
Abstract
To better understand the mechanisms underlying plant species responses to climate change, we compared transcriptional profiles of the co-dominant C4 grasses, Andropogon gerardii Vitman and Sorghastrum nutans (L.) Nash, in response to increased temperatures and more variable precipitation regimes in a long-term field experiment in native tallgrass prairie. We used microarray probing of a closely related model species (Zea mays) to assess correlations in leaf temperature (Tleaf) and leaf water potential (LWP) and abundance changes of ~10,000 transcripts in leaf tissue collected from individuals of both species. A greater number of transcripts were found to significantly change in abundance levels with Tleaf and LWP in S. nutans than in A. gerardii. S. nutans also was more responsive to short-term drought recovery than A. gerardii. Water flow regulating transcripts associated with stress avoidance (e.g., aquaporins), as well as those involved in the prevention and repair of damage (e.g., antioxidant enzymes, HSPs), were uniquely more abundant in response to increasing Tleaf in S. nutans. The differential transcriptomic responses of the co-dominant C4 grasses suggest that these species may cope with and respond to temperature and water stress at the molecular level in distinct ways, with implications for tallgrass prairie ecosystem function.
Similar content being viewed by others
Introduction
Predicted climate change includes increased average temperatures and more variable precipitation regimes1, which should result in increased water and temperature stress for many plant communities. Such chronic alterations in environmental conditions are expected to lead to phenotypic adaptations at the individual level2,3. For example, increased interannual variability in rainfall has been shown to select for greater morphological and physiological plasticity, which may result in evolutionary change4. Plant phenotypic responses to changing environmental conditions are achieved through the regulation of gene expression at transcriptional and post-transcriptional levels, which results in changes in metabolism and physiology. Thus, linking phenotype and gene expression could provide a more sensitive indicator of response to altered environmental conditions and stress. Furthermore, evaluating functional responses of plants at the genetic and biochemical level will increase our understanding of underlying genetic bases of responses that limit growth and survivorship, which can be related to fitness variation, selection and adaptation5,6. Individual responses also may cascade to affect community and ecosystem structure and function, particularly if the focal species is dominant in the system2,7.
Until recently, the molecular basis of plant responses to changing environmental conditions and stress in natural systems was relegated to an unknown ‘black box’8. However, genomic tools can elucidate molecular mechanisms and assess their importance in a real-world context8,9,10,11,12. Microarray profiling provides an analytical tool by which expression of thousands of genes can be studied simultaneously and has been used to monitor global gene expression in response to abiotic stresses in a number of higher plants13,14,15,16. However, most of these studies have focused on model species for which cDNA microarrays are readily available (e.g., Arabidopsis, maize, Populus, rice, and soybean) and have imposed relatively extreme conditions over short time frames and under controlled conditions (e.g., plants grown in pots exposed to drought or seedlings experiencing short-term stress). These studies provide insights into transcriptional profile changes involved in responses to various stresses and contribute to our understanding of the function of related genes. However, it is unclear how findings from these studies apply to non-model plants in natural systems. Environmental changes in natural systems may be more subtle with stress accumulating over time (allowing for acclimation). Plants in these systems are also exposed to greater variation in environmental conditions, as well as many other factors (e.g., competition, herbivory). All of these differences may alter the magnitude of variation in gene expression observed in more controlled settings8. Thus, if we are to improve our mechanistic understanding of plant responses to altered environmental conditions associated with climate change, a key step forward is to conduct genomic studies of non-model plant species under ecologically relevant field conditions.
Incorporating transcript profiling in field studies has been hampered by the lack of genomic resources available for non-model species10, although new resources are being generated at a rapid pace17. Most of the microarrays currently available for higher plants are for model and commercially important species8,17. While recent high-throughput genotyping tools, such as RAD-seq, negate having to sequence whole genomes, these techniques still require considerable time and/or resources (such as sequencing depth) to develop for non-model species. An alternative approach is to utilize microarrays designed for model species and apply them to closely related non-model species (i.e., heterologous or cross-species hybridization). Although shortcomings, such as loss of species-specific sequences and hybridization artifacts, do exist18,19, heterologous hybridizations have been successfully used to examine the transcriptomes of non-model plant species in response to a range of environmental stresses9,20,21,22,23,24, including competition25,26. However, studies of non-model plants in natural field settings are still uncommon (but see9,10).
Here, we present results from a field study designed to comparatively assess genomic responses of two native C4 grass species, Andropogon gerardii Vitman (big bluestem) and Sorghastrum nutans (L.) Nash (Indian grass), to two aspects of forecast climate change1: increased temperatures and increased variability in precipitation regimes. Previous studies reported that A. gerardii responded to experimentally altered precipitation regimes by down-regulating expression of genes related to photosynthesis, suggesting a productivity cost of drought stress adaptation or reduced performance due to drought stress10. When plants were exposed to both increased temperatures and altered precipitation regimes, A. gerardii shifted expression levels of genes in response to thermal stress rather than to water stress9. In the tallgrass prairie ecosystem, A. gerardii co-dominates with S. nutans and the two species interact to influence community and ecosystem structure and function27 and responses to climate change28,29,30. Both species are found throughout central and eastern North America, although historically S. nutans tended to dominate more mesic (southern) regions31. Despite close phylogenetic relatedness32,33, these two species fluctuate in abundance and differ in their ecophysiological responses to changes in temperature and water availability30,34,35,36,37. For example, net photosynthesis and stomatal conductance of A. gerardii was affected by increases in temperature, whereas these ecophysiological responses were more closely linked with alterations in water availability in S. nutans37. Ecophysiology of S. nutans also was more sensitive to extreme climate manipulations (heat waves) in the field30. In other words, under the same conditions, S. nutans exhibited a greater decline in ecophysiology compared to A. gerardii. Thus, these studies suggest that the two focal species presumed to be functionally similar ecologically may actually differ in important ways in their sensitivity to climate change.
As an initial step in understanding the potential molecular mechanisms underlying the differences in sensitivity observed at the ecophysiological level, we analyzed the transcriptional profiles of individuals of these co-dominant grass species exposed to both increased temperatures and altered precipitation regimes within natural field plots. Over a growing season (including a mid-July drought-recovery event), we used heterologous hybridizations with cDNA microarrays designed for a closely related model species, Zea mays, to characterize transcriptional profiles of each species in response to experimentally-induced temperature and water stress in the field. We expected that Zea mays would provide a suitable template due to conservation of stress response genes among plants38. We hypothesized that transcriptional responses would be differentially up- and down-regulated for different groups of genes, such as those involved in stress response11,16 and photosynthesis39, for A. gerardii and S. nutans. We expected that absolute differences in transcripts (sensitivity) would differ in response to temperature and water stress over the course of the growing season, with A. gerardii being more sensitive to increased temperatures and S. nutans being more sensitive to increasing water stress. These findings will help reveal mechanisms by which these two co-dominant species, and ultimately tallgrass prairie communities, respond to climate stress in a natural setting.
