
Abstract
Drug-induced liver injury (DILI) is the leading cause of acute liver failure (ALF). Continuous and prolonged hepatic cellular oxidative stress and liver inflammatory stimuli are key signatures of DILI. DEAD-box helicase 3, X-linked (DDX3X) is a central regulator in pro-survival stress granule (SG) assembly in response to stress signals. However, the role of DDX3X in DILI remains unknown. Herein, we characterized the hepatocyte-specific role of DDX3X in DILI. Human liver tissues of DILI patients and control subjects were used to evaluate DDX3X expression. APAP, CCl4 and TAA models of DILI were established and compared between hepatocyte-specific DDX3X knockout (DDX3XΔhep) and wild-type control (DDX3Xfl/fl) mice. Hepatic expression of DDX3X was significantly decreased in the pathogenesis of DILI compared with controls in human and mice. Compared to DDX3Xfl/fl mice, DDX3XΔhep mice developed significant liver injury in multiple DILI models. DDX3X deficiency aggravates APAP induced oxidative stress and hepatocyte death by affecting the pro-survival stress granule (SG) assembly. Moreover, DDX3X deficiency induces inflammatory responses and causes pronounced macrophage infiltration. The use of targeted DDX3X drug maybe promising for the treatment of DILI in human.
Similar content being viewed by others
Introduction
The liver is a major organ for drug metabolism and elimination [1]. Drug-induced liver injury (DILI) has become a leading cause of acute liver failure (ALF) and transplantation [2]. Acetaminophen (APAP) is the most well-studies drug which overdose induces hepatotoxicity and ALF [3, 4]. APAP is metabolized by cytochrome P450 isoforms into N-acetylbenzoquinoneimine (NAPQI) [5]. Excess NAPQI formation due to APAP overdose overwhelms the antioxidant glutathione (GSH) reserves of liver, causing accumulating of cytotoxic adducts and subsequent hepatotoxicity [6, 7]. Apart from APAP, established mice models also utilizes carbon tetrachloride (CCl4) or thioacetamide (TAA) to interpret the key aspects of DILI [7, 8]. Regardless of the etiology, profound hepatic cellular oxidative stress and persistent liver inflammation are key signatures of DILI, which aggravating hepatocyte injury and hepatocellular cell death [9,10,11]. However, the key molecular mechanisms that interprets these stress responses in the pathogenesis of DILI remains largely unknown.
The DEAD-box helicase 3, X-linked (DDX3X), a DEAD-box family member and helicase, has been implicated in human health and disease, regulates viral replication, innate immune response, and translational reprogramming [12,13,14]. DDX3X is a key molecule govern cell fate when exposed to stress stimuli by controlling the formation of pro-survival stress granule (SG) and pro-death NLRP3 activation [15, 16]. SGs are cytoplasmic messenger ribonucleoprotein (mRNP) aggregates that assemble during cellular stress (such as hypoxia, heatshock and oxidative stress) [17, 18]. Evidence of the DDX3X interacts with SG in hepatitis C virus infection has been documented [19], suggesting that DDX3X-mediated SG formation may be important for liver pathogenesis. However, the possible role of DDX3X in DILI is still unknown.
In this study, we used conditional hepatocyte DDX3X knockout mice (DDX3XΔhep) and DDX3X control mice (DDX3Xfl/fl) to explore the hepatocyte-specific role of DDX3X in DILI. Our results demonstrated that downregulation of DDX3X is found in human and multiple mice models of DILI. DDX3X deficiency impairs the capacity of SG assembly, leading to persistent and profound hepatic oxidative stress, cell death and subsequent inflammation. Therefore, targeting DDX3X may provide a novel strategy for treating drug-induced hepatotoxicity.
Materials and methods
Human liver tissues
Human DILI liver tissues (n = 16) were obtained from percutaneous liver biopsy from the Nanjing Drum Tower Hospital, Affiliated Hospital of Medical School, Nanjing University. Control liver samples (n = 16) originated from patients with hepatic hemangioma. Histological assessments were determined by two pathologists in a double-blind manner. All patients provided written informed consent. The study was approved by the Institutional Ethics Committee of Nanjing Drum Tower Hospital.
