
Abstract
In Egypt, Blastocystis sp. is not yet on the diagnostic list of parasitology reports, and information about its subtypes (STs) is scarce. This study investigated its prevalence and its STs/alleles, performed phylogenetic analysis, and considered the distribution of risk factors associated with Blastocystis sp. infections in West Ismailia, Ismailia governorate. Sociodemographic data, exposure factors, and previous parasitic infection status were recorded for symptomatic and asymptomatic individuals. Microscopy, polymerase chain reaction, sequencing, and phylogenetic analysis for Blastocystis sp. isolated from fecal samples were performed. Eighty Blastocystis sp.-infected individuals (15.3%) were examined. The age of the individuals ranged between 0.60 and 85.0 (mean 17.10 ± 15.70), the male/female ratio was 33/47, and the asymptomatic/symptomatic ratio was 55/25. The findings demonstrate clear evidence of direct contact with animals, poor water quality, and previous parasitic infections. Eleven samples yielded three Blastocystis STs (ST1: allele 4, ST2: alleles 9 and 12, and ST3: allele 34), with ST3 (45.5%) representing the most common subtype. Phylogenetic analysis with a robust bootstrap revealed three distinct clades for isolates of each subtype. This study updates the epidemiological knowledge of the distribution of Blastocystis sp. STs in Egypt and expands the current understanding of the prevalence, risk factor frequencies, and genetic diversity of this protist in the studied area.
Similar content being viewed by others


Introduction
Blastocystis sp. is a unicellular eukaryotic protist in the stramenopile family that lives in the guts of humans and animals. Blastocystis sp. has become a much more common issue for public health than previously thought because it is widely distributed with a high incidence and a high degree of genetic diversity1,2,3.
There are about 28 recognized Blastocystis sp. lineages. These subtypes (STs) are defined based on the genetic diversity of their small-subunit ribosomal RNA (SSU) genes. By 2013, 17 different STs (ST1 to ST17) had been identified. Although 11 further STs (ST18 to ST28) have been proposed, the validity of four of these (ST18, ST19, ST20, and ST22) remains contested3,4. The first nine subtypes and ST12 were isolated from the gastrointestinal tract of humans, with single instances of ST10, ST13, ST14, and ST16 also having been observed3,5,6,7,8,9,10. ST3, ST1, ST2, and ST4 are the most prevalent subtypes in humans. Rodents, birds, pigs, and other primates have all been colonized by different Blastocystis sp. subtypes3,5,6,7,8.
Blastocystis sp. is considered a parasite, and scientific consensus classifies it as a commensal and potentially even beneficial resident of the gut11,12. The hypothesis that certain strains within subtypes are pathogenic is under investigation10,13. Previous studies have speculated that Blastocystis sp. interacts with the host’s gut microbiota14,15. However, detailed insights remain lacking. Although Blastocystis sp. infections are generally asymptomatic in humans, common symptoms include nausea, anorexia, stomach discomfort, flatulence, and acute or chronic diarrhea. Such clinical manifestations have been suggested to result from the proteases and gut-microbiome dysbiosis caused by Blastocystis sp. colonization16.
Blastocystis sp. transmission is not clearly defined, but a human transmission cycle has been proposed17,18,19. Nonetheless, studies of family units in developed and developing countries have indicated that this pathway remains to be conclusively demonstrated20,21. Additional sources of infections appear to be contaminated water22,23,24,25, close contact with animals1, and contaminated soil20.
A lack of sanitation and clean water means that most Egyptian governorates are probably at high risk of Blastocystis infections, with several studies of the country linking Blastocystis sp. to urticaria, irritable bowel syndrome, asthma, and iron deficiency anemia, with infections diagnosed in both healthy (asymptomatic) and symptomatic individuals26,27,28.
Still, Blastocystis sp. is not listed as a pathogen in Egypt’s parasitological reports. According to a study on the diagnosis of gastrointestinal parasites by primary health care technicians in El-Kassassin, West Ismailia, Blastocystis sp. and Giardia duodenalis are completely missing from parasitological diagnosis results29.
Studies are lacking in Egypt concerning the epidemiology, molecular genetic diversity, and prevalence of Blastocystis sp. in carriers and non-carriers. Given the risk factors in Egypt and the current discussion of Blastocystis pathogenicity, this parasite should be included in the Egyptian “medical diagnostic radar.” Egypt has 27 governorates, one of which is Ismailia. The West Ismailia governorate has municipal divisions with rural areas, where domestic farm animals and birds share homes with residents. Inadequate hygiene is predicted, given the low socioeconomic status of the area. However, few molecular studies have been conducted to ascertain the incidence and subtype distribution of Blastocystis sp. in the governorate. Therefore, the present study aims to focus more closely on the prevalence, risk factor distribution, subtypes, and phylogenetics of Blastocystis sp. in a rural Egyptian community.
Results
Microscopy, polymerase chain reaction (PCR), sequencing, and phylogenetic analysis were performed for Blastocystis sp. from fecal samples (Fig. 1).

Fecal sample processing steps. (a) Blastocystis sp. vacuolar form and intermediate phase form using wet-mount examination. (b) Blastocystis sp. vacuolar form using iodine wet-mount examination. (c) PCR of some Blastocystis-positive samples. Lane 1 is 100 bp ladder (Promega), lane 2 is the positive control, lanes 3–4 are Blastocystis-positive samples, and lane 5 is the negative control. (d) Some sequences of Blastocystis sp. samples using MEGA software utilizing MUSCLE algorithm. (e) The phylogenetic tree of this study isolates’ sequences, cross-correlated with subtype allele.