Results
Variation in Tleaf and LWP over the growing season for A. gerardii and S. nutans
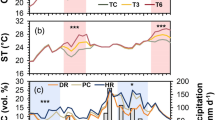
A. gerardii and S. nutans did not differ significantly in the response of leaf surface temperature (Tleaf; F1,25 = 0.62, p = 0.439) or leaf water potential (LWP; F1,26 = 1.74, p = 0.198) to the warming or altered precipitation treatments over the duration of the study, although S. nutans tended to exhibit lower (more negative) LWP than A. gerardii during mid-July. In general, Tleaf was lowest at the beginning of the growing season, peaked at the height of the drought (July 18), and then remained intermediate for the remainder of the growing season (Fig. 1A,B). In contrast, LWP became more negative as the growing season progressed (Fig. 1C,D), with LWP lowest in mid-July (July 18). For both species, Tleaf of individual plants in warmed plots was generally 1 or 2 °C higher than plants in unwarmed plots (Fig. 1, F1,6 = 8.02, p = 0.030 for A. gerardii; F1,17 = 0.23, p = 0.635 for S. nutans). But this difference in Tleaf diminished as the growing season progressed and air temperatures increased later in the growing season. Compared with plants exposed to the ambient precipitation treatment (rainfall immediately applied), plants experiencing the altered precipitation treatment (50% increase in time between ambient rainfall events) tended towards lower although not significantly different LWP and higher Tleaf (Fig. 1C,D, F1,13 = 0.64, p = 0.439 for A. gerardii; F1,40 = 0.49, p = 0.486 for S. nutans). The warming treatment decreased LWP of plants consistently compared to unwarmed plants under the altered precipitation treatment (Fig. 1C,D). As a consequence, the effects of altered precipitation vs. warming treatment were likely confounded, particularly as water became limiting (e.g., July and Aug dates). During the mid-July drought, plants had lower LWP (Fig. 1C,D) and exhibited drought and heat stress related phenotypes (e.g. dry leaf edges, leaf rolling/folding). Application of rain on July 19 to both the ambient and altered treatments resulted in recovery of LWP on July 24 (drought-recovery), but not to levels observed in June (Fig. 1C,D).

Response of leaf temperature (Tleaf) and leaf water potential (LWP) of Andropogon gerardii (A,C) and Sorghastrum nutans (B,D) to the warming and altered precipitation treatments for five sampling dates during 2006. U: un-warmed; W: warmed; AMB: ambient precipitation; ALT: altered precipitation. Box plots are shown, with median line and upper 75 and lower 25 percentiles. Error bars are the 5 and 95 percentiles of variation. Letters indicate significant difference of Tleaf or LWP among different treatments for an individual sampling date (p < 0.05).
Transcript expression patterns in response to Tleaf, LWP, and their interaction over the growing season
High-throughput regression analysis showed that the abundance of a subset of transcripts in leaf tissue of A. gerardii and S. nutans varied significantly with variation in Tleaf and LWP and the interaction of Tleaf and LWP across the sampling dates. To detect those transcripts responding separately to variation in Tleaf and LWP, we first identified transcripts correlated with the interaction of Tleaf and LWP and how many of these also responded to sampling date. Abundance of a small number of transcripts was significantly correlated with the interaction of Tleaf and LWP, with significantly more for A. gerardii (311) than S. nutans (54; χ2 = 185.46, p < 0.0001, Fig. 2A,B). Among these transcripts, 67 transcripts in A. gerardii and 11 transcripts in S. nutans also changed abundance in response to the sampling date (Fig. 2A,B).

The effect of the combination of variation in leaf temperature (Tleaf) and leaf water potential (LWP; Interaction), sampling date, and variation in Tleaf and LWP on the transcript abundance changes for individuals of Andropogon gerardii (A,C) and Sorghastrum nutans (B,D) exposed to the warming and altered precipitation treatments in 2006. The values represent the number of transcripts with significant abundance change (significant positive or negative slopes, q-value < 0.05). Number of transcripts with shared changed abundance between individual effects is shown in the overlapping areas. In all cases, the number of transcripts with abundance change was significantly different between the two species by Pearson χ2 tests (p < 0.0001).
After accounting for those transcripts significantly correlated with the interaction of Tleaf and LWP, a greater number of transcripts were correlated with variation in Tleaf for S. nutans (593) than A. gerardii (257, χ2 = 127.60, p < 0.0001, Fig. 2C,D). A small proportion of transcripts were correlated with variation in LWP for both species (1.5 vs. 0.1%), with again a greater number for S. nutans (101) than A. gerardii (11, χ2 = 64.86, p < 0.0001, Fig. 2C,D). In A. gerardii, there was no significant transcript level change correlated with both Tleaf and LWP, while the abundance of 2 and 31 transcripts changed by sampling date and variation in LWP and Tleaf, respectively. In S. nutans, 31 transcripts changed their abundance with both LWP and Tleaf, among which the abundance of 10 of these transcripts also were significantly correlated with sampling date. Eleven and 76 transcripts were significantly correlated with sampling date and variation in LWP and Tleaf, respectively.
Differential transcript abundance changes in A. gerardii and S. nutans in response to variation in Tleaf, LWP, their interaction and sampling date

(A) Number of transcripts with significant abundance change in response to variation of leaf temperature (Tleaf), leaf water potential (LWP), their interaction (Interaction) and sampling date for Andropogon gerardii (black bars) and Sorghastrum nutans (grey bars). In all cases, the number of transcripts with increased or decreased abundance change was significantly different between the two species by Pearson χ2 tests (or Fisher’s exact test) (p < 0.001). (B) The distribution of transcripts with significant abundance changes in different functional groups according to MapMan gene ontology in A. gerardii and S. nutans in response to variation in (a,b) leaf temperature (Tleaf), (c,d) leaf water potential (LWP), (e,f) the combination of Tleaf and LWP (Interaction), and (g,h) sampling date. Down = the number of transcripts with decreased abundance (negative slope). Up = the number of transcripts with increased abundance (positive slope) in response to increasing Tleaf and decreasing LWP.
-
1
Leaf temperature
Using an extension of the MapMan gene ontology from Arabidopsis to maize, transcripts with significant abundance changes were categorized into major functional groups and then further subdivided into specific functions. After accounting for statistical differences due to sampling date through multiple regression, the numbers of transcripts in A. gerardii that were correlated with variation in Tleaf positively (increased their abundance in response to increased Tleaf) vs. negatively (decreased their abundance in response to increased Tleaf) were similar (99 vs. 126, respectively; Fig. 3A). Conversely, more than two-thirds (383 of 486) of the transcripts in S. nutans were positively correlated to variation in Tleaf (Fig. 3A). Most of these were assigned to MapMan ontology cell, DNA, protein, redox, stress and transport groups (Fig. 3B-b; see Appendix S3, S4). Of the transcripts with changed abundance in response to variation in Tleaf, seven transcripts with functions in development, nucleotide metabolism, protein synthesis, redox and RNA transcriptional regulation, showed altered abundance in both A. gerardii and S. nutans (see Appendix S5). A notable difference between these two species is that more transcripts from the functional groups of stress, redox and cell changed their abundance in S. nutans compared to that in A. gerardii in connection with increasing Tleaf (Fig. 3B-a,b; Table 1). These transcripts in S. nutans were primarily represented by various heat shock proteins (see Appendix S5). In the redox group, the majority were superoxide dismutase and ascorbate peroxidase genes. All of these transcripts were positively correlated with variation in Tleaf (Table 1). Major intrinsic protein (MIP) and Tonoplast intrinsic protein (TIP), and p-/v- ATPases made up the majority of transcripts with changed abundance in the transport group for S. nutans, which again were all positively correlated with Tleaf. All the transcripts in the subgroup of “MIP” were positively correlated with Tleaf in S. nutans, but not in A. gerardii (Table 1, see Appendix S5). Similarly, in the signaling group, two transcripts related to calcium and two related to G-proteins in A. gerardii were negatively correlated with Tleaf, while most transcripts related to calcium, G-protein, receptor kinases and phosphinositides from S. nutans were positively correlated with Tleaf, except for one transcript (phosphinositides), which were negatively correlated with Tleaf (Table 1, see Appendix S5). In the photosynthesis group, only three transcripts were correlated with Tleaf in A. gerardii, one positively and two negatively, while three transcripts were all positively correlated with Tleaf in S. nutans (Table 1, see Appendix S5).
