
Abstract
Dietary information from aquatic organisms is instrumental in predicting biological interactions and understanding ecosystem functionality. In freshwater habitats, generalist fish species can access a diverse array of food sources from multiple food chains. These may include primary photosynthetic production and detritus derived from both oxic and anoxic decomposition. However, the exploitation of anoxic decomposition products by fish remains insufficiently explored. This study examines feeding habits of striped catfish (Pangasianodon hypophthalmus) at both adult and juvenile stages within a tropical reservoir, using stable carbon, nitrogen, and sulfur isotope ratios (δ13C, δ15N, and δ34S, respectively) and fatty acid (FA) analyses. The adult catfish exhibited higher δ15N values compared to primary consumers that feed on primary photosynthetic producers, which suggests ingestion of food sources originating from primary photosynthetic production-based food chains. On the other hand, juvenile catfish demonstrated lower δ15N values than primary consumers, correlating with low δ34S value and large proportions of bacterial FA but contained small proportions of polyunsaturated FA. This implies that juveniles utilize food sources from both anoxic decomposition and primary photosynthetic production-based food chains. Our results indicate that food chains based on anoxic decomposition can indeed contribute to the dietary sources of tropical fish species.
Similar content being viewed by others

Introduction
Understanding the feeding habits of aquatic animals is crucial for predicting biological interactions and comprehending ecosystem function. Freshwater ecosystems, characterized by diverse habitats and species, rely on multiple food chains that sustain complex food webs1,2. Algal primary production and microbial decomposition of autochthonous and allochthonous organic sources, such as detritus, serve as essential energy sources for food webs in freshwater ecosystems3. Furthermore, organic matter derived from autochthonous and allochthonous origins act as a substrate for anoxic decompositions by microorganisms, including methanogens4 and sulfate-reducing bacteria (SRB)5,6. The products of anoxic decomposition, such as methane and sulfide, can be transferred to higher trophic levels through chemoautotrophic microorganisms7,8. Generalist fish species often exploits a wide range of food sources, including phytoplankton and detritus9, but the contribution of anoxic decomposition products to fishes remains poorly understood.
Stable isotopes in animal tissues are widely used to determine the feeding habits and organic sources of aquatic organisms including generalist fish species. Stable carbon and nitrogen isotope ratios (δ13C and δ15N values, respectively) are particularly powerful tools for determining spatial/trophic niches and energy flows in the food webs of freshwater ecosystems10,11. δ13C value in consumer tissues discriminates between food webs based on planktonic (phytoplankton; δ13C = − 32 ± 2‰) or benthic algae (periphyton; δ13C = − 27 ± 3‰)12. On the other hand, δ15N value is used to identify trophic positions in a food web13,14,15. However, this conventional method may limit to define detritus-based food chains regardless of oxic and anoxic decomposition, considering that δ13C values of sedimentary detritus overlap among those of phytoplankton and terrestrial organic matters16. Fatty acid (FA) analysis is used to define the contribution of detritus and primary photosynthetic producers to aquatic consumers17,18 because the FA compositions of bacteria is distinct from that of phytoplankton19,20,21, and the FA composition in consumer tissues reflects that in their diet22,23. Furthermore, the stable sulfur isotope ratio (δ34S value) can be used to distinguish the origin of organic matter under oxic and anoxic conditions24. A difference in δ34S value likely results from a low δ34S value caused by SRB activity under anoxic conditions (ca. − 20‰)25,26, relative to δ34S values of sulfate and organisms from oxygenated conditions (0 to + 10‰ for freshwater)27.
The striped catfish (Pangasianodon hypophthalmus), an endemic species to the Mekong and Chao Phraya rivers in Southeast Asia, is a large freshwater fish that can grow up to 130 cm in body length28. Due to its popularity aquaculture, the catfish has been introduced into tropical inland waters in Southeast Asia28, South Asia29, and South America30. Often, released fish settle in the natural environments of these areas. Numerous studies have investigated aquaculture and physiological aspects of striped catfish using fingerlings31,32. In the Mekong River, the catfish can be found in various habitats, including mainstreams, floodplain, and lakes33. Annual migration patterns likely differ between juvenile and adult striped catfish33, potentially leading to variations in food sources across different growth stages. The catfish are considered generalists, feeding omnivorously on algae and detritus, owing to their relative gut length34. Furthermore, stomach content analysis has revealed the presence of phytoplankton, detritus, and zooplankton in their diets35. However, their trophic positions within food webs remain unclear.
This study aims to investigate the feeding habits of juvenile and adult striped catfish inhabiting the Kaeng Krachan Reservoir, Thailand (Fig. 1). In the reservoir, hatchery-reared striped catfish measuring 10 cm or less in total length (TL) has been released for fishery stock enhancement, and these fish have grown to reach 100 cm in TL. Due to fishery stock management practices, catfish reaching 30 cm or more in TL have been captured in the reservoir. Consequently, our investigation focused on juvenile (36–50 cm in TL) and adult (80–106 cm in TL). To assess the contribution of primary photosynthetic production and sedimentary detritus-based food chains, including oxic and anoxic decomposition, to their food sources, we analyzed three isotope ratios (δ13C, δ15N, and δ34S) and FA biomarkers in the tissues of juvenile and adult catfish. Additionally, we inferred the trophic positions of striped catfish within primary photosynthetic production-based food chains by comparing their δ13C and δ15N values with those of primary consumers that feed on primary photosynthetic producers and other sympatric fish species with well-documented feeding habits. Furthermore, we examined δ34S values in sulfate collected from water and sulfide obtained from sediment to establish the baselines for oxic and anoxic decomposition in the reservoir.

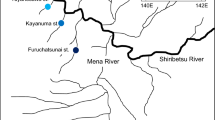
Map of the Kaeng Krachan Reservoir, Thailand. Closed circles represent the sampling sites. Bottom sediments were collected at sites 1–7 and 9–11. Water samples were collected at sites 1–8. Physico-chemical profiles were measured at sites 1–6.