Descriptive data concerning patient sociodemographics and the distribution of risk factors is illustrated for the 80 Blastocystis-positive samples (Tables 1 and 2). Microscopic and molecular examinations of Blastocystis-positive samples are shown in detail (Table 3, Fig. 2, and Table 4). Of the 15 samples sent for sequencing, 11 were successfully sequenced with high-quality yield (see "Methods" section for details). The genetic diversity and genotype distribution of Blastocystis sp. isolates are detailed in Table 5. Information regarding Blastocystis sp. STs in the current investigation, compared against studies of different Egyptian governorates, appears in Table 6. The current study’s sequence isolates were submitted to phylogenetic analysis (Fig. 3).

Parasitic stages associated with Blastocystis infection. (a) Blastocystis sp. vacuolar and intermediate-phase (arrowhead) form; (b) Chilomastix mesnili cyst (arrow) mixed with Blastocystis sp. cyst (arrowhead); (c) Entamoeba coli cyst; (d) Giardia duodenalis cyst; e: Entamoeba histolytica/dispar/moshkovskii cyst; (f) Hymenolepis nana egg. Photos a–e were taken with × 100; photo f was taken with × 40.

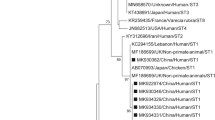
Phylogenetic analysis of the SSU rRNA (small subunit ribosomal RNA) nucleotide sequences of three subtypes of Blastocystis isolated from the West Ismailia population. The numbers above the branches indicate the percentages of bootstrap samplings. A solid circle indicates the Blastocystis species isolated in the current study, with Proteomonas lacerate used as an outgroup. RS: Reference sequence; IsKa: Ismailia–Kassassin isolates of the current study.
Blastocystis sp. was identified in a total of 80/520 (15.3%) fecal samples, according to direct light microscopy of smears (saline and iodine examination). The 80 Blastocystis-positive subjects included those with no available data (NAD = 47) and those with complete historical data available (SAD = 33). For NAD samples, the only available data were age, sex, and symptomatology status, with no information on exposure factors or previous parasitic infections (PPIs). Both SAD and NAD subjects were classified as “symptomatic” (gastrointestinal symptoms) or “asymptomatic” (no gastrointestinal symptoms), based on responses to a standardized questionnaire (Table 1).
The 80 Blastocystis-positive individuals ranged in age from 0.6 to 85 years. Most patients (55/80) had no gastrointestinal symptoms; those who did mostly complained of abdominal pain and diarrhea. Other non-specific symptoms such as vomiting, dehydration, and fever were also described (Table 1). Those aged between 5 and 18 (52/80) were most affected by Blastocystis sp. infection (Table 1). Approximately 60% (31) of this group were asymptomatic; the remainder were symptomatic. Females in this group represented 55.8% (29), and males represented 44.2% (23).
Most of the SAD subjects reported owning domestic animals (poultry, livestock, and pets). About 76.6% reported direct contact with their animals and that the floors of their animal farms were covered with sand. The water supplies at the studied subjects’ residences appeared mostly clear. However, some participants mentioned that the water sometimes became yellow and turbid with a smell (Table 1).
Almost all SAD individuals––31/33 (93.9%)––had recorded previous parasitic infections (PPI), including Entamoeba sp., Schistosoma sp., and Hymenolepis nana, in the previous year. Only 14 patients had received the anti-parasitic treatment they needed, with schistosomiasis patients, in particular, mostly not taking their medications or following their prescribed treatment regimens (Table 2).
According to microscopic wet-mount examinations of their fecal samples, 41.3% of participants recorded mixed parasitic infections (pathogenic and non-pathogenic). Polyparasitism included infections that were doubled, tripled, or even quadrupled. Other protozoan and helminthic parasites were found in Blastocystis-infected subjects, including Entamoeba coli, Entamoeba histolytica/dispar/moshkovskii, Chilomastix mesnili, Giardia duodenalis, and Hymenolepis nana (Table 3, Fig. 2).
Only 11 samples were successfully amplified, producing high-quality sequences that identified three STs (ST1, ST2, and ST3). Blastocystis ST3 was the most common ST in the West Ismailia population (45.5%), followed by ST2 and ST1 (27.3% each) (Table 4).
Tests performed using BLAST software revealed that each of the SSU rRNA gene sequences obtained in this study demonstrated a high degree of homology with previously reported sequences from other Blastocystis sp. isolates. Although ST1 and ST2 sequences showed some single-nucleotide polymorphism mutations against the reference strains, ST3 demonstrated 100% homology with the reference strain. Furthermore, 10/11 sequenced isolates belonged to asymptomatic subjects, with only one isolate (ST1) belonging to a symptomatic subject (Table 5).