-
2
Leaf water potential
Change in abundance of transcripts differed significantly in correlation with lower LWP (i.e., more negative) in both A. gerardii vs. S. nutans (Fig. 3A). Of the 9 transcripts with significant abundance changes in A. gerardii, 5 were negatively correlated with LWP (i.e., increased abundance in response to increasing water stress, or lower LWP; Fig. 3B-c), and these were primarily in the functional groups of DNA, lipid metabolism, protein and stress (see Appendix S3, S6). In contrast, more than 80% (51 of 59) of transcripts with changed abundance in S. nutans were positively correlated with LWP (i.e., decreased abundance in response to increasing water stress, Fig. 3A), with most in the functional groups of protein and RNA, followed by groups, such as cell, development, DNA and hormone metabolism (Fig. 3B-d, see Appendix S4, S6). No transcript was found changed in abundance in both species (see Appendix S6). No transcripts from the functional group of photosynthesis showed response to variation in LWP in either species (Table 1). Two transcripts from the groups of stress and transport were all negatively correlated with LWP in A. gerardii. One transcript from the group of signaling was negatively correlated with LWP in S. nutans (Table 1, see Appendix S6).
-
3
Interaction of Tleaf and LWP
In A. gerardii, about 65% (159 of 244) of transcripts were positively correlated with the interaction of Tleaf and LWP, while about 88% of transcripts (38 of 43) were negatively correlated with the interaction of Tleaf and LWP in S. nutans (Fig. 3A), and only one transcript (Spotid 13430, with 41% sequence similarity to a rice protein, Osr40c1 protein) showed decreased abundance in both species (see Appendix S7). For A. gerardii, most transcripts from DNA, protein and RNA synthesis groups were positively correlated with the interaction of LWP and Tleaf (Fig. 3B-e, see Appendix S7). The small number of transcripts with changed abundance in S. nutans were scattered among different functional groups (e.g., cell wall, DNA, hormone metabolism, protein and RNA), in which most transcripts in the groups of protein and RNA synthesis were decreased in abundance (Fig. 3B-f, see Appendix S7). No transcripts in the functional group of photosynthesis responded to the combined effect of changing LWP and Tleaf in S. nutans, while one transcript from A. gerardii was positively correlated to the combined effect (Table 1). Transcripts from the functional groups such as redox, signaling, stress and transport showed similar regulation with the number of transcripts increased vs. decreased abundance for S. nutans (1 vs. 1) and for A. gerardii (14 vs. 9) (Table 1, see Appendix S7).
-
4
Sampling date
Five dates (two June dates, two July dates and one August date) that encompassed most of the growing season (early June to mid-August) were chosen for sampling. A large proportion of variation in transcript abundance was attributable to sampling date (Fig. 3A). Among the transcripts with changed abundance for both species, the number of transcripts with increased or decreased abundance was similar, with 501 vs. 613 for A. gerardii, 995 vs. 736 for S. nutans (Fig. 3A). Transcripts with changed abundance in response to sampling date were distributed in the similar groups for the two grass species, such as cell, development, DNA, protein, RNA, transport (Fig. 3B-g,h, see Appendix S8). Among the transcripts with changed abundance in response to sampling date, 159 transcripts were common in A. gerardii and S. nutans, most of which were assigned in the groups of protein, transport, RNA, and DNA (Fig. 3B-g,h, see Appendix S8).
Response of transcript abundance to short-term drought recovery
Differences in transcript abundance between the two sampling dates in July were used to assess short-term drought-recovery responses of the two grass species because drought stress had been largely relieved in the second sampling date by a significant rainfall event to both the ambient and altered treatments. A. gerardii and S. nutans recover leaf water status rapidly after exposure to water deficits (Fig. 1), therefore the short recovery window allowed us to minimize other abiotic and biotic variables. For S. nutans, 1,639 transcripts (excluding 1,829 transcripts with significant abundance change to the interaction of sampling date and altered precipitation) were significantly changed in abundance between the two sampling dates, with 901 decreased (reduced expression in the second date) and 738 increased in abundance. However, only 42 transcripts (excluding 3,898 transcripts with significant abundance change to the interaction of sampling date and altered precipitation) were significantly changed in abundance between the drought (July 18) and recovery (July 24) dates for A. gerardii. Only five showed reduced abundance on the July 24 date compared to the July 18 date, with three transcripts annotated (see Appendix S9). The large number of transcripts with changed abundance in S. nutans were distributed in a wide range of functional groups, e.g. cell, DNA, protein, RNA, signaling, and stress groups (Fig. 4A, see Appendix S9). The number of transcripts with increased or decreased abundance was similar except in the groups protein and RNA in which the abundance of more transcripts were increased (Fig. 4A). More transcripts in the groups of redox, stress, and transport were decreased in abundance upon recovery from drought in S. nutans (47 vs. 20). These included transcripts associated with ascorbate peroxidase, thioredoxin, HSPs, DNA J-like proteins, and metabolite transporters (see Appendix S9). Transcripts whose abundance varied in A. gerardii were among the groups such as cell, protein, RNA and signaling, most of which increased in abundance (Fig. 4B).

The distribution of transcripts within major functional groups according to MapMan gene ontology with significant abundance change in leaf tissue of (A) Sorghastrum nutans and (B) Andropogon gerardii in response to recovery from a mid-July drought period. Positive numbers indicate the number of transcripts with increased abundance (positive slope) and negative numbers indicate the number of transcripts with reduced abundance (negative slope).
Comparison of the transcript level changes during drought-recovery in A. gerardii and S. nutans to those transcripts significantly correlated with variation in LWP (season-long drought) showed that there were no transcripts that changed abundance similarly under both long-term altered soil moisture conditions and short-term drought recovery in A. gerardii. However, abundance levels of 11 transcripts were affected by both long-term drought and short-term recovery in S. nutans (see Appendix S9), and these were dispersed across a range of functional groups (see Appendix S9).
Validation of microarray results using qRT-PCR
Results of the microarray experiment were confirmed by qRT-PCR of nine differentially expressed genes for each species (CB833735, CD651724, CB380843, CB833708, CD568792, DV622645, CD001262, DV621372 for both species and DV551311 for A. gerardii; CD001612 for S. nutans; see Appendix S2). Consistent results were obtained in the expression pattern (increased or decreased in abundance) of the selected clones. Although some variation in the relative amounts of transcript accumulation was observed between the two techniques, the two were significantly correlated overall (p = < 0.0001, rho = 0.481), within A. gerardii samples (p = < 0.0001, rho = 0.559), and within S. nutans samples (p = 0.0003, rho = 0.418, see Appendix S10).
