
Abstract
Birds have a diverse acoustic communication system, and the ability to recognise their own species’ song from a distance facilitates complex behaviours related to mate attraction and rival deterrence. However, certain species, including doves, do not learn songs and their vocal repertoires are much simpler than those of better-studied songbirds. In these so-called non-learning birds, relatively little is known about the role that bird song plays in intra- and interspecific interactions, and how such behaviours might be acquired (inherited or learned from experience). To investigate this question, we focused on two species of African wood doves whose long-range songs are used in a territorial context. Specifically, we examined the responses of sympatric and allopatric populations of male blue-headed wood-doves (Turtur brehmeri) and tambourine doves (Turtur tympanistria) to different types of simulated territorial intrusions, i.e. playback of conspecific, congeneric, and control songs. We aimed to assess (i) whether these species, which have similar songs, respond only to their own species' song or exhibit interspecific territoriality, and (ii) if the response pattern is affected by the presence or absence of congeners in the general area. We found that both species responded strongly to playback of their own species in both sympatric and allopatric populations. In allopatry, though, male tambourine doves misdirected their response and also approached the playback of congeneric songs. Our results indicate that, in areas where the studied Turtur doves live in sympatry, they do not exhibit consistent interspecific territoriality. However, we cannot exclude the possibility that the smaller tambourine dove avoids its larger congener during the process of territory establishment. The difference in tambourine doves’ response toward the song of present (sympatric) or absent (allopatric) congeners suggests that the ability to discriminate between songs of similarly singing potential competitors is acquired through earlier interactions and learning. This plasticity in response supports the misdirected aggression hypothesis, which argues that interspecific territorialism emerges as a maladaptive by-product of signal similarity. However, on an evolutionary timescale, such an ability could be considered an adaptive cognitive tool useful for resolving competing interests with congeners.
Similar content being viewed by others
Introduction
Acoustic communication is widespread in many animal taxa, but the evolution of complex sound signals has occurred in only a few groups. One such group is birds, whose songs exhibit an extraordinary degree of diversity and sophistication1,2. Apart from some basic pre-adaptations for vocal communication found in the ancestors of modern birds (e.g., breathing apparatus), the main factor that led to the evolution of elaborate songs was sexual selection. To wit, the most important functions of bird song are attracting a breeding partner and defending territory or other resources2. These two functions are not mutually exclusive and were historically assigned to males, as males were thought to be the dominant singing sex3. To fulfil these functions, songs must be species-specific as well as sufficiently individually unique to enable discrimination or recognition of individuals or classes of individuals (e.g., stranger vs neighbour)2,4. This two-level specificity of the song is then interpreted by the receiver for use in decision making, even in the absence of visual information5.
In species where only the male sings, the meaning of a song could be different from the perspectives of male and female receivers. When a female recognises the song of her own species, she can, in the next step, evaluate the quality (or other features like dialect origin) of the sender and decide whether to reproduce with him6,7. A male receiver, on the other hand, is usually focused on extracting information about the threat level expressed by the signal. For example, when territory boundaries are established, the song of a neighbour singing from his territory border is usually treated as less dangerous than the song of a stranger8. The evaluation of a potential rival's song enables the territory owner to make an appropriate decision about interacting with that rival9,10. Responding too aggressively may lead to a risk of injury or death in the worst case or wasted energy in the best case10,11. For the territory owner and potential mate, a long-distance signal carrying a message about species and individual identity is usually crucial and a prerequisite for territorial behaviour2.
Interspecific territoriality
Territoriality is widespread among animals12,13,14,15. Within a species, individuals prefer similar habitats and must compete for limited resources, but the exact pattern and level of aggressiveness exhibited during competition depend on their life histories (e.g., sedentary vs migratory lifestyle, breeding system)16,17. Initially, between-species interactions were assumed to be rare, costly, and the result of ancestral signal similarity. Surprisingly, recent comparative studies have revealed that interspecific territoriality is common in birds18,19. To explain the evolution of interspecies territorialism, four basic hypotheses have been proposed (reviewed in Cowen et al.20). The first is the resource competition hypothesis, which assumes that resources are partitioned spatially between dominant individuals of competing species21,22,23. The second hypothesis, asymmetric competition, is also based on strong resource rivalry but assumes that only dominant males of the dominant species succeed in interspecific territoriality24. The reproductive interference hypothesis explains the existence of territorial aggression between species as the result of competition not for the same resources but for a partner, due to incomplete reproductive isolation and the possibility of hybridisation25,26. The last hypothesis, misdirected aggression, posits that interspecies territorialism is maladaptive and a by-product of intraspecies territorialism15,27.
These hypotheses reflect field observations and are not mutually exclusive. To shed light on this phenomenon, then, researchers must search for evolutionary scenarios in which these different options are possible, rather than try to eliminate erroneous hypotheses. Interspecific territoriality is more likely if species have somewhat similar songs and can physically interact20. Hence, the first step for understanding the process of such territoriality is examining if a species detects the signal of a congeneric and recognises it as worthy of a response. The hypotheses presented above provide some background for when, why, and how this might happen. As predominantly territorial animals, birds are particularly interesting models for studying interspecific territoriality because different groups of species are diametrically opposed to each other in the way acoustic signals are transmitted between generations (inherited vs learned). In addition, territorial defence in birds involves processing signals heard from a distance, and at least the initial part of the response is evoked solely by the sound signal2.
Aims of the study
One interesting scenario for comparing potential interactions between species is when closely related species exploit similar resources and have similar signals. In such a case, interspecific interactions may be the result of competition, leading to interspecific territoriality and an intentional response to the other species (including avoidance of a 'stronger' competitor). However, interactions could also, at least potentially, be due to mistakes (similar signals). In addition, the character of such interactions might depend on whether the studied pair of species naturally occur in the same location (sympatry) or not (allopatry).