The alignment of Blastocystis sequences showed three to five nucleotide differences. Three samples––IsKa 7 (ST2), IsKa 38 (ST2), and IsKa 75 (ST1)––demonstrated three nucleotide differences. Despite these differences, the BLAST search revealed that the first two of these isolates belong to the same Blastocystis ST. One sample, IsKa 29 (ST1), showed four nucleotide differences, while two samples, IsKa 13 (ST1) and IsKa 56 (ST2), showed five nucleotide differences. All of the sequence data for Blastocystis nucleotides obtained from the current study were deposited in GenBank with accession numbers OL845600–OL845610 as follows: ST3 represented by five samples (OL845600–OL845604), ST2 represented by three samples (OL845605–OL845607), and ST1 represented by three samples (OL845608–OL845610) (Table 5). In terms of Blastocystis alleles, ST3 and ST1 produced completely homogeneous isolates with alleles 34 and allele 4, respectively. Meanwhile, ST2 exhibited low genetic variation in one isolate (IsKa 7), which corresponded to allele 12, with its remaining two isolates corresponding to allele 9.
A phylogenetic tree was constructed using eleven nucleotide sequences representative of the current study and GenBank database reference sequences, with Proteromonas lacertae (U37108) used as the outgroup. Three subtypes were distinguishable: ST1, ST2, and ST3 (Fig. 1).
Discussion
This study’s findings revealed Blastocystis sp. to be prevalent in 15.3% of the West Ismailia population, which is consistent with the observations of El-Badry et al. (2018). Other studies have revealed variations in Blastocystis prevalence27,30,31. PCR in Cairo revealed a prevalence of 35.5% for Blastocystis in both patient and control groups30. Patients with irritable bowel syndrome in Beni-Suef were found to have a prevalence of Blastocystis of 16.5% by microscopy and 19.1% by culture27. In the same governorate, 53.6% of patients with acute diarrhoea had Blastocystis31. Such disparities can be attributed to various epidemiological parameters, including the target population, detection method, and the presence or absence of symptoms.
In this study, Blastocystis colonization was more common among children aged 6 to 18 than among children under five years old and adults, which aligns with two previous Egyptian studies observing schoolchildren to be the most affected32,33. Infection with Blastocystis sp. has also been demonstrated by global surveys to be prevalent among schoolchildren18,34,35,36, likely owing to this age group’s lax hygiene standards. Blastocystis infection age disparities may be influenced by associated exposure risk factors, children’s immunity, and environmental variables34,35,36.
This study classifies Blastocystis infections as silent, with most infected subjects not exhibiting symptoms. Three factors may influence the asymptomatic status of Blastocystis infections among Egyptians: (a) awareness, which is limited in the rural Egyptian population, of the need to seek medical attention for diarrhea; (b) the diagnostic ability of Egyptian laboratories to identify Blastocystis sp. in primary care settings (i.e., primary health care units), leaving Blastocystis outside their diagnostic scope29; and (c) in the case of Blastocystis infection, the concept that it is a commensal protist, which encourages Egyptian physicians to disregard treating patients despite the presence of distressing symptoms29. These factors may enable Blastocystis sp. to colonize the host for an extended period without causing disease.
Most Egyptians, and particularly those living in rural areas, keep domestic animals and birds in and around their homes. This is especially evident in West Ismailia, where domestic animals, especially poultry, coexist with humans in traditional Egyptian residences without separate yards, exposing owners to high concentrations of Blastocystis infective stages for extended periods. This has led to the prediction that the risk of Blastocystis infection is tenfold greater in rural areas than in urban areas among the Egyptian population with irritable bowel syndrome27. Elsewhere, strong molecular evidence has confirmed zoonotic transmission between animals and their human caregivers37,38. Notably, humans in direct contact with animals were found to have the same Blastocystis STs in two studies conducted in Northern and Eastern Egypt (ST1, ST2, ST3, and ST4)37,39. Additionally, poultry can harbor the human-transmissible Blastocystis ST6 and ST740, although this has only been reported in a recent Egyptian study of colorectal cancer patients41.
The Sweet Water Canal serves as the primary water source for municipal divisions in West Ismailia and is used directly for animal bathing, dishwashing, and laundry. Consequently, water used for residential or recreational purposes becomes contaminated, creating a risk of transmission of gastrointestinal diseases to humans and animals alike. Rural communities in West Ismailia have been particularly hard-hit, with some settlements in remote locations lacking access to safe drinking water, which increases the possibility of contamination during transportation and processing. Numerous protozoan contaminants, including Blastocystis sp., have been detected in Egypt’s Dakahlia, Ismailia, and El-Minia governorates in potable water, water tanks, pumps, waterworks, and surface water (i.e., River Nile, ponds, and canals)24,42,43. Most SAD residents were observed to utilize tap water without a filter, with the water supplied appearing clear. However, because it is only used to remove rust, insects, and dust, the presence or absence of a filter would not substantially impact the purification of water from protozoa stages. Thus, even if the water appears clear, it may contain Blastocystis sp. Furthermore, in recent years, soil pollution has been identified as a source of Blastocystis sp. infection20; as a major farming region, soil is extremely likely to be a source of infection for the West Ismailia population.
PPI are prevalent in rural Ismailia (author’s observation, unpublished data); when questioned, this study’s participants were fully aware of this. Over half of participants failed to take their parasitic infection medications as prescribed and did not complete the entire course of treatment (dose and duration). Consequently, infected individuals have acted as carriers facilitating anthroponotic protist transmission, while also experiencing infection maintenance, chronicity, and consequences.
