
Abstract
The production of second-generation fuels from lignocellulosic residues such as sugarcane bagasse (SCB) requires the synergistic interaction of key cellulose-degrading enzymes and accessory proteins for their complete deconstruction to useful monomeric sugars. Here, we recombinantly expressed and characterized unknown GH5 xylanase from P. funiculosum (PfXyn5) in Pichia pastoris, which was earlier found in our study to be highly implicated in SCB saccharification. The PfXyn5 has a molecular mass of ~ 55 kDa and showed broad activity against a range of substrates like xylan, xyloglucan, laminarin and p-nitrophenyl-β-d-xylopyranoside, with the highest specific activity of 0.7 U/mg against xylan at pH 4.5 and 50 °C. Analysis of the degradation products of xylan and SCB by PfXyn5 showed significant production of xylooligosaccharides (XOS) with a degree of polymerization (DP) ranging from two (DP2) to six (DP6), thus, suggesting that the PfXyn5 is an endo-acting enzyme. The enzyme synergistically improved the saccharification of SCB when combined with the crude cellulase cocktail of P. funiculosum with a degree of synergism up to 1.32. The PfXyn5 was further expressed individually and simultaneously with a notable GH16 endoglucanase (PfEgl16) in a catabolite-derepressed strain of P. funiculosum, PfMig188, and the saccharification efficiency of the secretomes from the resulting transformants were investigated on SCB. The secretome of PfMig188 overexpressing Xyn5 or Egl16 increased the saccharification of SCB by 9% or 7%, respectively, over the secretome of PfMig188, while the secretome of dual transformant increased SCB saccharification by ~ 15% at the same minimal protein concentration.
Similar content being viewed by others

Introduction
The rapid increase in energy demand along with the continuous increase in the price of fossil fuels constantly necessitates the need to diversify the channels of energy generation for every growing economy1,2,3. The increasing shortage of non-renewable energy resources and huge environmental pressure from greenhouse gases released by the burning of fossil fuels continue to prompt growing studies in search of alternative production of cleaner fuels compared to petroleum-based fuels. Second-generation biofuels produced from lignocellulosic biomass continue to attract wide attention as bountiful sources as they neither compete with food requirements nor impact the food and feed markets4,5. Sugarcane bagasse (SCB) is one of the most abundant and promising biomass sources in the world which is obtained from the processing of sugarcane with a worldwide annual production of approximately 54 million tons6. On average, approximately 270 kg of bagasse (50% moisture) per metric ton of sugarcane is generated by the sugar factories as a major by-product7. SCB predominantly consists of cellulose, hemicellulose and lignin that are strongly associated to form the plant cell wall8,9.
The bioconversion process of this biomass requires several arrays of lignocellulolytic enzymes which act synergistically to deconstruct the tightly packed polymeric carbohydrate components into monomeric sugars that can subsequently be converted to fuels and valuable chemicals. Previous studies have shown that several biomass conversion processes rely on biomass pretreatment before enzymatic hydrolysis, to remove lignin and reduce the recalcitrance nature of the biopolymer10,11,12. However, this process often leads to a reduction in the hemicellulose content13. Owing to this challenge, several mild pretreatment techniques are getting developed to reduce the loss in hemicellulose content of lignocellulosic biomass thereby giving attention to more characterization of hemicellulases and notable accessory enzymes beneficial to biomass deconstruction10. Hemicellulases such as xylanases, lichenases and laminarases which are endoglucanases active on mixed-glucans have earlier been reported to improve the hydrolysis of xylan and cellulose thereby contributing to the reduction of enzyme dosage for complete biomass saccharification13,14. Xylanases (EC. 3.2.1.8), are crucial enzymes employed for the hydrolysis of β-1,4-d-xylosidic linkages in the xylan biomass, releasing xylooligosaccharides (XOS) and xylose. This enzyme has been reported to demonstrate wide application in several industrial processes such as in biorefinery, food and biobleaching of paper and pulp. It has been shown that xylanase treatment reduces the content of hexenuronic acid (HexA) in the pulp. HexA is a component formed during chemical pulping from glucuronic acid, which is part of hemicellulose polymers15. Based on the sequence and protein structure, xylanases are classified into many glycosyl hydrolase families (GH) such as 7, 8, 10, 11, 16, 30, 43, 51, 98, and 141, with most of the characterized fungal xylanases being GH 10 and 1116. While there are extensive studies on GH5 xylanases from bacterial hosts, there is a dearth of information on fungal GH5 xylanase till date. Furthermore, due to the presence of β-(1,3)- and β-(1,4)-d-glucans in plant cell walls, the requirements for mixed-linked glucanases such as endo-1,4-β-glucanase (EC 3.2.1.4), endo-1,3(4)-β-glucanase (EC 3.2.1.6), endo-1,3-β-glucanase (EC 3.2.1.39), and endo-1,3–1,4-β-glucanase (EC 3.2.1.73) in cellulolytic enzyme cocktails could also be important for better saccharification performance17.
In an attempt to develop effective technology for utilization of SCB in biorefineries, especially for production of second-generation biofuels, we previously identified the fungus Penicillium funiculosum NCIM1228 as a suitable host for the production of potent lignocellulolytic enzymes for the deconstruction of diverse pretreated cellulosic feedstocks as its secretome exhibited outstanding saccharification potential when compared with secretomes from other fungal strains18,19. Using a combination of both proteomic and biochemical approaches, we identified some relevant accessory enzymes such as xylanases (GH30), endo‑β‑1,3(4)‑glucanase (GH16), β‑1,3‑galactosidase (GH43), cutinase (CE5), and some glycoside hydrolases with unknown functions important for the conversion of SCB to monomeric sugars when the production media for cultivation of Penicillium funiculosum was modulated with SCB14. It was seen that the remarkable induction of these enzymes in the secretome of P. funiculosum facilitated complete saccharification of SCB although at a high enzyme dosage.
Therefore, this study was designed to further improve the quality of enzyme systems in the secretome of P. funiculosum for enhanced saccharification of SCB at low enzyme loading to reduce the overall enzyme requirement for SCB deconstruction. To achieve this, two accessory enzymes, i.e., a GH5 protein with an unknown function and a GH16 endoglucanase, both found to be highly upregulated in the secretome of P. funiculosum in the presence of SCB, were selected for overexpression in P. funiculosum. The unknown function enzyme from P. funiculosum belonging to glycosyl hydrolase family 5 (PfGH5) was first recombinantly expressed in Pichia pastoris and its characteristics were evaluated to decipher its role in biomass hydrolysis. Furthermore, its impact on SCB hydrolysis was investigated and then it was overexpressed along with a GH16 endoglucanase in the background of a catabolite-derepressed strain of P. funiculosum (PfMig188) to produce a more efficient cocktail specific for this biomass. The resulting enzyme cocktail from the engineered strain was then assessed for improved production of hemicellulolytic enzymes as well as for their saccharification efficiency toward SCB with minimal enzyme loading.
Results and discussions
Sequence analysis of an uncharacterized GH5 family protein of P. funiculosum
A unique GH5 family protein was one of the proteins that was found to be highly upregulated in our earlier study in the secretome of P. funiculosum in response to the pre-treated sugarcane bagasse and was implicated in the enhanced hydrolysis of the pretreated bagasse by the secretome14. The sequence analysis of this GH5 family protein of P. funiculosum, PfGH5, showed that this protein has not yet been characterized since the functions of the most similar genes that have been deposited in NCBI as well as in the CAZY database are unknown. As a result, we attempted to characterize and assess the functionality of this gene. The sequence analysis indicated that the PfGH5 contains 1561 nucleotides which encode a chain of 454 amino acid residues with a theoretical molecular weight of 50.1 kDa. Further analysis showed that there were four potential N-glycosylation sites (Asn-Ala-Ser) found in the PfGH5 sequence using the NetNGlyc1.0 server (http://www.cbs.dtu.dk/services/NetNGlyc/), while there was no potential O-glycosylation predicted using the NetOGlyc 4.0 server (http://www.cbs.dtu.dk/services/NetOGlyc/). The PfGH5 amino acid sequence revealed 100% similarity to an uncharacterized glycoside hydrolase family 5 protein from P. occitanis (accession number PCH04120.1), while 99% similarity was found with Talaromyces cellulolyticus (GAM37412.1) and Talaromyces pinophilus (KAF3403216.1) as seen also from the phylogenetic tree (Fig. 1). Surprisingly, the protein also exhibited 82% similarity with a Talaromyces islandicus GH5 protein annotated to be a β-xylosidase suggesting its possibility as a xylan-degrading enzyme. Further analysis using the InterPro scan tool revealed the presence of a conserved domain of cellulase super-family glycoside hydrolase 5 between positions 162–171. The nucleotide sequence of PfGH5 was deposited in the NCBI database with accession number OM160816.