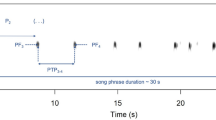
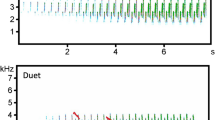
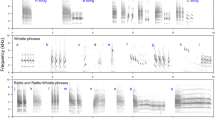
In this study, we experimentally tested the response of two non-learning, territorial bird species to conspecific and congeneric songs in both the sympatric and allopatric ranges. The main objectives were to see whether and how the studied species respond to each other's songs and how this response might change in areas in which the rival is not naturally found. Our models were the blue-headed wood-dove, Turtur brehmeri (hereafter brehmeri), and the tambourine dove, T. tympanistria (hereafter tympanistria). Genus Turtur contains a total of five species living in varied habitats of sub-Saharan Africa, and all have very similar songs that are used for long-distance communication. The songs consist of simple, low-frequency notes organised with generally similar syntax (songs have two parts differing in the duration of syllables, pauses, and peak frequency) and are quite different in comparison to other doves (Fig. 1). The model species both inhabit forests, but brehmeri has a range limited to central and western Africa, while tympanistria can be found in varied, forested habitats in almost all of sub-Saharan Africa28,29.

Spectrograms of the songs of focal bird species: blue-headed wood-dove (Turtur brehmeri) and tambourine dove (Turtur tympanistria), and species used as a control: yellow-billed touraco (Tauraco macrorhynchus) and narina trogon (Apaloderma narina).
In the area of sympatric occurrence, we tested both dove species. We expected birds to demonstrate a strong response to their own species’ song. If the two species indeed compete in these habitats, the smaller species should avoid the larger intruder. For this reason, we predicted that tympanistria males would not respond to the congener's song or might even exhibit extreme avoidance behaviours (like flying away and ceasing to sing). Instead, the larger brehmeri should attempt to chase away its smaller competitor. Thus, we also expected that brehmeri males might respond to the tympanistria song, but not necessarily as strongly as to that of conspecific rivals. If the species do not exhibit interspecific territoriality, however, we would not expect to observe any response to the songs of congeners.
For our allopatric sites, we conducted experiments in a forest where only tympanistria occurs. Again, we expected a strong response to the conspecific song. Regarding the response to the absent congener, we investigated the possibility of two potential scenarios. Males of tympanistria may not respond to an unknown congener’s song if it is not recognised as a potential competitor. Such a result would suggest that recognition of species-specific songs is inherited, and the relatively small differences between tympanistria and brehmeri songs facilitate efficient recognition even in the absence of any earlier experience with the congeneric species. However, if the allopatric tympanistria males respond to the song of a congener, it would support the misdirected aggression hypothesis, i.e., it is likely to be a case of mistaken identity due to the general similarity of both species' songs. It would also imply that any kind of response found between tympanistria and brehmeri in sympatry must result from earlier interactions between individuals of both species.
Methods
Study sites and species
The study was conducted in two sites in sub-Saharan Africa, one where brehmeri and tympanistria occur in sympatry and one where only tympanistria is present. The first site was in Kakum National Park, in the coastal region of southern Ghana (5.20–5.40 N, 1.30–1.51 W, altitude 135–250 m asl). The park covers an area of ~ 210 km2 of tropical moist evergreen forest with fragments of periodic or permanent swamp and riverine forests. Although logging operations between 1975 and 1989 decreased areas of dense primary forest, the logged areas have regenerated secondary forests characterised by denser vegetation. Kakum NP receives an annual average rainfall of 1460 mm, with most precipitation in September–October and March–June. Temperatures vary between 20 and 30 °C30,31. The second site was in Kibale National Park in Uganda (0.13–0.41 N, 30.19–30.32 E, altitude 1100–1600 m asl), a moist, evergreen, medium-altitude forest with a mosaic of primary and secondary forest, swamp, grassland, and thicket. Temperatures are very stable over the course of the year (daily fluctuation 15–27 °C) and the precipitation (annually around 760 mm) has a bimodal pattern, with more rainfall in March–April and September–November32. There appears to be no detectable bird seasonality in this area33.
In this study, we focused on two Turtur species that prefer strictly forest-like habitats. The more ubiquitous tympanistria is typically found in any type of forest or forest-like habitat, such as lowland and montane forests (even up to 3000 m asl), secondary forests, gardens, plantations, or even mangroves29. The range of brehmeri is restricted to western Africa and lowland primary forests up to 750 m asl only. Occasionally it is also found in old secondary forests, but generally, it avoids heavily disturbed forests28. With a body mass of 98.5–129.5 g (N = 10), brehmeri are significantly (t2,50 = 19.6, P < 0.0001) larger than tympanistria (51.5–81 g; N = 42; own measurements in the study area). From our own observations and the literature, we found no evidence of hybridisation between these two species34. Furthermore, a STRUCTURE analysis performed on 7000 SNP markers (obtained by sequencing RADseq libraries) did not detect any signal of admixture between brehmeri and tympanistria (own unpublished data based on 6 brehmeri and 26 tympanistria individuals). We did not observe any direct, physical interaction between males of these species in the sympatric area, although both species were heard singing relatively close to each other. However, the above observations do not exclude the possibility of interspecific territoriality completely.
Playback stimuli
We prepared three types of playback: (i) conspecific (brehmeri and tympanistria for brehmeri and tympanistria, respectively), (ii) congeneric (brehmeri for tympanistria and tympanistria for brehmeri), and (iii) a control (yellow-billed tauraco, Tauraco macrorhynchus, in Kakum NP and narina trogon, Apaloderma narina, in Kibale NP). The control species were chosen because their territorial calls have similar spectra to the territorial songs of doves. Both species are common in the studied areas but are not perceived as a threat by doves and do not interact with them. To create playback stimuli of doves, we selected recordings of territorial songs of local non-neighbouring males with a high signal-to-noise ratio. Each song sample was filtered (high-pass, 0.1 kHz; low-pass, 1.5 kHz). Each playback stimulus was created from a sample belonging to a different individual to avoid pseudo-replication. Dove playback consisted of a single song sample repeated 10 times (two songs per minute), which is the typical calling rate of both brehmeri and tympanistria. Comparable song durations and pacing were also maintained for the control treatments. The amplitude of each playback was set to 82 ± 2 dB SPL at a 1-m distance from the speaker, measured with a CHY 650 digital sound level meter (CHY Firemate Co., Ningbo, China). All playback stimuli were created with Raven Pro 1.6 (Cornell Lab of Ornithology, Ithaca, USA) and Avisoft SASLab Pro 5.2 (Raimund Specht, Berlin).