Mice and treatments
All animal studies were performed in accordance with the guidelines of the Institutional Animal Use and the Animal Experimentation Ethics Committee at the Northwest University. Mice were maintained in ventilated cages under 12 h light/dark cycles. The DDX3Xfl/fl mice (C57BL/6 strain) was generated as previously described [15]. DDX3X∆hep mice were established by crossing DDX3Xfl/fl mice with Alb-cre mice (C57BL/6 strain). Sex-matched mice at 8 weeks of age were used for all the experiments. To generate drug induced liver injury, DDX3X∆hep mice and DDX3Xfl/fl littermates were fasted overnight before intraperitoneally injected with APAP (350 mg/kg) or CCL4 (1 ml/kg) or TAA (300 mg/kg) as previously report [7, 8]. Serum and liver tissues were collected.
Statistical analysis
All experiments were repeated at least three times with consistent results. Statistical analysis was performed using the statistical software package SPSS 13.0 (SPSS, Chicago, IL, USA) and quantitative data were expressed as the Median (25th, 75th percentiles) or mean ± SD. All tests of significance were two-sided and p-values < 0.05 were considered statistically significant.
Additional material and methods are reported in the Supplementary materials.
Results
DDX3X expression is downregulated in human and mice with drug-induced liver injury (DILI)
To elucidate the role of DDX3X in DILI, we examined DDX3X expression in human liver sections with DILI. All patients with DILI gained higher serum levels of alanine aminotransferase (ALT) and aspartate transaminase (AST) compared with controls (Fig. 1A and Supplementary Table. S1). Immunohistochemical (IHC) staining revealed that DDX3X is widely expressed in hepatocytes and non-parenchymal cells of liver with control subjects, and hepatic expression of DDX3X was significantly decreased in the pathogenesis of DILI (Fig. 1B). Consistent with human data, downregulation of DDX3X was also found in multiple mice models of DILI (Fig. 1C–E), suggesting that DDX3X downregulation is involved in the pathogenesis of DILI in human and mice.

A Serum levels of ALT of control subjects and patients with DILI. Data are expressed as the Median (25th, 75th percentiles), ***p < 0.001. B Representative H&E and IHC sections of human liver tissues. Scale bars, 10 µm; C–E mRNA and protein expression of DDX3X in multiple DILI mouse models treated with APAP, CCL4, or TAA. Data are expressed as mean ± SD, n = 3-6/group. *p < 0.05, **p < 0.01.
Hepatocyte DDX3X protects against APAP induced liver injury
Given that hepatocyte is the major cell type involved in DILI, we investigated the role of hepatocyte DDX3X in the development of DILI by APAP treatment for 12 h in DDX3Xfl/fl and DDX3X∆hep mice, respectively (Fig. 2A). DDX3X mRNA and protein were readily expressed in DDX3Xfl/fl mice, but not in DDX3X∆hep mice (Fig. 2B, C). Histological analysis by H&E staining showed more severe liver injury in DDX3X∆hep mice compared with DDX3Xfl/fl controls induced with APAP (Fig. 2D). Consistently, ablation DDX3X causes elevated serum levels of ALT and AST (Fig. 2E). Corresponding to the protective role of hepatocyte DDX3X in DILI, survival assay showed significantly improved survival of DDX3Xfl/fl mice compared to DDX3X∆hep mice (Fig. 2F), indicating that DDX3X downregulation in hepatocytes is a major cause of liver injury in APAP induced mice.

A Schematic diagram of APAP induced DILI model; B, C mRNA and protein expression of DDX3X in mice treated with PBS or APAP for 12 h. n = 3/group; D Histological evaluation of liver sections from DDX3Xfl/fl and DDX3XΔhep mice treated with APAP or PBS for 12 h, respectively. Scale bars, 100 µm (top), 20 µm (bottom). n = 6/group; E Serum levels of ALT and AST of DDX3Xfl/fl and DDX3XΔhep mice with APAP induction. n = 6/group; F Survival assay of DDX3Xfl/fl and DDX3XΔhep mice treated with APAP or PBS for 72 h, respectively. n = 10/group. Data are expressed as mean ± SD. **p < 0.01.