Phylogenetic tree of PfXyn5 orthologs in fungi. Molecular phylogenetic analysis was performed using the maximum likelihood method and the JTT matrix-based model. The tree with the highest log likelihood (− 17,992.93) is shown. Initial trees for the heuristic search were obtained automatically by applying neighbor-joining and BioNJ algorithms to a matrix of pairwise distances estimated using the maximum composite likelihood (MCL) approach and then selecting the topology with a superior log likelihood value. The bootstrap support corresponding to the numbers on the tree branches was calculated per 1000 bootstrap replicates39. This analysis involved the catalytic domains of 18 closely related proteins. Evolutionary analyses were conducted using MEGA X software.
Recombinant expression, purification and functional characteristics of PfGH5 protein
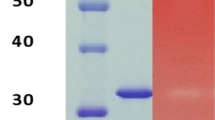
To understand the properties of this PfGH5 protein and its role in biomass deconstruction, the protein was first recombinantly expressed using Pichia pastoris as a host. P. pastoris holds a great advantage as a system for the production of recombinant proteins because of its ability to produce highly efficient enzymes in high abundance as well as for fast growth characteristics in inexpensive media20. In addition, its capability for appropriate folding and transportation of proteins renders it an excellent tool for recombinant protein production20. P. pastoris also secretes fewer native proteins thereby enabling easier purification and biochemical characterization of recombinant proteins in supernatant21,22. To express this protein, the alpha secretion signal peptide of the pPICZαA vector was used to direct the secretion of the protein under the AOX1 promoter. For this, the PfGH5 was fused with the α-secretion signal of the pPICZα vector to obtain the vector pPICZαA-PfGH5, designated as pOAO6. The recombinant pOAO6 vector was linearized with SacI restriction enzyme to enable its integration into the P. pastoris X-33 genome by homologous recombination. Linearized vector was then transformed into competent P. pastoris X-33 for expression and the transformants were selected with Zeocin selection marker (100 μg/ml) for 72 h. Some positive transformants were randomly picked up and re-cultivated on YPDA plates at 30 °C with different zeocin concentrations between 200 and 1000 μg/ml for screening multi-copy positive transformants. A strain with multi-copy integration of the gene that resisted 1000 μg/ml zeocin was further selected, confirmed by PCR (Supplementary Fig. S1) and then inoculated into 50 ml inducing media at 30 °C and 220 rpm for producing the recombinant PfGH5. The induction was monitored every 24 h by loading some of the culture supernatants on SDS-PAGE electrophoresis (Fig. 2a; Supplementary Fig. S2a) and the recombinant enzyme was finally recovered from Pichia pastoris culture supernatant at approximately 1.8 g/L protein after 96 h of induction.

Expression and characterization of recombinant PfXyn5 in P. pastoris (a) SDS-PAGE gel of the supernatant of PfXyn5 produced by P. pastoris over 96 h period of induction. Lanes 1–4 refers to induced cultures of PfXyn5 taken at 24, 48, 72 and 96 h time point while lane 5 refers to wild type culture of P. pastoris X-33 at 96 h time point. (b) SDS-PAGE gel of purified fractions of PfXyn5 by anion exchange chromatography using Q-Sepharose Fast Flow resin. Lane 1 refers to the crude enzyme while lanes 2 and 3 refer to fractions eluted with 25 and 50 mM NaCl gradient. (c) HPLC profile of hydrolysed products of beechwood xylan without PfXyn5 (control). (d) HPLC profile of hydrolysed products of beechwood xylan with PfXyn5. X1, X2, X3, X4, X5 and X6 refers to xylose, xylobiose, xylotriose, xylotetrose, xylopentaose and xylohexaose, respectively. (e) Effects of pH on PfXyn5 activity and stability. The relative activity (%) was calculated relative to the optimum activity obtained at pH 4.5 which was taken as 100%. (f) Effects of temperature on PfXyn5 activity. The relative activity (%) was calculated relative to the optimum activity obtained at 50 °C which was taken as 100% (g) Thermostability profile of PfXyn5. Stability at 40 °C, 50 °C, 60 °C, 70 °C and 80 °C were determined by incubating enzyme at specified temperatures for 300 min while samples were withdrawn every 30 min before assaying for residual activity according to the standard assay procedure.
PfGH5 was purified to apparent homogeneity by a single step of ion-exchange chromatography using Q-Sepharose Fast Flow (QSFF) and the purified enzyme was then used for all subsequent biochemical characterization (Supplementary Fig. S2b). The purified enzyme showed an intense single band on SDS-PAGE with a molecular mass close to 55 kDa (Fig. 2b), which was slightly higher than the theoretical mass of 50 kDa. The difference in sizes suggested that post-translational modifications occurred in PfGH5 during heterologous expression in P. pastoris and these post-translational modifications had effects on the migration rate of the PfGH5 on SDS-PAGE23. To gain insights into the action of this glycoside hydrolase in biomass deconstruction, the enzyme was screened on various polysaccharides, for its specificity on glycosidic bonds (Table 1). The PfGH5 exhibited the highest activity toward beechwood xylan with a specific activity of 0.7 U/mg protein followed by xyloglucan and lichenan. The results also showed that the PfGH5 could also hydrolyse laminarin, carboxymethyl cellulose and para-nitrophenyl-β-d-xylopyranoside (pNPX) although the efficiencies were much lower than with xylan while there was no activity detected on Avicel, β-glucan and para-nitrophenyl-β-d-glucopyranoside (pNPG) (Table 1).
Recognition of a broad range of substrates for catalysis indicated the promiscuous nature of the enzyme. However, the relatively higher specific activity of this enzyme towards both xylan and xyloglucan when compared with other substrates screened suggests the possibility of this PfGH5 be mainly a xylanase. Therefore, to further understand the mechanism of action of the enzyme and the nature of the products it produces, the hydrolysis reactions were set up with 5% xylan as well as with 5% pretreated SCB as the substrate and 25 mg/g protein dosage at 50 °C for 24 h and 72 h, respectively. Analysis of the hydrolysate showed significant production of xylooligosaccharides (XOS) with a degree of polymerization (DP) up to six (DP6) from the xylan in the test reaction containing the enzyme, while there was no trace of XOS in control reactions without enzyme in the reaction mixture after 24 h of incubation (Fig. 2c,d). We observed significant liberation of xylobiose and xylotriose with a very less amount of xylose in the hydrolysate. This mode of action was found to be similar to that of an endoxylanase from Trichoderma asperellum24, thus, suggesting that the PfGH5 is an endo-acting enzyme that randomly cleaves the β-linkages in xylan25. Furthermore, the chromatogram of the hydrolysate for the SCB test condition showed the presence of some glucose released in addition to xylose and XOS by PfGH5 (Supplementary Fig. S3a,b). The detected glucose in the SCB hydrolysates could be from the breakdown of xyloglucan or other mixed-linked glucans that are an integral component of the hemicellulose complex in SCB, as the enzyme showed some activities towards pure xyloglucan as well as mixed-linked glucan substrates (Table 1).
To further characterize the enzyme, the effect of pH on the PfGH5 xylanase activity was examined from pH 3.0 to 6.0. The results showed that the enzyme exhibited maximum activity at pH 4.5, while it had a relative activity of 76% and 60% at pH 4.0 and 5.0, respectively (Fig. 2e). The optimum pH activity observed with this enzyme is in contrast to earlier characterised GH5 xylanases mostly of bacterial origin which exhibited optimum activity between pH 5 and 626,27. The enzyme exhibited good stability at room temperature between pH 3.5–5.5 after 24 h of incubation (Fig. 2e). It was most stable at pH 4.5 and retained about 96% of its original activity after 24 h of incubation at this pH. It was remarkable that the enzyme had a residual activity of 90% at both pH 4.0 and 5.0 after 24 h of incubation (Fig. 2e). The high activity of this enzyme at acidic pH range and stability across a wider pH range indicates that this enzyme could be potentially relevant as most saccharification assays with fungal enzymes are mostly performed at acidic conditions between pH 4 and 528. Evaluation of the temperature optima of this enzyme showed that PfGH5 displayed maximal activity at 50 °C while it exhibited 84% and 73% of its activity at 40 °C and 60 °C, respectively (Fig. 2f). The thermostability of the enzyme was monitored by incubating enzyme at various elevated temperatures for 5 h and then measuring its residual activity. The enzyme was found most stable between 40 and 50 °C where it retained 85% of its original activity at 50 °C after 5 h of incubation (Fig. 2g). At more elevated temperatures (70 and 80 °C), the activity declined drastically (Fig. 2g). The half-lives of PfGH5 at 60, 70 and 80 °C were 192, 45 and 10 min, respectively (Table 2; Supplementary Fig. S4). Significant enzyme activity at elevated temperature is advantageous for industrial biomass processing, as it not only improves substrate solubility and decreases viscosity but also minimizes microbial contaminations29.