Playback experiment design and procedures
General procedures of experiments I–III
Each male was tested three times (conspecific, congeneric, and control species treatments) in a counterbalanced order. Experiments were only conducted when a male was observed and heard singing before the playback. In each trial with a given male, the speaker was placed in a slightly different location to avoid the habituation of the focal bird, but always within the same distance (~ 50 m) of the place from which the bird was singing before playback and at the same height above the ground, ca. 2 m. To broadcast the playback stimuli, we used either a Tascam DR-40X (TEAC Europe GmbH, Wiesebaden, Germany) or Sony PCM-D100, and JBL Charge 4 speakers (Harman International Industries, Stamford, Connecticut, USA). Before starting the 5-min playback, we recorded the focal male for at least 1 min. Recorded songs were then used for checking if the responding individuals were the same as before the start of playback. The identity check was based on comparing the time and frequency distribution of the initial syllables in songs (see Fig. S1). This method was developed for tympanistria and allows for even 96% classification efficacy, depending on the number of compared syllables35. Analogous measurements allow for differentiating between males of brehmeri at a similar level (unpublished own data). Playback was then followed by a 5-min post-playback phase.
Each trial was observed by two people positioned on opposite sides of the speaker, ca. 20 m away from it, in locations that minimised the possibility of disturbing a bird approaching the speaker. The person recording songs had a lavalier microphone connected to the second channel of the main recorder for dictating observations. The second observer was on the opposite side of the speaker in a place convenient for observing the whole experimental scene. The birds' behavioural responses were recorded by dictating observations into the additional recorder. Recordings of vocalisations and comments on physical behaviour were later time-aligned and combined into one audio file. All distances reported (as the closest distances to the speaker) were measured with a Leica DISTO D510 laser range finder.
Our preliminary work with the study species revealed they are shy and that observing their behaviour in the forest is difficult. Therefore, we were focused a priori on those aspects of behaviour that we were able to quantify with sufficient and repeatable certainty among all experiments. In the case of the vocal response, we used the number of songs sung by males. The physical behavioural responses of males were measured as the time spent within 25 m of the speaker (s), latency to approach the speaker (s), the closest approach to the speaker (m), and the number of flights.
Experiments in sympatric area
Playback for Experiment I (with brehmeri) and Experiment II (with tympanistria) was performed between November 12 and December 1 in 2021 between 6:04 and 12:43 local time (sunrise started 5:52–5:58) in Kakum NP. All males were tested three times, with the three types of songs (conspecific, congeneric, and control song of yellow-billed turaco) presented in a counterbalanced order. The time of testing reflected birds' activity and depended on the weather on a particular day (in practice, single trials were performed after 12:00 on two separate occasions when we waited longer for birds after two earlier tests). Altogether, we tested 19 males of brehmeri and 14 males of tympanistria. The average time between the following treatments with the same male was 32 min for Experiment I and 39 min for Experiment II.
Experiment in allopatric area
Playback for Experiment III was performed between June 28 and July 6 in 2022. We tested birds during their active time in the morning, 6:58–9:47 (sunrise started between 5:59 and 6:00), and in the late afternoon, 16:44–17:48 (sunset started between 18:03 and 18:05). Each male was tested only in the morning or the evening, as their pattern of activity was strictly bimodal and short due to weather conditions. Altogether, in Kibale NP we tested 15 males of tympanistria. The three songs of each trial were presented in a counterbalanced order, exactly as in Experiments I-II, and differed only in the species used for the control (narina trogon). The average time between subsequent treatments with the same male was 21 min.
Ethical note
To our knowledge, the individuals tested in the experiment reflected the population in a representative way with no potential biases resulting from social background, self-selection, habituation, or other factors as indicated in the STRANGE framework (Webster and Rutz 2020). This study was designed and performed in accordance with the ARRIVE guidelines36. Our experimental procedure adhered to the ASAB/ABS Guidelines for the care and use of animals (The Ethics Committee (ASAB) and the Animal Care Committee (ABS), 2019) and was approved by all responsible local bodies listed below as well as by the Polish Laboratory Animal Science Association (certificate no. 1952/2015 to TSO, conforming to Directive 2010/63/EU). Our experimental procedures were approved in Ghana by the Forestry Commission (Wildlife Division), permit no. WD/A.185/Vol.13/80, and in Uganda by Makerere University (College of Health Sciences; Makerere University Biological Field Station) and the Uganda Wildlife Authority, permit no. COD/96/05 and Research Material Transfer MTA no. 377, as well as the Uganda National Council for Science and Technology, permit no. NS256ES.
Statistical analysis
The original response variables we measured during experiments were partly correlated with each other, but the multicollinearity was moderate (variance inflation ratio VIF between 1.02 and 3.42). Therefore, to analyse the general strength of the response to playback, we used a principal component analysis (PCA) with varimax rotation and Kaiser normalisation (IBM SPSS Statistics 28.0.1.0). We extracted principal components separately for all three experiments. All three datasets were well suited for PCA (KMO and Bartlett's tests are given in Tables 1, 2, 3); the first two extracted components explained a similar percentage of the variance (PC1: 43.77–47.93% and PC2: 25.52–30.40%) and had a similar pattern of loadings from the original variables. All the first components had heavier loadings from variables related to the approach to the speaker (Tables 1, 2, 3), while all the second components had heavier loadings from the variables related to singing. Therefore, we refer hereafter to PC1 as the ‘Approaching’ component and to PC2 as the 'Vocal response' component (Tables 1, 2, 3). Higher values of PC1 indicate more flights during the playback, a closer approach to the speaker, and more time spent in its vicinity, hence, a stronger response. Higher values of PC2 indicate that responding birds sang more songs both during and after playback.
