
Abstract
Resistance to anoikis and Epithelial-mesenchymal transition (EMT) are two processes critically involved in cancer metastasis. In this study, we demonstrated that after anchorage deprival, hepatocellular carcinoma (HCC) cells not only resisted anoikis, but also exhibited EMT process. Microarray expression profiling revealed that expression of miR-424-5p was significantly decreased in anoikis-resistant HCC cells. Ectopic overexpression of miR-424-5p was sufficient to reverse resistance to anoikis, block EMT process and inhibit malignant behaviors of HCC cells. Target analysis showed that a potent β-catenin inhibitor, ICAT/CTNNBIP1 was a direct target of miR-424-5p. Further study demonstrated that miR-424-5p reversed resistance to anoikis and EMT of HCCs by directly targeting ICAT and further maintaining the E-cadherin/β-catanin complex on the cellular membrance. In vivo study further demonstrated that miR-424-5p significantly inhibited the tumorigenicity of HCC cells in nude mice. Clinical investigation demonstrated that miR-424-5p was significantly downregulated in HCC tissues compared with that of the non-cancerous liver tissues and this decreased expression of miR-424-5p was significantly correlated with higher pathological grades and more advanced TNM stages. Therefore, aberrant expression of miR-424-5p is critically involved in resistance to anoikis and EMT during the metastatic process of HCC and its downregulation significantly contributes to liver cancer progression.
Similar content being viewed by others

Introduction
Hepatocellular carcinoma (HCC) is one of the most common solid malignant tumors worldwide and is among the leading cause of cancer related death. The 5-year survival rate is rather low because of its rapid progression and high metastatic rate. Metastasis is a multistep process including dissociation of cancer cells from the primary sites, survival in the vascular system and proliferation at distant target organs. As a barrier to metastasis, cells normally undergo an apoptotic process known as anoikis, a type of cell death due to loss of contact with the extracellular matrix. Cancer cells acquire resistance to anoikis after detachment from the primary sites, thereafter travel through the circulatory systems to disseminate throughout the body1.
Epithelial-to-Mesenchymal Transition (EMT) was well recognized as a feature of embryogenesis, which is vital for morphogenesis during embryonic development. Recently increasing evidence suggests that EMT process is critically involved in tumor development and metastasis2,3,4. Because resistance to anoikis and EMT are tightly linked events involved in metastasis, exploring the relationship between these two progress, as well as the molecular mechanisms involved in it is thus a promising way for better understanding metastasis.
MicroRNAs(miRNAs) are 20–22 nucleotides non-coding RNAs, which regulate gene expressions at the posttranscriptional level5, subsequently control crucial physiological processes and thus play important roles in biological and pathological processes, including cancer6,7. Because of their abilities to exert functions similar to tumor suppressor or promoters, miRNAs are believed to play important roles in carcinogenesis and cancer progression. Thus it is not surprising that they are also implicated in critical events in metastasis, including resistance to anoikis and EMT8. Though the role of miRNA in metastasis of several cancers has been reported, its role in the critical events involved in metastasis of hepatocarcinoma is not completely clarified. Thus, in this study, we investigated the role of a key microRNA, miR-424-5p in resistance to anoikis and the related EMT process in the anchorage-deprived HCC cells. Deciphering the role of miR-424-5p in anchorage-independent HCC cells may offer new strategy to manipulate liver cancer metastasis.
Results
MiR-424-5p is down-regulated in detached anoikis-resistant HCC cells
Our previous studies have shown that anchorage-deprived HCC cells could resist anoikis and acquire more malignant property after detachment9,10. Although microRNA is reported to play a critical role in the progression of cancer, whether it is involved in the anoikis-resistance of HCC cells is not known and is investigated in this study. In order to identify miRNAs that are potentially involved in the anoikis-resistant process of HCC cells, we employed a miRNA microarray expression profiling and screened out a series of miRNAs which were differentially expressed between attached and detached HCC cells. Microarray analysis showed that miR-424-5p was among the top down-regulated hits (Fig. 1a) and its significant down-regulation in the anchorage-deprived HCC cells was further validated by quantitative real-time PCR (Fig. 1b).

Expression of miR-424-5p in detached hepatocarcinoma cells.
(a) HCC cells were plated in poly-HEMA coated and non-coated plates and allowed for growth for 24 h as detached and attached cells. MiRNA expression profiling of these two groups was analyzed and the statistically differently expressed microRNAs were selected. (b)BEL7402, SMMC7721 and HepG2 cells were plated in 6-well plates without or with poly-HEMA as attached and detached group. After 24 h of culture, cells were collected and the expression of miR-424-5p in BEL7402, SMMC7721 and HepG2 cells was detected by real-time PCR. Histogram represented the relative miR-424-5p expression levels normalized to U6. Data were presented as mean ± s.d. from triplicate analysis and were the representative data from at least three independent experiments. *P<0.05. **P<0.01.
Hepatocellular carcinoma cells undergo EMT after detachment
Both anoikis-resistant process and EMT are regarded as important events in the metastatic process of cancer cells. Our previous study showed that acquisition of resistance to anoikis conferred HCC cells more malignant behaviors, including resistance to apoptosis induction, resistance to extracellular stimuli and increased capability of invasion10. These properties are also known as features of EMT-transformed mesenchymal cells. Therefore it is intriguing to see whether these anchorage independent HCC cells also undergo EMT process. Thus we detected EMT related markers in these detached HCC cells by western blot, with β-actin as an internal control. Our data showed that expression levels of epithelial markers, including E-cadherin and β-catenin were significantly down-regulated, accompanied with up-regulation of mesenchymal markers, including N-cadherin and vimentin (Fig. 2a–b). The expression changes of these markers were also verified by quantitative real-time PCR (Fig. 2c). Snail, an identified transcription factor repressing epithelial genes via E-boxes in the corresponding promoters, was also analyzed. Our data showed that snail was significantly upregulated in these detached HCC cells (Fig. 2a–c). These data suggested that EMT was dramatically induced by acquisition of resistance to anoikis in the detached HCC cells. Moreover, our previous data showed that anchorage deprival also resulted in the acquisition of other properties involved in cancer progression, such as an increased ability to migrate, a higher resistance to apoptosis and a higher capability of colony formation11. Thus, all of these properties are in consistence with a more invasive and malignant behavior of EMT-transformed cells in cancer progression.

