
Abstract
Endothelial sprouting and proliferation are tightly coordinated processes mediating the formation of new blood vessels during physiological and pathological angiogenesis. Endothelial tip cells lead sprouts and are thought to suppress tip-like behaviour in adjacent stalk endothelial cells by activating Notch. Here, we show with genetic experiments in postnatal mice that the level of active Notch signalling is more important than the direct Dll4-mediated cell–cell communication between endothelial cells. We identify endothelial expression of VEGF-A and of the chemokine receptor CXCR4 as key processes controlling Notch-dependent vessel growth. Surprisingly, genetic experiments targeting endothelial tip cells in vivo reveal that they retain their function without Dll4 and are also not replaced by adjacent, Dll4-positive cells. Instead, activation of Notch directs tip-derived endothelial cells into developing arteries and thereby establishes that Dll4–Notch signalling couples sprouting angiogenesis and artery formation.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

References
Koch, S. & Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb. Perspect. Med. 2, a006502 (2012).
Ferrara, N., Gerber, H. P. & LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 9, 669–676 (2003).
Lobov, I. B. et al. Delta-like ligand 4 (Dll4) is induced by VEGF as a negative regulator of angiogenic sprouting. Proc. Natl Acad. Sci. USA 104, 3219–3224 (2007).
Hellstrom, M. et al. Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature 445, 776–780 (2007).
Zarkada, G., Heinolainen, K., Makinen, T., Kubota, Y. & Alitalo, K. VEGFR3 does not sustain retinal angiogenesis without VEGFR2. Proc. Natl Acad. Sci. USA 112, 761–766 (2015).
Suchting, S. et al. The Notch ligand Δ-like 4 negatively regulates endothelial tip cell formation and vessel branching. Proc. Natl Acad. Sci. USA 104, 3225–3230 (2007).
Jakobsson, L. et al. Endothelial cells dynamically compete for the tip cell position during angiogenic sprouting. Nat. Cell Biol. 12, 943–953 (2010).
Benedito, R. et al. Notch-dependent VEGFR3 upregulation allows angiogenesis without VEGF-VEGFR2 signalling. Nature 484, 110–114 (2012).
Siekmann, A. F. & Lawson, N. D. Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries. Nature 445, 781–784 (2007).
Tammela, T. et al. Blocking VEGFR-3 suppresses angiogenic sprouting and vascular network formation. Nature 454, 656–660 (2008).
Benedito, R. et al. The Notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell 137, 1124–1135 (2009).
Pedrosa, A. R. et al. Endothelial Jagged1 antagonizes Dll4 regulation of endothelial branching and promotes vascular maturation downstream of Dll4/Notch1. Arterioscler. Thromb. Vasc. Biol. 35, 1134–1146 (2015).
Kangsamaksin, T. et al. NOTCH decoys that selectively block DLL/NOTCH or JAG/NOTCH disrupt angiogenesis by unique mechanisms to inhibit tumor growth. Cancer Discov. 5, 182–197 (2015).
Arima, S. et al. Angiogenic morphogenesis driven by dynamic and heterogeneous collective endothelial cell movement. Development 138, 4763–4776 (2011).
Bentley, K., Gerhardt, H. & Bates, P. A. Agent-based simulation of notch-mediated tip cell selection in angiogenic sprout initialisation. J. Theor. Biol. 250, 25–36 (2008).
Bentley, K., Mariggi, G., Gerhardt, H. & Bates, P. A. Tipping the balance: robustness of tip cell selection, migration and fusion in angiogenesis. PLoS Comput. Biol. 5, e1000549 (2009).
Bentley, K. et al. The role of differential VE-cadherin dynamics in cell rearrangement during angiogenesis. Nat. Cell Biol. 16, 309–321 (2014).
Boas, S. E. & Merks, R. M. Tip cell overtaking occurs as a side effect of sprouting in computational models of angiogenesis. BMC Syst. Biol. 9, 86 (2015).
Sorensen, I., Adams, R. H. & Gossler, A. DLL1-mediated Notch activation regulates endothelial identity in mouse fetal arteries. Blood 113, 5680–5688 (2009).
Napp, L. C. et al. Extrinsic Notch ligand Δ-like 1 regulates tip cell selection and vascular branching morphogenesis. Circ. Res. 110, 530–535 (2012).
Tachibana, K. et al. The chemokine receptor CXCR4 is essential for vascularization of the gastrointestinal tract. Nature 393, 591–594 (1998).
Ara, T., Tokoyoda, K., Okamoto, R., Koni, P. A. & Nagasawa, T. The role of CXCL12 in the organ-specific process of artery formation. Blood 105, 3155–3161 (2005).