Results
Aqueous and sediment geochemistry
Water temperature (WT), chlorophyll a fluorescence, and dissolved oxygen (DO) were measured in the Kaeng Krachan Reservoir in September 2019. In the reservoir, WT and chlorophyll a fluorescence declined with increasing depth from the surface at all sampling sites. A thermocline was observed at approximately water depths of 5–10 m (sites 1–6; Figs. 1 and S1). We also observed the boundaries of DO concentrations at water depths of 5–10 m. The DO concentration was higher in the surface zone than in the profundal zone, but increased again near the reservoir bottom at all sites except for site 5 (Fig. S1). In September 2019, we also measured the sulfate ion concentration and δ34S values of sulfate ions (δ34Ssulfate) in water. The sulfate concentration was 8.4–53.8 µM and declined with increasing depth from the surface at all sampling sites. The δ34Ssulfate value was + 9.0‰ to + 12.3‰ (Fig. S2). Sediment samples were collected in September 2019 to analyze the δ34S values of acid-volatile sulfide (AVS) and total sulfur (TS) (δ34SAVS and δ34STS, respectively). TS and AVS were detected in sediments on the sites below water depths of 15.5 m (sites 2–6; Table S1). At all sites, δ34SAVS values (range + 6.3‰ to + 9.6‰) were intermediate between δ34Ssulfate and δ34STS values (range + 6.0‰ to + 8.4‰) (Table S1).
Stable isotope signatures of food web components in the Kaeng Krachan Reservoir
In the reservoir, we collected 25 juvenile striped catfish in August–September and November–December 2019, as well as 11 adult striped catfish in November–December 2019. The adult striped catfish exhibited higher values for three isotope signatures (δ13C = − 24.8 ± 1.4 [mean ± SD] ‰, δ15N = 9.1 ± 0.8‰, δ34S = 6.2 ± 0.9‰) compared to the juvenile catfish (δ13C = − 30.8 ± 1.1‰, δ15N = 2.2 ± 2.5‰, δ34S = 0.1 ± 2.0‰) (Table 1). We compared the δ13C and δ15N values among the catfish and other sympatric fish species collected in August–September and November–December 2019, which included the piscivore Hampala macrolepidota36, carnivore Hemibagrus nemurus37, insectivore Puntioplites proctozystron38, herbivore Hypsibarbus wetmorei39, and herbivore Oreochromis niloticus40 (cited from Medo et al.41). The Steel–Dwass test showed that the δ13C value of adult catfish did not differ from that of H. nemurus (δ13C = − 26.7 ± 1.4‰, p = 0.05) and H. wetmorei (δ13C = − 22.6 ± 3.0‰, p = 0.63), but was significantly higher than that of H. macrolepidota (δ13C = − 27.2 ± 0.5‰, p = 0.01), P. proctozystron (δ13C = − 27.3 ± 1.0‰, p = 0.02), and O. niloticus (δ13C = − 29.8 ± 0.6‰, p = 0.002). The Steel–Dwass test showed that the δ13C value of juvenile catfish was significantly lower than that of H. macrolepidota (p = 1.0e−4), H. nemurus (p = 4.5e−6), P. proctozystron (p = 1.0e−4), and H. wetmorei (p = 4.8e−5), but did not differ from that of O. niloticus (p = 0.22). The Steel–Dwass test showed that the δ15N value of adult catfish did not differ from that of P. proctozystron (δ15N = 9.3 ± 0.6‰, p = 1.00), H. wetmorei (δ15N = 10.5 ± 1.2‰, p = 0.18), and O. niloticus (δ15N = 9.1 ± 0.5‰, p = 1.00), but was significantly lower than that of H. macrolepidota (δ15N = 11.5 ± 0.5‰, p = 0.002) and H. nemurus (δ15N = 12.5 ± 0.6‰, p = 4.0e−4). The Steel–Dwass test showed that the δ15N value of juvenile catfish was significantly lower than that of H. macrolepidota (p = 1.0e−4), H. nemurus (p = 3.4e−6), P. proctozystron (p = 1.0e−4), H. wetmorei (p = 1.0e−4), and O. niloticus (p = 4.8e−5). We compared the δ15N values between juvenile catfish and primary consumers that feed on primary photosynthetic producers, Trichoptera (n = 14), collected in September and November 2019 (cited from Medo et al.41). The δ15N value of juvenile catfish was lower compared to Trichoptera (δ15N = 4.6 ± 0.6‰) (Table 1; Fig. 2). Furthermore, we compared the δ15N values among juvenile catfish and basal food sources, including periphyton (n = 26), particulate organic matter (POM; n = 48), and lake bottom sediments (n = 16), collected in September and November 2019 (cited from Medo et al.41). The δ15N value of juvenile catfish was lower than that of the basal sources (δ15N = 3.3 ± 1.4‰, 4.4 ± 1.2‰, and 5.6 ± 0.6‰ for POM, periphyton, and sediments) (Table 1; Fig. 2). We did not observe any horizontal and vertical variations in the δ13C and δ15N values in POM and sediments (Fig. S3), nor did we detect large seasonal variations in the δ13C and δ15N values in the food web components between August–September and November–December 2019 (Fig. S4).

Stable carbon and nitrogen isotope values (δ13C and δ15N) of all samples collected from the Kaeng Krachan Reservoir. Points and error bars indicate the mean and standard deviation of stable isotope ratios. POM indicates particulate organic matter. All data except for the striped catfish (Pangasianodon hypophthalmus) were cited from Medo et al., 202141.