Synergistic performance of PfGH5 with cellulases in saccharification of SCB
Since most of the characterized xylanases from the GH10 and GH11 families have been reported as notable accessory enzymes which work synergistically with cellulases in biomass deconstruction30,31, we next evaluated the impact of this new enzyme in hydrolysis of complex heterogeneous biomass together with the secretome from P. funiculosum (PfMig188). For this experiment, pretreated SCB was used as cellulosic substrate. The hydrolysis reaction was set-up at 5% solid loading and a total protein dosage of 10 mg/g dry biomass weight (DBW) that was modulated in different ratios by replacing a portion of the P. funiculosum cellulase cocktail with PfGH5 xylanase for 72 h at 50 °C as shown in Table 3. For the enzyme replacement approach, varying amounts of the PfMig188 cellulases were replaced with PfGH5 xylanase, while the total amount of enzyme added was kept constant on a protein basis at 10 mg/g DBW. Interestingly, the supplementation of the cellulase cocktail with varying amounts of PfGH5 xylanase increased the saccharification potential of P. funiculosum cellulase cocktail synergistically. The results showed substantial boosting effect in hydrolysis with the 80:20 and 90:10 enzyme combination ratio, while the 95:5 ratio showed maximum improvement and the highest degree of synergism (Table 3). The synergistic effect between this accessory enzyme and the cellulase cocktail could be due to the creation of more binding sites as a result of the removal of non-cellulosic polysaccharides by the xylanase for cellulase to act upon32. It was noticed that the total amount of monomeric sugars released with the 95:5 ratio increased by ~ 32% (10.78 g/L) as compared to the 100:0 ratio without the addition of PfGH5 (8.15 g/L), thus indicating that this enzyme holds great biotechnological prospect in improving the saccharification performance of P. funiculosum at reduced cellulase loading.
Vector construction and overexpression of PfXyn5 in catabolite derepressed strain of PfMig188
Upon validation of the function of this GH5 protein to be a xylanase, it was further designated as PfXyn5. Also, from the promising boosting characteristics it exhibits in SCB saccharification when combined with cellulase cocktail, we next decided to overexpress this gene in the background of the Mig1-repressor-deleted strain of P. funiculosum, i.e., PfMig188, as it has been shown to produce a larger amount of cellulolytic enzymes in our earlier study33. To overexpress the xylanase gene in PfMig188, the expression vector containing the endogenous gene along with its promoter and terminator was constructed using the backbone of pBIF binary vector as mentioned in the method section. The 3.6-kb region containing the Xyn5 gene (Fig. 3a) was amplified from the genome of the parent NCIM1228 strain and cloned into the pBIF vector to generate the pOAO7 plasmid. The pOAO7 recombinant plasmid was confirmed by restriction digestion (Supplementary Fig. S5a) before transformation into the PfMig188 strain using the agrobacterium-mediated transformation method (AMTM). The hygromycin-resistant transformants obtained (Fig. 3b) were subsequently analysed by performing PCR to check for the integration of the new expression cassette into the genome of the fungus (Fig. 3c). The amplification of the expected 5 kb fragment in three of the selected transformants (PfOAO6) and found to be absent in the control (PfMig188) confirmed the integration of the Xyn5 cassette into the genome of the fungus.

Construction of the PfXyn5 expression cassette and its overexpression in PfMig188. (a) Schematic diagram showing the assembly of the PfXyn5 cassette from the P. funiculosum NCIM1228 genome. (b) Transformants of pOAO7 after AMTM transformation in PfMig188. Transformants were selected on 100 µg/ml hygromycin. (c) Determination of PfXyn5 integration in the genome of PfMig188 transformants by PCR using the primers PgpdA-F and TrpC-R. The expected size is 5 kbp, corresponding to a fragment spanning the region of gpdA promoter and TrpC on the expression cassette. Lane M is DNA molecular mass marker; lane (1) is the pOAO7 plasmid which served as a positive control while lane (2) is the gDNA of PfMig188 which was the negative control. Lanes 3–7 are the transformants of PfOAO6. (d) The transcriptional expression of xylanase in NCIM1228, PfMig188, and all the three positive strains of PfOAO6 was measured by quantitative real-time PCR after growing the strains for 60 h in the presence of Avicel and SCB. The expression levels were normalized to those in NCIM1228 and plotted. (e) The xylanase, β-xylosidase and FPase activities in the fermentation broth of PfMig188 and three transformants of PfOAO6. (f) Total sugar released from hydrolysis of SCB by secretomes of PfMig188 and three transformants of PfOAO6. (g) Saccharification performance of PfOAO6 secretomes on SCB. Total xylose yield was estimated after 72 h hydrolysis and the hemicellulose conversion were determined and plotted (h) Percentage holocellulose conversion measured at 72 h saccharification period. ** shows the significant difference at α = 0.05 using Tukey’s multiple comparison test.
To understand the effect of increasing the copy number of this enzyme on its expression, the transcript abundance of the transformants was next examined at the mRNA level through quantitative PCR (qPCR). For this, cultures of NCIM1228, PfMig188, and the three positive transformants of PfOAO6 were grown in Mandel’s media containing 25 g/L Avicel and 15 g/L SCB for 60 h to obtain good mycelial growth. The transcript levels of all five strains under derepressing conditions were determined by real-time PCR (RT-PCR) with tubulin as a control, and the relative fold change was normalized to the level in NCIM1228 since it was the original strain from which all other mutant strains (PfMig188 and PfOAO6) were generated (Fig. 3d). The results under the overexpression showed about a 20-fold increase in Xyn5 transcripts for PfMig188, while there were about 35 to 40-fold increases in transcript levels for the PfOAO6 strains. To evaluate the expression level of the xylanase gene in the PfOAO6 transformants, the three transformants with increased transcripts were cultivated in a cellulase-inducing medium (CIM) for 5 days. The transformants were then screened based on the xylanase production capability of their culture supernatant and the results were compared with those for the parent strain, PfMig188 (Fig. 3e). The results showed a significant increase in xylanase activity in the range of 11 to 27%, in all the transformants compared to the parent strain, with the transformant T3 showing maximum xylanase production (Fig. 3e). Furthermore, we investigated if the overexpression of this gene will have an impact on the β-xylosidase content in the secretome since it is the last enzyme in the xylan degradation system that converts xylobiose to xylose. We found a close to ~ 9% increase in the β-xylosidase activity in two of the transformants (T3 and T5) while the β-xylosidase activity of transformants T1 was the same as the control strain (PfMig188). It was observed that these differences could be due to the differences in the integration loci of the expression cassette in the PfMig188 genome when performing random integration as seen in our earlier study34. As expected, we saw that there were no changes in the FPase activity of the secretomes in comparison with that of the parent strain (Fig. 3e), as the GH5 xylanase was found not to be active on a pure cellulosic substrate which is the main substrate in the standard filter paper assay reaction mixture (Table 1).
Since there was a significant improvement in the xylanase content of the transformants’ secretome as seen both from the transcriptional and translational studies, we next evaluated its corresponding effect on the digestion of pretreated SCB (Fig. 3f–h). For this, an SCB hydrolysis reaction was set up with a 15% solid load and minimal protein loading of 2.5 FPU/g DBW at 50 °C for 72 h. A low dose of 2.5 FPU/g was considered to avoid complete saccharification of the SCB, thereby enabling easy identification of the differences in the monomeric sugars released if any in the hydrolysate. The results obtained after hydrolysis showed no difference in the total glucose released from the secretomes of the transformants when compared with the native strain (Fig. 3f), while there was a noticeable improvement in the xylose content after 72 h as expected. The xylose yield with the secretomes of transformants T3 and T5 increased to 17 g/L which corresponded to 94% hemicellulose conversion when compared with the control that yielded 13.6 g/L xylose which corresponded to 76% hemicellulose conversion (Fig. 3g). Overall, the improvement in hemicellulose conversion in the transformants resulted in 44% total holocellulose conversion (Fig. 3h) which was ~ 9% improvement over that of the parent strain. The results indicate that the gene was successfully overexpressed and the integration of the additional copy of this gene in the fungus genome indeed helped in further improving the saccharification efficiency of its secretome in SCB deconstruction.