Discussion
As predicted, we found that A. gerardii and S. nutans differed significantly in the correlation of transcriptional responses with season-long variation in Tleaf and LWP, as well as their combined interaction, under field conditions. S. nutans had a greater number of transcripts that changed in abundance overall, with the majority of transcripts correlated with date and variation in Tleaf (Fig. 2). By contrast, A. gerardii showed similar numbers of transcripts changing abundance (both positive and negative) with variation in Tleaf and the combined variation in Tleaf and LWP, with few responsive to LWP. Interestingly, sampling date had the largest effect on both A. gerardii and S. nutans transcripts. This indicates that both species modify their gene expression according to changing abiotic and biotic conditions throughout the growing season regardless of the climate change treatments. Although this is difficult to distinguish from leaf maturation and ontogeny in these plants, we tried to control for this by selecting morphologically and developmentally similar individuals (vegetative, each with 3–5 leaves) at each sample date within each treatment. Among transcripts correlated with Tleaf and LWP interaction, most of these were reduced in abundance for S. nutans, while more were increased in abundance for A. gerardii (Fig. 3). During the mid-season drought-recovery period, A. gerardii and S. nutans again exhibited differential responses, during which the levels of thousands of transcripts varied in S. nutans, compared to less than 50 transcripts with significantly altered levels for A. gerardii (Fig. 4, Appendix S9), which suggests that S. nutans was more sensitive to drought conditions and responded more to alleviation from drought than A. gerardii. The finding of differential correlations of transcript responses with Tleaf and LWP, both season-long and during a mid-season drought-recovery period, is consistent with previous research showing evidence for differential sensitivity of transcript responses to temperature and water availability in A. gerardii9 and ecophysiological responses for the same individuals of A. gerardii and S. nutans sampled concurrently37 and in other experiments30.
We also observed different patterns of up- and down-regulation, and different functional groups of transcripts were found changed in abundance in A. gerardii vs. S. nutans. Moreover, rarely was there overlap between the two species in the identity of transcripts that responded significantly (Fig. 3; Appendix S3, S4). Differential regulation of multiple functional groups was observed between these two species in response to the combination of higher Tleaf and lower LWP (most transcripts in A. gerardii had increased transcript abundance levels vs. reduced abundance levels in S. nutans), LWP (lower LWP associated with increased transcripts abundance only in S. nutans), Tleaf (higher Tleaf associated with increased transcripts abundance only in S. nutans) and the drought-recovery period (increased transcript abundance associated with increased soil moisture in A. gerardii). These large differences were observed primarily in transcripts involved in different categories of processes, such as cell, glycolysis, protein, redox, RNA, signaling, stress, transport. In contrast, contrary to our expectations, we observed few responses of transcripts related to photosynthesis, and when there were significant responses, these were mostly related to variation in Tleaf rather than LWP. Other studies have shown that photosynthesis-related genes can be over-represented in response to drought recovery in several plant systems40. Previous research with A. gerardii found down-regulation of a number of photosynthesis-related genes to season-long drought10. The lack of response of photosynthesis-related genes in our study may be related to the relatively subtle nature of the experimental treatments and the lack of extreme water or temperature stress during the study period.
The differential patterns of regulation, particularly with respect to those genes involved in stress responses, suggest that the two species may use different strategies when coping with and/or responding to the increasing temperatures and water stress and their combined effects in the field. Even though no differential regulation of transcripts involved in stress recognition, such as genes in ABA synthesis pathway or transcription factor cascades, were found for the two species, we detected differential regulation of genes involved in stress avoidance and those associated with damage prevention and repair. For example, transcripts encoding sucrose synthase, which is related to osmotic adjustment for stress avoidance41, were only found in increased abundance in connection to LWP in A. gerardii. Antioxidant enzymes, such as glutathione reductase and superoxide dismutase, and HSPs play an important role in stress response, with antioxidant enzymes generally involved in the prevention of damage42 and HSP involved in damage repair by assisting with protein folding and stabilization43. In our study, transcripts encoding antioxidant enzymes and HSPs showed a consistent increase in response to increasing Tleaf in S. nutans, but not A. gerardii. The increased levels of these two groups of transcripts in response to increasing Tleaf in S. nutans suggests that this grass may be experiencing damage associated with temperature stress, whereas A. gerardii may not be experiencing stress or may responding to stress in different ways, such as through acclimation44 and osmotic adjustment.
Transcript abundance levels of aquaporins also changed significantly in S. nutans but not in A. gerardii in response to the variation of Tleaf, where generally there was increased abundance with the increase of Tleaf. Plant aquaporins, which serve as membrane water channels to increase water permeability, is a group of proteins generally thought to be involved in stress avoidance45. Aquaporins, mostly studied in Arabidopsis, have been associated with diverse abiotic stress responses, including water, nutrient, cold stress and other biotic stresses. However, the regulation of aquaporins in stress responses is inconsistent in the literature. ABA and water stress were found to increase aquaporin transcript abundance in some studies46,47, but decrease transcript levels in others47,48. As far as we know, studies of the regulation of aquaporins and their subgroups and isoforms in response to warming in plants are lacking. Regardless, the regulation of aquaporins in S. nutans but not in A. gerardii again suggests that one of the strategies for S. nutans to avoid temperature stress may be to increase expression level of aquaporins, while this was not the case for A. gerardii.
Overall, the patterns of transcript abundance levels associated with Tleaf and LWP, taken together with the ecophysiological performance of both species30,37, suggests that S. nutans is more sensitive and responsive to increasing temperatures with the increased transcripts abundance of HSPs, aquaporins, superoxide dismutase and ascorbate peroxidase. In contrast, A. gerardii appears less sensitive to the variation in temperature and water availability experienced in this experiment. A similar pattern in transcript abundance changes was observed for the short-term drought-recovery response for both species. In S. nutans, a large number of transcripts from multiple functional groups changed in abundance, with similar numbers of transcripts increased or decreased transcript abundance, while many fewer transcripts changed in abundance for A. gerardii. We cannot conclude which species is more responsive to short-term recovery from drought because it is not clear if both species responded at the same rate. This should be verified with more detailed time-course responses of both species to recovery from drought.
Plant stress response is a temporal process in which the stress is first recognized and then different strategies are adopted to either avoid the stress or respond to the stress by preventing and/or repairing the damage. Comparison of changes in levels of specific transcripts during season-long water stress and a short-term, mid-season drought-recovery period showed that, among the 1,639 transcripts changed in abundance during the drought-recovery period and the 101 transcripts that were related to the variation of LWP for S. nutans, only 11 were common between the two. Similarly, only 10% of the genes were in common when rapidly dehydrated (~6 h) barley roots were compared to gradually dehydrated (~7 d) barley roots49. This is partially due to different gene expression responses to acute versus chronic stress50,51,52. In our study, the stress gradually increased over time and encompassed a large portion of the growing season. This may have allowed for plant acclimation, which could explain the small number of transcripts regulated over the entire growing season when compared to the large number of transcripts regulated during a short-term drought-recovery period. For S. nutans, even though thousands of transcripts changed significantly in abundance with recovery from drought, many fewer responded during the season-long variation in water availability associated with the altered precipitation and warming treatments, and very few transcripts were common to the two types of stress.