To test for differences in the response to conspecific, congeneric, and control songs, we built generalised linear mixed-effects models (GLMM) using the 'lme4' package of R37 and checked model assumptions using the DHARMa package38. Our response variables were the measures of approaching (PC1) and vocal response (PC2) extracted separately for each experiment. We included in our models two main factors: (1) playback treatment (three levels: conspecific, congeneric, and control) and (2) playback order (three levels: first, second, or third). We included all first-order interaction terms and incorporated male identity as a random effect. For model selection, in all above analyses we adopted an information theoretic approach39. We ranked all possible models mentioned above according to their value of Akaike's information criterion corrected for small sample size (AICc), and obtained an averaged model by selecting the most supported ones (ΔAICc ≤ 6) after the exclusion of uninformative parameters using the MuMIn package in R40. Models were not over-dispersed (GLMMExperiment I,PC1: P = 0.936; GLMMExperiment I,PC2: P = 0.96; GLMMExperiment II,PC1: P = 0.92.; GLMMExperiment II,PC2: P = 1.0; GLMMExperiment III,PC1: P = 0.856; GLMMExperiment III,PC2: P = 0.88), no outliers were detected (all GLMMs: P = 1, except GLMMExperiment II,PC1: P = 0.285), and visual inspection of the Q-Q plots confirmed the normality of the residuals with a single exception at the marginal significance level (Kolmogorov–Smirnov test, GLMMExperiment I,PC1: P = 0.046; GLMMExperiment I,PC2: P = 0.671; GLMMExperiment II,PC1: P = 0.336; GLMMExperiment II,PC2: P = 0.964; GLMMExperiment III,PC1: P = 0.705; GLMMExperiment III,PC2: P = 0.570). We applied post-hoc contrasts to differentiate between the different levels of treatments. All P values reported are two-tailed.
Results
The two studied species exhibited a similar approaching response (PC1) to playback of their own species in both sympatry and allopatry (all Experiments I–III; Tables 4, 5, 6). The tested birds flew close to the speaker during playback, and the closer they approached, the longer they stayed close (Figs. 2, 3, 4). In the site where the two doves are sympatric, the response pattern to the conspecific, congeneric, and control playback was very similar in both species: males approached only when conspecific songs were played, and not those of congeners or the control species (all post-hoc comparisons are presented in Table S1). In the allopatric site (no brehmeri), instead, tympanistria males demonstrated similar approach responses, and we did not find statistically significant differences in responses to playback of songs of their own species and those of the absent congener (see post-hoc tests in Table S1; Fig. 4). Furthermore, the responses to both conspecific song (P = 0.0002) and congeneric song (P = 0.0005) were significantly different from the response to the control song (details in Table S1).

Experiment I: blue-headed wood-dove males responses to playback of conspecific, congeneric and control songs in sympatry (Ghana), measured with compound measures of response: PC1-approaching and PC2-vocal response.

Experiment II: tambourine dove males responses to playback of conspecific, congeneric and control songs in sympatry (Ghana), measured with compound measures of response: PC1-approaching and PC2-vocal response.

Experiment III: tambourine dove males responses to playback of conspecific, congeneric and control songs in allopatry (Uganda), measured with compound measures of response: PC1-approaching and PC2-vocal response.
The vocal responses (PC2) were less consistent among species and experiments. Males of brehmeri approaching the playback of their own species decreased their song rate during playback (Fig. 2, Table 4), and often continued searching for an intruder after the playback finished instead of resuming singing. Differences in vocal response to the playback of their own species compared to congeners and controls were highly significant (see Table S1). In the case of tympanistria, males' song behaviours were quite variable, and we found no significant differences between treatments (Tables 5, 6, S1). We found that the order of treatments had no significant influence on the response of tested birds; however, leaving the order out of certain models improved the results (Tables 4, 5, 6).
Discussion
In this study, we experimentally tested the effect of songs of conspecific, congeneric, and neutral control individuals on the territorial response of two closely related species of African wood doves living in sympatry and allopatry. We found that males of both species always responded strongly to the conspecific stimuli, while the single species tested in allopatry (tympanistria) also approached the speaker when the unfamiliar song of the absent congener was played. Neither species responded to the control song, providing a useful contrast to the responses to perceived incursions by conspecific and congeneric rivals.
Interspecific territoriality
The differences we observed between the responses to playback of conspecific song compared to congeneric song and to control song in sympatry and allopatry shed new light on the mechanisms underlying song recognition for members of the same species in addition to competition and potential interspecific territoriality in non-learning birds, represented here by African wood doves. Our results do not support the resource competition hypothesis, which posits that resources are partitioned, and defended, by dominant individuals of competing species20. Support for this hypothesis would be found in a strong response (with approaching as a proxy of aggression) by both species to both conspecific and congeneric song when they are found in sympatry. Instead, we found that, in sympatry, the studied Turtur doves only approached signals of their own species, while the response to congeneric song was similarly weak (or none) as to control song.
On the other hand, the lack of a strong response from larger brehmeri males to the songs of their smaller congener, together with the fact that the territories of the two species do not overlap within the sympatric range, could provide partial support for the asymmetric competition hypothesis, assuming that the larger species is dominant in interspecific territoriality18,20,24. As we have no information about possible hybridisation between species34 and differences in size and colouration between brehmeri and tympanistria are substantial28,29, we suspect that the reproductive interference hypothesis is unlikely to be relevant for this pair of species20. However, the apparent approaching response of tympanistria to the song of the absent brehmeri in allopatry does provide support for the misdirected aggression hypothesis15,20.