Overexpression of a GH16 endo‑β‑1,3(4)‑glucanase (PfEgl16) in PfMig188 significantly improved glucose yield from SCB
Since the overexpression of the PfXyn5 gene in PfMig188 from the earlier section improved only the saccharification of the hemicellulose component of SCB with almost no effect on the cellulose conversion, we next focused on overexpressing another gene, a GH16 endo‑β‑1,3(4)‑glucanase (Egl16) with accession number OM483862, which was also significantly upregulated in the presence of SCB from earlier proteomics study14. This enzyme is one of the β-glucanases typically active on complex polysaccharides containing mixed linkages such as β-(1,3)- and β-(1,4)-d-glucans in plant cell wall35. To overexpress the Egl16 gene of P. funiculosum in PfMig188 strain, the Egl16 nucleotide which consists of 1887 bp with one intron and 546 amino acids was constructed using the backbone of the pBIF binary vector along with its native promoter and terminator (Fig. 4a). The gene was amplified from the genome of the parent strain and first cloned into a PCR cloning vector pJET1.2 to generate the pOAO8 plasmid. The PstI/XbaI fragment was excised and cloned into the same sites in pBIF to create the pOAO9 vector. The pOAO9 plasmid was then digested with MunI and MluI restriction enzymes to confirm the presence of the Egl16 gene in the expression cassette and absence in the pBIF vector (Supplementary Fig. S5b). The confirmed pOAO9 plasmid was then transformed into PfMig188 strain using the ATMT procedure, and the hygromycin-resistant transformants (PfOAO7) obtained were analyzed for gene integration and protein expression (Fig. 4b). PCR was performed to confirm the integration of the Egl16 cassette into the genome as shown in Fig. 4c. PCR reactions performed with the primers PgpdA-F and TrpC-R, which were designed based on vector sequence, generated the anticipated amplification products (~ 5 kb fragment) in three out of the five transformants selected, but not in the control PfMig188, which confirmed the integration of the Egl16 cassette into the genome of PfMig188 strain (Fig. 4c).

Construction of the PfEgl16 expression cassette and its overexpression in PfMig188. (a) Schematic diagram showing the assembly of the PfEgl16 cassette from the P. funiculosum NCIM1228 genome. (b) Transformants of pOAO9 after AMTM transformation in PfMig188. Transformants were selected on 100 µg/ml hygromycin. (c) Determination of PfEgl16 integration in the genome of PfMig188 transformants by PCR using the primers PgpdA-F and TrpC-R. The expected size is 5 kbp, corresponding to a fragment spanning the region of gpdA promoter and TrpC on the expression cassette. Lane M is DNA molecular mass marker; lane (1) is the pOAO9 plasmid which served as a positive control while lanes 2–5 are the transformants of PfOAO7. Lane (6) is the gDNA of PfMig188 which was the negative control. (d) The transcriptional expression of endoglucanase in NCIM1228, PfMig188, and all the three positive strains of PfOAO7 was measured by quantitative real-time PCR after growing the strains for 60 h in the presence of Avicel and SCB. The expression levels were normalized to those in NCIM1228 and plotted. (e) The lichenase, laminarase and FPase activities in the fermentation broth of PfMig188 and three transformants of PfOAO7. (f) Saccharification performance of PfOAO7 secretomes on SCB. Total sugar released from hydrolysis of SCB by secretomes of PfMig188 and three transformants of PfOAO7. (g) Percentage holocellulose conversion measured at 72 h saccharification period. ** shows the significant difference at α = 0.05 using Tukey’s multiple comparison test.
For assessment of the expression level of the overexpressed Egl16 gene, cultures of NCIM1228, PfMig188, and the three positive transformants of PfOAO7 were grown in Mandel’s media containing 25 g/L Avicel and 15 g/L SCB as carbon sources for 60 h for mRNA isolation and RT-PCR. The transcript levels of all five strains under derepressing conditions were evaluated with tubulin as a control, and the relative fold change was normalized to the level in NCIM1228 (Fig. 4d). The results showed that the transcript level of Egl16 remained almost unchanged for PfMig188 despite the deletion of the carbon catabolite repressor gene in its genome while there were about threefold increases in transcript levels for two of the PfOAO7 strains. To evaluate the expression level of the endoglucanase gene in the PfOAO7 transformants, the three transformants obtained were cultivated in a cellulase-inducing medium (CIM) for 5 days and the supernatants of the harvested cultures were used for enzyme assays on mixed-linked glucan substrates. The results showed an enhancement in lichenase and laminarase activities in two of the transformants when compared with the parent strain while one of the transformants (T3) showed almost the same activity as that of the parent strain (Fig. 4e). About 26% and 18% increase in laminarase and lichenase activities were found with two of the transformants (T1 and T4) over that of PfMig188. The transformant T4 gave the maximum laminarase and lichenase activities of 9.5 U/ml and 65 U/ml, respectively, while PfMig188 yielded 7.5 U/ml and 54.8 U/ml for laminarase and lichenase, respectively. Evaluation of the impact of overexpressing this gene on the total cellulase activity showed that there were no changes in the FPase activity of the secretomes in comparison with that of the parent strain (Fig. 4e). This result was not surprising though as the overexpressed enzyme is an accessory enzyme that may show little or no activity on a pure cellulosic substrate such as Avicel17.
To evaluate the corresponding impact of overexpressing this gene on the quality of PfMig188 secretome towards SCB hydrolysis, a hydrolysis reaction was set-up with a 15% solid load of SCB and 2.5 FPU/g DBW at 50 °C for 72 h. Upon completion of the hydrolysis, the resulting hydrolysates were analysed accordingly and the results obtained showed a significant increase in the glucose content after 72 h for the PfOAO7 transformants while there was no difference in the total xylose released from the secretomes of the transformants when compared with the native strain (Fig. 4f), The results showed that the concentration of glucose released from SCB increased to 29.6 g/L and 28.5 g/L for transformants T1 and T4, respectively when compared with the control that yielded 26.1 g/L glucose. The approximate 13% increase in glucose yield with the PfOAO7 transformants led to 42.8% cellulose conversion in contrast to PfMig188 which was 37.7% cellulose conversion at this minimum enzyme dosage. Overall, the enhancement in saccharification performance of the PfOAO7 transformants resulted in 46% total holocellulose conversion (Fig. 4g) which was ~ 7% improvement over that of the parent strain (43%). These results, therefore, indicate that the introduction of an extra copy of the Egl16 gene into the genome of the PfMig188 strain enabled increased production of its corresponding enzyme for better hydrolysis on complex heterogeneous substrates such as SCB.
Double overexpression of Pfxyn5 and PfEgl16 results in an enriched accessory enzyme system for better saccharification of SCB residues
In the above sections, we found that individual overexpression of Pfxyn5 and PfEgl16 provided remarkable hydrolysis efficiency against SCB at low enzyme dosage. The results provided an insight that simultaneously overexpressing the two genes may further facilitate enhanced deconstruction of SCB over what was achieved when individually overexpressed. To engineer the fungal strain for the dual overexpression of both Xyn5 and Egl16, a systematic approach was utilized for the construction of the desired pOAO10 vector containing the Xyn5/Egl16 expression cassette (Fig. 5a), as described in “Materials and methods”. For fungal transformation, the pOAO10 plasmid was confirmed by restriction digestion before transformation into the PfMig188 strain (Supplementary Fig. S5c). The hygromycin-resistant transformants (PfOAO8) obtained (Fig. 5b) were analysed by PCR with the primers PgpdA-F and TrpC-R which were designed based on vector sequence. The results showed that all the transformants generated the anticipated amplification products (~ 7 kb fragment) which was absent in the native strain thereby confirming the integration of the Xyn5/Egl16 cassette into the genome of the fungus (Fig. 5c).

