of their cytoplasms were found. We suggest that the same evolutionary processes giving rise to individual differences among cytoplasmic male-sterility types might account for the absence of local adaptation between nuclear and cytoplasmic factors in this system, and also discuss the absence of local adaptation in the light of a recent model of local adaptation in host-parasite metapopulations.
Similar content being viewed by others

Introduction
Gynodioecy, the presence of hermaphrodite and male-sterile individuals in the same population, has been the focus of many empirical and theoretical studies both because it is considered a transitory stage from hermaphroditism to dioecy (Bawa, 1980; Schultz, 1994) and because both the nuclear and the mitochondrial genomes are involved in the genetic determination of gynodioecy in many species. Indeed, the existence of cytoplasmic male sterility had been already postulated in 140 species representing 47 genera and 20 families in 1972 (Kheyr-Pour, 1980). More recently, the genetic basis of sex determination has been shown to involve nuclear and cytoplasmic factors in gynodioecious crop species (Saumitou-Laprade et al., 1994) as well as in gynodioecious wild species such as Plantago lanceolata L. (van Damme, 1983), Origanum vulgare L. (Khey-Pour, 1980) and Thymus vulgaris L. (Couvet et al., 1986).
The evolutionary dynamic of cytoplasmic male-sterility factors with their nuclear restorer genes represents a coevolutionary conflict (Couvet et al., 1990; Gouyon et al., 1991) as is the relationship between hosts and parasites, although the genetic basis of the interaction may be different. In host-parasite interactions virulence factors in parasites select for resistance by their hosts (Frank, 1991), even when virulence and resistance do not follow a simple gene-for-gene relationship, and patterns of local adaptation of parasites to their sympatric hosts are expected (Ebert, 1994) and often observed (Parker, 1985; Lively & Jokela, 1996; but see Burdon & Jarosz, 1992). Recent theoretical developments that consider host-parasite interactions in a metapopulation, where systems of populations are connected by colonization and migration, suggest that locally adapted parasites should only occur under limited conditions. Where parasite populations exchange more genes than do those of their hosts, parasites should be locally adapted. When hosts have higher migration rates than parasites, on the other hand, parasites should be less capable of infecting sympatric than allopatric hosts, i.e. they are locally maladapted (Gandon et al., 1996). When new cytoplasmic factors for male sterility appear by mutation or migration in gynodioecious plant populations this induces selection for specific nuclear genes that restore male fertility (van Damme, 1982). This pattern of selection has been suggested to generate local adaptation of nuclear restorer genes to their particular cytoplasmic factors. Higher rates of restoration (a higher proportion of hermaphrodites) are therefore predicted for the progeny of crosses between two individuals from the same population compared with those between different populations (Couvet et al., 1986). This is consistent with the theoretical expectations outlined by Gandon et al. (1996), because nuclear genes, which are carried both in pollen and seeds, migrate more than cytoplasmic genes, which are present only in seeds if the inheritance is maternal as is generally the case (Corriveau & Coleman, 1988; Reboud & Zeyl, 1994). To our knowledge the existence of local adaptation of nuclear restorer genes to cytoplasmic factors has not yet been experimentally tested.
Here we test for this phenomenon using crosses of the gynodioecious species Thymus vulgaris L., Labiatae. We compare within-population crosses with those between populations and between regions for their rates of restoration and test for within-population variation in male-sterility cytoplasms in their restorability.
Materials and methods
Plant materials
Thymus vulgaris (thyme) is a small perennial entomophilous shrub characteristic of dry open Mediterranean vegetation. It is a pioneer species found in abandoned fields (Dommée et al., 1978) and burnt habitats (Belhassen et al., 1989) but remains in later stages of succession. In thyme the frequency of females varies between populations from 5 to 95% and decreases with population age (Assouad et al., 1978; Dommée et al., 1983; Belhassen, 1987). Plants are mainly pollinated by bees, which carry pollen over short distances (Dommée et al., 1983), but butterflies have been reported also to participate in pollination over longer distances (Tarayre & Thompson, 1997). Flowering occurs every year and lasts for two to four weeks. Each plant produces hundreds of small flowers with an ovary enclosing four ovules. The hermaphrodite flowers, which are larger than female flowers, also bear four stamens. These flowers are protandrous, the pistil becoming receptive a few days after the stamens' dehiscence. Sex in thyme is determined by the interaction of cytoplasmic male-sterility genes and nuclear restorer genes (Belhassen et al., 1991). The restoration of male fertility of a given cytoplasmic male sterility requires the presence of its specific restorer gene(s).