Ding, B. S. et al. Divergent angiocrine signals from vascular niche balance liver regeneration and fibrosis. Nature 505, 97–102 (2014).
Strasser, G. A., Kaminker, J. S. & Tessier-Lavigne, M. Microarray analysis of retinal endothelial tip cells identifies CXCR4 as a mediator of tip cell morphology and branching. Blood 115, 5102–5110 (2010).
Unoki, N. et al. SDF-1/CXCR4 contributes to the activation of tip cells and microglia in retinal angiogenesis. Invest. Ophthalmol. Vis. Sci. 51, 3362–3371 (2010).
Stone, J. et al. Development of retinal vasculature is mediated by hypoxia-induced vascular endothelial growth factor (VEGF) expression by neuroglia. J. Neurosci. 15, 4738–4747 (1995).
Detmar, M. et al. Hypoxia regulates the expression of vascular permeability factor/vascular endothelial growth factor (VPF/VEGF) and its receptors in human skin. J. Invest. Dermatol. 108, 263–268 (1997).
Lee, S. et al. Autocrine VEGF signaling is required for vascular homeostasis. Cell 130, 691–703 (2007).
Domigan, C. K. et al. Autocrine VEGF maintains endothelial survival through regulation of metabolism and autophagy. J. Cell Sci. 128, 2236–2248 (2015).
Fan, J. et al. Crim1 maintains retinal vascular stability during development by regulating endothelial cell Vegfa autocrine signaling. Development 141, 448–459 (2014).
del Toro, R. et al. Identification and functional analysis of endothelial tip cell-enriched genes. Blood 116, 4025–4033 (2010).
Rocha, S. F. et al. Esm1 modulates endothelial tip cell behavior and vascular permeability by enhancing VEGF bioavailability. Circ. Res. 115, 581–590 (2014).
Han, H. et al. Inducible gene knockout of transcription factor recombination signal binding protein-J reveals its essential role in T versus B lineage decision. Int. Immunol. 14, 637–645 (2002).
Diaz-Trelles, R. et al. Notch-independent RBPJ controls angiogenesis in the adult heart. Nat. Commun. 7, 12088 (2016).
Taylor, K. L., Henderson, A. M. & Hughes, C. C. Notch activation during endothelial cell network formation in vitro targets the basic HLH transcription factor HESR-1 and downregulates VEGFR-2/KDR expression. Microvasc. Res. 64, 372–383 (2002).
Williams, C. K., Li, J. L., Murga, M., Harris, A. L. & Tosato, G. Up-regulation of the Notch ligand Delta-like 4 inhibits VEGF-induced endothelial cell function. Blood 107, 931–939 (2006).
Harrington, L. S. et al. Regulation of multiple angiogenic pathways by Dll4 and Notch in human umbilical vein endothelial cells. Microvasc. Res. 75, 144–154 (2008).
Holderfield, M. T. & Hughes, C. C. Crosstalk between vascular endothelial growth factor, notch, and transforming growth factor-β in vascular morphogenesis. Circ. Res. 102, 637–652 (2008).
Ehling, M., Adams, S., Benedito, R. & Adams, R. H. Notch controls retinal blood vessel maturation and quiescence. Development 140, 3051–3061 (2013).
Liu, Q. et al. Genetic targeting of sprouting angiogenesis using Apln-CreER. Nat. Commun. 6, 6020 (2015).
Muzumdar, M. D., Tasic, B., Miyamichi, K., Li, L. & Luo, L. A global double-fluorescent Cre reporter mouse. Genesis 45, 593–605 (2007).
Xu, C. et al. Arteries are formed by vein-derived endothelial tip cells. Nat. Commun. 5, 5758 (2014).
Schepers, A. G. et al. Lineage tracing reveals Lgr5+ stem cell activity in mouse intestinal adenomas. Science 337, 730–735 (2012).
Ridgway, J. et al. Inhibition of Dll4 signalling inhibits tumour growth by deregulating angiogenesis. Nature 444, 1083–1087 (2006).
Krebs, L. T. et al. Notch signaling is essential for vascular morphogenesis in mice. Genes Dev. 14, 1343–1352 (2000).
Murtaugh, L. C., Stanger, B. Z., Kwan, K. M. & Melton, D. A. Notch signaling controls multiple steps of pancreatic differentiation. Proc. Natl Acad. Sci. USA 100, 14920–14925 (2003).
Del Monte, G., Grego-Bessa, J., Gonzalez-Rajal, A., Bolos, V. & De La Pompa, J. L. Monitoring Notch1 activity in development: evidence for a feedback regulatory loop. Dev. Dynam. 236, 2594–2614 (2007).