This study’s investigations also revealed polyparasitism in the West Ismailia population. Although most samples represented single Blastocystosis infections, the presence of pathogenic (G. duodenalis, E. histolytica, H. nana) and non-pathogenic (E. coli, C. mesnilli, non-pathogenic species of E. histolytica) parasites mixed with Blastocystis sp. infection suggests multiple sources of infection. Mixed parasitic infections are highly predicted in rural areas due to the presence of multiple risk factors, as has been previously documented44,45,46,47. Consumption of contaminated water and unwashed vegetables, lack of fingernail trimming and hand washing, children playing in the dirt, barefoot walking, low socioeconomic status, lack of sanitation, and large numbers of family members sharing a single room have all been observed among the residents of West Ismailia27,31,48,49. Furthermore, rats, cockroaches, fleas, ants, and flies were observed to spread in numerous locations, especially during hot weather, alongside sewage rash. Such behavioral, social, and sanitary factors are almost certainly implicated in developing mixed parasitic infections and perpetuating the life cycle of those infections. Thus, nearly all the transmission routes required for Blastocystis sp. appear open.
Three subtypes of Blastocystis sp. were characterized in the current investigation via the molecular analysis of isolates, namely, ST3, ST2, and ST1, with the latter two recording equal distribution. This study’s findings corroborate those of Souppart et al. (2010), who discovered that ST3 had the highest prevalence (61.9%) and that ST1 and ST2 had equal prevalence (19.05%).
Several Egyptian studies have subtyped and sequenced Blastocystis sp.39,41,50, with five STs (ST1, ST2, ST3, ST4 and ST7) identified at varying frequencies in distinct Egyptian groups using PCR sequenced-tagged sites, PCR restriction fragment length polymorphisms, and PCR sequencing (barcoding) (Table 6). The high prevalence of Blastocystis sp. ST1–ST4 in the Egyptian community suggests that most infections are transmitted from person to person.
The current investigation has revealed that ST3 is responsible for the vast majority of Blastocystis infections in West Ismailia, aligning with observations for other Egyptian governorates across 12 other studies (Table 6), which have revealed ST3 to be the most prevalent Blastocystis subtype in six distinct Egyptian locations; furthermore, it is the ST most closely related to various gastrointestinal symptoms (Table 6). Other subtypes (ST1, ST2, ST4 and ST7) have been detected in the Egyptian community, with varying frequencies depending on the sample size and testing technique used. ST1 and ST2 have been identified as relevant STs in a smaller number of Egyptian studies28,39. Notably, STs and their relative frequencies appear to vary significantly within a single country (Table 6).
Almost all the isolates sequenced in this investigation were asymptomatic, except for one patient who suffered diarrhea and abdominal pain and was subtyped as ST1, an observation consistent with the findings of other studies10,55,56,57. Blastocystis is more frequent in healthy individuals, with its existence also linked to altered composition and increased richness of the bacterial gut microbiota14,15. It is unclear whether Blastocystis directly promotes a healthy gut and microbiome or whether it prefers to colonize and persist in a healthy gut environment. A study of Blastocystis sp. ST3 indicated that Blastocystis sp. may modify the gut ecosystem in a protective manner and facilitate faster recovery from disturbances12. The presence of Blastocystis among healthy individuals has also been linked to reduced levels of fecal calprotectin, a sign of intestinal inflammation according to a comparative investigation conducted in Mexico56. In contrast, some researchers have suggested that particular Blastocystis sp. isolates may produce an imbalance of the gut microbiota58,59,60,61,62,63. The context of the environment and hosts must be considered when discussing whether Blastocystis is a pathogen or a mutualist.
The present study’s genetic analysis reveals that all 11 isolates detected in Blastocystis subtypes cross-corresponded to previously observed alleles. Intriguingly, ST3 isolates produced the highest frequency of isolates matching allele 34, the most common variant found in humans worldwide, with ST2 isolates exhibiting low levels of genetic diversity and multiple nucleotide substitutions corresponding to two different alleles (9 and 12). Although ST1 isolates demonstrated limited genetic diversity, all isolates corresponded to allele 4. In Egyptian isolates from Cairo, genetic diversity was detected in three subtypes in the same pattern, with ST1 and ST2 exhibiting nucleotide differences ranging from 1 to 11 and ST3 exhibiting reduced genetic variability of up to four nucleotide differences50. However, no further allelic analysis was conducted. On the contrary, there was no evidence of genetic diversity in the Blastocystis subtypes (ST3 and ST1) isolated from individuals with irritable bowel syndrome in the Beni-Suef governorate27.
A phylogenetic tree demonstrated that the 11 nucleotide sequences in this study clustered into the same subtype cluster, with high bootstrap support, and could be classified into three subtypes: ST1, ST2, and ST3. Each Blastocystis sp. ST formed a distinct clade, implying that the West Ismailian Blastocystis population can be divided into three subgroups. Another Egyptian study found the same phylogenetic distribution pattern for ST1 and ST3 subtype clusters in patients with irritable bowel syndrome27.
Among the drawbacks of the present investigation is the infeasibility of sequencing every isolate. Because samples were collected via a large-scale survey, there were no epidemiological data for some participants, which hindered presenting comprehensive information.
Blastocystis sp. infections are significantly under- and mis-diagnosed in Egypt, particularly in rural and remote areas such as West Ismailia. Additional research can illuminate the epidemiological situation of Blastocystis sp. in Egypt, enabling more effective control efforts against Blastocystis sp. infections and other parasitic disorders.