Geographical origin of parental plants
Seeds were collected from three natural populations of the region of Languedoc (Vialaret: 3°40'E, 43°49'N; Pompignan Haut: 3°52'E, 43°54'N; La Jasse: 3°46'E, 43°47'N) and two populations of Provence (Rustrel: 5°29'E, 43°55'N; Le Barroux: 5°05'E, 44°07'N) in the south of France. The three populations from the Languedoc are 10–50 km apart. The two Provence populations are situated more than 300 km from the Languedoc populations. The frequencies of hermaphrodites in these populations at the time the seeds were collected were 0.90, 0.50 and 0.55 for Pompignan Haut, La Jasse and Vialaret and 0.77 and 0.58 for Rustrel and Le Barroux, respectively, whereas the average frequency of hermaphrodites within the species is about 0.4. The frequency of hermaphrodites was measured by recording the sex of all individuals in each population. All populations contained between 100 and 200 plants. We chose populations with higher than average frequencies of hermaphrodites because hermaphrodite frequency is known to increase with increasing population age (Manicacci et al., 1996), and coevolutionary processes that could lead to local adaptation should be evident in old populations where male-sterility cytoplasms have coevolved with their nuclear restorer genes.
Seeds were sown in the greenhouse and seedlings were transplanted into an experimental field. When these plants flowered, 12 females and 50 hermaphrodites were chosen for the experiment as follows. Four females originating from each of the three Languedoc populations (12 altogether), each from a different mother, were chosen at random and transplanted to an insect-free greenhouse. Fifty hermaphrodites (10 from each of the five populations) were chosen at random to be used as pollen donors the following spring.
Crossing protocol
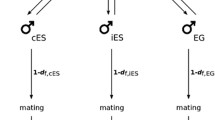
Three types of crosses were performed on each of the 12 females (for exact protocol see Fig. 1): (i) within-population crosses with pollen from the same population as the female; (ii) between-population crosses with pollen from a population different from the female but of the same region (Languedoc); (iii) between-region crosses with pollen from a population located in a different region (Provence).

Design of the experiment specifying the paternal populations used for each cross type.
Each female individual was divided into three equivalent parts using screens. The flowers of each part received pollen from the 10 hermaphrodites from each chosen paternal population, to simulate the pollen cloud of this population. To avoid pollen competition, each female flower was pollinated with pollen from a single hermaphroditic flower. Approximately 200 flowers were pollinated per female per treatment.
Seed families
Seeds produced from each of the 36 (12 females ×three treatments) combinations were collected two months later, germinated and cultivated in pots in the greenhouse. Individuals originating from each combination shared a common mother and 10 fathers coming from a single population; they will be referred to as a family. We obtained 20–50 offspring per family. One year later, at flowering, the frequency of hermaphrodites and females was scored in each family.
Statistical analysis
All statistical analyses were conducted using the GLIM statistical package (Baker, 1987). We performed two independent analyses. The first tested the effects of cross type (within population, between populations within region, and between regions), maternal population, maternal individual and their interactions on the numbers of hermaphrodites and females within each progeny using a logistic regression. The second analysis investigated the effect of paternal population, testing for specific interactions between maternal individual, maternal population and paternal population, without considering the distance between populations. In both analyses maternal individual was nested within maternal population. Cross type effect was declared as a fixed factor. It was possible to calculate the interactions between maternal plant and cross type or paternal population to determine whether different mothers reacted differently to the three types of cross and to the five paternal populations, respectively.