Nowotschin, S., Xenopoulos, P., Schrode, N. & Hadjantonakis, A. K. A bright single-cell resolution live imaging reporter of Notch signaling in the mouse. BMC Dev. Biol. 13, 15 (2013).
Luche, H., Weber, O., Nageswara Rao, T., Blum, C. & Fehling, H. J. Faithful activation of an extra-bright red fluorescent protein in “knock-in” Cre-reporter mice ideally suited for lineage tracing studies. Eur. J. Immunol. 37, 43–53 (2007).
Lawson, N. D. et al. Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 128, 3675–3683 (2001).
Zhong, T. P., Childs, S., Leu, J. P. & Fishman, M. C. Gridlock signalling pathway fashions the first embryonic artery. Nature 414, 216–220 (2001).
Duarte, A. et al. Dosage-sensitive requirement for mouse Dll4 in artery development. Genes Dev. 18, 2474–2478 (2004).
Carlson, T. R. et al. Endothelial expression of constitutively active Notch4 elicits reversible arteriovenous malformations in adult mice. Proc. Natl Acad. Sci. USA 102, 9884–9889 (2005).
Wang, H. et al. Genome-wide analysis reveals conserved and divergent features of Notch1/RBPJ binding in human and murine T-lymphoblastic leukemia cells. Proc. Natl Acad. Sci. USA 108, 14908–14913 (2011).
Liefke, R. et al. Histone demethylase KDM5A is an integral part of the core Notch-RBP-J repressor complex. Genes Dev. 24, 590–601 (2010).
Hasan, S. S. et al. Endothelial Notch signalling limits angiogenesis via control of artery formation. Nat. Cell Biol. 19, http://dx.doi.org/10.1038/ncb3574 (2017).
Limbourg, A. et al. Notch ligand Delta-like 1 is essential for postnatal arteriogenesis. Circ. Res. 100, 363–371 (2007).
Nielsen, C. M. et al. Deletion of Rbpj from postnatal endothelium leads to abnormal arteriovenous shunting in mice. Development 141, 3782–3792 (2014).
Koch, U. et al. Delta-like 4 is the essential, nonredundant ligand for Notch1 during thymic T cell lineage commitment. J. Exp. Med. 205, 2515–2523 (2008).
Claxton, S. et al. Efficient, inducible Cre-recombinase activation in vascular endothelium. Genesis 46, 74–80 (2008).
Pitulescu, M. E., Schmidt, I., Benedito, R. & Adams, R. H. Inducible gene targeting in the neonatal vasculature and analysis of retinal angiogenesis in mice. Nat. Protoc. 5, 1518–1534 (2010).
Hozumi, K. et al. Delta-like 1 is necessary for the generation of marginal zone B cells but not T cells in vivo. Nat. Immunol. 5, 638–644 (2004).
Gerber, H. P. et al. VEGF is required for growth and survival in neonatal mice. Development 126, 1149–1159 (1999).
Ema, M., Takahashi, S. & Rossant, J. Deletion of the selection cassette, but not cis-acting elements, in targeted Flk1-lacZ allele reveals Flk1 expression in multipotent mesodermal progenitors. Blood 107, 111–117 (2006).
Haigh, J. J. et al. Cortical and retinal defects caused by dosage-dependent reductions in VEGF-A paracrine signaling. Dev. Biol. 262, 225–241 (2003).
Tokoyoda, K., Egawa, T., Sugiyama, T., Choi, B. I. & Nagasawa, T. Cellular niches controlling B lymphocyte behavior within bone marrow during development. Immunity 20, 707–718 (2004).
Wang, Y. et al. Ephrin-B2 controls VEGF-induced angiogenesis and lymphangiogenesis. Nature 465, 483–486 (2010).
Gong, S., Kus, L. & Heintz, N. Rapid bacterial artificial chromosome modification for large-scale mouse transgenesis. Nat. Protoc. 5, 1678–1696 (2010).
Brooker, R., Hozumi, K. & Lewis, J. Notch ligands with contrasting functions: Jagged1 and Delta1 in the mouse inner ear. Development 133, 1277–1286 (2006).
Yang, X. et al. Notch activation induces apoptosis in neural progenitor cells through a p53-dependent pathway. Dev. Biol. 269, 81–94 (2004).
Oswald, F. et al. A phospho-dependent mechanism involving NCoR and KMT2D controls a permissive chromatin state at Notch target genes. Nucleic Acids Res. 44, 4703–4720 (2016).