Hepatocyte DDX3X protects against CCl4 or TAA induced liver injury
The effects of hepatocyte DDX3X on DILI were further validated in CCl4 and TAA induced DILI model (Fig. 3A and D). Extensively liver injury, elevated ALT and AST levels were presented in DDX3X∆hep mice compared with DDX3Xfl/fl controls after CCl4 induction (Fig. 3B, C). Consistently, DDX3X∆hep mice with TAA induction also showed extensive liver injury, increased ALT and AST levels compared with DDX3Xfl/fl controls with TAA treatment (Fig. 3E, F), further validating that hepatocyte DDX3X plays a protective role against DILI.

A Schematic diagram of CCl4 induced DILI model; B histological evaluation of liver sections from DDX3Xfl/fl and DDX3XΔhep mice treated with CCl4 for 48 h, respectively. Scale bars, 100 µm (left), 20 µm (right); C serum levels of ALT and AST of DDX3Xfl/fl and DDX3XΔhep mice with CCl4 induction; D schematic diagram of TAA induced DILI model; E Histological evaluation of liver sections from DDX3Xfl/fl and DDX3XΔhep mice treated with TAA for 24 h, respectively. Scale bars, 100 µm (left), 20 µm (right); F serum levels of ALT and AST of DDX3Xfl/fl and DDX3XΔhep mice with TAA induction. Data are expressed as mean ± SD, n = 6/group. *p < 0.05, **p < 0.01, ***p < 0.001.
Hepatocytes DDX3X is required for stress granules formation and cell survival in APAP-induced liver injury
DDX3X is a key regulator driving pro-survival SGs assembly to respond to changes in homeostatic flux [15, 20]. To demonstrate whether the deteriorated liver injury of DDX3X∆hep mice is due to the impairment of SGs formation, we examined the role of DDX3X in hepatocyte SG assembly under APAP treatment. Confocal microscopy imaging of G3BP1 (a marker of SGs) in DDX3Xfl/fl hepatocytes showed obviously SG assembly after APAP induction (Fig. 4A, B). However, DDX3XΔhep hepatocytes gained less SG formation compared to DDX3Xfl/fl hepatocytes (Fig. 4A, B). In line with that, more SYTOX green+ hepatocytes were found in APAP-treated DDX3X∆hep mice compared with APAP-treated DDX3Xfl/fl mice (Fig. 4C, D), suggesting that hepatocyte DDX3X deletion induces severe DILI may at least partly due to deficiency of pro-survival SG formation, thus impairing the capacity of hepatocytes to respond to stress stimuli.

A Negative control of primary hepatocytes were treated with PBS; and positive control of SG in primary hepatocytes were first primed with LPS for 4 h, then induced by 50 μM sodium (meta) arsenite for 30 min; B representative IF stained sections of G3BP1 (a marker of SGs) in primary hepatocytes isolated from DDX3Xfl/fl or DDX3XΔhep mice with in vitro stimulation of APAP for 12 h. Scale bars, 10 µm; C representative IF stained sections of SYTOX green in primary hepatocytes isolated from DDX3Xfl/fl or DDX3XΔhep mice with in vitro stimulation of APAP or PBS for 12 h, respectively. Scale bars, 20 µm; D Quantitative data of SYTOX green+ cells of primary hepatocytes; Data are expressed as mean ± SD, n = 3/group. ***p < 0.001.
Hepatocyte DDX3X ablation aggravated oxidative stress
Extensive production of reactive oxygen species (ROS) during drug metabolism is a prominent feature of APAP-induced hepatotoxicity [21, 22]. SGs formation is a key antioxidant process to protects cellular homeostasis in response to oxidative stress [23]. To characterize whether the deteriorated liver injury caused by DDX3X deficiency is due to impairment of antioxidant process, we examined the ROS production. Dihydroethidium (DHE) staining revealed that APAP-treated DDX3X∆hep mice displayed more extensive ROS production compared with APAP-treated DDX3Xfl/fl mice (Fig. 5A, B). In agreement with the ROS induction, higher level of APAP-metabolizing enzyme cytochromeP450, family2, subfamilyE, polypeptide1(CYP2E1) was seen in hepatocytes isolated from DDX3X∆hep mice compared with hepatocytes isolated from DDX3Xfl/fl mice in APAP medium (Fig. 5C, D). Taken together, these results indicates that hepatocyte DDX3X deletion aggravated oxidative stress in APAP-induced liver injury.