The total model was somewhat underdispersed, as demonstrated by the error mean deviance of less than one. Because testing against this error term would artificially inflate the F-value, for those effects that would be tested against the error mean deviance we compared the deviance directly against the χ2 expectation with appropriate degrees of freedom. These effects were Mother (Maternal population) and the interactions between this factor and Type of cross in the first analysis and Paternal population in the second analysis.
In addition, to examine the relationship between sex ratios produced by each mother in the different cross types we calculated the correlation between sex ratio produced by the within-population cross and sex ratio produced by the other crosses. This tests whether the restoration by one cross type is a good predictor of restoration by other cross types.
Results
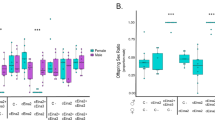
The proportion of hermaphrodites (the sex ratio) observed in the 36 families was highly variable (Table 1), ranging from 0 to 1. Although, on average, the proportion of hermaphrodites was larger in within-population crosses than in between-population or between-region crosses (0.36±0.08, 0.26±0.09 and 0.24±0.09, respectively; mean ±SE; see Table 1), the logistic regressions showed that variation in numbers of hermaphrodites and females within families was explained by differences between mothers within maternal populations and the interaction between mothers within maternal populations and both the type of cross in the first analysis and the particular paternal population in the second (Table 2). However, no main effects of cross type (the distance between maternal and paternal populations), paternal nor maternal population of origin were found.
Progeny sex ratios generated in the within-population crosses were very good predictors of progeny sex ratios from both other cross types (Fig. 2). This demonstrates that some maternal parents were well restored in all crosses whereas some were poorly restored overall. Between-population crosses restored better than between-region crosses for some maternal plants although for others the opposite was true, as demonstrated by the significant interactions in Table 2. In addition, most data points lay below the diagonal for equal restoration for all three cross types, showing that most within-population crosses restored somewhat better than between-population or between-region crosses (Fig. 2).

Sex ratios from between-population and between-region crosses of Thymus vulgaris plotted against the sex ratio from the within-population cross for each mother. The diagonal line represents the line of equal restoration for the three cross types. Pearson product-moment correlations were significant both for between-population and between-region crosses (N=12, r=0.81, P=0.0014 and N=12, r=0.74, P=0.0056, respectively).
Discussion
In this study, using females of T. vulgaris crossed with pollen from sympatric and more or less distant allopatric populations, we found no evidence for local adaptation but strong differences between cytoplasms in their restoration ability. The existence of different cytoplasmic male-sterility types within a single population has been reported in several gynodioecious species: Plantago lanceolata (van Damme, 1982), Plantago coronopus (Koelewijn & van Damme, 1995) and Beta vulgaris ssp. maritima (Boutin et al., 1987; Saumitou-Laprade et al., 1993). In thyme, the existence of at least four cytoplasmic male-sterility types within populations has been demonstrated (Manicacci et al., 1997), but this probably underestimates population variation. Even more polymorphism is suggested by the distribution of sexual phenotypes within natural populations, and by the high level of molecular polymorphism observed for cytotypes (Belhassen et al., 1993; Manicacci et al., 1996). Although it is difficult to establish a clear relationship between molecular and functional diversity, several studies (Belhassen et al., 1993; Cuguen et al., 1994; Ronfort et al., 1995) have shown that they are linked.
In our thyme populations some mothers carried a cytoplasmic male-sterility type that was readily restored whereas others carried a poorly restored cytoplasmic male-sterility type (Table 1). Indeed, some mothers (mothers 10 and 2, for example) produced a high rate of hermaphrodites regardless of the paternal population they were mated to, but some others (mothers 8 and 3, for example) always produced a low sex ratio (Table 1). This was observed in all maternal populations and no significant differences were obtained. In addition to this difference in general restoration ability, mothers also differed in the nature of their response to the different cross types. Some mothers were equally well or poorly restored by all cross types whereas others varied greatly (Table 1). For the latter, restoration was generally better by sympatric pollen (Fig. 2), but because few females showed this pattern, there was no significant overall effect of local adaptation. Comparable results have been obtained for a larger selection of populations from the Languedoc Region (Manicacci et al., 1997).