Fatty acid compositions of the striped catfish
The most abundant FA was saturated FA (SAFA) in adult and juvenile catfish (44.8 ± 2.9% and 45.8 ± 7.4%, respectively; Table 2). The second dominating FA in the adult and juvenile catfish was monounsaturated FA (MUFA) (31.0 ± 5.1% and 35.1 ± 9.6%, respectively). Bacterial FA (BAFA) and MUFA were significantly higher in juvenile catfish than that in adult catfish (Wilcoxon rank-sum test, p = 1.2e−7 and 0.04, respectively) (Fig. 3). The abundance of i- and a- branched FAs, as well as methane-oxidizing bacteria (MOB) specific FAs19, was less than 1% of all FAs (Table 1). Polyunsaturated FA (PUFA) was significantly higher in adult catfish than in juvenile catfish (Wilcoxon rank-sum test, p = 0.001) (Fig. 3).

Composition of fatty acid (FA) groups (% relative to total FA) in adult and juvenile in the striped catfish (Pangasianodon hypophthalus). Asterisk indicates a significantly difference at **p < 0.001; *p < 0.05.
Relationships among stable isotope signatures and fatty acid compositions
There was a significant correlation between δ15N and δ34S in adult catfish (Pearson's correlation coefficient [r] = 0.80, p = 0.003). Likewise, there were significant correlations between δ15N and δ34S (r = 0.91, p = 2.0e−10), and δ15N and BAFA (r = − 0.82, p = 4.5e−7), for juvenile catfish (Fig. 4).

Correlations among three stable isotope ratios (carbon [δ13C], nitrogen [δ15N], and sulfur [δ34S]), bacterial fatty acid (BAFA), and total length (cm) in the striped catfish (Pangasianodon hypophthalmus). Solid lines indicate significant correlations at Pearson's correlation coefficient (p < 0.05).
Discussion
This study indicates that the striped catfish exploited food sources across multiple food chains, ranging from anoxic decomposition by bacteria to primary photosynthetic production by algae. The stable isotope signatures and FA compositions were distinct between juvenile and adult catfish. Our δ13C and δ15N analyses indicated that the primary producers contributed to the food sources for adult catfish but not entirely for juvenile catfish. The lower δ15N values relative to those of primary consumers in juvenile catfish were associated with low δ34S values and high proportions of BAFA, indicating the contribution of anoxic decomposition-based food chains to their diet. Although striped catfish were previously thought to feed omnivorously on phytoplankton, detritus, and zooplankton32, this study suggests that the feeding habits of catfish differ between adults and juveniles.
Since the δ15N value of adult catfish did not significantly differ from that of the herbivore O. niloticus40 and insectivore P. proctozystron38, it is likely that adult catfish were primary to secondary consumers in primary photosynthetic production-based food chains. The FA composition of adult catfish was characterized by high proportions of PUFA, which are synthesized by phytoplankton such as diatoms, dinoflagellates, and cryptophytes, in freshwater ecosystems22,23. In addition to algal sources, long-chain PUFA can also be obtained from freshwater zooplankton owing to their selective accumulation42. Thus, adult catfish may ingest zooplankton as well as algae.
Low δ15N values in juvenile catfish suggests that they exploited food sources from anoxic decomposition-based food chains. Given a trophic discrimination factor of ca. + 3‰ between a prey item and consumer43, the δ15N values of consumers on primary photosynthetic production-based food chains should be higher than those of primary producers, such as periphyton and POM (δ15N = ca. + 4‰). Although primary consumers that feed on periphyton and phytoplankton can serve as an isotopic baseline for primary photosynthetic production-based food chains10, the δ15N values of primary consumers, Trichoptera, did not cover those observed in juvenile catfish. The decrease in δ15N values with increasing BAFA proportions and decreasing δ34S values in juvenile catfish indicates that the low δ15N values were caused by bacterial activity under anoxic conditions. Extremely low δ15N values have been observed in methanotrophic and/or chemoautotrophic communities in freshwater44,45, caves46, and the deep sea47, which likely results from extensive nitrogen isotope fractionation during the assimilation of dissolved inorganic nitrogen (i.e., ammonium and nitrate) by bacteria under anoxic conditions. However, despite the high proportion of BAFA, juvenile catfish also contained PUFA that are typically absent in bacteria19,48. Therefore, it is likely that juvenile catfish also exploited food sources from primary photosynthetic production-based food chains.
The δ13C values of juvenile catfish were the lowest among fish but were similar values to those of primary consumers, Trichoptera. Furthermore, specific FAs to MOB, such as 16:1n-8 and 18:1n-819, were not detected in the tissues of juvenile catfish. These findings suggest limited contributions of methanogens to the diet of juvenile catfish. In freshwater ecosystems, phytoplankton exhibits δ13C values in the range of − 35 to − 25‰49, which was consistent with our results of POM. Because biogenic methane shows extremely low δ13C values (δ13Cmethane = − 110 to − 50‰)50, it is thought that low δ13C values in consumer tissues are evidence that consumers incorporate methane-derived carbon via methane-oxidizing bacteria4,7,8. Alternatively, chemosynthetic bacteria such as sulfur- and ammonia-oxidizing bacteria can also be a low δ13C source (δ13C > − 40‰; e.g.,51). In contrast, Sanseverino et al.52 reported that MOB-specific FAs were found in freshwater fishes despite a relatively high δ13C value in tissues (δ13C = − 37 to –28‰), which suggests consumers that obtained the compounds from anoxic conditions do not necessarily present extremely low δ13C values if they obtained the compounds from anoxic conditions. It is possible that we overlooked the bacterial contributions to high trophic levels using bulk δ13C analysis alone.
According to previous reports, ingestions of anoxic decomposition-derived sources, evidenced by low δ13C values, could be associated with fishes (i) feeding on detritus52 and/or (ii) feeding on bacterivorous organisms such as zooplankton and chironomid larvae53,54. Moreover, BAFAs have been found in detritivores55 and benthivores18. Given the results of gut morphometry for adult and juvenile striped catfish34, it seems that the main food sources for striped catfish in both growth stages was detritus rather than animal sources. These findings indicate that juvenile catfish ingested detritus at the bottom of the reservoir to obtain anoxic decomposition-derived sources. Exploiting such organic sources has been observed in chironomid larvae and fish with high hypoxia-tolerance4, which is consistent with the physiological traits of striped catfish, known for their elevated hypoxia tolerance56.