Acknowledgements
We thank G. de Luxán for help with the active NICD immunostaining protocol and H. J. Fehling (Institute of Immunology, University Clinics Ulm, Germany) for providing ROSA26RFP mice. The Max Planck Society, the University of Münster, the Cells in Motion (CiM) graduate school, the DFG Research Unit 2325 and the DFG cluster of excellence ‘Cells in Motion’ have supported this study.
Author information
Authors and Affiliations
Contributions
M.E.P., S.F.R. and R.H.A. designed the study. M.E.P., I.S., T.A. and F.B. performed experiments, B.D.G., F.F. and T.B. performed and analysed ChIP experiments, M.E. initiated the NICD rescue experiments, D.B. performed the R26R-Confetti clonal expansion and part of the RbpjiΔTC experiments, H.P. and U.H.L. contributed with image acquisition and quantification, M.S. performed the flow cytometry sorting of retinal ECs, S.F.R., T.N. and N.F. generated mutant mice, and M.E.P. and R.H.A. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Effect of Notch signalling on CXCR4 and VEGF-A in retinal ECs.
(a) Isolectin B4 (IB4, white) stained retinas of Dll4iΔEC/ΔEC, Dll1iΔEC/+ Dll4iΔEC/ΔEC, Dll1iΔEC/ΔEC Dll4iΔEC/ΔEC and control mice. Circles indicate similar vessel outgrowth. (b) Quantitation of retinal vessel progression (n = 10 control, 8 Dll4iΔEC/ΔEC, 14 Dll1iΔEC/+ Dll4iΔEC/ΔEC and 6 Dll1iΔEC/ΔEC Dll4iΔEC/ΔEC retinas) and EC area per field (n = 9 control, 5 Dll4iΔEC/ΔEC, 14 Dll1iΔEC/+ Dll4iΔEC/ΔEC and 6 Dll1iΔEC/ΔEC Dll4iΔEC/ΔEC retinas). Data represent mean ± s.e.m. P values, one-way ANOVA with Tukey’s multiple comparison post-hoc test. (c) Dll4 (white) and IB4 (green) whole-mount staining of control, Dll4iΔEC/ΔEC and Dll4iΔEC/ΔEC NICDiOEC/OEC retinas. Endothelial Dll4 protein can be seen in control but not in mutant samples (arrowheads). (d) Quantitative analysis of VEGF-A immunosignal in IB4-labelled vessels and total retinas of the indicated mutants at P6 (n = 8 control, 5 Dll4iΔEC/ΔEC, 6 Dll4iΔEC/ΔEC NICDiOEC/+ and 5 Dll4iΔEC/ΔEC NICDiOEC/OEC retinas). Increased VEGF-A levels in Dll4iΔEC/ΔEC ECs were normalized after expression of active Notch. Data represent mean ± s.e.m. P values, one-way ANOVA with Tukey’s multiple comparison post-hoc test. (e) Representative images of VEGF-A protein in Dll4iΔEC/ΔEC NICDiOEC/OEC retinas. ECs were visualized by IB4. Images on the right and in centre depict higher magnifications of insets. Rescue efficiency correlates with endothelial VEGF-A expression (arrowheads). (f) RT-qPCR analysis of Hes1, Hey1 and Efnb2 transcripts in sorted retinal ECs of the indicated mutants and littermate controls (n = 5 control, 6 Dll4iΔEC/ΔEC, 3 Dll4iΔEC/ΔEC NICDiOEC/+ and 3 Dll4iΔEC/ΔEC NICDiOEC/OEC mice from two independent tamoxifen injections). Notch target gene upregulation in Dll4 iΔEC/ΔEC NICDiOEC/+ ECs was reduced close to control expression in Dll4iΔEC/ΔEC NICDiOEC/OEC ECs. Data represent mean ± s.e.m. P values, one-way ANOVA with Tukey’s multiple comparison post-hoc test. Comparisons where P value was not shown are not significant. (g) Elevated ESM1 (blue/white) in IB4-stained (green) Dll4iΔEC/ΔEC ECs relative to control littermates. Arrowheads mark ESM1 + sprouts. ESM1 immunostaining behind the angiogenic front was greatly reduced in Dll4iΔEC/ΔEC NICDiOEC/OEC double mutant retinas (arrows).
Supplementary Figure 2 Endothelial VEGF signalling in Notch-dependent angiogenesis.