Conclusions
The current study updates the epidemiological situation and distribution of Blastocystis STs in Egypt. Phylogenetic analysis has revealed three distinct clades for isolates pertaining to each subtype, adding to our current understanding of Blastocystis’s prevalence and genetic diversity. The widespread presence of ST3 in the West Ismailia population and throughout Egypt necessitates subtyping analysis, which has become indispensable for elucidating the relationship between Blastocystis subtypes and pathogenicity in the Egyptian population.
We highlight the need to invest in parasite education programs, specific to Blastocystis sp., that involve the general public along with doctors and laboratory technicians. Moreover, further studies are needed in the underrepresented areas of Egypt to verify the distribution of Blastocystis sp. throughout the country.
Methods
Study area and sample collection
The Ministry of Health and Population in West Ismailia’s municipal divisions (villages around the Sweet Water Canal’s geographical line) conducted a screening survey of 598 fecal samples for schistosomiasis eradication campaigns. Samples were collected randomly, without regard for age or gender, from the nearest sampling area.
Thus, this descriptive study used data from screens for gastrointestinal parasites of individuals in the municipal divisions of West Ismailia (El-Kassassin, El-Mahsama, El-Talelkbeer, and Abu-Suwayr). A total of 80 fecal samples positive for Blastocystis sp. were analyzed. The flowchart in Fig. 4 describes the current study’s process.

Flowchart describing the origin of the current study’s samples. SAD: Subjects with available data; NAD: Subjects with no available data; MOHP: Ministry of Health and Population; GIPs: Gastrointestinal parasites; SCU: Suez Canal University.
Five hundred and twenty samples were selected after considering their suitability based on inclusion and exclusion criteria. The 520 fecal samples and their questionnaire forms were sent to Suez Canal University’s Parasitology Laboratory for ovum and parasite examination, using wet mount and iodine microscopy to screen for gastrointestinal parasites. Blastocystis-positive samples were separated and selected for the current study analysis. The amount of the received fecal samples (a full tablespoon, i.e., 15–20 gr) was used as a guideline for rolling in or rolling out the received fecal samples.
To collect and transfer the fecal samples, participants were given a clean, labeled plastic container with an applicator stick. Patients were given verbal explanations in Arabic regarding the collection and transfer processes and the amount of stool sample required. Stool samples were excluded if contaminated with urine and water or if the amount was too small (less than a full tablespoon, i.e., 15–20 gr). Subsequently, the parasitology laboratory at El-Mahsama Family Practice Center divided the stool samples into two parts: one part for the eradication program campaign (primary health care unit at the parasitology laboratory) and one part for processing at the Suez Canal University’s Medical Department’s parasitology laboratory in Ismailia.
Microscopy of fecal samples
A teaspoon-sized fecal sample was combined with 50 mL saline. In a 15 mL conical plastic tube, the mixture was strained using gauze. One mL of the filtered fecal mixture was pipetted into Eppendorf tubes and frozen. For Blastocystis form diagnosis, microscopic examination was conducted using a direct smear. To view other parasitic stages under the microscope, the formalin-ethyl acetate sedimentation procedure was employed to concentrate the strained mixture64. Wet-mount analysis was performed and Lugol’s iodine added to the slides for microscopic examination.
Extraction of genomic DNA
Before extraction, the preserved 1 mL fecal samples were washed and centrifuged several times in phosphate-buffered saline until the supernatant became clear. Then, the supernatant was discarded, and 200 µL of the 1 mL sediment was exposed to the InhibitEX lysis buffer from the Qiagen DNA Stool Mini Kit (Qiagen, Germany, GmbH) according to the manufacturer’s protocol. Using 100 µL of elution buffer, the protocol was slightly modified, and the extracted DNA was stored at -20 °C for further molecular investigation.
PCR amplification and sequencing
Financial restrictions meant that only 15 samples were sent to the Istituto Superiore di Sanità in Rome, Italy, for molecular characterization. For amplification of the SSU rRNA region in Blastocystis isolates, Blasto F (5′-TCTGGTTGATCCTGCCAGT-3′) and Blasto R (5′-AGCTTTTTAACTGCAACAACG-3′) primers were used according to the protocol described by Meloni et al.65. A proofreading enzyme (GoTaq Promega) was used to amplify the PCR. The Promega Wizard SV Gel and PCR Clean-Up System kit was used to purify the obtained 600 bp amplicon, and purified products were sent for MGW Sanger sequencing.
The sequences obtained were compared with other Blastocystis SSU rRNA gene sequences available in the National Center for Biotechnology Information database using the BLAST tool (http://www.ncbi.nlm.nih.gov). Multiple nucleotide sequence alignments were performed using the MUSCLE algorithm (https://www.megasoftware.net/web_help_10/Part_II_Assembling_Data_For_Analysis/Building_Sequence_Alignments/MUSCLE/About_Muscle.htm) of the MEGA software (https://www.megasoftware.net/)3,66. Additionally, Blastocystis alleles were identified by determining their closest similarity to known Blastocystis sequences using the Blastocystis PubMLST database (https://pubmlst.org/organisms/blastocystis-spp).
Phylogenetic analysis
Molecular and Evolution Genetic Analysis software (https://www.megasoftware.net/) was used to produce a phylogenetic tree for nucleotide sequences using the maximum likelihood method. One thousand bootstrap replicates were used to test the phylogenetic tree’s reliability and the statistical support for the topology. Evolutionary distances were calculated using the Tamura-3 parameter model.