Ontogenetic dietary shifts are widely observed in fish species and are probably driven by several factors, such as habitat use, morphological constraints, and swimming ability57. Our results on the differences of stable isotopes and FA between adult and juvenile catfish can be linked to habitat use at each growth stage, and perhaps juvenile access to lentic systems in the reservoir. Annual migration patterns are likely different between juvenile and adult striped catfish; juveniles spend time in the nursery and feeding grounds of the floodplain, while adults migrate between feeding and spawning grounds of the mainstream33. The δ13C and δ34S values of basal food sources often vary depending on the habitats, even within a lake12,15. Low δ13C and δ34S values, which suggest that exploitations of anoxic decomposition-derived compounds by aquatic consumers, is typically observed in restricted sites, such as the profundal zones of lakes and in the vegetation area of shallow lakes and rivers7,8,24,58. Although horizontal and vertical variations in δ13C and δ34S values were slight in basal sources at the reservoir, sulfide (i.e., AVS), which is an indicator of anoxic conditions, was found at restricted sites, including lentic areas and profundal zone of the reservoir.
In this study, we did not observe low δ15N values in sediments, like those found in juvenile catfish. Our results also indicated that δ34STS extracted from the sediments did not reflect the δ34S value of the organic sources of juvenile catfish. The inconsistency in stable isotope values between deposit feeders and sediments can be explained by the selection of diet particles from sedimentary organic matter59. Although there was no direct evidence of potential organic sources for juvenile catfish, low stable isotope ratios in catfish compared to bulk sediments suggests selective assimilation of organic matter from bulk sediments.
However, it is important to acknowledge the limitations of this study. Tropical fishes in Southeast Asia exhibit seasonal dietary shifts60, which were not considered in this study due to sample collections lacking seasonality. Therefore, our dietary analyses may not fully capture the feeding habits of striped catfish. Additionally, the δ13C and δ15N values in food web components, especially in short-lived organisms, are highly variable depending on the season and month61. Due to sample size constraints, we could not account for seasonal variations between September and November. Consequently, stable isotope data from these two months were combined into a single dataset, potentially overlooking seasonal fluctuations in food web baselines and the feeding habits of catfish during that period. Finally, it is crucial to consider that trophic discrimination factors and turnover rates of stable isotopes in animal tissues can vary depending on body size and age62,63. Future analyses that encompass there parameters will offer a more comprehensive understanding of the differences in feeding habits between adult and juvenile catfish, surpassing the limitations of solely utilizing their bulk stable isotope data.
Overall, stable isotope and FA composition analyses indicated that the striped catfish ingested food sources derived from anoxic decomposition and primary photosynthetic production-based food chains during juvenile stages. Meanwhile, the striped catfish exploited food sources derived from primary photosynthetic production-based food chains during adult stages. While the feeding habits of freshwater fishes are commonly described based on primary photosynthetic production and oxic decomposition-based food chains, our research demonstrated that anoxic decomposition-based food chains also contribute to the food sources of tropical freshwater fishes. Nonetheless, it remains unclear how exploiting the food sources derived from anoxic decompositions benefits fish growth and survival. Foraging at sites where dissolved oxygen is depleted may mitigate resource competitions and provide refuge areas from predators, whereas ingesting sedimentary organic matter, including bacteria, can have negative effects on somatic growth and reproduction owing to a poor-quality food3,64. To test these hypotheses, future studies should investigate habitat use and trade-offs between food quality and quantity.
Methods
Ethics statement
All experiments and sample collection described in this paper agreed with the policies of the Animal Experiment Committee of Graduate School of Informatics, Kyoto University. All fieldwork and sample processing were conducted under the permission of the National Research Council of Thailand. All fish used in this study were caught and sold by fishermen and fish markets under the permission of the Inland Fisheries Patrol Unit, Kaeng Krachan District, Department of Fisheries, Thailand. All fish tissues used in our experiments were purchased from the fish markets, and the fish were already dead when we purchased them. We did not kill the fishes ourselves.
Study site
This study was conducted during a monomictic lake, the Kaeng Krachan Reservoir, Phetchaburi Province, western Thailand (12° 54ʹ N, 99° 36ʹ E; Fig. 1). The Kaeng Krachan Dam impounds the Phetchaburi River, which originated in Myanmar in 1966. The climate in this area can be divided into two seasons: the wet (May–October) and the dry seasons (November–April). The annual maximum depth of the reservoir is approximately 45 m during the wet season. A thermocline in the reservoir was observed at a depth of 5–15 m from April to November and an annual overturn occurred in December65. The productivity of the reservoir is classified as mesotrophic during the dry season and eutrophic during the wet season66. The Department of Fisheries, Thailand, has released hatchery-reared striped catfish < 10 cm in TL into the reservoir for fishery stock enhancement. In addition to released catfish, there is recruitment from the natural reproduction of stocked fish in the reservoir. The Department of Fisheries, Thailand imposes fishing restrictions on the striped catfish during the spawning seasons of this species, the middle of May to the middle of August. Furthermore, it is unlikely for fishermen in the reservoir to target the catfish below 30 cm in TL.
Sample collection
Sample were collected from the end of August to the beginning of September (the end of wet season) and from the end of November to the beginning of December (the beginning of dry season) in 2019. All striped catfish used in this study were caught by a local fisherman with permission from the Inland Fisheries Patrol Unit, Kaeng Krachan District, Department of Fisheries, Thailand. We used 36 individuals of the striped catfish (36–106 cm in TL). Based on the mature size of the catfish (> 54 cm in body length)28, we divided them into juveniles (n = 25; 36–50 cm in TL) and adults (n = 11; 80–106 cm in TL). Consequently, 25 juvenile catfish were collected in August–September and November–December 2019, while 11 adult catfish were collected in November–December 2019. Catfish samples were collected from the dorsal and ventral muscles for stable isotope and FA analyses, respectively. In September 2019, sediment samples were collected at a few centimeters of depth, using an Ekman-Birge grab at depths of 3–30 m at ten sites (sites 1–7 and 9–11; Fig. 1). Water samples were collected in September 2019 using a Van Dorn water sampler (water capacity, 6.0 L: Miyamoto Riken Ind. Co, Ltd., Osaka, Japan) at four depths (0, 5, 10, and 2 m above the bottom of the reservoir) and seven sites (sites 1–7; Fig. 1). All samples were stored in a freezer at − 20 °C until further processing.