(a,b) Representative overview (a) and confocal images (b) of IB4-stained VegfaiΔEC/+ and control retinas treated with vehicle or with DAPT. Circles indicate vessel outgrowth in control. (c) Representative overview pictures of IB4-stained VegfaiΔEC/ΔEC and control retinas treated with vehicle or with DAPT. Circles indicate vessel outgrowth in control. (d,e) Confocal images of IB4 (green), ESM1 (blue) and ERG (red) stained vehicle (d) and DAPT-treated (e) VegfaiΔEC/ΔEC and control P6 retinas. Note residual ESM1 staining (arrowheads) in VegfaiΔEC/ΔEC EC sprouts but impaired DAPT-induced upregulation in the plexus (arrows) relative to control. (f) Confocal images of IB4 (green), VEGF-A (blue) and CXCR4 (red) staining of P6 retina. Note correlation (arrowheads) between CXCR4 and VEGF-A immunosignals.
Supplementary Figure 3 Role of endothelial CXCR4 in retinal angiogenesis.
(a) Representative overview confocal images showing IB4-labelled Cxcr4iΔEC/ΔEC and Cxcr4iΔEC/KO retinas together with the corresponding controls. Circles indicate vessel outgrowth in the control. (b) Combined ERG (EC nuclei; red), IB4 (green) and EdU (blue) staining shows reduced ERG + EdU + EC numbers in Cxcr4iΔEC/ΔEC retinal vessels compared to littermate control. EdU pulse-labelling was done 2 h prior to analysis. (c) Representative confocal images of CXCR4 (red/white) and IB4 (green) stained Cxcr4iΔEC/KO and control retinas. Arrowheads mark sprouts at the angiogenic front. (d) ESM1 (white) and IB4 (green) staining of Cxcr4iΔEC/ΔEC and control retinas. ESM1 expression is not overtly changed. (e) Quantitative analysis of EC proliferation for Cxcr4iΔEC/ΔEC and control littermates (n = 7 control and 8Cxcr4iΔEC/ΔEC retinas from two independent tamoxifen injections). Graphs indicate reduction of Cxcr4iΔEC/ΔEC ERG + EdU + cells per EC area at the angiogenic front relative to control, while the number of ERG + cells was not significantly changed because of the impaired outgrowth. Data represent mean ± s.e.m. P values, two-tailed unpaired t-test. (f) Quantitative analysis of vascular parameters for Cxcr4iΔEC/KO, Cxcr4KO/+ and control retinas: retinal vessel progression, EC area, branching points (n = 7 control, 10 Cxcr4KO/+ and 6 Cxcr4iΔEC/KO retinas), and sprouts (n = 5 control and 6 Cxcr4iΔEC/KO retinas). Note absence of significant differences between Cxcr4KO/+ heterozygotes and controls. Data represent mean ± s.e.m. P values, two-tailed unpaired t-test and one-way ANOVA with Tukey’s multiple comparison post-hoc test. (g) RT-qPCR analysis for Cxcr4 (n = 6 control and 7 Cxcr4iΔEC/ΔEC mice), Vegfa(n = 6 control and 12 Cxcr4iΔEC/ΔEC mice) and Esm1 (n = 5 control and 7 Cxcr4iΔEC/ΔEC mice) in sorted Cxcr4iΔEC/ΔEC and control retinal ECs. Data represent mean ± s.e.m. P values, two-tailed unpaired t-test. (h) RT-qPCR expression analysis of the Notch pathway components Dll4 (n = 7 control and 9 Cxcr4iΔEC/ΔEC mice), Jag1 (n = 8 control and 12 Cxcr4iΔEC/ΔEC mice) Hey1 (n = 4 control and 10 Cxcr4iΔEC/ΔEC mice) and Hes1 (n = 5 control and 12 Cxcr4iΔEC/ΔEC mice) in sorted Cxcr4iΔEC/ΔEC and control retinal ECs. Data represent mean ± s.e.m.P values, two-tailed unpaired t-test.
Supplementary Figure 4 Role of CXCR4 in Notch-controlled angiogenesis.