Statistical analysis
A descriptive analysis was used to report sociodemographic characteristics and the frequency and distribution of Blastocystis subtypes among the affected subjects. After uploading all questionnaire data, the IBM SPSS software package version 26.0 was used to analyze the data (IBM Corporation, Armonk, NY). Numbers and percentages were used to represent categorical data; mean and standard deviation were used to represent numerical data. The chi-squared test was used to examine the relationship between a PPI and its treatment as two qualitative variables. The p-value was statistically significant at 0.05.
Ethical considerations
Fecal sample collection and medical history questionnaires were reviewed and approved by the Scientific Research Committee and Bioethics Board of Suez Canal University, Faculty of Medicine, Egypt (approval no. 5089). All methods were performed in accordance with the relevant guidelines. Participants in the study were asked to sign a written informed consent form that clearly detailed in Arabic the study’s objectives, sociodemographic questionnaire, symptomatology, exposure factors, and procedures. The collected data were kept private during and after the collection and analysis. The study’s participants were informed that they could withdraw at any time. Patients whose fecal specimens were positive for gastrointestinal parasites were referred to physicians for further treatment.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Ahmed, S. A. & Karanis, P. Blastocystis spp., ubiquitous parasite of human, animals and environment. in Reference Module in Earth Systems and Environmental Sciences 1–6 (Elsevier, 2019). https://doi.org/10.1016/B978-0-12-409548-9.10947-9.
Scanlan, P. D. et al. The microbial eukaryote Blastocystis is a prevalent and diverse member of the healthy human gut microbiota. FEMS Microbiol. Ecol. 90, 326–330 (2014).
Stensvold, C. & Clark, C. G. Pre-empting pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 36, 229–232 (2020).
Hublin, J. S. Y., Maloney, J. G. & Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 135, 260–282 (2021).
Maloney, J. G., da Cunha, M. J. R., Molokin, A., Cury, M. C. & Santin, M. Next-generation sequencing reveals wide genetic diversity of Blastocystis subtypes in chickens including potentially zoonotic subtypes. Parasitol. Res. 120, 2219–2231 (2021).
Maloney, J. G., Jang, Y., Molokin, A., George, N. S. & Santin, M. wide genetic diversity of Blastocystis in white-tailed deer (Odocoileus virginianus) from Maryland, USA. Microorganisms 9, (2021).
Maloney, J. G., Molokin, A., da Cunha, M. J. R., Cury, M. C. & Santin, M. Blastocystis subtype distribution in domestic and captive wild bird species from Brazil using next generation amplicon sequencing. Parasite Epidemiol. Control 9, (2020).
Higuera, A. et al. Identification of multiple Blastocystis subtypes in domestic animals from Colombia using amplicon-based next generation sequencing. Front. Vet. Sci. 8, 932 (2021).
Ramírez, J. D. et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 41, 32–35 (2016).
Tito, R. Y. et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 68, 1180–1189 (2019).
Mülayim, S. et al. investigation of isolated Blastocystis subtypes from cancer patients in Turkey. Acta Parasitol. 66, 584–592 (2021).
Billy, V. et al. Blastocystis colonization alters the gut microbiome and, in some cases, promotes faster recovery from induced colitis. Front. Microbiol. 12, (2021).
Betts, E. L. et al. Metabolic fluctuations in the human stool obtained from Blastocystis carriers and non-carriers. Metabolites 11, 883 (2021).
Andersen, L. O. brie. & Stensvold, C. R. Blastocystis in health and disease: Are we moving from a clinical to a public health perspective? J. Clin. Microbiol. 54, 528 (2016).
Stensvold, C. R. et al. Differentiation of Blastocystis and parasitic archamoebids encountered in untreated wastewater samples by amplicon-based next-generation sequencing. Parasite Epidemiol. Control 9, e00131 (2020).
Mirjalali, H. et al. Distribution and phylogenetic analysis of Blastocystis sp. subtypes isolated from IBD patients and healthy individuals in Iran. Eur. J. Clin. Microbiol. Infect. Dis. 36, 2335–2342 (2017).
Paulos, S. et al. Occurrence and subtype distribution of Blastocystis sp. in humans, dogs and cats sharing household in northern Spain and assessment of zoonotic transmission risk. Zoonoses Public Health 65, 993–1002 (2018).
Muadica, A. S. et al. Molecular diversity of Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp in asymptomatic school children in Leganés Madrid Spain. Microorganisms 8, 466 (2020).
Khaled, S. et al. Blastocystis sp. prevalence and subtypes distribution amongst Syrian refugee communities living in North Lebanon. Microorganisms 9, 1–13 (2021).
Jinatham, V., Maxamhud, S., Popluechai, S., Tsaousis, A. D. & Gentekaki, E. Blastocystis one health approach in a rural community of Northern Thailand: Prevalence, subtypes and novel transmission routes. Front. Microbiol. 12, 746340 (2021).
Lhotská, Z. et al. A study on the prevalence and subtype diversity of the intestinal protist Blastocystis sp in a gut-healthy human population in the Czech Republic. Front. Cell. Infect. Microbiol. 10, 544335 (2020).
Taamasri, P. et al. Transmission of intestinal blastocystosis related to the quality of drinking water - PubMed. Southeast Asian J Trop Med Public Heal. 31, 112–117 (2000).
Leelayoova, S. et al. Evidence of waterborne transmission of Blastocystis hominis–PubMed. Am. J. Trop. Med. Hyg. 70, 658–662 (2004).