This experiment therefore did not support the hypothesis of local adaptation between nuclear and cytoplasmic sex determination factors in T. vulgaris (Couvet et al., 1986). We found no evidence that nuclear restorer genes were better at restoring their sympatric cytoplasms than they were at restoring allopatric cytoplasms. Nor was there an effect of distance between the populations of origin for maternal or paternal parents. Unfortunately our statistical power was limited for this analysis, with only four maternal plants for each of three populations. Indeed, we would have been able to find local adaptation only if at least 10 of the 12 females had been better restored by sympatric than allopatric pollen, using a conservative sign test. Possibly therefore our inability to find local adaptation might have resulted from low detection ability. Alternatively, we might have sampled at an inappropriate geographical scale, and local effects may have been found if we had chosen a finer one.
If local adaptation is truly lacking in this system, this could result from several causes. First, as stated above, the fact that different females responded differently to the three cross types suggests that there is no general pattern of local adaptation. These differences between mothers could result from the fact that some cytoplasmic types, on account of the biochemical basis of male sterility, are more easily restored than others, as has been shown for Plantago lanceolata (van Damme, 1983). Such male-sterility cytoplasms may be restored by many different unspecific restorer genes and therefore by a diversity of cross types or paternal populations. Another possibility is that if there is little cost of restorer genes, the restorer genes for cytoplasmic types that have existed for a long time can be expected to spread even to populations where the specific cytoplasm is absent. This process generates a loss of local adaptation with increasing age of a cytoplasmic type. Sampling a range of ages of cytoplasmic male-sterility types would therefore generate the large differences among mothers for their response to cross type and thus their degree of local adaptation.
A different possibility to explain the lack of local adaptation between nuclear restorer and cytoplasmic male-sterility factors is that the same cytoplasmic types exist over a broad geographical range, as has been shown for Beta maritima (Boutin et al., 1987; Saumitou-Laprade et al., 1993). In this case appropriate restorer genes would also be selected throughout, leading to an absence of geographical pattern when pollinations are performed with pollen mixtures. This situation might result from repeated mutation to the same male-sterility factor, or from a high rate of migration of cytoplasms, through high seed flow. In fact, in coevolutionary conflicts local adaptation is only predicted when migration rates or, similarly, mutation rates have particular restricted values per generation for both partners (Gandon et al., 1996). Even if migration rates are appropriate for the evolution of local adaptation, observations from a particular moment during the coevolutionary cycle can show lag phases where the more rapidly evolving player trails the slower one in a transitory manner (Nee, 1989). However, the parallel between host-parasite and cytoplasmic male-sterility restorer genes warrants further examination, because cytoplasmic male sterility and restorer genes modify the fitness of their bearers in a different way than do virulence and resistance genes.
Conclusion
Here we present a number of hypotheses that might explain the lack of local adaptation between male-sterility cytoplasms and their nuclear restorer genes. These hypotheses are not mutually exclusive, and a combination of these effects could generate the patterns observed. High variation for neutral markers associated with male-sterility cytoplasms has been found in thyme (Manicacci et al., 1996), suggesting that widespread cytoplasms are uncommon. Therefore, it is unlikely that local adaptation is absent because the same cytoplasm is found over a large geographical area. Clearly the strong maternal effect together with the highly significant interactions of maternal effect with both distance between maternal and paternal populations and paternal population itself indicate that cytoplasms differ in how easily they are restored by different nuclear restorer genes. At the same time, low cost of restoration could allow restorer alleles to accumulate. Estimates of the real costs of restoration have proved difficult to obtain, so low or even absent costs can reasonably be supposed (de Haan et al., 1997). Therefore, we consider that a combination of the last two factors would be the most likely explanation for a lack of local adaptation.