(a) Representative overview images showing DAPT-treated Cxcr4iΔEC/ΔEC and control retinas. Circles indicate vessel outgrowth in control. (b) Confocal images showing vehicle- or DAPT-treated Cxcr4iΔEC/ΔEC and control retinas. Note the decrease sprouting (arrowheads) in DAPT-treated Cxcr4iΔEC/ΔEC compared to control littermates. (c) Overview images of IB4-stained untreated or AMD3100-treated Dll4iΔEC/ΔEC retinas. Circles indicate vessel outgrowth in control (d) (e) Representative overview pictures. (d) and high magnification confocal images (e) of IB4-stained Cxcr4iΔEC/KO RbpjiΔEC/ΔEC and Cxcr4iΔEC/+ RbpjiΔEC/ΔEC retinas. Sprouting (arrowheads) was reduced in Cxcr4iΔEC/KO RbpjiΔEC/ΔEC retinas compared to Cxcr4iΔEC/+ RbpjiΔEC/ΔEC control retinas. (f) Representative confocal images of IB4-stained Cxcr4iΔEC/ΔEC Dll4iΔEC/ΔEC and Dll4iΔEC/ΔEC retinas. Sprouting (arrowheads) was reduced in Cxcr4iΔEC/ΔEC Dll4iΔEC/ΔEC retinas relative to Dll4iΔEC/ΔEC control retinas. (g) Quantitative analysis of vascular parameters for Cxcr4iΔEC/ΔEC Dll4iΔEC/ΔEC, Dll4iΔEC/ΔEC and control (Cre negative) retinas: retinal vessel progression, EC area and sprouting (n = 8 control, 6 Dll4iΔEC/ΔEC and 6 Cxcr4iΔEC/ΔEC Dll4iΔEC/ΔEC retinas). Note significant differences in sprouting but not in vessel progression and EC area between Cxcr4iΔEC/ΔEC Dll4iΔEC/ΔEC and Dll4iΔEC/ΔEC retinas. Data represent mean ± s.e.m. P values, one-way ANOVA with Tukey’s multiple comparison post-hoc test.
Supplementary Figure 5 Notch and VEGF-A-controlled processes.
(a) ChIP analysis of modified and total H3 at the Hes1, Vegfa, Esm1 and Cxcr4 loci in MS1 cells. A region located on chromosome X (Chr. X) was used as control. Shown is mean ± SD of 3 independent experiments for H3K4me1, H3K4me3 and total H3 and 2 independent experiments, measured twice each, for H3K9ac and H3K27me3 ([NS] not significant, [∗]P < 0.05, [∗∗] P < 0.01, [∗∗∗] P < 0.001, two-tailed unpaired t-test). Statistics source data for are shown in Supplementary Table 3. (b) Representation of regions (red) analysed by ChIP with distance in kilobases (kb) relative to transcriptional start sites (black arrows). (c) RT-qPCR analysis of the indicated genes in MS1 cells treated with DMSO or DAPT for 3 h (n = 6 DMSO-treated MS1 and 6 DAPT-treated MS1; individual experiments in triplicate). Data represent mean ± s.e.m. P values, two-tailed unpaired t-test. (d) Elevated VEGF-A (white) in the P7 RbpjiΔEC/ΔEC vascular plexus (arrows) and adjacent avascular tissue (arrowheads). Images are representative of three mice analysed. (e) RT-qPCR analysis of Esm1 (n = 6 control and 8 NICDiOEC/OEC mice), Vegfa (n = 5 control and 9 NICDiOEC/OEC mice), Cxcr4 (n = 6 control and 10 NICDiOEC/OEC mice) and Dll4 (n = 6 control and 9 NICD iOEC/OEC mice) in sorted NICDiOEC/OEC and control retinal ECs. Data represent mean ± s.e.m.P values, two-tailed unpaired t-test. (f) RT-qPCR analysis of Vegfr2 (n = 6 control and 10 Flk1iΔEC/KO mice), Vegfa (n = 6 control and 6 Flk1iΔEC/KO mice), Cxcr4 (n = 6 control and 6 Flk1iΔEC/KO mice) and Dll4 (n = 7 control and 5 Flk1iΔEC/KO mice) in sorted Flk1 iΔEC/KO and control ECs. Data represent mean ± s.e.m. P values, two-tailed unpaired t-test. (g) Upregulation of VEGF-A (white) in P6 NICDiOEC/OEC ECs (white arrows) and avascular retina (yellow arrowheads). (h) Increased expression of ESM1 (white) in NICDiOEC/OEC sprouts (arrowheads) and plexus (arrows). (i) Increased expression of Dll4 (blue) in NICDiOEC/OEC capillaries, whereas CXCR4 (red) did not overtly change in mutant sprouts (arrowheads) and plexus (arrows). (j) Downregulation of VEGFR2 and Dll4 in Flk1iΔEC/KO vessels with some residual signal (arrowheads). k, Lost ESM1 (white) staining in Flk1iΔEC/KO sprouts (arrowheads) and ectopic plexus expression (arrows). Panels in centre and bottom rows show higher magnifications and ESM1 signal of insets. (l) Cleaved caspase 3 (CASP3; red) detects EC death (arrows) in Flk1iΔEC/KO retinal vessels. Panels in centre and bottom rows show higher magnifications and CASP3 signal of insets. (m) DAPT-induced ESM1 (white) expression in control (arrowheads) but not in Flk1iΔEC/KO vascular plexus. (n) Quantitation of VEGF-A in P6 total retina (n = 4 control and 4 NICDiOEC/OEC retinas) and ESM1 in IB4 + vessels sprouts (S) and plexus (P) (n = 5 control and 5NICDiOEC/OEC retinas).