Expression of EMT related markers in attached and detached HCC cells.
(a) BEL7402, SMMC7721 and HepG2 cells were seeded in 6-well plates without or with poly-HEMA coating as attached and detached HCC cells. After 24 h of culture, cells were collected and the expression of EMT markers in BEL7402, SMMC7721 and HepG2 cells was detected by western blot. β-actin was used as an internal control. (b) Quantitation of band intensities from western blot analysis was analyzed densitometrically using Image J software and normalized to β-actin. (c) After the detached and attached cells were cultured for 24 h, expression of EMT related markers were detected by real-time PCR and the histograms represented the relative mRNA expression levels normalized to β-actin. Presented data were representative one from at least three independent experiments. *P<0.05. **P<0.01.
Ectopic overexpression of miR-424-5p reversed resistance to anoikis and EMT of anchorage-deprived HCC cells
MiR-424-5p was among the top significantly down-regulated microRNAs in the detached HCC cells and previous reports indicated its involvement in carcinogenesis12,13,14. Thus we are interested in defining its role in resistance to anoikis and in EMT process of these anchorage-deprived HCC cells. To clarify this issue, we transfected miR-424-5p into the detached HCC cells and its enforced expression efficiency was verified by real-time PCR (Fig. 3a). Our data showed that after anchorage deprival, cell viabilities of the miR-424-5p transfected cells were significantly decreased compared with the mock control (Fig. 3b–c). Further analysis showed a significant activation of caspase 3 in the miR-424-5p transfected detached HCC cells (Fig. 3d–g). Because cleavage of caspase 3 was a definite evidence of apoptosis, these data indicated that massive apoptosis was induced in the mir-424-5p transfected cells. Apoptosis occurrence in the miR-424-5p transfected detached HCC cells was further confirmed by ANNEXIN V-FITC apoptosis assay (Fig. 3h). Altogether, these data indicated that miR-424-5p transfection could reverse resistance to anoikis in the anchorage-deprived HCC cells.

Enforced miR-424-5p expression in HCC cells reversed anoikis resistance of HCC cells.
(a) BEL7402, SMMC7721 and HepG2 cells were transiently transfected with 50 nM miR-424-5p or random mock control, respectively. MiR-424-5p level in the transfected HCC cells was detected by real-time PCR and normalized to internal control U6. (b–c) The miR-424-5p transftected HepG2 cells were cultured in anchorage-deprived condition for 24 h; and cell viabilities were determined by trypan blue staining(b) and CCK8 kit(c). (d) The miR-424-5p transfected HepG2 cells were transferred to the poly-HEMA coated wells and cultured as anchorage-deprived cells for another 24 h. Caspase 3 activation was determined by caspase 3 substrate assay kit. (e–g) The miR-424-5p transfected HepG2 and SMMC7721 cells were cultured in the poly-HEMA coated wells as detached cells for another 24 h, caspase 3 activity was detected by western blot (e). The band densities were analyzed by Image J software and normalized to β-actin(f–g). (h) Apoptotic rate of the miR-424-5p transfected detached HCC cells were determined by ANNEXIN V-PI apoptotic assay kit followed by flow cytometry analysis. Presented data were representative one from at least three independent experiments. * P < 0.05, **P < 0.01.
Besides conquering resistance to anoikis, ectopic overexpression of miR-424-5p was also accompanied with decreased expression of mesenchymal markers (N-cadherin, Vimentin) and increased expression of epithelial markers(E-cadherin, β-catenin) (Fig. 4a), which indicated a reversion of EMT in the detached HCC cells as well. Furthermore, other malignant behaviors accompanied with resistance to anoikis, including colony formation and invasion was also significantly inhibited by overexpression of miR-424-5p (Fig. 4b–c). Taken together, these data indicated a critical role of miR-424-5p involved in HCC progression by suppressing anoikis and reversing EMT related malignant behaviors of detached HCC cells.

Enforced miR-424-5p expression reversed EMT-like features and malignant behaviors of HCC cells.
(a) HepG2 cells were transfected with miR-424-5p or miR-NC and cultured in anchorage deprived condition for 24 h before expression analysis of the EMT related markers. (b) After cultured in anchorage deprived condition for 24 h, miR-424-5p transfected HCC cells were seeded into normal 6-well plate at the density of 1,000 cells/well and incubated for 7 days. The resulting colonies were stained with crystal violet and counted. (c) MiR-424-5p transfected BEL7402, SMMC7721 and HepG2 cells were cultured in anchorage deprived condition for 24 h before invasion activities were measured. Photos were representative fields of invasive cells on the membrane. All of the presented data are representative one from at least three independent experiments. *P < 0.05,**P < 0.01, *** P < 0.001.
ICAT is a direct target of miR-424-5p
Having identified miR-424-5p as a regulator of EMT, we were then interested in identifying the target gene of miR-424-5p responsible for these effects. We evaluated the potential targets of miR-424-5p using available public algorithms15 and the target gene annotations revealed that ICAT/CTNNBIP1, a novel β-catenin-interacting protein16,17 was the predicted target of miR-424-5p. Through sequestering β-catenin, ICAT inhibits E-cadherin based cell adhesion and Wnt responsive gene expression, thus leads to an avenue for ICAT to play a role in tumorigenesis. Intriguingly, ICAT is overexpressed in some tumors, though its exact role in cancer progression has not been clarified18. Because ICAT is a regulator of E-cadherin/β-catenin complex of the cell adhesion, we hypothesized that up-regulation of ICAT in response to reduced level of miR-424-5p might be involved in EMT process of these detached HCC cells. To test this hypothesis, we firstly verified that ICAT was significantly up-regulated in detached HCC cells at both the mRNA and protein level (Fig. 5a–b). We further verified that ICAT expression was strongly inhibited by ectopic overexpression of miR-424-5p, while its expression was significantly enhanced by miR-424-5p inhibitor (Fig. 5c–e). Then we cloned a reporter construct containing the 3′-UTR of ICAT downstream of a luciferase open reading frame (Fig. 5f). This dual-luciferase reporter assay showed that co-transfection of ICAT-3′-UTR recombinant and miR-424-5p significantly reduced the relative luciferase activity in HEK293 cells, while transfection of mutant recombinant abrogated inhibition by miR-424-5p (Fig. 5g). These data suggested that ICAT was a direct target of miR-424-5p.