In this study, we investigated the transcriptional profiles of the co-dominant tallgrass prairie C4 grass species, A. gerardii and S. nutans, exposed to variable rainfall regimes and increased temperatures under field conditions. Although our use of heterologous hybridization with maize cDNA microarrays (which was the most accessible technique available at the time of the experiment) has limitations resulting from the sequence divergence between the focal species and the model species8,18,19, we used stringent criteria to minimize these. First, during the quality control process, spots with signal to noise ratios of less than 10 were excluded from further analysis to decrease the inclusion of cross-hybridization artifacts53. As a result of these screening, approximately 70% of the transcripts with above-background detection remained for statistical analysis for both species (see Appendix S1), which suggests heterologous hybridization is feasible for analysis of transcript regulation for these two species without losing generality. Second, the microarray results of nine transcripts from each species were generally consistent with species-specific qRT-PCR results (see Appendix S10), confirming the expression changes observed with the heterologous hybridizations. Moreover, a number of significantly up- or down-regulated genes common to stress response or damage repair according to MapMan ontology were discovered in the two species responding to our imposed environmental stresses. The detection of these specific transcript changes provides confidence in the results from heterologous hybridizations. Finally, the cDNA sequences of the maize microarray SAM1.2 (18862 sequences) we used in this experiment were searched against a non-redundant transcripts data set of A. gerardii and S. nutans (unpublished data). With an E-value cutoff of <1E-10, ~75% and 73% of the probes had one or more hit to A. gerardii and S. nutans transcripts data set, respectively. This roughly corresponds to the percentage of the hybridized features to maize, with 75.14% and 78.60% for A. gerardii and S. nutans, respectively. The hybridization data suggests that there is likely conservation of stress response genes across these three species. RNA-seq techniques may provide further insight to gene expression changes, including transcripts conserved across species, as well as species specific transcripts. Although species-specific transcripts may provide insight into unique adaptations in these native grasses, it less likely that the annotated function of such transcripts is known.
In summary, the two focal species in this study, A. gerardii and S. nutans, co-dominate tallgrass prairie plant communities. They are close phylogenetic relatives32,33 who are generally considered ecologically and functionally equivalent and similarly adapted to high heat and drought conditions. However, our study found that these grasses differ substantially in their transcriptional responses to changing environmental conditions; these differences included the number, pattern and the identity of functional groups of transcripts with significant changes in abundance in response to variation in leaf temperature and leaf water potential. In particular, transcripts related to stress response, as well as other important function groups (cell function, signaling, transport), were consistently increased in abundance in S. nutans leaf tissue in response to both decreasing water availability (e.g., LWP in the season-long stress) and increasing temperatures. Little overlap was shown between the two species in the types of genes being regulated in correlation with variation in leaf-level temperature and water content. The different sensitivities of A. gerardii and S. nutans to individual stressors, which is in line with differential sensitivities observed at the ecophysiological, whole plant and community levels (e.g.,30,34,35,37), imply that these two closely related species may utilize different strategies for coping with and/or responding to environmental stressors. The differential transcriptomic responses of these co-dominant species could influence individual performance (physiological and growth) and ultimately have consequences for the structure and function of the tallgrass prairie ecosystem, as the two species contribute disproportionately to ecosystem function and structure27,54. Because this study was conducted in the field, it provides valuable insight to the mechanisms these co-dominant species use in a natural setting. We suspect that increased sensitivity by S. nutans may contribute to maladaptive plasticity55, which could lead to greater dominance in the community by A. gerardii. It is unknown whether selection for drought-resistant genotypes had already occurred in this field experiment when individuals were sampled, though is evidence for selection of genotypes over the long term56. Regardless of whether selection occurred, we posit that both plasticity within genotypes and different responses by selected genotypes will be important for understanding these species’ responses to gradual climate change.
Given that the precipitation and warming treatments represent relatively subtle changes in temperature and water availability and that it is often difficult to distinguish effects of temperature vs. water stress, the next steps to understanding how these two grass species differ in their sensitivity to climate change are to impose more severe temperature and water stress at a range of treatment levels57 and integrate proteomic and metabolomic analyses, together with the application of next generation sequencing techniques (e.g., RNAseq). This will allow us to improve our mechanistic understanding of the effects of increasing temperature vs. water stress on responses of the co-dominant grasses, and to determine whether there are thresholds in their responses.
Methods
Site description
The study was conducted at the Rainfall Manipulation Plots (RaMPs), a long-term climate change field experiment established maintained since 1998 and located at the Konza Prairie Biological Station in northeastern Kansas58. The RaMPs experiment consists of 12 location-fixed shelters (14 × 9 m) located over 6 × 6 m native tallgrass prairie plots, which are divided into four 2 × 2 m plots, two of which have infrared heating lamps installed. The RaMPs shelters allow for the manipulation of growing season precipitation in either an ambient (rainfall immediately applied) or altered pattern (50% increase in time between ambient rainfall events). On average, the altered precipitation treatment reduces volumetric soil moisture by ~15%58. Warming treatments (unwarmed, warmed) are nested within the precipitation treatments and are applied year-round to achieve on average a + 2 °C increase in ambient air temperature58. Thus, both treatments result in moderate changes in water availability and temperature in the experimental plots.
Focal species and field sampling
We focused on the two co-dominant tallgrass prairie species, A. gerardii and S. nutans, both of which are C4 grasses that reproduce primarily vegetatively via buds on belowground rhizomes59, forming dense intermixed stands of tillers (individuals). As clones grow and spread, they sever their root and rhizome connections60, and as a consequence individual tillers of sufficient distance apart are independent of each other. Because the RaMPs experiment is located in an annually burned area, tillers typically emerge in early-mid April (after burning) and grow vegetatively until mid-July/August when some (but not all) individuals initiate flowering61. Individuals typically increase in size vegetatively from mid-Apr to mid- to late-June (unpubl. data). We did not control for plant genotype due to an interest in average species-level response, although this approach can create additional noise in transcriptional response.