Supplementary Figure 6 Genetic tracking and manipulation of tip cells in vivo.
(a) Confocal images of IB4 (blue) and ESM1 (green) stained retinas at the indicated postnatal stages. Note continuous presence of ESM1 in angiogenic sprouts. (b) Examples of clonal expansion of recombined ECs in Esm1-CreERT2T/+ R26R-ConfettiT/+ double transgenics. IB4 labels ECs (blue), while nuclear GFP and cytoplasmic YFP mark recombined cell clones in arteries or at the angiogenic front (encircled areas ECs). (c) Example of quantitation approach used to analyse the percentage of recombined GFP + sprout area (green; orange in bottom image) per total angiogenic front area (orange in top image) and to analyse the percentage of recombined GFP + cells in arteries (green; orange in right image) per total arterial EC area (orange in left image). (d) Quantitation of Cxcr4iΔTC/KO, Cxcr4iΔTC/+ and Cxcr4+/KO GFP + cells per total endothelial area (n = 10 Cxcr4+/KO, 10 Cxcr4iΔTC/+ and 8 Cxcr4iΔTC/KO retinas) and of GFP + sprouts per front area (n = 10 Cxcr4+/KO, 10 Cxcr4iΔTC/+ and 8 Cxcr4iΔTC/KO retinas) at 96 h after 4-OHT injection. Data represent mean ± s.e.m. P values, two-tailed unpaired t-test. (e) Confocal images of IB4 (blue), GFP (green) and ESM1 (white) staining in P6 Notch1iΔTC/ΔTC (Esm1-CreERT2T/+ Notch1loxlox R26-mTmGT/+) and control (Esm1-CreERT2T/+ R26-mTmGT/+) retinas at 96 h after 4-OHT injection. Note presence of ESM1 immunosignal in GFP + Notch1iΔTC/ΔTC tip cells (arrowheads) and capillaries (arrows). (f) IB4 (blue), GFP (green) and CXCR4 (red) staining of Esm1-CreERT2T/+ R26-mTmGT/+ retinas at 96 h after 4-OHT injection. Recombined GFP + cells found within the plexus area (arrowheads) are positive for CXCR4.
Supplementary Figure 7 Control of tip cell behaviour by Dll4 and Jag1.
(a) Representative overview and high magnification confocal images of GFP-labelled (green) Dll4iΔTC/ΔTC (Esm1-CreERT2T/+ Dll4loxlox R26-mTmGT/+) and control (Esm1-CreERT2T/+ R26-mTmGT/+) ECs in the retinal vasculature (IB4, blue) at 48 h after 4-OHT administration. Arrowheads indicate Dll4-deficient GFP + tip cells displaying increased extension of protrusions and filopodia. (b) Confocal images of IB4 (blue), GFP (green) and Dll4 (white) staining in P6 Dll4iΔTC/ΔTC and control retinas at 96 h after 4-OHT injection. Note absence of Dll4 immunosignal in GFP + Dll4iΔTC/ΔTC tip cells ECs (white arrowheads) and reduced expression of Dll4 in non-recombined ECs (yellow arrowheads). (c) IB4 (blue), GFP (green) and CXCR4 (red) staining in P6 Dll4iΔTC/ΔTC and control retinas at 96 h after 4-OHT injection. Note increased CXCR4 immunosignal in GFP + Dll4iΔTC/ΔTC cells in plexus area (arrows) but not in sprouts (arrowheads) as compared to control GFP + cells. (d) IB4 (blue), GFP (green) and Jag1 (white) staining in P6 Dll4iΔTC/ΔTC and control retinas at 96 h after 4-OHT injection. Note reduced Jag1 expression in GFP + Dll4iΔTC/ΔTC cells plexus area (arrowheads) as compared to control GFP + cells. (e) Confocal images of IB4 (blue), GFP (green) and CXCR4 (red) staining in P6 Jag1iΔTC/ΔTC (Esm1-CreERT2T/+ Jag1loxlox R26-mTmGT/+) and control (Esm1-CreERT2T/+ R26-mTmGT/+) retinas at 96 h after 4-OHT injection. Note no significant difference for CXCR4 immunosignal between Jag1iΔTC/ΔTC recombined sprouts (white arrowheads) and non-recombined tip cells (yellow arrowheads). (f) IB4 (blue), GFP (green) and Dll4 (white) staining in P6 Jag1iΔTC/ΔTC and control retinas at 96 h after 4-OHT injection. Note no obvious difference for Dll4 immunosignal between Jag1iΔTC/ΔTC recombined sprouts (white arrowheads) and non-recombined tip cells (yellow arrowheads). Images in a–f are representative of three mice analysed.