References
Assouad, M. W., Dommée, B., Lumaret, R. and Valdeyron, G. (1978). Reproductive capacities in the sexual forms of the gynodioecious species Thymus vulgaris L. Bot J Linn Soc, 77: 29–39.
Baker, R. J. (1987). GLIM 3.77. Reference manual, 2nd edn. Numerical Algorithms Group, Oxford.
Bawa, K. S. (1980). Evolution of dioecy in flowering plants. Ann Rev Ecol Syst, 11: 15–39.
Belhassen, E. (1987). Dissémination et voisinage chez une espèce gynodioïque: le cas de Thymus vulgaris L. Génét Sél Évol, 19: 307–320.
Belhassen, E., Trabaud, L., Couvet, D. and Gouyon, P. -H. (1989). An example of nonequilibrium processes: gynodioecy of Thymus vulgaris in burned habitats. Evolution, 43: 662–667.
Belhassen, E., Dommée, B., Atlan, A., Gouyon, P. -H., Pomente, D., Assouad, M. W. and Couvet, D. (1991). Complex determination of male sterility in Thymus vulgaris L.: genetic and molecular analysis. Theor Appl Genet, 82: 137–143.
Belhassen, E., Atlan, A., Couvet, D., Gouyon, P. -H. and Quetier, F. (1993). Mitochondrial genome of Thymus vulgaris L. (Labiatae) is highly polymorphic within and among natural populations. Heredity, 71: 462–472.
Boutin, V., Pannenbecker, G., Ecke, W., Schewe, G., Saumitou-Laprade, P., Jean, R. et al. (1987). Cytoplasmic male sterility and nuclear restorer genes in a natural population of Beta maritima: genetical and molecular aspects. Theor Appl Genet, 73: 625–629.
Burdon, J. J. and Jarosz, A. M. (1992). Temporal variation in the racial structure of flax rust (Melampsora lini) populations growing on natural stands of wild flax (Linum marginale): local vs. metapopulation dynamics. Pl Pathol, 41: 165–179.
Corriveau, J. L. and Coleman, A. W. (1988). Rapid screening method to detect potential biparental inheritance of plastid DNA and results for over 200 angiosperm species. Am J Bot, 75: 1443–1458.
Couvet, D., Bonnemaison, F. and Gouyon, P.-H. (1986). The maintenance of females among hermaphrodites: the importance of nuclear-cytoplasmic interactions. Heredity, 57: 325–330.
Couvet, D., Atlan, A., Belhassen, E., Gliddon, C., Gouyon, P. -H. and Kjellberg, F. (1990). Coevolution between two symbionts: the case of cytoplasmic male sterility in higher plants. In: Futuyma, D. & Antonovics, J. (eds) Oxford Surveys in Evolutionary Biology, vol. 7, pp. 225–249. Oxford University Press, Oxford.
Cuguen, J., Wattier, R., Saumitou-Laprade, P., Forcioli, D., Morchen, M., Van D ijk, H. and Vernet, P. (1994). Gynodioecy and mitochondrial DNA polymorphism in natural populations of Beta vulgaris ssp. maritima Génét Sél Évol, 26: 87–101.
De Haan, A. A., Hundscheid, M. P. J. and Van Hinsberg, A. (1997). Effects of CMS types and restorer alleles on plant performance in Plantago lanceolata L: an indication for cost of restoration. J Evol Biol, 10: 803–820.
Dommée, B., Assouad, M. W. and Valdeyron, G. (1978). Natural selection and gynodioecy in Thymus vulgaris L. Bot J Linn Soc, 77: 17–28.
Dommée, B., Guillerm, J. L. and Valdeyron, G. (1983). Régime de reproduction et hétérozygotie des populations de thym Thymus vulgaris L., dans une succession post-culturale. C r Acad Sci Paris, 296: 111–114.
Ebert, D. (1994). Virulence and local adaptation of a horizontally transmitted parasite. Science, 265: 1084–1086.