A Representative DHE stained liver sections of DDX3Xfl/fl and DDX3XΔhep mice treated with APAP or PBS. Scale bars, 25 µm; B calculation of DHE+ cells of liver sections; C, D protein expression of CYP2E1 in primary hepatocytes treated with APAP for 12 h. Data are expressed as mean ± SD, n = 3/group. *p < 0.05, **p < 0.01, ***p < 0.001.
Hepatic DDX3X deletion exacerbates macrophage infiltration and inflammation
To explain the pronounced inflammation in the livers of DDX3XΔhep mice, we evaluated hepatic mRNA expression of pro-inflammatory cytokines (including TNF-α, IL-1β and HMGB1) from liver tissues of DDX3XΔhep mice, and found these cytokines were all significantly higher in DDX3XΔhep mice than in DDX3Xfl/fl mice with APAP treatment (Fig. 6A). Consistently, higher mRNA expression of TNF-α, IL-1β and HMGB1 were also found from primary hepatocytes isolated from DDX3XΔhep mice compared with DDX3Xfl/fl mice (Fig. 6B). Considering that upregulation of these cytokines is tightly associated with macrophage-mediated inflammation, we evaluated hepatic macrophage infiltration by F4/80 immunostaining. DDX3XΔhep mice showed numerous and obvious macrophage infiltration in the livers compared with DDX3Xfl/fl mice in APAP-induced DILI model (Fig. 6C). In addition, DDX3Xfl/fl macrophages co-cultured with DDX3XΔhep hepatocytes showed increased numbers of infiltrated macrophage compared to DDX3Xfl/fl macrophages cocultured with DDX3Xfl/fl hepatocytes in vitro (Fig. 6D, E). Taken together, these data demonstrated that loss of DDX3X in hepatocytes induces macrophage infiltration, thus facilitating the inflammatory responses in APAP-induced DILI.

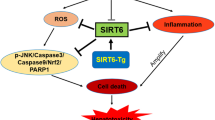
A mRNA expression levels of IL-1β, HMGB1 and TNF-α in liver tissues and primary hepatocytes of DDX3Xfl/fl and DDX3XΔhep mice; B Representative IHC stained sections of F4/80 in liver tissues of APAP induced DILI. Scale bars, 100 µm (left), 25 µm (right); (C) Schematic diagram of DDX3Xfl/fl macrophage co-cultured with DDX3Xfl/fl or DDX3XΔhep hepatocytes in APAP medium; D, E Numbers of infiltrated macrophages were evaluated by co-culture system. Scale bars, 100 µm; F Schematic diagram of DDX3X in DILI. DDX3X knockout in the hepatocytes of mice significantly exacerbated histological severity in multiple DILI murine models. This effect is through regulation of SG formation to protect hepatocytes from oxidative stress and macrophage inflammation. Data are expressed as mean ± SD, n = 3-6/group. *p < 0.05, **p < 0.01, ***p < 0.001.
Discussion
The first novel finding in present study is that DDX3XΔhep mice administrated with APAP, CCl4 or TAA showed seriously hepatotoxicity compared with DDX3Xfl/fl mice induced with same drugs. These findings were corroborated by aggregated liver histology and elevated serum ALT and AST. Moreover, DDX3X deletion was associated with a significant induction of oxidative stress and cell death. This change was tightly associated with impaired SG formation of hepatocytes, thus losing protection from ROS insults induced by APAP overdose. In addition, seriously hepatotoxicity caused by DDX3X deficiency induces pro-inflammatory cytokines (such as HMGB1, IL-1β and TNF-α) production and recruitments macrophage infiltration, which perpetuate of liver injury.