Supplementary Figure 8 Notch activation directs tip cell progeny into arteries.
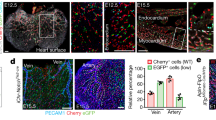
(a) Maximum intensity projection of IB4 (red), Dll4 (white) and NICD (GFP, green) staining in NICDiOTC/OTC retinas at 96 h after 4-OHT administration. Arrowheads mark recombined nuclear GFP + cells; arteries (A) are indicated. Arrowheads indicate GFP + ECs with Dll4 immunostaining in and around arteries (top panels). Images are representative of two mice analysed. (b,c) Confocal images showing IB4 (red), NICD (GFP, green) and Dll4 (white) (b) or CXCR4 (white) (c) staining in NICDiOTC/OTC retinas at 24 h after 4-OHT administration. Arrowheads mark recombined GFP + ECs with Dll4 (b) and CXCR4 (c) immunosignal in capillaries at or near the angiogenic front. Whereas all NICDiOTC/OTC-expressing cells from three mice analysed express Dll4, only 72% of NICDiOTC/OTC-expressing cells from three mice analysed show CXCR4 immunostaining. (d) Representative images showing arterial incorporation of recombined GFP + cells in Dll4iΔTC/ΔTC, Jag1iΔTC/ΔTC, Notch1iΔTC/ΔTC, RbpjiΔTC/ΔTC and Cxcr4iΔTC/KO mice and their respective controls. Note reduced arterial contribution of GFP + ECs in Dll4iΔTC/ΔTC, Jag1iΔTC/ΔTC and Notch1iΔTC/ΔTC at 96 h after 4-OHT injection. This phenotype was also seen in the RbpjiΔTC/ΔTC at 72 h but not in the Cxcr4iΔTC/KO vasculature at 96 h after 4-OHT injection. Arrowheads point to GFP + ECs incorporated into arteries. (e) Whole-mount retina immunostaining of IB4 (green) and active NICD (NICD Val1744, red). Confocal images show the angiogenic growth front on top of the artery and central vascular plexus. NICD signal is detected in ECs (white arrowheads) and perivascular cells (yellow arrowheads). High NICD signal decorates arteries and ECs in the peri-arterial plexus (bottom). Bottom row panels show examples of sprouting ECs without (yellow arrows) or with high active NICD (white arrows). (f) Additional combinations of maximum intensity projections (same regions as in Figure 8d) for CXCR4 (blue), ERG (purple), RFP (red, ESM1 recombined cells) and active endogenous Notch (YFP/Venus, green) immunostaining in CBF:H2B Venus Notch reporter (Esm1-CreERT2T/+ CBF:H2B VenusT/+ ROSA26-tdRFPT/+). White arrows point to endothelial cells with high Notch activity, while white arrowheads indicate ERG + cells with no Notch activity. Yellow arrows indicate ERG + endothelial cells from capillary front, with detectable Notch activity and reversed CXCR4 polarization.
Supplementary information
Supplementary Information
Supplementary Information (PDF 34328 kb)
Supplementary Table 1
Supplementary Information (XLSX 46 kb)
Supplementary Table 2
Supplementary Information (XLSX 33 kb)
Supplementary Table 3
Supplementary Information (XLSX 22 kb)
Rights and permissions
About this article

Cite this article
Pitulescu, M., Schmidt, I., Giaimo, B. et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat Cell Biol 19, 915–927 (2017). https://doi.org/10.1038/ncb3555
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3555
This article is cited by
-
Crosstalk between colorectal CSCs and immune cells in tumorigenesis, and strategies for targeting colorectal CSCs
Experimental Hematology & Oncology (2024)
-
Eph-ephrin signaling couples endothelial cell sorting and arterial specification
Nature Communications (2024)
-
Parenchymal cues define Vegfa-driven venous angiogenesis by activating a sprouting competent venous endothelial subtype
Nature Communications (2024)
-
Progress in the study of mechanisms and pathways related to the survival of random skin flaps
Updates in Surgery (2024)
-
Targeting adenosine A2A receptors for early intervention of retinopathy of prematurity
Purinergic Signalling (2024)