We sampled a single tiller of each species within the control (unwarmed) and a randomly selected warmed subplot in paired (based on adjacency) ambient and altered RaMPs. Sampling occurred five times during the growing season in 2006: June 1, June 7, July 18, July 24 and August 22. June 1 was chosen to capture early growing season conditions when water and temperature stress are typically minimal. However, due to less than ideal conditions (lower PAR (photosynthetically active radiation)), June 7 also was chosen to capture early growing season conditions with higher light availability. The two dates in July were chosen specifically to capture a drought and recovery period; the first (July 18) when soil moisture was low and the second (July 24) five days after rainfall events were applied to both the altered and ambient precipitation treatments. Finally, August 22 was chosen to capture late growing season conditions. For each sampling date, morphologically similar vegetative (non-flowering) tillers (3–5 fully expanded leaves) of each species were identified in each of the subplots. At each sampling date, one individual of each species was selected from each replicate plot subjected to the precipitation (n = 2, ambient, altered) and warming (n = 2, control, warmed) treatments, for n = 2 species × 4 treatment combinations × 3 replicates per treatment combination = 24 individuals per five sample periods, n = 120 total. Because timing discrepancies may affect gene expression12, leaf tissue was collected from individuals located in paired ambient and altered RaMPs within five minutes of each other. The first or second fully expanded leaf was randomly selected for genomic analysis from each individual to ensure leaves had finished developing but were not old enough to be senescing or have reduced cellular activity. The entire leaf was clipped and immediately flash-frozen and stored in liquid nitrogen until brought to the laboratory. Immediately after collection of the first leaf, we measured leaf surface temperature (Tleaf) on the remaining leaf, as part of intensive physiological measurements reported elsewhere (see Nippert et al.37). The whole leaf was then collected for determination of leaf water potential (LWP) using a Scholander-type pressure chamber (PMS Instruments, Inc., Corvallis, OR, USA). LWP reflects the xylem pressure required to continue the flow of water from roots to leaves, where more negative water potentials generally represent plant water stress. We focused on Tleaf as an indicator of the warming treatment and temperature stress, as it is highly correlated with canopy temperature37 and was significantly elevated in response to the warming treatment (Fig. 1A,B). Focusing on LWP as an indicator of the precipitation treatment and water stress was due to its highly correlation with soil water content at 25, 50 and 75 cm (r = 0.64, 0.61 and 0.57, respectively, with p-values < 0.0001), which was measured on a bi-weekly basis with neutron probes located in the warmed and unwarmed subplots within each RaMP. In addition, LWP was significantly reduced with the altered precipitation treatment (Fig. 1C,D). The ranges of both Tleaf and LWP captured included levels considered stressful and potentially inhibitory for ecophysiology for the two species30.
Mixed-model ANOVAs were implemented in SAS (SAS/STAT Software version 9.1.3, SAS Institute, Cary, NC, USA) to assess the effects of sampling date (five dates during the growing seaon) and the precipitation (ambient, altered) and warming (control, warmed) treatments on Tleaf and LWP between the two grass species (Fig. 1). Precipitation was treated as a fixed effect, with warming treated as a random effect nested within precipitation. Sampling date was treated as a random effect.
RNA preparation and microarray hybridization
Leaf tissue samples were stored in an −80 °C freezer for less than one year prior to RNA extraction. Total RNA was extracted from each of the 240 leaf samples (two species) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA)62, and purified with the RNeasy kit (Qiagen, Valencia, CA, USA). RNA quantity was measured by a NanoDrop spectrophotometer (Nanodrop products, Thermo Scientific, Wilmington, DE, USA). The verification of RNA quality, preparation of cDNA, and subsequent steps leading to hybridization and array scanning were performed by the Keck Biotechnology Resource Facility at Yale University (http://keck.med.yale.edu/). We used maize spotted cDNA arrays (SAM 1.2, GEO platform GPL4521) produced by the Center for Plant Genomics at Iowa State University for hybridization. The arrays included 15,680 maize cDNA clones (14,118 informative) isolated from maize ear tissue. The sample pairing for microarray hybridization followed Design C of Milliken et al.63, in which pairs were assigned within a level (ambient vs. altered precipitation) of the whole plot treatment and across levels (unwarmed vs. warmed) of the subplot treatment.
Quality control of heterologous hybridizations
In total, there were hybridizations for 47 leaf samples of A. gerardii and 49 leaf samples of S. nutans. Some samples had to be discarded due to low RNA quality. Array image data were collected using GenePix software (Version 6, Molecular Devices, Inc., Sunnyvale, CA, USA) and normalized using the ratio of medians. Prior to the normalization, features with obvious abnormality and saturated signal were flagged as uninformative and excluded from statistical analysis. Because the signal intensity of some spots was low as a consequence of the heterologous hybridization, stringent criteria concerning signal to noise ratios were applied during quality control of the images. Features with signal to noise ratios (for both channels, 635 nm and 532 nm) less than 10 were excluded from further normalization. After quality control (see Appendix S1 in Supporting Information), there were 10,607 features in total for further analysis for A. gerardii, while 11,095 features were included for S. nutans, among which 10,329 were common for both species. Considering the different hybridization efficiencies (Appendix S1), we focused the following statistical analysis on only the common features of A. gerardii and S. nutans, discarding 278 unique transcripts from A. gerardii and 766 from S. nutans.
Statistical analysis of microarray data
The microarray data were analyzed for each species separately using high-throughput regression9 to assess how gene expression varied among individual plants as a consequence of altered precipitation and temperatures over the entire growing season. Specifically, multiple regression analysis was used to capture the linear relationship between gene expression levels, as the dependent variable, and Tleaf, LWP, together with their interaction, and date (representing the five sampling dates) as the independent variables. The inclusion of date was used to distinguish responses of the transcriptome that are due to changes in temperature or water availability from those due to sampling time. Prior to regression analysis, the microarray data were normalized following Travers et al.9.
Regression analysis was conducted for each species separately with the residuals from the normalization analysis. Data was subset so only the common hybridized features (10,329) represented by all five sampling dates were included in the regression analyses: 10,309 (99.8% of the common hybridized features) features in total for both species. Statistical significance of either positive (increased transcript abundance associated with higher Tleaf or decreased transcript abundance associated with lower LWP) or negative (decreased transcript abundance associated with higher Tleaf or increased transcript abundance associated with lower LWP) slopes was tested against the null hypothesis that the slope was zero. Significant differences in the number of transcripts with abundance change in response to Tleaf, LWP, their interaction, and sampling date between the two species were tested by Pearson χ2 test.
To assess genes responding to drought vs. recovery, we subsetted the data focusing on the two sampling dates in July. Specifically, mixed-model ANOVA was used to assess alteration of transcript levels under drought conditions (July 18) vs. drought recovery (July 24). There were 8,662 and 10,776 (82% and 97% of hybridized features) transcripts analyzed for A. gerardii and S. nutans respectively, among which 7869 features were common for both species. Similarly, we focused on these common features in the drought-recovery analysis. The normalized expression data (residuals from the normalization analyses above) were used as the dependent variable, with precipitation and warming treatments, sampling date and their interactions as independent variables (fixed effects), and RaMP as random effect.
The normalization analyses, high-throughput regression analyses, mix-model ANOVA for drought recovery evaluation, and Pearson χ2 test were implemented in SAS (version 9.1.3). For regression analyses and mix-model ANOVA, statistical significance was further evaluated using the q-statistic to control the experiment-wise false discovery rate (FDR)64. A q-value of less than 0.05 was chosen to identify significant changes in transcript levels. The transcripts with significant abundance changes were grouped into major functional groups using an extension of MapMan gene ontology65 to maize using protein sequence identity to Arabidopsis.
Quantitative Real-time Polymerase Chain Reaction (qRT-PCR)
We used qRT-PCR to validate the microarray results for the same RNA samples used in the heterologous hybridization (see Appendix S2). Because leaf tissue of A. gerardii and S. nutans was often limited and a relatively large amount of RNA was utilized for the microarray hybridizations, RNA samples were not available for qRT-PCR analysis for all individuals or for all sampling dates. Therefore, we focused on only those sampling dates in which there were adequate samples available for a subset of individuals (June 7 and July 18 for A. gerardii, June 7 and August 22 for S. nutans). Nine annotated transcripts with significant response to variation in LWP or Tleaf according to microarray analysis and which have been shown to be involved in stress response (e.g., superoxide dismutase, HSP 90) were selected and cloned (Appendix S2). Expression levels of these transcripts were analyzed for each species with qRT-PCR, with β-tubulin and Elongation factor (EF) 1α as the internal standard for A. gerardii and S. nutans, respectively. The range of the primer amplification efficiency was between 90% and 98%, and correlation coefficiency was between 99.1% and 100.0% as determined by serial dilution of cDNA templates.