It is not unusual to find asymmetry between the response to members of one’s own species and the response to related species, or even different dialects of the same species. For example, Hamao41 found that sympatric, related species differ in their responses to conspecific song dialects, using as models the Japanese tit (Parus minor) and the varied tit (Poecile varius). In a later study, he also found that the responses of varied tits to heterospecific songs were much weaker in sympatric populations than in allopatric ones42. The difference between Hamao's studies41,42 and the present work is that the species he tested were not congeners but confamilial (Parus vs Poecile), and that, unlike those two species, wood doves do not learn their songs2. The exact degree of phylogenetic distance between interacting species is likely not very important. As revealed by Drury et al. (2020), in the context of interspecific territoriality, song as a territorial signal acts at a deeper (even between-family) evolutionary timescale. Moreover, interspecific territoriality is known to occur even between unrelated species if they are forced by local conditions to compete for resources, as was observed for the chaffinch (Fringilla coelebs) and great tit (Parus major) on islands near Scotland43. However, interspecific interactions between non-learning birds are less known and require deeper consideration.
Is song learning important for territoriality?
Song production in doves does not involve cultural transmission44,45. However, it is worth remembering that there are different forms of learning in acoustic communication is worth remembering. Janik and Slater46 distinguished between production learning and two forms of contextual learning, where individuals may produce an existing signal in a new context (usage learning) or a receiver extracts a novel meaning from a signal (comprehension learning). In both cases of contextual learning, the experience based on interactions with other individuals is crucial. So it is worth considering how potential competitors might perceive the presence or absence of a species with similar requirements and signals. Although the differences in song structure between Turtur spp. are small, they are consistent and allow for acoustic discrimination between species. In the case of brehmeri and tympanistria, their songs overlap in duration, number of notes, and frequency, and both species' songs can be divided into two distinct parts (own unpublished data). However, they clearly differ in the pattern of within-song note production over time (Fig. 1). In brehmeri songs, pauses between notes decrease constantly with time, while in tympanistria, pauses in the initial part of songs are variable in length. Moreover, tympanistria individuals differ greatly from one another in these between-note intervals, which allows for individual recognition35. We do not yet understand the mechanisms of species recognition in doves, particularly whether the response to conspecific songs is inherited or learned. However, pigeons are known to pay attention to the intervals between pulses, and it is likely that the above-mentioned differences in the time pattern of notes are important for species recognition47,48. For example, the perch-coo vocalisations of Streptopelia doves are long-range signals that are used for species recognition based solely on acoustic cues, and their temporal parameters were found to be the most salient features for recognition49,50,51. The song of the Turtur doves we studied here seems to be the functional equivalent of the perch-coo of Streptopelia doves, i.e. a signal that is produced to reach a receiver in the distance, who at the moment of calling is usually out of sight52.
Doves are known to use the same brain areas for recognition of species-specific vocalisations as songbirds53,54. Experiments on Streptopelia doves indicated that they are capable of learning discrimination tasks in an operant set-up based on conspecific and heterospecific vocalisations49,55. Here, the contrast we observed between tympanistria’s lack of response to congeneric songs in sympatry and the strong response in allopatry suggests that tympanistria living without related species nearby cannot differentiate between the song of their own species and the song of congeners. In the allopatric site, we observed a clear movement response to brehmeri playback, comparable to the response to conspecifics. Hence, our results suggest that individuals with no previous experience of interacting with the congeneric species categorised songs of similar general structure as meriting a response. At least at first, when they heard the song of the congener, they did not appear to perceive it as different from their own. Consistently, this also means that the lack of tympanistria response to brehmeri song in sympatry is a form of a learned 'not responding' by the meaning of contextual learning sensu Janik and Slater46.
Similar experiments on non-learning bird species are scarce. However, one interesting example was reported for flycatchers belonging to genus Empidonax. These members of the Tyrannidae family (and Suboscines) are known to develop species-specific songs without learning56,57. Initial studies on alder (Empidonax alnorum) and willow (E. trailii) flycatchers revealed little response to heterospecific song playback in sympatry58. However, when both species were tested in areas where they shared habitats (i.e., overlap in micro-scale), they were found to respond aggressively to heterospecific songs59.
Even more remarkable was the recent finding on sympatric rallids, which are also non-learners (phylogenetically even more distant from any song-learning bird taxa than Tyrannids). Jedlikowski et al. (2022) showed that the water rail, Rallus aquaticus, and the little crake, Zapornia parva, are able to distinguish not only each other but also specific individuals of the other species. This work provided an example of a dear-enemy effect in different species. Based on these studies, it seems that being a non-learner in the context of the acquisition of vocalisation (socially learned vs inherited) does not necessarily limit a bird’s abilities to diversify its responses toward different individuals of the same and even other, potentially competing, species.
The response to heterogenic song was also tested experimentally in other African dove taxa, although in a different context than in our study: hybridisation. In Uganda, de Kort et al.52,61 studied two closely related doves (vinaceous dove, Streptopelia vinacea, and ring-necked dove, S. capicola) as well as hybrids between the two. The authors focused on perch-coo and bow-coo signals, which both play a role in inter- and intra-sexual signalling but are produced in behaviourally different contexts (long vs short distance communication). They found that allopatric populations showed a stronger response to conspecific than to heterospecific perch-coos, but equal responses to bow-coos of either species. Instead, hybrids exhibited no clear pattern between their own coo structure and that of the species to which they responded most strongly, indicating a lack of behavioural coupling. Interestingly, in allopatry, ring-necked doves responded more strongly to perch-coos of vinaceous doves than vice versa, which the authors explained based on their ecological history. Namely, the ring-necked dove had expanded into the vinaceous dove’s range, thus creating a context in which rapid learning of a novel (heterospecific) competitor’s song might be favoured52.
Overall, then, research on other non-learning bird species has revealed flexibility in their response patterns to conspecific and congeneric (or even more evolutionary distant) individuals depending on the ecological and evolutionary context.