Physico-chemical analysis
The WT (°C), DO (mg L−1), and chlorophyll a fluorescence (µg L−1) were measured at six sites (sites 1–6; Fig. 1) during September 2019. The data were observed using a conductivity-temperature-depth instrument (AAQ-RINKO, AAQ170; JFE-Advantech, Kobe, Japan). For ion concentration analysis, the water samples were vacuum filtered through a glass microfiber GF/F filter (0.7 µm mesh or nominal pore size; Whatman, Maidstone, UK) and then through a PTFE syringe filter (0.45 µm pore size; RephiLe Bioscience, Ltd., Boston, USA). We measured the sulfate ion concentration of the water column (µM) using an ion chromatography system (Dionex ICS-1100, Thermo Fisher Scientific, MA, USA) at Kyoto University.
Stable isotope analysis
Samples of the striped catfish were oven-dried at 50 °C for 48–72 h, then crushed by scissors. The crushed samples were delipidated using a chloroform–methanol solution (2:1 v/v). The TS in the sediment samples was converted to sulfate by heating from 80 to 120 °C in a 35.5% hydrogen peroxide solution. The AVS of the sediments was liberated by anaerobic acidification in 47% concentrated sulfuric acid during active distillation (pure nitrogen gas). The liberated H2S was collected as a CdS precipitate in traps containing 0.5 M cadmium acetate solution. The yellow CdS precipitate was oxidized with a few drops of hydrogen peroxide solution (35.5%). These resulting sulfates derived from TS and AVS were recovered as a BaSO4 precipitate by adding a 0.5 M BaCl2 solution. The resulting BaSO4 precipitate and dried sediment fraction were precisely weighted, and the TS and AVS concentration was calculated in μmol-S g−1 of dry sediment. Sulfate in water was also recovered as a BaSO4 precipitate after removing major cations using a cation-exchange resin.
δ13C and δ15N values of the striped catfish were measured using EA-IRMS (Delta V equipped with an elemental analyzer Flash EA 2000, Thermo Fisher Scientific, MA, USA) in the Center for Ecological Research, Kyoto University (CERKU). δ34S value of the striped catfish and inorganic samples (i.e., BaSO4 from the sulfate of the water column and sulfide of the sediment) were measured using an EA-IRMS (Delta V equipped with an elemental analyzer Flash EA 2000, Thermo Fisher Scientific, MA, USA at the Research Institute for Humanity and Nature [RIHN] and IsoPrime EA, GV Instruments, Manchester, UK, at the Tokyo University of Marine Science and Technology [TUMSAT], respectively). All samples for the δ34S were mixed with excess V2O5.
The δ13C, δ15N, and δ34S (in ‰) were expressed as the deviation from standards according to the following equation:
where X is 13C, 15N, or 34S; Rsample is the ratio (13C/12C, 15N/14N or 34S/32S) in the sample; and Rstandard is the ratio in the standard. The standard reference materials were PDB carbonate, atmospheric N2, and Vienna Canyon Diablo Troilite (VCDT) for the carbon, nitrogen, and sulfur samples, respectively. We calibrated the δ13C and δ15N values using three working standards (CERKU-02, 03, and 0567) and calibrated the δ34S value using five working standards (IAEA-S-1, IAEA-S-2, and IAEA-S-3 at RIHN, and MSS-2 and MSS-368 at TUMSAT). The overall analytical precision (standard deviation, SD) for our analyses based on working standards was within ± 0.1 and ± 0.2‰ for δ13C and δ15N in CERKU, respectively, and within ± 0.3‰ for δ34S in RIHN and TUMSAT.
Fatty acid analysis
All catfish muscle samples were freeze-dried for 24–48 h and crushed using scissors. We used 100 mg of crushed sample (dry mass) for lipid extraction using the Bligh-Dyer method. We then conducted saponification and methylation of the free lipids using HCl–methanol (one-step method). The FAME composition was quantitatively analyzed using gas chromatography (GC) coupled with a flame ionization detector (FID; GC-14B, Shimadzu Scientific Instruments, Kyoto, Japan). The structures of the individual FAMEs were separately identified by using GC with a mass spectrometer (MS; GCMS-QP5050, Shimadzu Scientific Instruments, Kyoto, Japan) and scanning electron impact mass spectra. Each piece of equipment had an Omegawax capillary column (30 m, 0.25 mm, i.d., 0.25-µm film thickness, Supelco, USA). The following temperature gradient and instrument settings were used: the GC columns initial temperature was 140 °C; this was held for 2.5 min then ramped at 4 °C/min to 240 °C and then kept for 15 min, giving a total runtime of 42.5 min. Nitrogen and helium gases were used as carriers for FID and MS, respectively. Individual FAMEs were identified by comparing their retention times with those of authentic standard mixtures, Supelco FAME mix 37 components, Supelco Bacterial Acid Methyl Ester Mix (Sigma-Aldrich, St. Louis, MO, US), and with spectrographic patterns from the mass spectral library (NIST107.LIB and NIST21.LIB). The proportion of fatty acids (%) was determined as the ratio of each FAME peak area to the total areas of all the peaks in the FID analysis. The FAs were abbreviated following the standard nomenclature X:Yn-Z, where X is the number of carbons, Y is the number of double bonds, and Z is the position of the first double bond counted from the end of the methyl group in the FA (e.g. 20:5n-3). We summarized the FA groups including the sum of SAFA, MUFA, PUFA, and BAFA. Here, BAFA included 15:0, 17:0, iso-, and anteiso- (i- and a-) branched FAs (i.e., i-15:0, a-15:0, i-17:0, and a-17:0), 16:1n-7, and 18:1n-7, as these FAs are widely observed in bacteria69,70.