ICAT is a direct target of miR-424-5p.
(a–b) Expression of ICAT mRNA and protein in detached HepG2 cells was analyzed by real-time PCR (a) and western blot (b), respectively. (c) The HepG2 cells were transfected with miR-424-5p or its inhibitor; and ICAT mRNA expression was detected by real-time PCR and quantified by normalization to β-actin. (d–e) The BEL7402, SMMC7721 and HepG2 HCC cells were transfected with 50 nM of miR-424-5p or nonsense control, the endogenous protein levels of ICAT in HCC cells were detected by western blot (d) and quantified by the band intensity of ICAT normalized to β-actin (e). (f) miR-424-5p binding site is located in the ICAT/CTNNBIP1-3′-UTR and mutation was generated in the ICAT/CTNNBIP1-3′-UTR sequence by mutating 3 nucleotides recognized by miR-424-5p. Either wild-type (WT) or mutant CTNNBIP1-3′-UTR was subcloned into the dual-luciferase reporter vector. (g) The luciferase reporter vector containing WT ICAT/CTNNBIP1-3′-UTR or mutant 3′-UTR was cotransfected with miR-424-5p or miR-NC into HEK293 cells. Firefly luciferase activities were determined 48 h post-transfection and normalized to Renilla luciferase. Presented data were representative one from at least three independent experiments. *P < 0.05, **P < 0.01,***P < 0.001.
MiR-424-5p reversed EMT by directly targeting ICAT
We have shown that ectopic overexpression of miR-424-5p inhibited EMT process of these anchorage-deprived cells; then we further performed immunoprecipitation analysis to verify the direct binding between E-cadherin and β-catenin in the miR-424-5p or HA-ICAT transfected cells. Our data showed that ectopic overexpression of miR-424-5p enhanced the binding between E-cadherin and β-catenin, while ICAT overexpression inhibited their interactions (Fig. 6a–b). These data further verified that miR-424-5p-mediated downregulation of ICAT is responsible for this deregulation of E-cadherin/β-catenin complex on the cell membrane. In addition, three siRNAs specifically targeting ICAT were synthesized and western blot verification showed that Si-1 and Si-3 had significant inhibitory effect on ICAT (Fig. 6c). Thus the following experiments were done with the mixture of these two SiRNAs. Our data showed that Si-ICAT transfection significantly inhibited EMT markers in the detached HCC cells (Fig. 6d), while overexpression of ICAT rescued the effect of miR-424-5p on EMT of these detached HCC cells (Fig. 6e–f). Thus these data confirmed that deregulation of miR-424-5p, through targeting ICAT, is involved in EMT process of the detached HCC cell. Moreover, we are interested in clarifying whether the malignancy linked with EMT process was also mediated by miR-424-5p-induced downregulation of ICAT. Our data showed that knockdown of ICAT could indeed photocopy the observed effect on invasion upon overexpression of miR-424-5p to a great extent (Fig. 6g–h). These data indicated that the involvement of miR-424-5p in EMT could take effect, at least to a great degree, via directly targeting ICAT.

miR-424-5p disrupted the cell-cell adhesion complex by directly targeting ICAT.
(a) HEK293 cells were transfected with 40 nM, 80 nM and 120 nM of miR-424-5p and cultured for 24 h. Cell lysates were extracted and subjected to immunoprecipitation with anti-β-catenin antibody followed by western blot analysis with anti-E-cadherin antibody. Proteins in whole-cell lysates were used as input control for analysis by western blot with anti-E-cadherin antibody and anti-ICAT antibody, respectively. (b) HEK293 cells were transfected with 0.5 μg, 1 μg and 2 μg of HA tagged ICAT recombinant and cultured for 24 h. Cell lysates were extracted and subjected to immunoprecipitation with anti-β-catenin antibody followed by western blot analysis with anti-E-cadherin antibody. Proteins in whole-cell lysates were used as input control. Similar results were got from three independent experiments. (c) HepG2 cells were transfected with 50 nM of 3 synthesized siRNAs targeting ICAT or nonsense control.48 h after transfection, the inhibitory effect of siRNA were analyzed by western blot. (d) The HepG2 cells were transfected with a mixture of Si-1 and Si-3 targeting ICAT and cultured in anchorage deprived condition for 24 h before analysis of EMT related markers by real-time PCR. (e–f) HepG2 cells were transfected with miR-424-5p or HA-tagged ICAT recombinant and cultured in anchorage deprived condition for 24 h. Expression of E-cadherin, N-cadherin and ICAT was determined by western blot (e) and quantified by band intensity analysis of these proteins normalized to that of β-actin (f). (g–h) BEL7402, SMMC7721 and HepG2 HCC cells transfected with miR-424-5p, miR-424-5p inhibitor or Si-ICAT were cultured as anchorage deprived cells for 24 h before being harvested and analyzed for their invasion capability (g). The invaded cells were quantified and presented histogram were representative one from three independent experiments performed in duplicate (h). *P<0.05, **P < 0.01, ***P < 0.001.