Elshazly, A., Elsheikha, H., Soltan, D., Mohammad, K. & Morsy, T. Protozoal pollution of surface water sources in Dakahlia Governorate, Egypt–PubMed. J. Egypt. Soc. Parasitol. 37, 51–74 (2007).
Angelici, M. C., Nardis, C., Scarpelli, R. & Ade, P. Blastocystis hominis transmission by non-potable water: A case report in Italy–PubMed. New Microbiol. 41, 173–177 (2018).
Abdel-Hameed, D. M. A., Hassanin, O. M. & Zuel-Fakkar, N. M. Association of Blastocystis hominis genetic subtypes with urticaria. Parasitol. Res. 108, 553–560 (2011).
El-Badry, A. A., Abd El Wahab, W. M., Hamdy, D. A. & Aboud, A. Blastocystis subtypes isolated from irritable bowel syndrome patients and co-infection with Helicobacter pylori. Parasitol. Res. 117, 127–137 (2018).
Deeb El, H. K. & Khodeer, S. Blastocystis spp: Frequency and subtype distribution in iron deficiency anemic versus non-anemic subjects from Egypt. J. Parasitol. 99, 599–602 (2013).
Ahmed, S. A., Mohamed, S. F., Fouad, A. M. & Karanis, P. Gastrointestinal parasites diagnoses at the primary health care units: A comparative analysis of diagnostic abilities of parasitology staff technicians versus medical parasitologists in Ismailia, Egypt. R. Soc. Trop. Med. Hyg. trac072 (2022).
Zuel-Fakkar, N. M., Abdel Hameed, D. M. & Hassanin, O. M. Study of Blastocystis hominis isolates in urticaria: A case-control study. Clin. Exp. Dermatol. 36, 908–910 (2011).
Hamdy, D. A., Abd El Wahab, W. M., Senosy, S. A. & Mabrouk, A. G. Blastocystis spp. and Giardia intestinalis co-infection profile in children suffering from acute diarrhea. J. Parasit. Dis. 44, 88–98 (2020).
Abu El-Fetouh, N., Abdelmegeed, E., Attia, R., El-Dosoky, I. & Azab, M. Genotyping of Blastocystis hominis symptomatic isolates and kinetics of associated local CD3 and CD20 cell infiltrate. Parasitol. United J. 8, 122 (2015).
El Saftawy, E. A., Amin, N. M., Hamed, D. H., Elkazazz, A. & Adel, S. The hidden impact of different Blastocystis genotypes on C-3 and IgE serum levels: A matter of debate in asthmatic Egyptian children. J. Parasit. Dis. 43, 443–451 (2019).
Bertozzo, T. V., David, É. B., Oliveira-Arbex, A. P., Victória, C. & Guimarães, S. Frequency, spatial distribution, and genetic diversity of Blastocystis among referred individuals to a clinical laboratory: First report of subtype 9 in Brazil. Acta. Trop. 234, (2022).
Khaled, S. et al. Prevalence and subtype distribution of Blastocystis sp. in Senegalese school children. Microorganisms 8, 1408 (2020).
Cinek, O. et al. Blastocystis in the faeces of children from six distant countries: Prevalence, quantity, subtypes and the relation to the gut bacteriome. Parasit. Vectors 14, (2021).
Mokhtar, A. & Youssef, A. Subtype analysis of Blastocystis spp isolated from domestic mammals and poultry and its relation to transmission to their incontact humans in Ismailia governorate, Egypt. Parasitol. United J. 11, 1687–7942 (2018).
Udonsom, R. et al. Blastocystis infection and subtype distribution in humans, cattle, goats, and pigs in central and western Thailand. Infect. Genet. Evol. 65, 107–111 (2018).
Abdo, S. M. et al. Detection and molecular identification of Blastocystis isolates from humans and cattle in northern Egypt. J. Parasit. Dis. 45, 738–745 (2021).
Rauff-Adedotun, A. A., Mohd Zain, S. N. & Farah Haziqah, M. T. Current status of Blastocystis sp. in animals from Southeast Asia: A review. Parasitol. Res. 119, 3559–3570 (2020).
Ali, S. H. et al. an association between Blastocystis subtypes and colorectal cancer patients: A significant different profile from non-cancer individuals. Acta Parasitol. 67, 752–763 (2022).
Rayan, H. Z., Eida, O. M., El-hamshary, E. M. & Ahmed, S. A. Detection of human Cryptosporidium species in surface water sources in Ismailia using polymerase chain reaction. Parasitol. United J. 2, 119–126 (2009).
Khalifa, R. M. A., Ahmad, A. K., Abdel-Hafeez, E. H. & Mosllem, F. A. Present status of protozoan pathogens causing water-borne disease in Northern part of El-Minia governorate Egypt. J. Egypt. Soc. Parasitol. 44, 559–566 (2014).
Fernández-Niño, J. A. et al. Profiles of intestinal polyparasitism in a community of the Colombian Amazon region. Biomedica 37, 368–377 (2017).
Pagheh, A. S. et al. A cross-sectional analysis of intestinal parasitic infections among the general population in north of Iran. J. Infect. Dev. Ctries. 12, 120–126 (2018).
Dawaki, S., Al-Mekhlafi, H. M. & Ithoi, I. The burden and epidemiology of polyparasitism among rural communities in Kano State Nigeria. Trans. R. Soc. Trop. Med. Hyg. 113, 169–182 (2019).