Vector construction for the simultaneous overexpression of PfXyn5 and PfEgl16 in PfMig188. (a) Schematic diagram showing the construction cassette for the dual overexpression of the Xyn5 and Egl16 genes from P. funiculosum NCIM1228. Abbreviations: Pro, promoter; SP, signal peptide; Ter, terminator. (b) Transformants of pOAO10 after AMTM transformation in PfMig188. Transformants were selected on 100 µg/ml hygromycin. (c) Determination of integration of the dual cassette (Xyn5/Egl16) in the genome of PfMig188 transformants by PCR using the primers PgpdA-F and TrpC-R. The expected size is 7 kbp, corresponding to a fragment spanning the region of gpdA promoter and TrpC on the expression cassette. Lane M is DNA molecular mass marker; lane (1) is the pOAO10 plasmid which served as a positive control while Lane (2) is the gDNA of PfMig188 which was the negative control. Lanes 3–7 are the transformants of PfOAO8. (d) The transcriptional expression of endoglucanase in NCIM1228, PfMig188, and all the five positive strains of PfOAO8 was measured by quantitative real-time PCR after growing the strains for 60 h in the presence of Avicel and SCB. The expression levels were normalized to those in NCIM1228 and plotted. (e) The xylanase, β-xylosidase, lichenase, laminarase and FPase activities in the fermentation broth of PfMig188 and the five transformants of PfOAO8. (f) Saccharification performance of PfOAO8 secretomes on SCB. Total sugar released from hydrolysis of SCB by secretomes of PfMig188 and three transformants of PfOAO8. (g) Percentage holocellulose conversion measured at 72 h saccharification period. ** shows the significant difference at α = 0.05 using Tukey’s multiple comparison test.
A further evaluation of the transcript level of the overexpressed genes (Xyn5 and Egl16) showed increased xylanase abundance with the PfOAO8 transformants over that of PfMig188 when grown in Mandel’s media containing 25 g/L Avicel and 15 g/L SCB as carbon sources (Fig. 5d). Transformants T1 showed the highest increased Xyn5 transcript which was twofold higher than that of PfMig188 while transformants T3 and T5 both showed a 1.8-fold increment in Xyn5 transcripts (Fig. 5d). For Egl16, the results obtained showed about three to ninefold increases in its transcript level after 60 h with transformants T5 having the highest Egl16 level (Fig. 5d).
To evaluate the expression level of the two overexpressed genes at the translational level, the positive transformants of PfOAO8 were cultivated in a cellulase-inducing medium (CIM) for 5 days and the supernatants of the harvested cultures were used for enzyme assays as stated in “Materials and methods”. Analysis of the results showed significant improvement in enzyme activities across all the transformants when compared with the parent strain which was also in accordance with the mRNA levels (Fig. 5e). The PfOAO8 transformants showed ~ 17% and ~ 7% increase in xylanase and β-xylosidase activities, respectively, compared to PfMig188. Similarly, we found about 43% and 37% increases in lichenase and laminarase activities, respectively when compared to PfMig188 while the FPase activities in all the transformants remained unchanged just as it was observed for the PfOAO6 and PfOAO7 strains.
For the evaluation of the corresponding impact of simultaneously overexpressing these two genes on the quality of PfMig188 secretome towards SCB hydrolysis, another set of hydrolysis reactions was set up with a 15% solid load of SCB and 2.5 FPU/g DBW at 50 °C for 72 h. Upon completion of the hydrolysis, the resulting hydrolysates were analysed accordingly and the results obtained showed a significant increase in both the glucose and xylose contents of the hydrolysates for the PfOAO8 transformants when compared with the native strain (Fig. 5f). The results showed between 11 and 17% increase in the concentration of glucose released for the PfOAO8 transformants when compared with the control. Furthermore, the concentration of xylose in the hydrolysates of the PfOAO8 transformants increased by 11%. The saccharification efficiency (holocellulose conversion) for PfOAO8 strains when analysed was found to be considerably higher than the individual overexpressing strains (PfOAO6 and PfOAO7) as they exhibited between 45 and 49% holocellulose conversion in contrast to the native strain (PfMig188) where ~ 40% holocellulose conversion was achieved (Fig. 5g). These results demonstrated that the Xyn5–Egl16 double overexpression provided a highly significant increase in SCB saccharification efficiency when compared to the individual ones and the engineered strains developed could potentially be used as promising bioresources needed for the production of more balanced cellulase and accessory enzyme cocktails required for the low-cost production of lignocellulose-based biofuels34,36.
In conclusion, an unknown glycosyl hydrolase 5 xylanase from the hypercellulolytic P. funiculosum was recombinantly expressed and characterized and its mechanism of action in the deconstruction of lignocellulose especially SCB was further deciphered in this study. The notable biochemical properties of PfXyn5 suggest it is a suitable candidate for various industrial applications most especially in bioenergy. Using a combined application of various fungal genetic tools, a new fungal strain was also created for simultaneous overexpression of notable accessory enzymes whose secretome demonstrated enhanced saccharification performance on pretreated SCB with minimal protein loading.
Materials and methods
Plasmids and microbial strains
All the strains and plasmids used in this study are listed in Table 4, while all the primer sets used in the study are itemized in Table 5. The nucleotide and protein sequences for Xyn5 and Egl16 for the study have been deposited in the NCBI database with accession numbers OM160816 and OM483862, respectively. Escherichia coli DH5α was used for plasmid propagation throughout the experiments. The Agrobacterium tumefaciens LBA4404 strain used for fungal transformation was maintained on a low-sodium LB medium (10 g/L tryptone, 5 g/L yeast extract, 5 g/L sodium chloride) containing 100 µg/ml kanamycin and 30 µg/ml rifampicin. The pBIF vector which was used as the backbone vector for fungal transformation, contains hygromycin and kanamycin resistance genes as selective markers for the selection of transformants (35). The pJET vector containing the ampicillin resistance gene was used as a shuttle vector for the sub-cloning of the Egl16 gene before re-cloning into pBIF for overexpression. P. funiculosum NCIM1228 and its derivative P. funiculosum Mig188 (PfMig188), the fungal strains used for this study, were routinely cultivated on Petri dishes containing low-malt extract–peptone (LMP) agar for approximately 14 days until full sporulation. For the selection and maintenance of fungal transformants, an LMP medium supplemented with hygromycin B at 100 µg/ml was used.
Cellulosic substrate and pretreatment
NaOH-treated Sugarcane bagasse was used as the cellulosic substrate in this study. The SCB was kindly provided by Natems Sugars Private Limited, Hyderabad, Telangana, India. SCB was washed, dried in a convection oven at 60 °C to constant weight, and then knife-milled using a 1 mm sieve in the mill. The alkaline pretreatment of SCB was done at 90 °C using 0.4 M NaOH at 10% solid loading for 6 h. After pretreatment, the solids were washed three times was then repeatedly washed until the pH became neutral. Compositional analysis conducted according to National Renewable Energy Laboratory (NREL) procedure TP510-4261838 on the pretreated sugarcane bagasse yielded a cellulose content of 46.1%, a hemicellulose content of 24.5% and a lignin content of 17%.
Computational analysis of Xyn5 from P. funiculosum (PfXyn5)
The Xyn5 sequence was retrieved from the draft genome sequence of P. funiculosum NCIM1228 available in our laboratory. The amino acid sequences of some closely similar glycoside hydrolase 5 proteins from other species were retrieved from the NCBI database, and molecular phylogenetic analysis by the maximum likelihood method and the Tamura-Nei model was conducted using MEGA X software39. Multiple-sequence alignment of PfXyn5 with other GH5 proteins obtained from the phylogenetic analysis was performed using the Clustal Omega multiple-sequence alignment program40. The signal peptide and conserved domains were analyzed at Signal P 5.0 server (http://www.cbs. dtu.dk/services/SignalP/) and ScanProsite (http://www.expasy.ch/tools/ScanProsite), respectively. N-Glycosylation and O-glycosylation sites were predicted using NetNGlyc 1.0 (http://www.cbs.dtu.dk/services/NetNGlyc/) and NetOGlyc 4.0 server (http://www.cbs.dtu.dk/services/NetOGlyc/), respectively.
Construction of PfXyn5 expression vector for recombinant expression and purification in P. pastoris
The xylanase sequence used in the present study from P. funiculosum was synthesized by GenScript USA, Inc. (New Jersey, USA) with codon optimization for expression in P. pastoris. The synthesized sequence was ligated into the vector pPICZαA, named pPICZαA-PfGH5 (or pOAO6). The pOAO6 vector was transformed into Escherichia coli (DH5α), and subsequently, the positive clone was screened, and the plasmid was extracted using Qiagen mini-prep kit. The pOAO6 plasmid was linearized using SacI restriction endonuclease and transformed into P. pastoris X-33 competent cells using the lithium chloride method described by Kumar et al.41. The transformation liquid was plated on a YPD plate containing 100 µg/ml zeocin, and the positive transformants were further reselected on a higher concentration of zeocin (1000 µg/ml) for 48 h before being screened using polymerase chain reaction (PCR) with the AOXI-F and AOX1-R primer pairs (Table 5). The selected positive transformants were subsequently inoculated into buffered glycerol-complex medium (BMGY) for 24 h before transferring into methanol-complex medium (BMMY) for xylanase expression with the addition of 1% methanol every 12 h till the end of the fermentation process. The induction of the expressed protein was monitored by collecting samples from the cultures every 24 h and loaded on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).