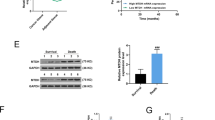
Ectopic overexpression of miR-424-5p suppressed the tumorigenesis in nude mice
To further assess the effect of miR-424-5p on HCC tumor growth in vivo, the xenografted tumor growth model was made by subcutaneous injection with SMMC7721 HCC cells in nude mice. The formed tumors were then transfected with pCMV-MIR-424-5p plasmid or mock control before the mice were sacrificed on day 18 after transplantation. Our data showed that the average size of tumors transfected with miR-424-5p plasmid was significantly smaller than those transfected with mock control on day 12 after transfection (Fig. 7a–b). The average weight of tumors in the miR-424-5p transfected group was significantly less than those in the mock control group (Fig. 7c). Quantitative real-time PCR showed that expression of miR-424-5p in tumors transfected with miR-424-5p plasmid was indeed significantly higher than those transfected with mock control (Fig. 7d); while expression of its target gene, ICAT was significantly lower in the miR-424-5p transfected tumors (Fig. 7e). Correlation analysis showed that the expression level of ICAT was negatively correlated with that of miR-424-5p (Fig. 7f), which further confirmed that ICAT was a target of miR-424-5p in HCC tissues. We further analyzed the expression of EMT related markers and our data showed that Epithelial marker E-cadherin and β-catenin was significantly upregulated while mesenchymal marker N-cadherin was significantly downregulated in miR-424-5p transfected group (Fig. 7g–i). These in vivo data was consistent with our in vitro data, showing that miR-424-5p inhibited HCC progression through its downregulation of ICAT, which further inhibited EMT through maintaining the E-cadherin/β-catenin complex on the cell membrane.

miR-424-5p inhibited the tumorigenicity of HCC cells in vivo in BALB/c nude mice.
(a) The 5-week BALB/c nude mice were subcutaneously injected with SMMC7721 cells. Images presented were the isolated tumors transfected with miR-424-5p plasmid or mock control on day 12 after transfection. (b–c) The volume (b) and weight(c) of the miR-424-5p plasmid or mock control transfected tumors were assayed on day 18 after transplantation of HCC cells. (d) Expression of miR-424-5p in the miR-424-5p plasmid or mock control transfected HCC tumors was analyzed by real-time PCR and normalized to U6. (e) Expression of ICAT in HCC tumors transfected with the miR-424-5p or mock plasmid was analyzed by real-time PCR and normalized to β-actin. (f) The correlation analysis between the expression levels of miR-424-5p and ICAT. (g–i) The expression of E-cadherin(g), N-cadherin(h) and β-catenin(i) in the HCC tumors transfected with miR-424-5p plasmid or mock control. *P<0.05, **P<0.01.
Downregulation of miR-424-5p in HCC tissues was significantly correlated with advanced disease progression in HCC patients
To investigate the role of miR-424-5p in HCC patients, we analyzed the association between miR-424-5p expression and clinical pathological features of hepatocacinoma patients. Our data showed that miR-424-5p expression was significantly reduced in the liver cancer tissues compared with that of the corresponding non-cancerous liver tissues (Fig. 8a); while the expression of ICAT was significantly upregulated in cancer tissue compared with that of the non-cancerous liver tissues (Fig. 8b). Spearman Correlation analysis showed that in both cancer tissues and non-cancerous liver tissues, expression of miR-424-5p was negatively correlated with expression of ICAT (Fig. 8c–d). Because EMT process is tightly linked with metastasis, expression of miR-424-5p and ICAT was also investigated in the HCC tissues from metastatic patients and non-metastatic patients. Our data showed that miR-424-5p was significantly downregulated while ICAT was significantly upregulated in the HCC tissues from metastatic patients compared with those from non-metastatic patients (Fig. 8e–f). Expression of miR-424-5p and ICAT was significantly negatively correlated in HCC tissues from both metastatic patients and non-metastatic patients (Fig. 8g–h). Statistical analysis showed that the more advanced HCC patients usually have lower miR-424-5p expression (Table 1). In addition, the expression of miR-424-5p was positively correlated with epithelial markers including E-cadherin and β-catenin while negatively correlated with ICAT expression (Table 2). These data further demonstrated that loss of miR-424-5p expression contributed to EMT related behaviors of HCC cells, probably through decreasing ICAT expression.

Clinical investigation of miR-424-5p in the liver cancer tissues and sera samples from HCC patients.
(a–b) The expression of miR-424-5p (a) and ICAT (b) in HCC tissues and corresponding non-cancerous liver tissues were determined by real-time PCR. (c–d) Correlation between the expression of miR-424-5p and ICAT in both cancer tissues(d) and non-cancerous tissues (d) were analyzed by Spearman correlation analysis. (e–f) Expression of miR-424-5p(e) and ICAT(f) in metastatic HCC tissues and non-metastatic HCC tissues were determined by real-time PCR. (g–h) Correlation between the expression of miR-424-5p (g) and ICAT (h) in metastatic HCC tissues and non-metastatic tissues were analyzed. (i) miR-424-5p expression in the sera from metastatic HCC patients, non-metastatic HCC patients and healthy controls were analyzed by real-time PCR. (j) In those patients whose sera and liver tissue specimen were both available, expression of miR-424-5p in both the sera and liver tissues were detected; and the correlation of miR-424-5p expression in sera and that in liver tissues were further analyzed. *P<0.05, **P<0.01,***P<0.001.
Downregulation of miR-424-5p in sera from clinical metastatic liver cancer patients
It is well recognized that human sera and other body fluids are rich resources of circulating mature miRNAs, which is closely linked with disease progression and pathological status19. These miRNAs are strikingly stable under harsh conditions including boiling, low/high pH, extended storage and multiple freeze-thaw cycles20. Since our previous data have identified miR-424-5p as an anti-oncomir to suppress tumor metastasis, we are interested in defining whether it also has indicative value in the human sera to predict disease progression and metastatic status of HCC patients. Thus we detected the expression of miR-424-5p in the sera from metastatic patients, non-metastatic patients and healthy controls. Our data showed that expression miR-424-5p was significantly downregulated in sera from liver cancer patients compared with those from healthy controls and its expression in metastatic cancer patients is significantly decreased compared with that of the non-metastatic cancer patients (Fig. 7I). Furthermore, those patients with poorer tumor differentiation and advanced metastatic status were prone to have lower expression of miR-424-5p (Table 3).In those patients whose HCC tissues and sera samples were both available, spearman correlation analysis was conducted. Correlation analysis showed that expression levels of miR-424-5p in the HCC tissues and in sera were statistically correlated(Fig. 7J), which further demonstrated that miR-424-5p played a significant role in HCC progression. These results verified our in vitro and in vivo data and further confirmed the anti-oncogenic function of miR-424-5p in HCC patients.