An ecological perspective on interactions between wood doves
In the sympatric area, where brehmeri and tympanistria co-occur and their territories are often adjacent, we observed no approaching response to both congeners' songs and control songs in our experiments. From several observation posts, males of both species could be heard simultaneously, but the distance between song posts was typically more than 50–100 m, and their territories seemed not to overlap. In several patches, only one of the two species was present. These observations have two potential explanations: (i) both species can discriminate between the songs of conspecifics and those of congeners, or (ii) the weaker competitor can discriminate, and it uses this information to make decisions regarding settlement in sympatry. Our field data suggest that in Kakum NP, brehmeri males choose territories deeper in the forest, containing the highest trees close to streams, while tympanistria were often found close to forest edges or partly open areas, e.g., in tree fall gaps (or secondary forest). In Kibale NP, instead, tympanistria were most often found in sites like those preferred by brehmeri in Kakum NP. This suggests that, in sympatry, smaller tympanistria males may avoid settlement close to (or within the territory of) brehmeri, perhaps as the result of initial competitive interactions with their congeners. Such a mechanism would support the asymmetric competition hypothesis, with the caveat that the birds do not have to compete all the time. Data on the life history of wood doves are scarce but based on re-captures and recording the same birds in the same territories year after year, it appears that their territories are stable for a long time, maybe even their whole lives. Like many birds in the tropics, they are likely to be long-lived if they succeed in reaching adulthood62. This may suggest that the response pattern to congeners (and members of their own species) is settled when young males try to establish their territories for the first time. Based on this, and on the observations we made in our experiments, we hypothesise that when the two species are found in sympatry, young tympanistria likely interact with brehmeri males but are chased away by the stronger rival, and thus avoid such confrontations later in life. The ability to discriminate between conspecific and congeneric individuals is thus acquired through life experience, enabling the bird to save time and energy later in life. This is a hypothetical but probable scenario for the coexistence of these two species in sympatry, which also explains the pattern of responses observed in allopatry.
Evolutionary perspective (functionality of response to congener's song in allopatry)
Our results suggest that tambourine doves may have some inherited template memory, but that response control is likely to develop while listening and interacting with potential rivals and/or mates. Therefore, we can assume that in sympatry, tympanistria must have learned not to respond to a song of a stronger (congeneric) rival (comprehension learning)46. This ability to learn responses to songs similar to their own opens the door for between-species interactions and territoriality in case of changes in species range. From this point of view, we should reconsider whether or not responding to a sister species in the allopatric zone can truly be considered maladaptive, as argued by the misdirected aggression hypothesis. In Kibale NP, where the congener is not naturally found, this response could indeed be viewed as maladaptive. Still, from a long-term and broad-scale perspective, this pattern may represent a plastic ability to find the best response in a changing environment that may or may not contain individuals of both species. Therefore, the misdirected aggression hypothesis may not be, by definition, maladaptive. Species ranges can change as a result of many factors, including in some dove species63,64, and the ability to adapt (i.e., to respond functionally) to a new competitor may be crucial on the evolutionary time-scale. In this context, our experimental results are consistent with a recent comparative study that revealed, through a large-scale phylogenetic analysis, that interspecific territoriality is widespread in birds and is strongly associated with hybridisation and breeding habitat overlap18.
The pair of Turtur species studied here seems to be a good model for investigating responses to heterospecific song under different forms of allopatry. T. tympanistria has a larger species range and less-specific habitat preferences than brehmeri; it can thus be found at a distance of hundreds of kilometres to the closest brehmeri population, as well within basically the same area but at different elevations, such as on Mount Cameroon28,29,65. It would be extremely valuable to understand how tympanistria from different allopatric populations respond to the song of congeners. For example, a study on sibling species of African sunbirds revealed that responses to heterospecific song could be strongly different among allopatric populations66. Conversely, another tropical species, the white-eared ground-sparrow (Melozone leucotis) used information encoded in vocalisations to discriminate competitor from noncompetitor species even in the absence of previous experience (understood as living in sympatry or allopatry with Prevost’s ground-sparrow, Melozone biarcuatum)67. From these previous studies and the current work, it appears that the responses of birds to heterospecific song are diverse and cannot be generalised to a single common pattern.
Conclusions
Two species of Turtur wood doves living in sympatry, where they do not exhibit interspecific territoriality (at least as adults with established territories), responded strongly to songs of their own species but did not approach the song of congeners and control songs. In allopatry, instead, tympanistria males 'misdirected' their response to the congeneric dove song, even though they had not previously had any earlier contact with the species. Their approach response to the playback of bremeri song was similar to that of their own species, while they did not approach control songs. We suggest that this is not necessarily a totally maladaptive behaviour as, from an evolutionary perspective, such an ability could be useful for resolving competing interests with congeners. It is more adaptive to be plastic in the response to a potential competitor than to respond or not respond in a fixed way.
Data availability
The datasets analysed during the current study are available from the corresponding author on reasonable request (Tomasz S. Osiejuk, email: osiejuk@amu.edu.pl).
References
Bradbury, J. W. & Vehrencamp, S. L. Principles of animal communication, 2nd ed. (Sinauer Press, 2011).
Catchpole, C. J. & Slater, P. J. B. Bird song: Biological themes and variations (Cambridge University Press, 2008).
Schmidt, R., Kunc, H. P., Amrhein, V. & Naguib, M. Aggressive responses to broadband trills are related to subsequent pairing success in nightingales. Behav. Ecol. 19, 635–641 (2008).
Kroodsma, D. E. & Byers, B. E. The function(s) of bird song. Am. Zool. 31, 318–328 (1991).
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. Female eavesdropping on male song contests in songbirds. Science 296, 873–873 (2002).
Dalziell, A. H., Welbergen, J. A. & Magrath, R. D. Male superb lyrebirds mimic functionally distinct heterospecific vocalizations during different modes of sexual display. Anim. Behav. 188, 181–196 (2022).
Wang, D. et al. Machine learning reveals cryptic dialects that explain mate choice in a songbird. Nat. Commun. 13, 1630 (2022).
Temeles, E. J. The role of neighbours in territorial systems: when are they ‘dear enemies’?. Anim. Behav. 47, 339–350 (1994).
Davies, N. B. & Halliday, T. R. Deep croaks and fighting assessment in toads Bufo bufo. Nature 274, 683–685 (1978).
Logue, D. M. et al. Does signalling mitigate the cost of agonistic interactions? A test in a cricket that has lost its song. Proc. R. Soc. B. 277, 2571–2575 (2010).