Given the downregulation of DDX3X in human and mice liver of DILI, we addressed its importance by generating hepatocyte specific DDX3X knockout mice [15]. The DDX3XΔhep mice demonstrated no obvious liver phenotype under basal conditions compared with DDX3Xfl/fl mice. However, DDX3XΔhep mice displayed deteriorate liver injury and pronounced liver inflammation compared with DDX3Xfl/fl mice under treatment of series drugs. Considering that APAP is the most widely used drug to explore the key aspects of DILI, we select APAP to explain the molecular mechanisms by which DDX3X exerts its functions in DILI. SGs formation is a major antioxidant process to protects cellular homeostasis in response to oxidative stress [23]. As a key decision maker governing the cell fate, DDX3X plays an important role to form the pro-survival SG [15]. We found that under APAP treatment, DDX3X∆hep hepatocyte displayed less SG assumable compared with DDX3Xfl/fl hepatocyte. Moreover, DDX3X∆hep hepatocyte is more prone to die compared with APAP-treated DDX3Xfl/fl hepatocyte, suggesting that hepatocyte DDX3X deletion induces severe DILI may at least partly due to impairing the capacity of hepatocytes SGs formation to response to stress stimuli. In addition, compared with APAP-treated DDX3Xfl/fl mice, more pronounced ROS production and elevated level of APAP-metabolizing enzyme CYP2E1 were also found in the liver of DDX3X∆hep mice, implying that loss of DDX3X make hepatocyte more sensitive to oxidative stress. Moreover, we noticed an increased expression of pro-inflammatory cytokines and infiltration of macrophage of DDX3X∆hep mice compared with DDX3Xfl/fl mice, indicating that lack of DDX3X might enhance the hepatocyte production of pro-inflammatory cytokines and macrophage infiltration, thus facilitating the inflammatory responses in APAP-induced DILI.
In conclusion, these observations demonstrate for the first time that DDX3X plays an essential role in the development of DILI. Further, the mechanism of this effect is through regulation of SG formation to protect hepatocytes from oxidative stress and macrophage inflammation. The use of targeted DDX3X drug maybe promising for the treatment of DILI in human.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Lammert C, Bjornsson E, Niklasson A, Chalasani N. Oral medications with significant hepatic metabolism at higher risk for hepatic adverse events. Hepatology. 2010;51:615–20.
Bernal W, Wendon J. Acute liver failure. N. Engl J Med. 2013;369:2525–34.
Larson AM, Polson J, Fontana RJ, Davern TJ, Lalani E, Hynan LS, et al. Acetaminophen-induced acute liver failure: results of a United States multicenter, prospective study. Hepatology. 2005;42:1364–72.
Jaeschke H. Acetaminophen: Dose-dependent drug hepatotoxicity and acute liver failure in patients. Dig Dis. 2015;33:464–71.
Krenkel O, Mossanen JC, Tacke F. Immune mechanisms in acetaminophen-induced acute liver failure. Hepatobiliary Surg Nutr. 2014;3:331–43.
McGill MR, Sharpe MR, Williams CD, Taha M, Curry SC, Jaeschke H. The mechanism underlying acetaminophen-induced hepatotoxicity in humans and mice involves mitochondrial damage and nuclear DNA fragmentation. J Clin Invest. 2012;122:1574–83.
Chauhan A, Sheriff L, Hussain MT, Webb GJ, Patten DA, Shepherd EL, et al. The platelet receptor CLEC-2 blocks neutrophil mediated hepatic recovery in acetaminophen induced acute liver failure. Nat Commun. 2020;11:1939.
Kolodziejczyk AA, Federici S, Zmora N, Mohapatra G, Dori-Bachash M, Hornstein S, et al. Acute liver failure is regulated by MYC- and microbiome-dependent programs. Nat Med. 2020;26:1899–911.
Ghallab A, Hassan R, Hofmann U, Friebel A, Hobloss Z, Brackhagen L, et al. Interruption of bile acid uptake by hepatocytes after acetaminophen overdose ameliorates hepatotoxicity. J Hepatol. 2022;77:71–83.
Lawson JA, Farhood A, Hopper RD, Bajt ML, Jaeschke H. The hepatic inflammatory response after acetaminophen overdose: role of neutrophils. Toxicol Sci. 2000;54:509–16.