Statistical analysis
We used δ13C and δ15N values that were measured from several organisms in our previous study41; five fish species including piscivore Hampala macrolepidota36, carnivore Hemibagrus nemurus37, insectivore Puntioplites proctozystron38, herbivore Hypsibarbus wetmorei39, and herbivore Oreochromis niloticus40; other organisms and substrates were periphyton (n = 26), POM (n = 48), lake bottom sediments (n = 16), chironomids larvae (n = 24), aquatic insects (including Trichoptera, Anisoptera larvae, and Cymothoidae, n = 41), and shrimp (n = 7) (Table 1). We considered that POM represented the δ13C and δ15N values of phytoplankton in the reservoir. The POM was collected by vacuum filtering water samples through a glass microfiber GF/F filter (0.7 µm mesh or nominal pore size; Whatman, Maidstone, UK) after the water samples were collected using a Van Dorn water sampler (water capacity, 6.0 L: Miyamoto Riken Ind. Co, Ltd., Osaka, Japan)41. We collected the aquatic insects hiding in periphyton. All samples were collected during August–September and November–December 2019.
All statistical analyses were performed using the R software ver. 4.0.171 at a statistical significance level of 0.05. Using Steel–Dwass test (pSDCFlig function in NSM3 package72), we compared δ13C and δ15N values among fish species (i.e., the juvenile and adult catfish and other fishes). We investigated the correlations in δ13C and δ15N between three factors: (i) δ34S, (ii) BAFA, and (iii) TL using Pearson's correlation analyses (cor.test function).
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Thompson, R. M., Dunne, J. A. & Woodward, G. Freshwater food webs: towards a more fundamental understanding of biodiversity and community dynamics. Freshw. Biol. 57, 1329–1341 (2012).
Borrelli, J. J. & Relyea, R. A. A review of spatial structure of freshwater food webs: Issues and opportunities modeling within-lake meta-ecosystems. Limnol. Oceanogr. 67, 1746–1759 (2022).
Brett, M. T., Kainz, M. J., Taipale, S. J. & Seshan, H. Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc. Natl. Acad. Sci. 106, 21197–21201 (2009).
Jones, R. I. & Grey, J. Biogenic methane in freshwater food webs. Freshw. Biol. 56, 213–229 (2011).
Holmer, M. & Storkholm, P. Sulphate reduction and sulphur cycling in lake sediments: A review. Freshw. Biol. 46, 431–451 (2001).
Zak, D. et al. Sulphate in freshwater ecosystems: A review of sources, biogeochemical cycles, ecotoxicological effects and bioremediation. Earth-Sci. Rev. 212, 103446 (2021).
Trimmer, M. et al. River bed carbon and nitrogen cycling: State of play and some new directions. Sci. Total Environ. 434, 143–158 (2012).
Grey, J. The incredible lightness of being methane-fuelled: Stable isotopes reveal alternative energy pathways in aquatic ecosystems and beyond. Front. Ecol. Evol. 4, 8. https://doi.org/10.3389/fevo.2016.00008 (2016).
Higgins, K. A., Vanni, M. J. & González, M. J. Detritivory and the stoichiometry of nutrient cycling by a dominant fish species in lakes of varying productivity. Oikos 114, 419–430 (2006).
Post, D. M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 83, 703–718 (2002).
Layman, C. A. & Allgeier, J. E. Characterizing trophic ecology of generalist consumers: A case study of the invasive lionfish in The Bahamas. Mar. Ecol. Prog. Ser. 448, 131–141 (2012).
France, R. L. Differentiation between littoral and pelagic food webs in lakes using stable carbon isotopes. Limnol. Oceanogr. 40, 1310–1313 (1995).
Deniro, M. J. & Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 45, 341–351 (1981).
Minagawa, M. & Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 48, 1135–1140 (1984).
Peterson, B. J. & Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 18, 293–320 (1987).
Del Giorgio, P. A. & France, R. L. Ecosystem-specific patterns in the relationship between zooplankton and POM or microplankton del13C. Limnol. Oceanogr. 41, 359–365 (1996).
Strandberg, U. et al. Selective transfer of polyunsaturated fatty acids from phytoplankton to planktivorous fish in large boreal lakes. Sci. Total Environ. 536, 858–865 (2015).
Chaguaceda, F., Eklöv, P. & Scharnweber, K. Regulation of fatty acid composition related to ontogenetic changes and niche differentiation of a common aquatic consumer. Oecologia 193, 325–336 (2020).
Taipale, S. J., Brett, M. T., Pulkkinen, K. & Kainz, M. J. The influence of bacteria-dominated diets on Daphnia magna somatic growth, reproduction, and lipid composition. FEMS Microbiol. Ecol. 82, 50–62 (2012).
Taipale, S. et al. Fatty acid composition as biomarkers of freshwater microalgae: Analysis of 37 strains of microalgae in 22 genera and in seven classes. Aquat. Microb. Ecol. 71, 165–178 (2013).
Galloway, A. W. E. et al. A fatty acid based Bayesian approach for inferring diet in aquatic consumers. PLoS One 10, e0129723. https://doi.org/10.1371/journal.pone.0129723 (2015).
Iverson, S. J. Tracing aquatic food webs using fatty acids: from qualitative indicators to quantitative determination. in Lipids in Aquatic Ecosystems (ed. Arts, M. T., Brett, M. T., & Kainz, M. J.) 281–308 (Springer New York, 2009).
Tocher, D. R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 11, 107–184 (2003).
Croisetière, L., Hare, L., Tessier, A. & Cabana, G. Sulphur stable isotopes can distinguish trophic dependence on sediments and plankton in boreal lakes. Freshw. Biol. 54, 1006–1015 (2009).