Additional Information
How to cite this article: Smith, M. D. et al. Gene expression patterns of two dominant tallgrass prairie species differ in response to warming and altered precipitation. Sci. Rep. 6, 25522; doi: 10.1038/srep25522 (2016).
Accession codes
References
Stocker, T. F. et al. IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (2013).
Smith, M. D., Knapp, A. K. & Collins, S. A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecology 90, 3279–3289 (2009).
Matesanz, S., Gianoli, E. & Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 1206, 35–55 (2010).
Lázaro-Nogales, A. et al. Environmental heterogeneity leads to higher plasticity in dry-edge populations of a semiarid Chilean shrub: insights into climate change responses. J. Ecol. 103, 338–350 (2015).
Pratt, J. D. & Mooney, K. A. Clinal adaptation and adaptive plasticity in Artemisia californica: Implications for the response of a foundation species to predicted climate change. Glob. Chang. Biol. 19, 2454–2466 (2013).
Gibson, G. The environmental contribution to gene expression profiles. Nat. Rev. Genet. 9, 575–581 (2008).
Gustafson, D. J. et al. Genetic sorting of subordinate species in grassland modulated by intraspecific variation in dominant species. PLoS One 9, 1–7 (2014).
Leakey, A. D. B. et al. Gene expression profiling: Opening the black box of plant ecosystem responses to global change. Glob. Chang. Biol. 15, 1201–1213 (2009).
Travers, S. E. et al. Variation in gene expression of Andropogon gerardii in response to altered environmental conditions associated with climate change. J. Ecol. 98, 374–383 (2010).
Travers, S. E. et al. Ecological genomics: making the leap from model systems in the lab to native populations in the field. Front. Ecol. Environ. 5, 19–24 (2007).
Johnson, S. M. et al. Transcriptomic analysis of Sorghum bicolor responding to combined heat and drought stress. BMC Genomics 15, 456, doi: 10.1186/1471-2164-15-456 (2014).
Meyer, E. et al. Integrating transcriptional, metabolomic, and physiological responses to drought stress and recovery in switchgrass (Panicum virgatum L.). BMC Genomics 15, 527, doi: 10.1186/1471-2164-15-527 (2014).
Nakashima, K. et al. Comparative functional analysis of six drought-responsive promoters in transgenic rice. Planta 239, 47–60 (2014).
Fernandez, P. et al. Transcriptomic identification of candidate genes involved in sunflower responses to chilling and salt stresses based on cDNA microarray analysis. BMC Plant Biol. 8, 1–18 (2008).
Gullì, M. et al. Comparison of Drought Stress Response and Gene Expression between a GM Maize Variety and a Near-Isogenic Non-GM Variety. PLoS One 10, 1–21 (2015).
Wilkins, O., Waldron, L., Nahal, H., Provart, N. J. & Campbell, M. M. Genotype and time of day shape the Populus drought response. Plant J. 60, 703–715 (2009).
Alvarez, M., Schrey, A. W. & Richards, C. L. Ten years of transcriptomics in wild populations: what have we learned about their ecology and evolution? Mol. Ecol. 24, 710–725 (2015).
Bar-Or, C., Czosnek, H. & Koltai, H. Cross-species microarray hybridizations: a developing tool for studying species diversity. Trends Genet. 23, 200–207 (2007).
Whitehead, A. Comparative genomics in ecological physiology: toward a more nuanced understanding of acclimation and adaptation. J. Exp. Biol. 215, 884–891 (2012).
Gong, Q., Li, P., Ma, S., Indu Rupassara, S. & Bohnert, H. J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 44, 826–839 (2005).
Hammond, J. P. et al. A comparison of the Thlaspi caerulescens and Thlaspi arvense shoot transcriptomes. New Phytol. 170, 239–260 (2006).
Sharma, N., Cram, D., Huebert, T., Zhou, N. & Parkin, I. A. P. Exploiting the wild crucifer Thlaspi arvense to identify conserved and novel genes expressed during a plant’s response to cold stress. Plant Mol. Biol. 63, 171–184 (2006).
Horvath, D., Anderson, J. V., Chao, W. S., Foley, M. E. & Doğramaci, M. In Advances in the Understanding of Biological Sciences Using Next Generation Sequencing (NGS) Approaches (eds. Sablok, G., Kumar, S., Ieno, S., Kuo, J. & Varotto, C. ) 63–78 (Springer International Publishing, 2015). doi: 10.1007/978-3-319-17157-9_5.
Chelaifa, H., Mahe, F. & Ainouche, M. Transcriptome divergence between the hexaploid salt-marsh sister species Spartina maritima and Spartina alterniflora (Poaceae). Mol. Ecol. 19, 2050–2063 (2010).
Horvath, D. P., Llewellyn, D. & Clay, S. A. Heterologous Hybridization of Cotton Microarrays with Velvetleaf (Abutilon Theophrasti) Reveals Physiological Responses Due to Corn Competition. Weed Sci. 55, 546–557 (2007).
Broz, A. K., Manter, D. K., Callaway, R. M., Paschke, M. W. & Vivanco, J. M. A molecular approach to understanding plant–plant interactions in the context of invasion biology. Funct. Plant Biol. 35, 1123–1134 (2008).
Smith, M. D. & Knapp, A. K. Dominant species maintain ecosystem function with non-random species loss. Ecol. Lett. 6, 509–517 (2003).
Knapp, A. K. et al. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 298, 2202–2205 (2002).
Fay, P. A., Carlisle, J. D., Knapp, A. K., Blair, J. M. & Collins, S. L. Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 137, 245–251 (2003).
Hoover, D. L., Knapp, A. K. & Smith, M. D. Contrasting sensitivities of two dominant C4 grasses to heat waves and drought. Plant Ecol. 215, 721–731 (2014).
Brown, D. A. Early Nineteenth-Century Grasslands of the Midcontinent Plains. Ann. Assoc. Am. Geogr. 83, 589–612 (1993).
Morrone, O. et al. Phylogeny of the Paniceae (Poaceae: Panicoideae): integrating plastid DNA sequences and morphology into a new classification. Cladistics 28, 333–356 (2012).
Giussani, L. M., Cota-Sanchez, J. H., Zuloaga, F. O. & Kellogg, E. A. A molecular phylogeny of the grass subfamily Panicoideae (Poaceae) shows multiple origins of C4 photosynthesis. Am. J. Bot. 88, 1993–2012 (2001).
Silletti, A. & Knapp, A. K. Long-term responses of the grassland co-dominants Andropogon gerardii and Sorghastrum nutans to changes in climate and management. Plant Ecol. 163, 15–22 (2002).
Silletti, A. M., Knapp, A. K. & Blair, J. M. Competition and coexistence in grassland codominants: responses to neighbour removal and resource availability. Can. J. Bot. 82, 450–460 (2004).