Discussion
Metastatic disease is responsible for the majority of cancer related deaths. In the metastatic process, escaping from detachment induced cell death is the prerequisite for disseminated cancer cells to survive in the circulating system and further lodge at secondary organs to form a metastatic cancer lesion1,21. EMT processes, characterized by diminished epithelial characteristics and increased mesenchymal attributes, are believed to facilitate cancer metastasis3,4. More and more evidences showed that resistance to anoikis and EMT are tightly linked, though the involved mechanisms are not clarified. In this work, we showed for the first time that acquisition of anoikis-resistance, accompanied with miR-424-5p deregulation, conferred hepatocarcinoma cells with EMT related changes.
The present study showed that miR-424-5p was significantly downregulated in the anoikis-resistant hepatocarcinoma cells while compared with their attached counterpart. In coordination with anchorage deprival, an EMT process was observed in the detached anoikis-resistant HCC cells. Meanwhile, downregulation of miR-424-5p in the detached HCC cells was accompanied with a significant upregulation of its predicted target gene, ICAT. Ectopic overexpression of miR-424-5p in the anchorage-deprived hepatocarcinoma cells reversed EMT process through up-regulation of ICAT, which further induced E-cadherin/β-catenin complex restoration on the cell membrane. Further luciferase reporter gene analysis revealed that ICAT is a direct target of miR-424-5p. RNAi-mediated knockdown of ICAT decreased E-cadherin/β-catenin complex on the HCC cell membrane and inhibited EMT features of anoikis-resistant HCC cells, which properly photocopied the effect of miR-424-5p. The in vivo study using the xenograft tumor model in nude mice supported our in vitro data and confirmed the anti-oncogenic role of miR-424-5p in HCC and its negative regulation of its target, ICAT. Furthermore, our clinical investigation showed that miR-424-5p was significantly downregulated in HCC tissues and sera from metastatic patients and the decreased expression of miR-424-5p were significantly correlated with poorer tumor differentiation and more advanced clinical stages. All of our in vitro, in vivo and clinical data indicated that aberrant expression of miR-424-5p was critically involved in HCC progression.
It is more and more accepted that oncogenic EMT plays an important role in tumor progression22, thus the focus of the present study is to understand the molecular basis of the tight correlation between resistance to anoikis and the oncogenic EMT. Our data showed that anoikis-resistant process of HCC cells was also accompanied with EMT process, while miR-424-5p could prevent HCC progression by reversing both of these two critical processes involved in metastasis. In consideration of our previous data showing that acquisition of resistance to anoikis also confers HCC cell more invasive phenotype and act as a selective pressure for the metastasis of HCC9, it is interesting to see whether miR-424-5p could also reverse accompanied malignant phenotype during acquisition of resistance to anoikis. Indeed, our data showed that miR-424-5p inhibited the invasion and colony formation capability of anoikis-resistant HCC cells as well, which further verified its anti-oncogenic role in preventing HCC progression.
MiR-424 is located on human chromosome Xq26.3 and has recently been classified in a large cluster together with miR-15/miR-1623. The role of miR-424 has been investigated in several types of physiological and pathological conditions, which revealed the diverse function of miR-424 in different situations. It was reported that miR-424 was implicated in maintaining the physiology of vascular remodeling and angiogenesis24, remodeling of the epithelium in the mammary gland25, protecting against permanent focal cerebral ischemia injury26, promoting differentiation of monocytes27 and skeletal muscle-derived progenitor cells28. Recently, an interesting study showed that miR-424 was involved in APLN-dependent miRNA-FGF signaling axis needed for the maintenance of pulmonary vascular homeostasis29. Thus, the role of miR-424 in differentiation and maintaining tissue homeostasis has been well established. A recent report showed that miR-424, together with 6 other microRNAs could induced hMSC to converse into functionally mature hepatocytes30, which identified its physiological role in liver and provided the foundation for understanding that its abnormal expression is related with morbidity, such as liver cancer.
In recent years, multiple investigations have reported the role of miR-424 in cancer. Downregulation of miR-424 has been observed in cervical cancer31, ovarian cancer13, colon cancer32, lung cancer33 and it act as tumor suppressor to inhibit cancer progression in these investigated cancers. Whereas miR-424-5p was reported to be up-regulated and act as a tumor promoter in pancreatic cancer cells34. These functional discrepancies probably are because of the different roles that miR-424 plays in different tissues and organs. In the revision time of this study, an investigation reported that miR-424 suppressed malignant behaviors of the primary HCC cells, which is consistent with our data (Online supplement materials) and supported our proposal that miR-424-5p act as a suppressor of liver cancer. However, their study focused on the role of miR-424 in primary HCC and their identified target is c-Myb; whereas our study focused on the effect of miR-424-5p on anchorage-independent metastatic HCC and the identified target is ICAT. In this study, we demonstrated for the first time that miR-424-5p inhibited EMT process and HCC progression by targeting ICAT in anchorage-independent hepatocellular carcinoma cells.
A common hallmark of EMT is the breakdown of expression or function of E-cadherin35, which further disrupts the E-cadherin/β-catenin complex and suffices to circumvent anoikis in some contexts. This E-cadherin/β-catenin complex plays a crucial role in epithelial cell-cell adhesion and maintenance of tissue architecture. The direct target of miR-424-5p, ICAT is a potent inhibitor of β-catenin, which can inhibit the E-cadherin/β-catenin interface and disrupt the intercellular adhesion function of the cells16,36. β-catenin is crucial for establishing a connection between E-cadherin and actin filaments and maintaining epithelial properties. Both of our in vitro and in vivo data showed that miR-424-5p could maintain the stability of E-cadherin/β-catenin complex by directly targeting ICAT, the β-catenin inhibitor.