To purify the protein, the culture supernatants were recovered by pelleting the cells by centrifugation at 4000 rpm for 5 min, 4 °C and filtered on 0.45-µm filters (Millipore, Molsheim, France) to remove any remaining cells. The recovered supernatant was dialyzed against 50 mM sodium phosphate buffer, pH 7.5 overnight with two equal changes of buffer at 12 h intervals until the pH becomes 7.5. After adjusting the pH to 7.5, the supernatants were filtered once more on 0.2-µm filters and loaded onto a FPLC HiTrap Q Sepharose XL 5 ml column (GE Healthcare) that was equilibrated with 50 mM sodium phosphate buffer (pH 7.5). Proteins were eluted using a linear gradient of NaCl (25–1000 mM) in the same buffer mentioned above at flow rate of 2.0 ml/min. Fractions containing the enzyme activity were pooled and subjected to SDS-PAGE to check protein purity. Protein concentrations were determined using the bicinchoninic acid (BCA) method using bovine serum albumin (BSA) as standard14.
Biochemical characterization of recombinant PfXyn5
To characterize the recombinant PfXyn5, the protein was screened on various polysaccharides which include Avicel, beechwood xylan, xyloglucan, laminarin, sodium carboxymethyl cellulose, lichenan, β-glucan, p-nitrophenyl-β-d-glucopyranoside (pNPG) and p-nitrophenyl-β-d-xylopyranoside (pNPX). The substrates were incubated with the appropriate amount of the enzyme at 50 °C for 30 min and the amount of reducing sugar liberated was quantitated using the DNS method where appropriate. For determination of the mechanism of action of the enzyme, hydrolysis reactions were set up using 5% xylan and 5% pretreated SCB as substrates and protein dosage of 25 mg/g DBW at 50 °C for 24 h and 72 h, respectively. The resulting hydrolysates obtained were analysed using an Agilent 1260 series HPLC instrument coupled with a 1290 Infinity evaporative light scattering detector (ELSD). The degradation products were estimated using a Rezex RSO-oligosaccharide Ag+ (4%) column (200 by 10 mm) with a Rezex RSO-oligosaccharide Ag+ (4%) guard column (60 by 10 mm) (Phenomenex), with a mobile phase of 100% water. Throughout the analysis, the column was kept at 80 °C, and the mobile phase flow rate was maintained at 0.3 ml/min under isocratic conditions for 60 min. The chromatographic run was initiated by first equilibrating the column for 5 min followed by injecting 20 µl of the sample solution into the column. The ELSD was maintained at 45 °C throughout. The nebulizer (nitrogen) gas pressure was set at 2.8 × 105 Pa, and the detector gain was set at 9 × 105 Pa. The concentration of different XOS generated was calculated from calibration curves of external standards purchased from Megazyme. The pH dependence of PfXyn5 was determined by using 50 mM citrate–phosphate buffer (pH 3.0–6.0) containing purified enzyme and xylan at 50 °C as described above. To determine the effect of pH on the stability of PfXyn5, the enzyme was incubated in relevant buffers of varying pH (3.0–6.0) without substrate for 24 h at room temperature. The residual xylanase activity was determined according to the standard assay procedure earlier described. The temperature dependence was determined by incubating the reaction with the optimal pH at 40–80 °C. To determine the dependence on pH and temperature, the maximum activity obtained was defined as 100%. The thermal stability of PfXyn5 was analysed by preincubating it at 40–80 °C for 5 h and its residual activity was then determined.
Effect of PfXyn5 xylanase supplementation on the performance of secretome from the P. funiculosum (PfMig188) during biomass hydrolysis
The effect of supplementing the cellulase cocktail present in the secretome of P. funiculosum with PfXyn5 in SCB saccharification was determined by setting up hydrolysis reactions with pretreated 5% solid loading and a total protein dosage of 10 mg/g DBW at 50 °C for 72 h with constant shaking. The 10 mg/g protein concentration was maintained at different combination ratios of 60:40, 70:30, 80:20, 90:10, 95:05 and 100:0 for P. funiculosum to PfXyn5. After hydrolysis the samples were centrifuged to separate the solid residue from the digested biomass. Supernatants that were recovered after enzymatic hydrolysis of the pretreated SCB were analysed by high-performance liquid chromatography with an Aminex HPX-87H anion-exchange column (Bio-Rad, USA) and a refractive index (RI) detector to analyse the released monosaccharides (glucose and xylose) by anion-exchange chromatography. The concentration of each monosaccharide was calculated from calibration curves of external standards (xylose and glucose) purchased from Absolute Standards Inc. The degree of synergism between the PfXyn5 and cellulase was calculated following the method described by Goncalves et al. and Li et al.13,32 with the formula:
where α and β correspond to the mass ratio of the enzymes in each reaction. Y1+2, indicates the yield of reducing sugar achieved by the two enzymes working simultaneously, and Y1 and Y2 indicate the yields of reducing sugar, achieved by each enzyme working separately.
Engineering of P. funiculosum Mig188 for overexpression of Xyn5 and Egl16
All vectors for fungal expression described in this study were constructed on the backbone of the pBIF vector that was previously constructed using the binary vector pCAMBIA1300 as a backbone37. Binary vectors for the overexpression of the Xyn5 and Egl16 genes from P. funiculosum were constructed as follows. The endogenous gene encoding Xyn5 was amplified from the genome of P. funiculosum NCIM1228 using the primers PfXyn5-F and PfXyn5-R (Table 5). The primers were designed according to the Xyn5 sequence containing both its native promoter and terminator, which was obtained from the draft genome sequence of the strain. The PCR product obtained was digested with the restriction enzymes PstI and BamHI before being ligated into the pBIF vector previously digested with the same enzyme sets to generate the pBIF-Xyn5 (pOAO7) vector. For the overexpression of Egl16, the Egl16 gene was amplified from the genome of P. funiculosum NCIM1228 using the primers PfEgl16-F1 and PfEgl16-R1 spanning its native promoter and terminator (Table 5). The PCR product was first subcloned into pJET1.2 to obtain the pOAO8 vector. The PstI/XbaI fragment containing the Egl16 fragment was excised and cloned into the same sites in pBIF to create the pBIF-Egl16 vector (pOAO9) vector. The ligated products of pOAO7 and pOAO9 were then transformed into E. coli DH5α cells and selected on 50 µg/ml kanamycin. The resulting colonies were screened for positive transformants by colony PCR, followed by restriction digestion of the corresponding plasmids. To create a strain for the combined overexpression of both the Xyn5 and Egl16 genes, the pOAO7 vector was used as the base vector upon which the Egl16 gene was cloned. The Egl16 gene with its native promoter and terminator was re-amplified from the genome of P. funiculosum NCIM1228 using the primers PfEgl16-F2 and PfEgl16-R2 containing the ApaI and BamHI restriction sites, respectively. The PCR product obtained was digested with the ApaI and BamHI restriction enzymes before being ligated into the corresponding sites of the pOAO7 vector to obtain the pBIF-Xyn5/Egl16 (pOAO10) vector. The verified pOAO7, pOAO9, and pOAO10 plasmids were transformed into the PfMig188 fungal strain according to the agrobacterium-mediated transformation method (AMTM) as previously described42. After transformation, the resulting hygromycin-resistant transformants were screened according to the method described previously by Fang and Xia43. The transformants were verified by PCR for the integration of the Xyn5 and Egl16 gene expression cassettes. For PCR analysis, the primers PgpdA-F and TrpC-R were used to screen for Xyn5 and Egl16 integration, while transformants of Xyn5/Egl16 were screened using the primer set PgpdA IR-F and TrpC-R (Table 5).
Expression analysis of cellulolytic enzymes via real-time PCR
For real-time PCR experiments, cultures of all strains were grown in minimal Mandel’s medium containing 25 g/L Avicel and 15 g/L SCB for 60 h44. Mycelia were harvested by filtration and frozen in liquid nitrogen. RNA was extracted using an RNeasy kit (Qiagen) according to the manufacturer’s instructions. RNA was treated with DNase (Invitrogen) before cDNA synthesis. One microgram of RNA was used as the template for each quantitative real-time PCR (qRT-PCR). A cDNA synthesis control was performed to ensure the absence of DNA contamination. qRT-PCR was carried out using iTaq universal SYBR green supermix (Bio-Rad) and a Bio-Rad CFX96 qPCR detection system. Primers for transcripts to be tested were designed using the boundary sequence of two exons to avoid any amplification from contaminant genomic DNA. qRT-PCR was performed in biological triplicates with tubulin as the endogenous control. Relative expression levels were normalized to the level of tubulin, and the fold changes in RNA levels were calculated as the ratios of the relative expression levels in PfMig188 and the corresponding transformants of Xyn5 and Egl16 to that in NCIM1228 under cellulase-inducing conditions45.