Parker, G. A. Assessment strategy and the evolution of fighting behaviour. J. Theor. Biol. 47, 223–243 (1974).
Emlen, S. T. & Oring, L. W. Ecology, sexual selection, and the evolution of mating systems. Science 197, 215–223 (1977).
Maher, C. R. & Lott, D. F. A review of ecological determinants of territoriality within vertebrate species. Am. Midl. Nat. 143, 1–29 (2000).
Maher, C. R. & Lott, D. F. Definitions of territoriality used in the study of variation in vertebrate spacing systems. Anim. Behav. 49, 1581–1597 (1995).
Orians, G. H. & Willson, M. F. Interspecific territories of birds. Ecology 45, 736–745 (1964).
Davies, N. B., Krebs, J. R. & West, S. A. An introduction to behavioural ecology (Wiley-Blackwell, 2012).
Dugatkin, L. A. Antipredation Behavior. in Encyclopedia of Ecology 218–221 (Elsevier, 2008). https://doi.org/10.1016/B978-008045405-4.00004-5.
Drury, J. P., Cowen, M. C. & Grether, G. F. Competition and hybridization drive interspecific territoriality in birds. Proc. Natl. Acad. Sci. U.S.A. 117, 12923–12930 (2020).
Losin, N., Drury, J. P., Peiman, K. S., Storch, C. & Grether, G. F. The ecological and evolutionary stability of interspecific territoriality. Ecol. Lett. 19, 260–267 (2016).
Cowen, M. C., Drury, J. P. & Grether, G. F. Multiple routes to interspecific territoriality in sister species of North American perching birds. Evolution 74, 2134–2148 (2020).
Cody, M. L. Character convergence. Annu. Rev. Ecol. Syst. 4, 189–211 (1973).
Cody, M. L. Convergent characteristics in sympatric species: A possible relation to interspecific competition and aggression. The Condor 71, 223–239 (1969).
Grether, G. F., Losin, N., Anderson, C. N. & Okamoto, K. The role of interspecific interference competition in character displacement and the evolution of competitor recognition. Biol. Rev. 84, 617–635 (2009).
MacArthur, R. H. Geographical ecology: patterns in the distribution of species (Princeton University Press, 1984).
Drury, J. P., Okamoto, K. W., Anderson, C. N. & Grether, G. F. Reproductive interference explains persistence of aggression between species. Proc. R. Soc. B. 282, 20142256 (2015).
Payne, R. B. Behaviour and songs of hybrid parasitic finches. Auk 97, 118–134 (1980).
Murray, B. G. The ecological consequences of interspecific territorial behavior in birds. Ecology 52, 414–423 (1971).
Baptista, L. F., Trail, P. W., Horblit, H. M. & Boesman, P. F. D. Blue-headed Wood-Dove (Turtur brehmeri). in Birds of the World (eds. Billerman, S. M., Keeney, B. K., Rodewald, P. G. & Schulenberg, T. S.) (Cornell Lab of Ornithology, 2020). https://doi.org/10.2173/bow.bhwdov1.01.
Kirwan, G. M., Baptista, L. F., Trail, P. W., Horblit, H. M. & Boesman, P. F. D. Tambourine Dove (Turtur tympanistria). in Birds of the World (eds. Billerman, S. M., Keeney, B. K., Rodewald, P. G. & Schulenberg, T. S.) (Cornell Lab of Ornithology, 2021). https://doi.org/10.2173/bow.tamdov1.02.
Appiah-Opoku, S. Using protected areas as a tool for biodiversity conservation and ecotourism: A case study of Kakum National Park in Ghana. Soc. Nat. Resour. 24, 500–510 (2011).
Tsai, Y. H. et al. Monitoring forest cover change within different reserve types in southern Ghana. Environ. Monit. Assess 191, 281 (2019).
Isbell, L. A. Re-evaluating the ecological constraints model with red colobus monkeys (Procolobus rufomitratus tephrosceles). Behav 149, 493–529 (2012).
Wallis, S. J. Sexual behavior and reproduction of Cercocebus albigena johnstonii in Kibale forest Western Uganda. Int. J. Primatol. 4, 153–166 (1983).
McCarthy, E. M. Handbook of avian hybrids of the world. (Oxford University Press, 2006).
Osiejuk, T. S., Żbikowski, B., Wheeldon, A. & Budka, M. Hey mister Tambourine Dove, sing a song for me: Simple but individually specific songs of Turtur tympanistria from Cameroon. Avian Res. 10, 14 (2019).
Percie du Sert, N. et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol 18, e3000410 (2020).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 67, (2015).
Hartig, F. DHARMa: Residual diagnostics for hierarchical (multi-level/mixed) regression models (2020).
Burnham, K. P. & Anderson, D. R. Model selection and multimodel inference (Springer-Verlag, 2002).
Bartoń, K. MuMIn: Multi-Model Inference. R package version 1.15.6. Retrieved from https://cran.r-project.org/web/packages/MuMIn/index.html (2016).
Hamao, S. Asymmetric response to song dialects among bird populations: The effect of sympatric related species. Anim. Behav. 119, 143–150 (2016).
Hamao, S. Effect of sympatry on discrimination of heterospecific song by varied tits. Anim. Behav. 180, 307–314 (2021).
Reed, T. M. Interspecific territoriality in the chaffinch and great tit on islands and the mainland of Scotland: Playback and removal experiments. Anim. Behav. 30, 171–181 (1982).
Lade, B. I. & Thorpe, W. H. Dove songs as innately coded patterns of specific behaviour. Nature 202, 366–368 (1964).
Nottebohm, F. & Nottebohm, M. E. Vocalizations and breeding behaviour of surgically deafened ring doves (Streptopelia risoria). Anim. Behav. 19, 313–327 (1971).
Janik, V. M. & Slater, P. J. B. The different roles of social learning in vocal communication. Anim. Behav. 60, 1–11 (2000).