Swemmer, A. M., Knapp, A. K. & Smith, M. D. Growth responses of two dominant C4 grass species to altered water availability. Int. J. Plant Sci. 167, 1001–1010 (2006).
Nippert, J. B., Fay, P. A., Carlisle, J. D., Knapp, A. K. & Smith, M. D. Ecophysiological responses of two dominant grasses to altered temperature and precipitation regimes. Acta Oecologica 35, 400–408 (2009).
D’Orso, F. et al. Conservation of AtTZF1, AtTZF2, and AtTZF3 homolog gene regulation by salt stress in evolutionarily distant plant species. Front. Plant Sci. 6, 1–16 (2015).
Chaves, M. M., Flexas, J. & Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 103, 551–560 (2009).
Priest, H. D. et al. Analysis of Global Gene Expression in Brachypodium distachyon Reveals Extensive Network Plasticity in Response to Abiotic Stress. PLoS One 9, 1–15 (2014).
Van den Ende, W. & El-Esawe, S. K. Sucrose signaling pathways leading to fructan and anthocyanin accumulation: A dual function in abiotic and biotic stress responses? Environ. Exp. Bot. 108, 4–13 (2014).
Rizhsky, L., Liang, H. & Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol 130, 1143–1151 (2002).
Kapoor, M. & Roy, S. S. In Abiotic Stresses in Crop Plants (eds. Chakraborty, U. & Chakraborty, B. N. ) 1–22 (CAB International, 2015).
Barua, D. et al. Variation in heat-shock proteins and photosynthetic thermotolerance among natural populations of Chenopodium album L. from contrasting thermal environments: implications for plant responses to global warming. J. Integr. Plant Biol. 50, 1440–1451 (2008).
Li, G., Santoni, V. & Maurel, C. Plant aquaporins: Roles in plant physiology. Biochim. Biophys. Acta 1840, 1574–1582 (2014).
Mirzaei, M., Pascovici, D., Atwell, B. J. & Haynes, P. A. Differential regulation of aquaporins, small GTPases and V-ATPases proteins in rice leaves subjected to drought stress and recovery. Proteomics 12, 864–877 (2012).
Maurel, C. et al. Aquaporins in Plants. Physiol. Rev. 95, 1321–1358 (2015).
Katsuhara, M., Tsuji, N., Shibasaka, M. & Panda, S. K. Osmotic stress decreases PIP aquaporin transcripts in barley roots but H2O2 is not involved in this process. J. Plant Res. 127, 787–792 (2014).
Talamè, V., Ozturk, N. Z., Bohnert, H. J. & Tuberosa, R. Barley transcript profiles under dehydration shock and drought stress treatments: A comparative analysis. J. Exp. Bot. 58, 229–240 (2007).
Deyholos, M. K. Making the most of drought and salinity transcriptomics. Plant. Cell Environ. 33, 648–654 (2010).
Zhang, J.-Y. et al. Global reprogramming of transcription and metabolism in M edicago truncatula during progressive drought and after rewatering. Plant. Cell Environ. 37, 2553–2576 (2014).
Cramer, G. R., Urano, K., Delrot, S., Pezzotti, M. & Shinozaki, K. Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biology 11, 1–14 (2011).
Verdnik, D., Handran, S. & Pickett, S. In DNA arrray image analysis: nuts and bolts (ed. Kamberova, G. ) 83–98 (DNA Press, 2002).
Smith, M. D., Wilcox, J., Kelly, T. & Knapp, A. K. Dominance not richness determines invasibility of tallgrass prairie. Oikos 2, 253–262 (2004).
Miner, B. G., Sultan, S. E., Morgan, S. G., Padilla, D. K. & Relyea, R. a. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 20, 685–92 (2005).
Avolio, M. L., Beaulieu, J. M. & Smith, M. D. Genetic diversity of a dominant C4 grass is altered with increased precipitation variability. Oecologia 171, 571–581 (2013).
Smith, M. D. An ecological perspective on extreme climatic events: a synthetic definition and framework to guide future research. J. Ecol. 99, 656–663 (2011).
Fay, P. A. et al. Relative effects of precipitation variability and warming on tallgrass prairie ecosystem function. Biogeosciences 8, 3053–3068 (2011).
Benson, E. J. & Hartnett, D. C. The Role of Seed and Vegetative Reproduction in Plant Recruitment and Demography in Tallgrass Prairie. Plant Ecol. 187, 163–178 (2006).
Weaver, J. E. & Fitzpatrick, T. J. The Prairie. Ecological Monographs 4, 109–295 (1934).
La Pierre, K. J. et al. Explaining temporal variation in above-ground productivity in a mesic grassland: the role of climate and flowering. J. Ecol. 99, 1250–1262 (2011).
McCarty, D. R. A simple method for extraction of RNA from maize tissue. Maize Genet. Coop. Newsl. 60, 61 (1986).
Milliken, G. A., Garrett, K. A. & Travers, S. E. Experimental Design for Two-Color Microarrays Applied in a Pre-Existing Split-Plot Experiment. Stat. Appl. Genet. Mol. Biol. 6, 1–21 (2007).
Storey, J. D. & Tibshirani, R. Statistical significance for genome wide studies. Proc. Natl. Acad. Sci. 100, 9440–9445 (2003).
Thimm, O. et al. MAPMAN: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 37, 914–939 (2004).
Acknowledgements
We thank J. Bai, K.A. Garrett, S.H. Hulbert, J.E. Leach, A.A. Saleh, and S.E. Travers for their contributions to the study. We particularly thank S. Yuan for conducting the statistical/bioinformatics analyses, preparing all tables, and contributing to writing the first draft of the manuscript. Thanks to C. Anderson, E. Frank, D. Hoover, N. Lu, K. Wagner and C. Yaege for field, laboratory, and bioinformatics assistance. Additional thanks to A. Monteiro, S. Dellaporta, J. Townsend and G. Wagner for support in the real-time PCR experiment. A.K. Knapp and J.M. Blair conceived the rainfall manipulation experiment. Support was provided to M.D.S. by the US Department of Energy, Office of Environment and Science (Grant #DE-FG02-04ER63892). Additional support was provided by the USDA CREES Ecosystem Studies Program, and the NSF Ecosystems and LTREB programs.
Author information
Authors and Affiliations
Contributions
M.D.S. conceived the study. M.D.S. wrote the first draft of the manuscript. M.D.S., A.M.H. and M.L.A. edited subsequent versions of the manuscript. A.M.H. prepared Figure 3 and 4.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Microarray data files were submitted to GEO-NCBI data bank under the series ID number GSE35492 (GSE35492).
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Smith, M., Hoffman, A. & Avolio, M. Gene expression patterns of two dominant tallgrass prairie species differ in response to warming and altered precipitation. Sci Rep 6, 25522 (2016). https://doi.org/10.1038/srep25522
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep25522
This article is cited by
-
Variation and heritability of morphological and physiological traits among Leymus chinensis genotypes under different environmental conditions
Journal of Arid Land (2019)
-
Linking gene regulation, physiology, and plant biomass allocation in Andropogon gerardii in response to drought
Plant Ecology (2018)
-
Effects of extreme changes in precipitation on the physiology of C4 grasses
Oecologia (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