Cellulolytic secretome preparation
Penicillium funiculosum NCIM1228, PfMig188, and the resulting PfOAO6, PfOAO7, and PfOAO8 transformants were cultivated on Petri dishes containing low-malt extract agar until full sporulation. After 14 days of incubation, spores were recovered with sterile water, filtered through sterile Mira cloth, and quantified using a hemocytometer. The primary culture of each strain was prepared by culturing 107 conidiophores in potato dextrose broth (PDB) for 36 h. Primary cultures of the strains were added to a cellulase-inducing medium in Erlenmeyer flasks at a final concentration of 10%. Cellulase-inducing medium (CIM) contained soya peptone (24 g/L), wheat bran (14 g/L), microcrystalline cellulose (MCC) (16 g/L), pretreated SCB (15 g/L), KH2PO4 (12.4 g/L), K2HPO4 (2.68 g/L), (NH4)2SO4 (0.28 g/L), CaCO3 (2.5 g/L), corn steep liquor (1%), urea (0.52 g/L), and yeast extract (0.05 g/L), with the final pH adjusted to 5.0. The flasks were kept at 28 °C for 5 days with orbital shaking at 150 rpm (Innova 44; Eppendorf AG, Germany). Induced cultures were centrifuged at 9000 rpm for 10 min at 4 °C, and the cellulolytic supernatants were collected and stored at 4 °C until use.
Determination of the enzyme activities of the engineered secretome
All enzymatic activities performed in this study were routinely determined following standard assay procedures. Endoglucanase, xylanase, lichenase, laminarase, β-glucosidase and β-xylosidase activities were determined by incubating appropriate dilution of the enzyme with 2% CMC (Sigma), 2% beechwood xylan (HiMedia), 1% lichenan (Megazyme), 1% laminarin (Sigma) and p-nitrophenyl-β-d-xylopyranoside (Megazyme), respectively, for 30 min, after which the amount of reducing sugars released was measured as previously reported12. One unit of CMCase, xylanase, lichenase and laminarase activity is defined as the amount of enzyme releasing 1 µmol of reducing sugar per min while one unit of β-xylosidase activities was defined as the amount of protein that released 1 μmol of p-nitrophenol (pNP) per min. Total cellulase activity in the secretome was measured in terms of filter paper units (FPU) per milliliter of original (undiluted) enzyme solution. The assay requires a fixed degree of conversion of the substrate, from 50 mg of filter paper within 60 min at 50 °C. One FPU is defined as the amount of enzyme required to produce 2 mg of glucose from 50 mg of filter paper within 60 min of incubation46. To quantitate the amount of secreted proteins in the various PfMig188 secretomes as well as its derivatives, the crude secretomes were first buffer exchanged with the help of a 10-kDa cut-off membrane using citrate– phosphate buffer (pH 4.0) and then the total protein of each secretome was estimated by the bicinchoninic acid (BCA) method using bovine serum albumin (BSA) as standard.
Biomass saccharification and quantification of fermentable sugars
The saccharification efficiency of the secretomes of all the strains used in the study with pretreated SCB was determined according to the method described previously by Ogunyewo et al.34, with some modifications. The performance of the secretomes toward NaOH-treated SCB was evaluated at 15% dry weight of biomass using an enzyme concentration of 2.5 FPU/g DBW. Saccharification was performed in 50 ml screw-cap Falcon tubes in an incubator shaker at 50 °C for 72 h. The reaction mixture included NaOH-treated SCB under 15% dry weight loading in a 5 ml final reaction volume. The total cellulase activities of each secretome of all the fungal strains tested were measured, and the appropriate volume of the desired protein concentration (2.5 FPU/g DBW) was added to the reaction mixture. The reactions were set up in 50 mM citrate phosphate buffer (pH 4.0), and the mixtures were incubated at 50 °C with constant shaking at 220 rpm for 72 h. Samples were collected at the end of the 72 h incubation period and analysed for the production of fermentable sugars. Control experiments were carried out under the same conditions, using substrates without enzymes (enzyme blank) and enzymes without substrates (substrate blank); a substrate-free negative control was set up by filling the Falcon tubes with 50 mM citrate phosphate buffer (pH 4.0), and the background of soluble sugars present in the respective biomass was determined by incubating each biomass in the absence of the enzyme. Following the completion of hydrolysis at each time point, the Falcon tubes were centrifuged at 3500 rpm for 10 min in a swinging-bucket centrifuge (Eppendorf, Germany) to separate the solid residue from the digested biomass. Supernatants that were recovered after enzymatic hydrolysis of the pretreated SCB were analysed by high-performance liquid chromatography with an Aminex HPX-87H anion-exchange column (Bio-Rad, USA) and a refractive index (RI) detector to analysed the released monosaccharides (glucose and xylose) by anion-exchange chromatography. The filtered mobile phase (4 mM H2SO4) was used at a constant rate of 0.3 ml/min with the column, and the RI detector temperature was maintained at 35 °C. The concentration of each monosaccharide was calculated from calibration curves of external standards (xylose and glucose) purchased from Absolute Standards Inc. The theoretical conversions of cellulose and hemicellulose (in percentage) into monomeric sugars were calculated using the equations provided in NREL’s LAP TP-510-43630 as earlier reported19.
Data and statistical analysis
All experiments were performed in triplicate, and the results are presented as the means and standard deviations. The data were compiled in a Microsoft Excel spreadsheet, where the averages and standard errors of the means were determined. All graphs were created using GraphPad Prism 8.0 software. The data were further evaluated by one-way analysis of variance (ANOVA) and multiple t-tests using GraphPad Prism 8.0 software where appropriate.
Data availability
The nucleotide and protein sequences for the PfXyn5 and PfEgl16 genes used for this study have been deposited to the NCBI database with accession numbers OM160816 and OM483862, respectively.
References
Armstrong, Z., Mewis, K., Strachan, C. & Hallam, S. J. Biocatalysts for biomass deconstruction from environmental genomics. Curr. Opin. Chem. Biol. 29, 18–25 (2015).
Chundawat, S. P. S., Beckham, G. T., Himmel, M. E. & Dale, B. E. Deconstruction of lignocellulosic biomass to fuels and chemicals. Annu. Rev. Chem. Biomol. Eng. 2, 121–145 (2011).
Gupta, A. & Verma, J. P. Sustainable bio-ethanol production from agro-residues: A review. Renew. Sustain. Energy Rev. 41, 550–567 (2015).
Rastogi, M. & Shrivastava, S. Recent advances in second generation bioethanol production: An insight to pretreatment, saccharification and fermentation processes. Renew. Sustain. Energy Rev. 80, 330–340 (2017).
Steindorff, A. S. et al. Insights into the lignocellulose-degrading enzyme system of Humicola grisea var. thermoidea based on genome and transcriptome analysis. Microbiol. Spectr. 9, e01088-21 (2021).
Ahmadi, F., Zamiri, M. J., Khorvash, M., Ziaee, E. & Polikarpov, I. Pre-treatment of sugarcane bagasse with a combination of sodium hydroxide and lime for improving the ruminal degradability: Optimization of process parameters using response surface methodology. J. Appl. Anim. Res. 44, 287–296 (2015).
Xu, F., Zhong, X. C., Sun, R. C. & Lu, Q. Anatomy, ultrastructure and lignin distribution in cell wall of Caragana Korshinskii. Ind. Crops Prod. 24, 186–193 (2006).
Jafari, V., Labafzadeh, S. R., Jeihanipour, A., Karimi, K. & Taherzadeh, M. J. Construction and demolition lignocellulosic wastes to bioethanol. Renew. Energy 36, 2771–2775 (2011).
Junior, W. G. M. et al. Sugarcane bagasse saccharification by enzymatic hydrolysis using endocellulase and β-glucosidase immobilized on different supports. Catalysts 11, 340 (2021).
de Oliveira Gorgulho Silva, C. et al. Mild hydrothermal pretreatment of sugarcane bagasse enhances the production of holocellulases by Aspergillus niger. J. Ind. Microbiol. Biotechnol. 46, 1517–1529 (2019).
Sheng, Y. et al. Enzymatic conversion of pretreated lignocellulosic biomass: A review on influence of structural changes of lignin. Bioresour. Technol. 324, 124631 (2021).
Zhao, X., Song, Y. & Liu, D. Enzymatic hydrolysis and simultaneous saccharification and fermentation of alkali/peracetic acid-pretreated sugarcane bagasse for ethanol and 2,3-butanediol production. Enzyme Microb. Technol. 49, 413–419 (2011).