Frank, S. A. (1991). Ecological and genetic models of host-pathogen coevolution. Heredity, 67: 73–83.
Gandon, S., Capowiez, Y., Dubois, Y., Michalakis, Y. and Olivieri, I. (1996). Local adaptation and gene-for-gene coevolution in a metapopulation model. Proc R Soc B, 263: 1003–1009.
Gouyon, P. -H., Vichot, F. and Van Damme, J. M. M. (1991). Nuclear-cytoplasmic male sterility: single-point equilibria vs. limit cycles. Am Nat, 137: 498–514.
Khey-Pour, A. (1980). Nucleo-cytoplasmic polymorphism for male sterility in Origanum vulgare L. J Hered, 71: 253–260.
Koelewijn, H. P. and Van Damme, J. M. M. (1995). Genetics of male sterility in gynodioecious Plantago coronopus I. Cytoplasmic variation. Genetics, 139: 1749–1758.
Lively, C. M. and Jokela, J. (1996). Clinal variation for local adaptation in a host-parasite interaction. Proc R Soc B, 263: 891–897.
Manicacci, D., Atlan, A. and Couvet, D. (1997). Spatial structure of nuclear factors in sex determination in the gynodioecious Thymus vulgaris L. J Evol Biol, 10: 889–907.
Manicacci, D., Couvet, D., Belhassen, E., Gouyon, P.-H. and Atlan, A. (1996). Founder effects and sex-ratio in the gynodioecious Thymus vulgaris L. Mol Ecol, 5: 63–72.
Nee, S. (1989). Antagonistic coevolution and the evolution of genotypic randomization. J Theor Biol, 140: 499–518.
Parker, M. A. (1985). Local population differentiation for compatibility in an annual legume and its host-specific fungal pathogen. Evolution, 39: 713–723.
Reboud, X. and Zeyl, C. (1994). Organelle inheritance in plants. Heredity, 72: 132–140.
Ronfort, J., Saumitou-Laprade, P., Cuguen, J. and Couvet, D. (1995). Mitochondrial DNA diversity and male sterility in natural populations of Daucus carota ssp. carota Theor Appl Genet, 91: 150–159.
Saumitou-Laprade, P., Rouwendal, P., Cuguen, J., Krens, F. A. and Michaelis, G. (1993). Different CMS sources found in Beta vulgaris ssp. maritima mitochondrial variability in wild populations revealed by a rapid screening procedure. Theor Appl Genet, 85: 529–535.
Saumitou-Laprade, P., Cuguen, J. and Vernet, P. (1994). Cytoplasmic male sterility in plants: molecular evidence and the nucleocytoplasmic conflict. Trends Ecol Evol, 9: 431–435.
Schultz, S. T. (1994). Nucleo-cytoplasmic male sterility and alternative routes to dioecy. Evolution, 48: 1933–1945.
Tarayre, M. and Thompson, J. D. (1997). Population genetic structure of the gynodioecious Thymus vulgaris L. (Labiatae) in southern France. J Evol Biol, 10: 157–174.
Van Damme, J. M. M. (1982). Gynodioecy in Plantago lanceolata L. I. Polymorphism for plasmon type. Heredity, 49: 303–318.
Van Damme, J. M. M. (1983). Gynodioecy in Plantago lanceolata L. II. Inheritance of three male sterility types. Heredity, 50: 253–273.
Acknowledgements
We wish to thank M. Beltran for help with the experimental work, D. Manicacci, D. Couvet and P.-H. Gouyon for valuable comments on an earlier version of the manuscript, O. Kaltz and B. Schmid for useful advice on statistical analyses and two anonymous reviewers for constructive comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gigord, L., Lavigne, C., Shykoff, J. et al. No evidence for local adaptation between cytoplasmic male sterility and nuclear restorer genes in the gynodioecious species Thymus vulgaris L.. Heredity 81, 156–163 (1998). https://doi.org/10.1046/j.1365-2540.1998.00388.x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.1998.00388.x
- Springer Nature Switzerland AG