Kaplan, I. R. & Rittenberg, S. C. Microbiological fractionation of sulphur isotopes. J. Gen. Microbiol. 34, 195–212 (1964).
Canfield, D. E. Isotope fractionation by natural populations of sulfate-reducing bacteria. Geochim. Cosmochim. Acta 65, 1117–1124 (2001).
Nehlich, O. The application of sulphur isotope analyses in archaeological research: A review. Earth-Sci. Rev. 142, 1–17 (2015).
Van Zalinge, N., Lieng, S., Ngor, P. B., Heng, K., & Valbo-Jørgensen, J. Status of the Mekong Pangasianodon hypophthalmus recourses, with special reference to the stocks shared between Cambodia and Vietnam. MRC Technical Paper No. 1. (Mekong River Commission, Phnom Penh, 2002).
Singh, A. K. & Lakra, W. S. Culture of Pangasianodon hypophthalmus into India: Impacts and present scenario. Pak. J. Biol. Sci. 15, 19–26 (2011).
Garcia, D. A. Z. et al. The same old mistakes in aquaculture: the newly-available striped catfish Pangasianodon hypophthalmus is on its way to putting Brazilian freshwater ecosystems at risk. Biodivers. Conserv. 27, 3545–3558 (2018).
Kumar, N. et al. Nano-zinc enhances gene regulation of non-specific immunity and antioxidative status to mitigate multiple stresses in fish. Sci. Rep. 13, 1–20 (2023).
Kumar, N. Dietary riboflavin enhances immunity and anti-oxidative status against arsenic and high temperature in Pangasianodon hypophthalmus. Aquaculture 533, 736209 (2021).
So, N., Van Houdt, J. K. J. & Volckaert, F. A. M. Genetic diversity and population history of the migratory catfishes Pangasianodon hypophthalmus and Pangasius bocourti in the Cambodian Mekong River. Fish. Sci. 72, 469–476 (2006).
Medo, A. et al. Gut morphometry represents diet preference to indigestible materials in the largest freshwater fish, Mekong giant catfish (Pangasianodon gigas). Zool. Sci. 37, 444–449 (2020).
Purnomo, K., Kartamihardja, E. S. & Koeshendrajanal, S. Pertumbuhan, mortalitas, dan kebiasaan makan ikan patin siam (Pangasius hypophthalmus) introduksi di waduk wonogiri. J. Penelit. Perikan. Indones. 9, 13–20 (2017).
Siaw-Yang, Y. Food resource utilization partitioning of fifteen fish species at Bukit Merah Reservoir Malaysia. Hydrobiologia 157, 143–160 (1988).
Musrafa-Kamal, A. S., Kamaruddin, I. S., Christianus, A., Daud, S. K. & Yu-Abit, L. Feeding habits of fishes in the Pengkalan Gawi-Pulau Dula section of Kenyir Lake Terengganu, Malaysia. Asian Fish. Sci. 25, 144–157 (2012).
Sangpradub, N., Pankai, P. & Hanjavanit, C. Diets of three Cyprinid species from Huai Pa Dang Reservoir Thailand. Trop. Nat. Hist. 15, 127–136 (2015).
Baird, I. G. Probarbus jullieni and Probarbus labeamajor: The management and conservation of two of the largest fish species in the Mekong River in southern Laos. Aquat. Conserv. Mar. Freshw. Ecosyst. 16, 517–532 (2006).
Shalloof, K. & Khalifa, N. Stomach contents and feeding habits of Oreochromis niloticus (L.) from Abu-Zabal Lakes, Egypt. World Appl. Sci. J. 6, 1–5 (2009).
Medo, A. et al. Stable carbon and nitrogen isotope ratios and fatty acid profiles in fishes in the Kaeng Krachan Reservoir, Thailand. Kyoto Univ. Res. Inf. Repos. https://doi.org/10.14989/262474 (2021)
Hessen, D. O. & Leu, E. Trophic transfer and trophic modification of fatty acids in high Arctic lakes. Freshw. Biol. 51, 1987–1998 (2006).
Caut, S., Angulo, E. & Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 46, 443–453 (2009).
Kohzu, A. et al. Stream food web fueled by methane-derived carbon. Aquat. Microb. Ecol. 36, 189–194 (2004).
Grey, J., Kelly, A. & Jones, R. I. High intraspecific variability in carbon and nitrogen stable isotope ratios of lake chironomid larvae. Limnol. Oceanogr. 49, 239–244 (2004).
Sarbu, S. M., Kane, T. C. & Kinkle, B. K. A chemoautotrophically based cave ecosystem. Science 272, 1953–1954 (1996).
MacAvoy, S. E., Morgan, E., Carney, R. S. & Macko, S. A. Chemoautotrophic production incorporated by heterotrophs in Gulf of Mexico hydrocarbon seeps: An examination of mobile benthic predators and seep residents. J. Shellfish Res. 27, 153–161 (2008).
Taipale, S. J. et al. Differing Daphnia magna assimilation efficiencies for terrestrial, bacterial, and algal carbon and fatty acids. Ecology 95, 563–576 (2014).
Vuorio, K., Meili, M. & Sarvala, J. Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshw. Biol. 51, 807–822 (2006).
Whiticar, M. J. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chem. Geol. 161, 291–314 (1999).
Zerkle, A. L., House, C. H. & Brantley, S. L. Biogeochemical signatures through time as inferred from whole microbial genomes. Am. J. Sci. 305, 467–502 (2005).
Sanseverino, A. M., Bastviken, D., Sundh, I., Pickova, J. & Enrich-Prast, A. Methane carbon supports aquatic food webs to the fish level. PLoS One 7, e42723. https://doi.org/10.1371/journal.pone.0042723 (2012).
Harrod, C. & Grey, J. Isotopic variation complicates analysis of trophic relations within the fish community of Plußsee: A small, deep, stratifying lake. Arch. für Hydrobiol. 167, 281–299 (2006).