Though ICAT is found to be significantly deregulated in several human tumors18,37, whether it is involved in tumorigenesis has not been clarified. Our data showed that miR-424-5p could block EMT process of anchorage-independent HCC cells by directly targeting ICAT and disrupting the combination of E-cadherin/β-catenin complex. Perturbation of this complex further caused loss of epithelial properties and appearance of mesenchymal characteristics, therefore allowed release of single cell due to weak cell-cell adhesion and conferred invasive properties to tumor cells38, which is accompanied with EMT process. Altogether, ICAT contributed to EMT by inhibiting the adherence of β-catenin with E-cadherin, whereas miR-424-5p reversed this EMT process by directly targeting ICAT. Thus, our data showed for the first time that in the metastatic process, miR-424-5p reversed EMT process of anoikis-resistant HCC cell by directly targeting ICAT and further reconstituted the E-cadherin/β-catenin complex on the cell membrane. A recent study proposed that miR-424-5p promoted mesenchymal-epithelial transition in a metastatic prostate cancer model39, which is obviously in discrepancy with our data. These seemingly conflicting observations could be due to the distinct functions of miR-424-5p in different tissues and cell types. One of the distinguished characteristics of microRNA is its space and time specific functions, which may explain these divergent and even contrasting reports of the same microRNA.
In summary, the present results showed that miR-424-5p was significantly downregulated in metastatic anchorage-independent HCC cells, which was accompanied by EMT process and metastatic properties. Moreover, clinical investigation indicated that miR-424-5p expression was significantly decreased in both liver cancer tissues and sera from metastatic HCC patients compared with those from non-metastatic HCC patients. We also observed that the patients with higher pathological grades and more advanced TNM stages were prone to have lower expression levels of miR-424-5p. These data, together with our data showing restoration of miR-424-5p reversed malignant behaviors of HCC, further verified the anti-cancer function of miR-424-5p in HCC. In conclusion, our results identified a crucial anti-oncogenic role of miR-424-5p in the liver cancer progression and suggested that miR-424-5p might be a potential prognostic predictor and a therapeutic target for HCC patients.
Methods
Patients, liver tissues and sera samples
For the liver tissue analysis, pathologically confirmed HCC specimens were obtained from 50 HCC patients undergoing curative resections in Provincial Hospital Affiliated to Shandong University from April 2012 to May 2013. No patients received any preoperative anticancer treatment. Cell differentiation based on tumor grading was determined by the Edmondson Grading System. Clinical stages were classified according to the International Union against Cancer TNM classification system. Summary of the clinical parameters of these patients were shown in Table 1. For miR-424-5p expression analysis in the sera, 62 HCC patients attended the Provincial Hospital Affiliated to Shandong University from April 2012 to September 2013 were included in this study. Tumor grades and disease stages were determined by Edmondson Grading System and International Union against Cancer TNM classification system, respectively. The clinical characteristics of these patients were shown in Table 3. Sera from 40 age and gender matched healthy persons were analyzed as healthy controls. The surgically excised liver tissues and collected sera specimen were put in −80°C freezer until use and analysis. This study was performed with ethical approval of the Human Ethics Committee of Shandong University in accordance with the Declaration of Helsinki and written informed consents were obtained from all of the enrolled subjects.
Cell model construction and transfection
BEL7402, SMMC7721 and HepG2 hepatocarcinoma cells were cultured in RPMI1640 medium supplemented with 10% fetal bovine serum. Anchorage-deprived cell model was constructed as previously described9,10. Human HEK293 cells were obtained from American Type Culture Collection and were cultured at 37°C with 5% CO2 in DMEM supplemented with 10% FCS. The transient transfections for cell models were performed with Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, USA). For miRNA transfection experiments, exponentially growing HCCs were plated in 6-well plates at a density of 2 × 105 cells/well and were grown overnight to 80% confluence. An miRNA mimic/inhibitor or a nonsense control (GenePharma Co., Shanghai, China) was put to the culture medium at a final concentration of 100 nM according to the manufacturer's recommendations. For the ICAT/CTNNBIP1 inhibition and overexpression studies, siRNA (GenePharma Co., Shanghai, China) against ICAT or HA-tagged ICAT plasmid (a kind gift from Dr. Tetsu Akiyama from University of Tokyo) was introduced to the HCC cells at the final concentration of 50 nM according to the protocol.
Isolation of microRNA from HCC cells, liver tissues and sera
For microRNA isolation from HCC cells and tissues, microRNAs were prepared with an All-in-One™ miRNA qRT-PCR Detection Kit (GeneCopoeia, Maryland, USA) according to the manufacturer's instructions. For microRNA isolation from sera samples of HCC patients, sera samples were collected and stored at −80°C freezer until use. MicroRNA was isolated from 0.4 mL of serum using miRNA isolation kit (Bioteke, Beijing, China) according to the manufacturer's protocol. Purity and concentration of RNA were determined by using a dual beam UV spectrophotometer (Eppendorf AG, Hamburg, Germany).
Real-time PCR analysis of mRNA and microRNA
Real-time PCR analysis of mRNA of HCC cells and tissues was done as previously described40. For real-time PCR analysis of microRNA expression in liver tissues and HCC cells, microRNAs were prepared with an All-in-One™ miRNA qRT-PCR Detection Kit (GeneCopoeia, Maryland, USA) according to the manufacturer's instructions. U6 was used as an endogenous normalization control for miRNA detection. The PCR cycling conditions were as follows: pre-denaturing at 95°C for 10 min, followed by 40 cycles of denaturing at 95°C for 10 s, annealing at 58°C for 20 s and extension at 72°C for 10 s.