Weerakoon, K. G. et al. Co-parasitism of intestinal protozoa and Schistosoma japonicum in a rural community in the Philippines. Infect. Dis. Poverty 7, (2018).
Elmonir, W. et al. Prevalence of intestinal parasitic infections and their associated risk factors among preschool and school children in Egypt. PLoS ONE 16, e0258037 (2021).
Yones, D., Zaghlol, K., Abdallah, A. & Galal, L. Effect of enteric parasitic infection on serum trace elements and nutritional status in upper Egyptian children. Trop. Parasitol. 5, 35 (2015).
Souppart, L. et al. Subtype analysis of Blastocystis isolates from symptomatic patients in Egypt. Parasitol. Res. 106, 505–511 (2010).
Abaza, S., Rayan, H., Soliman, R., Nemr, N. & Mokhtar, A. Subtype analysis of Blastocystis spp. isolates from symptomatic and asymptomatic patients in Suez Canal University Hospitals, Ismailia. Egypt. Parasitol. United J. 7, 56–67 (2014).
El-Taweel, H. et al. Restriction fragment length polymorphism RFLP analysis of Blastocystis spp. in symptomatic and asymptomatic individuals from Alexandria. Egypt. Parasitol. United J. 13, 164–171 (2020).
Fouad, S. A., Basyoni, M. M. A., Fahmy, R. A. & Kobaisi, M. H. The pathogenic role of different Blastocystis hominis genotypes isolated from patients with irritable bowel syndrome. Arab J. Gastroentrol. 12, 194–200 (2011).
Hussein, E. M., Hussein, A. M., Eida, M. M. & Atwa, M. M. Pathophysiological variability of different genotypes of human Blastocystis hominis Egyptian isolates in experimentally infected rats. Parasitol. Res. 102, 853–860 (2008).
Mardani Kataki, M., Tavalla, M. & Beiromvand, M. Higher prevalence of Blastocystis hominis in healthy individuals than patients with gastrointestinal symptoms from Ahvaz, southwestern Iran. Comp. Immunol. Microbiol. Infect. Dis. 65, 160–164 (2019).
Nieves-Ramírez, M. E. et al. Asymptomatic intestinal colonization with protist Blastocystis is strongly associated with distinct microbiome ecological patterns. mSystems 3, e00007-18 (2018).
Kesuma, Y., Firmansyah, A., Bardosono, S., Sari, I. P. & Kurniawan, A. Blastocystis ST-1 is associated with irritable bowel syndrome-diarrhoea (IBS-D) in Indonesian adolescences. Parasite Epidemiol. Control 6, (2019).
Yason, J. A., Liang, Y. R., Png, C. W., Zhang, Y. & Tan, K. S. W. Interactions between a pathogenic Blastocystis subtype and gut microbiota: In vitro and in vivo studies. Microbiome 7, 30 (2019).
Stensvold, C. R., Arendrup, M. C., Nielsen, H. V., Bada, A. & Thorsen, S. Symptomatic infection with Blastocystis sp. subtype 8 successfully treated with trimethoprim-sulfamethoxazole. Ann. Trop. Med. Parasitol. 102, 271–274 (2008).
Stensvold, C. R., Lebbad, M. & Verweij, J. J. The impact of genetic diversity in protozoa on molecular diagnostics. Trends Parasitol. 27, 53–58 (2011).
Mattiucci, S., Crisafi, B., Gabrielli, S., Paoletti, M. & Cancrini, G. Molecular epidemiology and genetic diversity of Blastocystis infection in humans in Italy. Epidemiol. Infect. 144, 635–646 (2016).
Gabrielli, S. et al. Molecular Subtyping of Blastocystis sp. isolated from farmed animals in Southern Italy. Microorganisms 9, (2021).
Stensvold, C. R., Alfellani, M. & Clark, C. G. Levels of genetic diversity vary dramatically between Blastocystis subtypes. Infect. Genet. Evol. 12, 263–273 (2012).
Cociancic, P., Rinaldi, L., Zonta, M. L. & Navone, G. T. Formalin-ethyl acetate concentration, FLOTAC pellet and anal swab techniques for the diagnosis of intestinal parasites. Parasitol. Res. 117, 3567–3573 (2018).
Meloni, D. et al. Molecular subtyping of Blastocystis sp. isolates from symptomatic patients in Italy. Parasitol. Res. 109, 613–619 (2011).
Stensvold, C. R. et al. Terminology for Blastocystis subtypes–a consensus. Trends Parasitol. 23, 93–96 (2007).
Acknowledgements
The study was supported by Taif University Researchers Supporting Project (TURSP-2020/152), Taif University, Taif, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
S.A. conception of the study, study design, methodology, formal analysis and investigation, interpretation of the results, writing original draft of manuscript; H.E., S.M., M.A., K.H., P.K. methodology, formal analysis and investigation, interpretation of the results, editing manuscript; S.A., H.E., S.M., M.A and T.S. resources of the experiment. P.K. and T.S language profiling. P.K. mentoring, manuscript editing-revision, supervision. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ahmed, S.A., El-Mahallawy, H.S., Mohamed, S.F. et al. Subtypes and phylogenetic analysis of Blastocystis sp. isolates from West Ismailia, Egypt. Sci Rep 12, 19084 (2022). https://doi.org/10.1038/s41598-022-23360-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-23360-0
This article is cited by
-
Mapping gut parasitism patterns in a cohort of Egyptians
Scientific Reports (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