Ravinet, M., Syväranta, J., Jones, R. I. & Grey, J. A trophic pathway from biogenic methane supports fish biomass in a temperate lake ecosystem. Oikos 119, 409–416 (2010).
Goedkoop, W., Sonesten, L., Markensten, H. & Ahlgren, G. Fatty acid biomarkers show dietary differences between dominant chironomid taxa in Lake Erken. Freshw. Biol. 40, 135–143 (1998).
Lefevre, S., Huong, D. T. T., Wang, T., Phuong, N. T. & Bayley, M. Hypoxia tolerance and partitioning of bimodal respiration in the striped catfish (Pangasianodon hypophthalmus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 158, 207–214 (2011).
Sánchez-Hernández, J., Nunn, A. D., Adams, C. E. & Amundsen, P. A. Causes and consequences of ontogenetic dietary shifts: A global synthesis using fish models. Biol. Rev. 94, 539–554 (2019).
Karube, Z., Okada, N., & Tayasu, I. Sulfur stable isotope signature identifies the source of reduced sulfur in benthic communities in macrophyte zones of Lake Biwa, Japan. Limnology (2012).
Proulx, I. & Hare, L. Differences in feeding behaviour among Chironomus species revealed by measurements of sulphur stable isotopes and cadmium in larvae. Freshw. Biol. 59, 73–86 (2014).
Pool, T. et al. Seasonal increases in fish trophic niche plasticity within a flood-pulse river ecosystem (Tonle Sap Lake, Cambodia). Ecosphere 8, e01881. https://doi.org/10.1002/ecs2.1881 (2017).
Gu, B., Chapman, A. D. & Schelske, C. L. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a softwater eutrophic lake. Limnol. Oceanogr. 51, 2837–2848 (2006).
Overman, N. C. & Parrish, D. L. Stable isotope composition of walleye: 15N accumulation with age and area-specific differences in δ13C. Can. J. Fish. Aquat. Sci. 58, 1253–1260 (2011).
Vander Zanden, M. J., Clayton, M. K., Moody, E. K., Solomon, C. T. & Weidel, B. C. Stable isotope turnover and half-life in animal tissues: A literature synthesis. PLoS One 10, e0116182. https://doi.org/10.1371/journal.pone.0116182 (2015).
Taipale, S. J., Kahilainen, K. K., Holtgrieve, G. W. & Peltomaa, E. T. Simulated eutrophication and browning alters zooplankton nutritional quality and determines juvenile fish growth and survival. Ecol. Evol. 8, 2671–2687 (2018).
Yokoyama, A., Arai, N., Mitamura, H., Mitsunaga, Y., Yamane, H., A., & Viputhanumas, T. Horizontal distribution and diel movements of 0-year-old hatchery-reared Mekong giant catfish Pangasianodon gigas in Kaeng Krachan reservoir, Thailand, obtained by acoustic telemetry. Nippon Suisan Gakkaishi, 85, 575–584 (2019).
Meesukko, C., Gajaseni, N., Peerapornpisa, Y. L. & Voinov, A. Relationships between seasonal variation and phytoplankton dynamics in Kaeng Krachan Reservoir, Phetchaburi Province Thailand. Trop. Nat. Hist. 7, 131–143 (2007).
Tayasu, I., Hirasawa, R., Ogawa, N. O., Ohkouchi, N. & Yamada, K. New organic reference materials for carbon- and nitrogen-stable isotope ratio measurements provided by Center for Ecological Research, Kyoto University, and Institute of Biogeosciences, Japan Agency for Marine-Earth Science and Technology. Limnology 12, 261–266 (2011).
Yanagisawa, F., Miyoshi, T., Ueda, A., & Sakai, H. Comparison of sulfur isotope ratio measurements by various techniques, and the δ34S values of some sulur standards. Inst. Therm. Spr. Res., Okayama Univ. 53, 77–84 (1983).
Zhang, C. L. et al. Lipid biomarkers and carbon isotope signatures of a microbial (Beggiatoa) mat associated with gas hydrates in the Gulf of Mexico. Appl. Environ. Microbiol. 71, 2106–2112 (2005).
He, R., Wooller, M. J., Pohlman, J. W., Tiedje, J. M. & Leigh, M. B. Methane-derived carbon flow through microbial communities in arctic lake sediments. Environ. Microbiol. 17, 3233–3250 (2015).
R Core Team. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. (2020).
Schneider, G., Chicken, E., & Becvarik, R. NSM3: Functions and Datasets to Accompany Hollander, Wolfe, and Chicken -Nonparametric Statistical Methods, Third Edition. R package version 1.16. https://CRAN.R-project.org/package=NSM3 (2021).
Acknowledgements
We thank the staff of the Inland Fisheries Patrol Unit, Kaeng Krachan District, Department of Fisheries, Thailand for their assistance with the fish sampling. Stable isotope analyses were conducted using an Isotope Ratio Mass Spectrometer in Joint Research Facilities of the Center for Ecological Research, Kyoto University and Research Institute for Humanity and Nature. We thank Dr. Masanori Katsuyama (Kyoto Prefectural University) for assistance with ion concentration analysis by chromatography. We would also like to thank Dr. Hideyuki Doi (Kyoto University) for available comments on writing this manuscript.
Author information
Authors and Affiliations
Contributions
Conceptualization: A.M., N.O., Y.O., K.K., N.A., T.V., H.M. Developing methods: Y.M., T.S., A.S.G., T.Y. Conducting the research: A.M., H.K., T.N., D.K., Y.O., Y.M., M.K., A.Y., H.M. Data analysis, figure, and table preparation: A.M. Funding acquisition: A.M., H.M. Data interpretation and writing: A.M., Y.O., K.K., H.N., N.O. with substantial help from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Medo, A., Ohte, N., Kajitani, H. et al. Striped catfish (Pangasianodon hypophthalmus) exploit food sources across anaerobic decomposition- and primary photosynthetic production-based food chains. Sci Rep 13, 13992 (2023). https://doi.org/10.1038/s41598-023-41209-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-41209-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