Computational target prediction and luciferase reporter assays
The computational target prediction tools, including TargetScan (http://www.targetscan.org), PicTar (http://www.pictar.mdc-berlin.de) and miRanda (http://www.microrna.org) were used to assess the potential targets of miR-424-5p. The human ICAT 3′-UTR sequence was ligated into the pMIR-GLO luciferase vector (Promega, Madison, WI, USA) to get pMIR-ICAT-3′-UTR recombinant. Another pMIR-GLO luciferase construct containing the ICAT 3′-UTR sequence with a mutation in the putative miR-424-5p seed region (UGCUGCU to UGACGCU) was generated as a negative control and named Mut-ICAT-3′-UTR. ICAT-3′-UTR primers were as follows: forward 5′-CCAAGTGCGCTTTGCAATA-3′; reverse 5′-ACTCTCCAGGTGGACCCTCA-3′. Mutagenesis primers for the miR-424-5p target site in the ICAT 3′-UTR were as follows: forward 5′-AAAGAACTTGCGCCTGCCTTCACAT-3′; reverse 5′-ATGTGAAGGCAGGCGCAAGTTCTTT-3′. HEK-293 cells were plated in 96-well plates and allowed to reach 60%–80% confluence before being co-transfected with a reporter construct (pMIR-GLO REPORT plasmid, pMIR-GLO-ICAT 3′-UTR, pMIR-GLO-ICAT 3′-UTR-Mut) and miR-424-5p. Cells co-transfected with pMIR-GLO-ICAT-3′-UTR plasmid and a non-specific sequence of miRNA was used as a mock control. After 24 hours of incubation, cells were harvested and luciferase activities were measured using the Dual-Luciferase Reporter Assay System(Promega, Madison, WI, USA)according to the manufacturer's recommendations. Data were normalized for transfection efficiency by division of firefly luciferase activity with that of Renilla luciferase.
Immunoprecipitation and western blot analysis
For immunoprecipitation (IP), whole-cell extracts were collected and lysed in IP buffer (Merck, New Jersey, USA). After centrifugation at 14,000 rpm for 10 min, supernatants were collected and incubated with protein G plus-agrose immunoprecipitation reagent together with anti-β-catenin antibody (proteintech, Chicago, USA). After 16 h of incubation, beads were washed five times with IP buffer. Immunoprecipitates were eluted by boiling with 1% (wt/vol) SDS sample buffer. Western blot analysis was done as described before40 using specific antibodies against ICAT(Epitomics, Burlingame, CA),β-catenin(proteintech, Chicago, USA),E-cadherin(BD Biosciences, San Jose, CA), N-cadherin(BD Biosciences, San Jose, CA), snail(Immunoway, Newark, USA), fibronectin (BD Biosciences, San Jose, CA) and vimentin (Cell Signaling Technology, Beverly, USA). Expression of β-actin was assessed as an internal control using its specific antibody (Abcam, Cambridge, MA).
Cell viability and apoptosis assay
Cell viabilities were analyzed by CCK-8 kit (Dojindo Laboratories, Kumamoto, Japan) and trypan blue staining, respectively. For apoptosis assay, HCC cells were analyzed with Annexin-V apoptosis assay kit (Biouniquer, Nanjing, China) according to the manufacture's assay. Apoptosis marker, caspase 3 activities were detected by Caspase-3/CPP32 Colorimetric Assay Kit (BioVision, CA, USA) according to its protocol; and cleavage of caspase 3 was detected by western blot using its specific antibody(Cell Signaling Technology, Beverly, USA).
Colony formation and cell invasion assay
BEL7402, SMMC7721 and HepG2 cells were plated in poly-HEMA-coated plates and further cultured for another 24 h before being transferred to the normal 6-well plates at a density of 1,000 cells/well. For colony formation assay, plates were incubated for 7 days and the resulting colonies were stained with crystal violet. Only colonies containing more than 50 cells were regarded as positive colonies and were counted under the microscope. Invasion assay of HCC cells was tested using a transwell system as previously described9.
In vivo tumor growth assay
5-week-old immunodeficient male BALB/c athymic nude mice (Huafukang Biotechnology Ltd, Beijing, China) were bred in aseptic conditions and maintained in Shandong University Experimental Animal Center. All mice were subcutaneously injected with SMMC7721 HCC cells (1 × 107 cells) in the left flank region. When visible tumor appeared, the mice were randomly divided into miR-424-5p transfection group and mock control group. Tumors were injected with 10 μg of pCMV-miR-424-5p plasmid (Origene Technologies, Rockville, MD) or empty pCMV vector control in 50 μl transfection reagent (Polyplus-transfection Inc, New York, USA) according to the manufacturer's instructions. This injection was performed once every 3 days for a total of 12 days before the mice were sacrificed and the tumors were isolated. The tumor volume (V) was obtained by measuring the length (L) and width (W) with a caliper and calculated with the formula: Volume (mm3) = [width2 (mm2) × length (mm)]/2. Mice were maintained in accordance with guidelines of the Institutional Animal Care and Use Committee and all the animal studies performed were approved Shandong University Institutional Animal Care and Use Committee.
Statistical analysis
Data were assessed using software packages SPSS version 16.0 (SPSS Inc., IL, USA). χ2 tests were applied to compare categorical variables and continuous variables were analyzed using the t test. Spearman's rank correlation test was used to evaluate correlations between variables. A P value < 0.05 was considered statistically significant and a two-tailed P value was used in all analyses.
References
Simpson, C. D., Anyiwe, K. & Schimmer, A. D. Anoikis resistance and tumor metastasis. Cancer Lett 272, 177–185 (2008).
Polyak, K. & Weinberg, R. A. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nature reviews. Cancer 9, 265–273 (2009).
Zeisberg, M. & Neilson, E. G. Biomarkers for epithelial-mesenchymal transitions. J Clin Invest 119, 1429–1437 (2009).
Kalluri, R. & Weinberg, R. A. The basics of epithelial-mesenchymal transition. J Clin Invest 119, 1420–1428 (2009).
Bandres, E., Agirre, X., Ramirez, N., Zarate, R. & Garcia-Foncillas, J. MicroRNAs as cancer players: potential clinical and biological effects. DNA and cell biology 26, 273–282 (2007).
Schoof, C. R., Botelho, E. L., Izzotti, A. & Vasques Ldos, R. MicroRNAs in cancer treatment and prognosis. Am J Cancer Res 2, 414–433 (2012).
Kong, Y. W., Ferland-McCollough, D., Jackson, T. J. & Bushell, M. microRNAs in cancer management. Lancet Oncol 13, e249–258 (2012).
Gregory, P. A. et al. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nature cell biology 10, 593–601 (2008).