Hagmann, C. E. & Cook, R. G. Discrimination of dynamic change and constancy over time by pigeons. Psychon. Bull. Rev. 18, 697–704 (2011).
ten Cate, C., Spierings, M., Hubert, J. & Honing, H. Can birds perceive rhythmic patterns? A review and experiments on a songbird and a parrot species. Front. Psychol. 7, (2016).
Beckers, G. J. L. & Ten Cate, C. Perceptual relevance of species-specific differences in acoustic signal structure in Streptopelia doves. Anim. Behav. 62, 511–518 (2001).
de Kort, S. R. & ten Cate, C. Repeated decrease in vocal repertoire size in Streptopelia doves. Anim. Behav. 67, 549–557 (2004).
Slabbekoorn, H. & Cate, C. T. Comparative analysis of perch-coo vocalizations in Streptopelia Doves. Auk 116, 737–748 (1999).
de Kort, S.R., den Hartog, P. & ten Cate, C. Vocal signals, isolation and hybridization in the vinaceous dove (Streptopelia vinacea) and the ring-necked dove (S. capicola). Behav. Ecol. Sociobiol. 51, 378–385 (2002).
Terpstra, N. J., Bolhuis, J. J., Riebel, K., van der Burg, J. M. M. & den Boer-Visser, A. M. Localized brain activation specific to auditory memory in a female songbird. J. Comp. Neurol. 494, 784–791 (2006).
Terpstra, N. J., Bolhuis, J. J., den Boer-Visser, A. M. & ten Cate, C. Neuronal activation related to auditory perception in the brain of a non-songbird, the ring dove. J. Comp. Neurol. 488, 342–351 (2005).
Beckers, G. J. L., Suthers, R. A. & Cate, C. ten. Mechanisms of frequency and amplitude modulation in ring dove song. J. Exp. Biol. 206, 1833–1843 (2003).
Lowther, P. E. Alder Flycatcher (Empidonax alnorum). in Birds of the World (eds. Billerman, S. M., Keeney, B. K., Rodewald, P. G. & Schulenberg, T. S.) (Cornell Lab of Ornithology, 2020). https://doi.org/10.2173/bow.aldfly.01.
Sedgwick, J. A. Willow Flycatcher (Empidonax traillii). in Birds of the World (eds. Billerman, S. M., Keeney, B. K., Rodewald, P. G. & Schulenberg, T. S.) (Cornell Lab of Ornithology, 2020). https://doi.org/10.2173/bow.wilfly.01.
Barlow, J. C. & McGillivray, W. B. Foraging and habitat relationships of the sibling species Willow Flycatcher (Empidonax traillii) and Alder Flycatcher (E. alnorum ) in southern Ontario. Can. J. Zool. 61, 1510–1516 (1983).
Prescott, D. R. C. Territorial responses to song playback in allopatric and sympatric populations of Alder (Empidonax alnorum) and Willow (E. traillii) flycatchers. Wilson Bull. 99, 611–619 (1987).
Jedlikowski, J., Polak, M. & Ręk, P. Dear-enemy effect between two sympatric bird species. Anim. Behav. 184, 19–26 (2022).
de Kort, S. R., den Hartog, P. M. & ten Cate, C. Diverge or merge? The effect of sympatric occurrence on the territorial vocalizations of the vinaceous dove Streptopelia vinacea and the ring-necked dove S. capicola. J. Avian Biol. 33, 150–158 (2002).
Stutchbury, B. J. M. & Morton, E. S. Behavioral ecology of tropical birds. (Academic Pres, 2022).
de Kort, S., den Hartog, P. & ten Cate, C. Vocal signals, isolation and hybridization in the vinaceous dove (Streptopelia vinacea) and the ring-necked dove (S. capicola). Behav. Ecol. Sociobiol. 51, 378–385 (2002).
van den Bosch, F., Hengeveld, R. & Metz, J. A. J. Analysing the velocity of animal range expansion. J. Biogeogr. 19, 135 (1992).
Vokurková, J., Motombi, F. N., Ferenc, M., Hořák, D. & Sedláček, O. Seasonality of vocal activity of a bird community in an Afrotropical lowland rain forest. J. Trop. Ecol. 34, 53–64 (2018).
McEntee, J. P. Reciprocal territorial responses of parapatric African sunbirds: Species-level asymmetry and intraspecific geographic variation. Behav. Ecol. 25, 1380–1394 (2014).
Sandoval, L., Méndez, C. & Mennill, D. J. Different vocal signals, but not prior experience, influence heterospecific from conspecific discrimination. Anim. Behav. 85, 907–915 (2013).
Acknowledgements
We greatly appreciate Samuel Kojo Annan-Riverson (Manager of Kakum NP) and Innocent Kato (Makerere University Biological Field Station) for logistic help and organisation of the fieldwork, and dr. Yaw Boakye Agyemang for help in organising permits for research. Park rangers, Jonas Bannor and Emmanuel Kwakye (in Kakum NP) and Swaibu Katusabe, Perez Rukiri and Martin Mukasa (in Kibale NP) were our field assistants and made our work safe. We thank Magda Herdegen-Radwan for RADseq library analyses, which will be a subject of a separate study, but support knowledge about lack of hybridization of the studied species. We are grateful to K. Lindsay Higgins for professional language editing. The study was funded by the Polish National Science Centre’s (grant no. 2018/31/B/NZ8/00482 to T.S.O.).
Author information
Authors and Affiliations
Contributions
T.S.O. conceived this project; M.N., P.S., M.B. and T.S.O. collected data and performed experiments. M.N. and T.S.O. performed the bioacoustics and statistical analyses; P.S. was responsible for collecting all permits and contact with local authorities; M.N. and T.S.O. drafted the manuscript and all authors contributed to the editing of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Niśkiewicz, M., Szymański, P., Budka, M. et al. Response of forest Turtur doves to conspecific and congeneric songs in sympatry and allopatry. Sci Rep 13, 15948 (2023). https://doi.org/10.1038/s41598-023-43035-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-43035-8
This article is cited by
-
Neighbour–stranger discrimination in an African wood dove inhabiting equatorial rainforest
Scientific Reports (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