Tacke F. Targeting hepatic macrophages to treat liver diseases. J Hepatol. 2017;66:1300–12.
Ryan CS, Schröder M. The human DEAD-box helicase DDX3X as a regulator of mRNA translation. Front Cell Dev Biol. 2022;10:1033684.
Heaton SM, Gorry PR, Borg NA. DExD/H-box helicases in HIV-1 replication and their inhibition. Trends Microbiol. 2022;31:393–404.
Li Q, Pène V, Krishnamurthy S, Cha H, Liang TJ. Hepatitis C virus infection activates an innate pathway involving IKK-α in lipogenesis and viral assembly. Nat Med. 2013;19:722–9.
Samir P, Kesavardhana S, Patmore DM, Gingras S, Malireddi RKS, Karki R, et al. DDX3X acts as a live-or-die checkpoint in stressed cells by regulating NLRP3 inflammasome. Nature. 2019;573:590–4.
Fox D, Man SM. DDX3X: stressing the NLRP3 inflammasome. Cell Res. 2019;29:969–70.
Hu K, Relton E, Locker N, Phan NTN, Ewing AG. Electrochemical measurements reveal reactive oxygen species in stress granules*. Angew Chem Int Ed Engl. 2021;60:15302–6.
Arimoto K, Fukuda H, Imajoh-Ohmi S, Saito H, Takekawa M. Formation of stress granules inhibits apoptosis by suppressing stress-responsive MAPK pathways. Nat Cell Biol. 2008;10:1324–32.
Pène V, Li Q, Sodroski C, Hsu CS, Liang TJ. Dynamic interaction of stress granules, DDX3X, and IKK-α mediates multiple functions in hepatitis C virus infection. J Virol. 2015;89:5462–77.
Cui BC, Sikirzhytski V, Aksenova M, Lucius MD, Levon GH, Mack ZT, et al. Pharmacological inhibition of DEAD-Box RNA Helicase 3 attenuates stress granule assembly. Biochem Pharm. 2020;182:114280.
Li L, Wang H, Zhang J, Sha Y, Wu F, Wen S, et al. SPHK1 deficiency protects mice from acetaminophen-induced ER stress and mitochondrial permeability transition. Cell Death Differ. 2020;27:1924–37.
Levada K, Guldiken N, Zhang X, Vella G, Mo FR, James LP, et al. Hsp72 protects against liver injury via attenuation of hepatocellular death, oxidative stress, and JNK signaling. J Hepatol. 2018;68:996–1005.
Wu S, Jiang L, Lei L, Fu C, Huang J, Hu Y, et al. Crosstalk between G-quadruplex and ROS. Cell Death Dis. 2023;14:37.
Acknowledgements
We thank members of the Department of Gastroenterology and Hepatology, Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University for helpful discussions and technical support.
Funding
This work was supported by grants from National Natural Science Foundation of China (Nos. 82000548, 82100621, 81402337), the Natural Science Foundation of Jiangsu Province (No. BK20210147), the Health Commission Fund of Jiangsu Province (No. LK2021025), the Central Guiding Local Science and Technology Development Special Project (No. XZ202202YD0018C), the Xijing Hospital Boost Plan (No. XJZT19MJ23), the Research Project of Chinese People’s Liberation Army (No. CLJ21J020), and the fundings for Clinical Trials from the Affiliated Drum Tower Hospital, Medical School of Nanjing University (No. 2021-LCYJ-PY-4).
Author information
Authors and Affiliations
Contributions
TL, SY, TZ and HZ were involved in study design, conducted the experiments, and analyzed the data; CC, XS, DC, W, KJ, DX, MC, JL and WL performed the experiments or provided material support; WX, LZ, MJ and BX designed, supervised the study, and wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Professor Alessandro Finazzi-Agrò
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Luo, T., Yang, S., Zhao, T. et al. Hepatocyte DDX3X protects against drug-induced acute liver injury via controlling stress granule formation and oxidative stress. Cell Death Dis 14, 400 (2023). https://doi.org/10.1038/s41419-023-05913-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-023-05913-x