Gonçalves, G. A. L. et al. Synergistic effect and application of xylanases as accessory enzymes to enhance the hydrolysis of pretreated bagasse. Enzyme Microb. Technol. 72, 16–24 (2015).
Ogunyewo, O. A. et al. Accessory enzymes of hypercellulolytic Penicillium funiculosum facilitate complete saccharification of sugarcane bagasse. Biotechnol. Biofuels 14, 1–17 (2021).
Immerzeel, P. & Fiskari, J. Synergism of enzymes in chemical pulp bleaching from an industrial point of view: A critical review. Can. J. Chem. Eng. https://doi.org/10.1002/cjce.24374 (2022).
Joshi, N., Sharma, M. & Singh, S. P. Characterization of a novel xylanase from an extreme temperature hot spring metagenome for xylooligosaccharide production. Appl. Microbiol. Biotechnol. 104, 4889–4901 (2020).
Li, J. et al. Overexpression and characterization of a novel endo-β-1,3(4)-glucanase from thermophilic fungus Humicola insolens Y1. Protein Expr. Purif. 138, 63–68 (2017).
Ogunmolu, F. E. et al. Proteomics insights into the biomass hydrolysis potentials of a hypercellulolytic fungus Penicillium funiculosum. J. Proteome Res. 14, 4342–4358 (2015).
Ogunyewo, O. A. et al. Engineered Penicillium funiculosum produces potent lignocellulolytic enzymes for saccharification of various pretreated biomasses. Process Biochem. 92, 49–60 (2020).
Katsimpouras, C., Dedes, G., Thomaidis, N. S. & Topakas, E. A novel fungal GH30 xylanase with xylobiohydrolase auxiliary activity. Biotechnol. Biofuels 12, 120 (2019).
Miao, T. et al. Improved production of xylanase in Pichia pastoris and its application in xylose production from xylan. Front. Bioeng. Biotechnol. 9, 780 (2021).
Kao, M.-R., Yu, S.-M. & Ho, T.-H. Improvements of the productivity and saccharification efficiency of the cellulolytic β-glucosidase D2-BGL in Pichia pastoris via directed evolution. Biotechnol. Biofuels 14, 126 (2021).
Bernardi, A. V., Gerolamo, L. E., Uyemura, S. A. & Dinamarco, T. M. A thermophilic, pH-tolerant, and highly active GH10 xylanase from Aspergillus fumigatus boosted pre-treated sugarcane bagasse saccharification by cellulases. Ind. Crops Prod. 170, 113697 (2021).
Zheng, F. et al. An endoxylanase rapidly hydrolyzes xylan into major product xylobiose via transglycosylation of xylose to xylotriose or xylotetraose. Carbohydr. Polym. 237, 116121 (2020).
Basit, A. et al. Highly efficient degradation of xylan into xylose by a single enzyme. ACS Sustain. Chem. Eng. 7, 11360–11368 (2019).
Hurlbert, J. C. & Preston, J. F. Functional characterization of a novel xylanase from a corn strain of Erwinia chrysanthemi. J. Bacteriol. 183, 2093–2100 (2001).
Gallardo, Ó. et al. Characterization of a family GH5 xylanase with activity on neutral oligosaccharides and evaluation as a pulp bleaching aid. Appl. Environ. Microbiol. 76, 6290–6294 (2010).
Xie, J. et al. A novel thermophilic β-mannanase with broad-range pH stability from Lichtheimia ramosa and its synergistic effect with α-galactosidase on hydrolyzing palm kernel meal. Process Biochem. 88, 51–59 (2020).
Yu, T. et al. Hyperthermostable Thermotoga maritima xylanase XYN10B shows high activity at high temperatures in the presence of biomass-dissolving hydrophilic ionic liquids. Extremophiles 20, 515–524 (2016).
Hu, J., Arantes, V. & Saddler, J. N. The enhancement of enzymatic hydrolysis of lignocellulosic substrates by the addition of accessory enzymes such as xylanase: Is it an additive or synergistic effect?. Biotechnol. Biofuels 4, 1–14 (2011).
Sun, F. F. et al. Accessory enzymes influence cellulase hydrolysis of the model substrate and the realistic lignocellulosic biomass. Enzyme Microb. Technol. 79–80, 42–48 (2015).
Li, J. et al. Synergism of cellulase, xylanase, and pectinase on hydrolyzing sugarcane bagasse resulting from different pretreatment technologies. Bioresour. Technol. 155, 258–265 (2014).
Randhawa, A., Ogunyewo, O. A., Eqbal, D., Gupta, M. & Yazdani, S. S. Disruption of zinc finger DNA binding domain in catabolite repressor Mig1 increases growth rate, hyphal branching, and cellulase expression in hypercellulolytic fungus Penicillium funiculosum NCIM1228. Biotechnol. Biofuels 11, 15 (2018).
Ogunyewo, O. A. et al. Synergistic action of a lytic polysaccharide monooxygenase and a cellobiohydrolase from Penicillium funiculosum in cellulose saccharification under high-level substrate loading. Appl. Environ. Microbiol. 86, 1–21 (2020).
Chen, X. et al. High-level expression of a novel Penicillium endo-1,3(4)-β-d-glucanase with high specific activity in Pichia pastoris. J. Ind. Microbiol. Biotechnol. 39, 869–876 (2012).
Qian, Y. et al. Production of highly efficient cellulase mixtures by genetically exploiting the potentials of Trichoderma reesei endogenous cellulases for hydrolysis of corncob residues. Microb. Cell Fact. 16, 207 (2017).
Nizam, S., Singh, K. & Verma, P. K. Expression of the fluorescent proteins DsRed and EGFP to visualize early events of colonization of the chickpea blight fungus Ascochyta rabiei. Curr. Genet. 56, 391–399 (2010).
Sluiter, A. et al. Determination of structural carbohydrates and lignin in biomass: Laboratory Analytical Procedure (LAP) (Revised July 2011). http://www.nrel.gov/biomass/analytical_procedures.html (2008).
Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549 (2018).
Madeira, F. et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 47, W636–W641 (2019).
Kumar, S., Mannil, A. & Mutturi, S. Modified chemical method for efficient transformation and diagnosis in Pichia pastoris. Protein Expr. Purif. 174, 105685 (2020).
Michielse, C. B., Hooykaas, P. J. J., van den Hondel, C. A. M. J. J. & Ram, A. F. J. Agrobacterium-mediated transformation of the filamentous fungus Aspergillus awamori. Nat. Protoc. 3, 1671–1678 (2008).
Fang, H. & Xia, L. High activity cellulase production by recombinant Trichoderma reesei ZU-02 with the enhanced cellobiohydrolase production. Bioresour. Technol. 144, 693–697 (2013).
Mandels, M. & Reese, E. T. Induction of cellulase in Trichoderma viride as influenced by carbon sources and metals. J. Bacteriol. 73, 269–278 (1957).
Kitchen, R. R., Kubista, M. & Tichopad, A. Statistical aspects of quantitative real-time PCR experiment design. Methods 50, 231–236 (2010).
Ghose, T. K. Measurement of cellulase activities. Pure Appl. Chem. 59, 257–268 (1987).
Acknowledgements
This study was funded by the Department of Biotechnology, Government of India via Bioenergy Centre Grants BT/IN/Indo-UK/SSY/07/2018-19 & BT/PR/Center/03/2011-Phase II. OAO acknowledges kind financial support via Arturo Falaschi Pre-Doctoral Fellowship from the ICGEB. The authors would like to thank Dr. Leonardo D. Gómez for providing the pretreated sugarcane bagasse used for this study and Mr Raubins Kumar for the technical support during enzyme purification.
Author information
Authors and Affiliations
Contributions
O.A.O. and S.S.Y. conceptualized the idea, designed and coordinated the study, O.A.O. and O.E.O. performed fungal genetic modification experiments and biochemical characterization. O.A.O. and S.K. performed enzyme expression experiments in Pichia pastoris. S.S.Y. supervised and generated fund for the study. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ogunyewo, O.A., Okereke, O.E., Kumar, S. et al. Characterization of a GH5 endoxylanase from Penicillium funiculosum and its synergism with GH16 endo-1,3(4)-glucanase in saccharification of sugarcane bagasse. Sci Rep 12, 17219 (2022). https://doi.org/10.1038/s41598-022-21529-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-21529-1
This article is cited by
-
Identification and Characterization of Two Novel Extracellular β-Glucanases from Chaetomium globosum against Fusarium sporotrichioides
Applied Biochemistry and Biotechnology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