Cao, L. et al. Involvement of anoikis-resistance in the metastasis of hepatoma cells. Exp Cell Res 315, 1148–1156 (2009).
Zhang, Z. et al. Acquisition of anoikis resistance reveals a synoikis-like survival style in BEL7402 hepatoma cells. Cancer Lett 267, 106–115 (2008).
Chen, Y. C. et al. Oct-4 expression maintained cancer stem-like properties in lung cancer-derived CD133-positive cells. PloS one 3, e2637 (2008).
Long, X. H. et al. Tumor suppressive microRNA-424 inhibits osteosarcoma cell migration and invasion via targeting fatty acid synthase. Exp Ther Med 5, 1048–1052 (2013).
Xu, J. et al. Suppressed miR-424 expression via upregulation of target gene Chk1 contributes to the progression of cervical cancer. Oncogene 32, 976–987 (2013).
Faraoni, I. et al. MiR-424 and miR-155 deregulated expression in cytogenetically normal acute myeloid leukaemia: correlation with NPM1 and FLT3 mutation status. J Hematol Oncol 5, 26 (2012).
Miranda, K. C. et al. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 126, 1203–1217 (2006).
Gottardi, C. J. & Gumbiner, B. M. Role for ICAT in beta-catenin-dependent nuclear signaling and cadherin functions. Am J Physiol Cell Physiol 286, C747–756 (2004).
Graham, T. A., Clements, W. K., Kimelman, D. & Xu, W. The crystal structure of the beta-catenin/ICAT complex reveals the inhibitory mechanism of ICAT. Mol Cell 10, 563–571 (2002).
Koyama, T. et al. Mutation and expression of the beta-catenin-interacting protein ICAT in human colorectal tumors. Jpn J Clin Oncol 32, 358–362 (2002).
Wu, W. K. et al. MicroRNA dysregulation in gastric cancer: a new player enters the game. Oncogene 29, 5761–5771 (2010).
Chen, X. et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 18, 997–1006 (2008).
Luanpitpong, S., Iyer, A. K., Azad, N., Wang, L. & Rojanasakul, Y. Nitrosothiol Signaling in Anoikis Resistance and Cancer Metastasis. For Immunopathol Dis Therap 3, 141–154 (2012).
Frisch, S. M., Schaller, M. & Cieply, B. Mechanisms that link the oncogenic epithelial-mesenchymal transition to suppression of anoikis. J Cell Sci 126, 21–29 (2013).
Chamorro-Jorganes, A. et al. MicroRNA-16 and microRNA-424 regulate cell-autonomous angiogenic functions in endothelial cells via targeting vascular endothelial growth factor receptor-2 and fibroblast growth factor receptor-1. Arterioscler Thromb Vasc Biol 31, 2595–2606 (2011).
Ghosh, G. et al. Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF-alpha isoforms and promotes angiogenesis. J Clin Invest 120, 4141–4154 (2010).
Llobet-Navas, D. et al. The miR-424(322)/503 cluster orchestrates remodeling of the epithelium in the involuting mammary gland. Genes Dev 28, 765–782 (2014).
Zhao, H. et al. MiRNA-424 protects against permanent focal cerebral ischemia injury in mice involving suppressing microglia activation. Stroke 44, 1706–1713 (2013).
Rosa, A. et al. The interplay between the master transcription factor PU.1 and miR-424 regulates human monocyte/macrophage differentiation. Proc Natl Acad Sci U S A 104, 19849–19854 (2007).
Oishi, T. et al. Osteogenic differentiation capacity of human skeletal muscle-derived progenitor cells. PLoS One 8, e56641 (2013).
Kim, J. et al. An endothelial apelin-FGF link mediated by miR-424 and miR-503 is disrupted in pulmonary arterial hypertension. Nat Med 19, 74–82 (2013).
Cui, L. et al. A set of microRNAs mediate direct conversion of human umbilical cord lining-derived mesenchymal stem cells into hepatocytes. Cell Death Dis 4, e918 (2013).
Cheung, T. H. et al. Dysregulated microRNAs in the pathogenesis and progression of cervical neoplasm. Cell Cycle 11, 2876–2884 (2012).
Oneyama, C. et al. MiR-424/503-mediated Rictor upregulation promotes tumor progression. PLoS One 8, e80300 (2013).
Berghmans, T. et al. Identification of microRNA-based signatures for response and survival for non-small cell lung cancer treated with cisplatin-vinorelbine A ELCWP prospective study. Lung Cancer 82, 340–345 (2013).
Wu, K. et al. MicroRNA-424-5p suppresses the expression of SOCS6 in pancreatic cancer. Pathol Oncol Res 19, 739–748 (2013).
Zavadil, J., Haley, J., Kalluri, R., Muthuswamy, S. K. & Thompson, E. Epithelial-mesenchymal transition. Cancer research 68, 9574–9577 (2008).
Stow, J. L. ICAT is a multipotent inhibitor of beta-catenin. Focus on “role for ICAT in beta-catenin-dependent nuclear signaling and cadherin functions”. Am J Physiol Cell Physiol 286, C745–746 (2004).
Imai, M., Nakamura, T., Akiyama, T. & Horii, A. Infrequent somatic mutations of the ICAT gene in various human cancers with frequent 1p-LOH and/or abnormal nuclear accumulation of beta-catenin. Oncol Rep 12, 1099–1103 (2004).
Nelson, K., Helmstaedter, V. & Lage, H. The influence of tamoxifen on growth behavior and cell-cell adhesion in OSCC in vitro. Oral oncology 43, 720–727 (2007).
Banyard, J. et al. Regulation of epithelial plasticity by miR-424 and miR-200 in a new prostate cancer metastasis model. Sci Rep 3, 3151 (2013).
Wei, Q. et al. Deregulation of the NLRP3 inflammasome in hepatic parenchymal cells during liver cancer progression. Lab Invest 94, 52–62 (2014).
Acknowledgements
We thank Dr. Tetsu Akiyama from University of Tokyo for providing the HA-tagged ICAT plasmid. This study was supported by the National Nature Science Foundation of China (No. 81172352, No.81472269 and No. 30700357) and the Science and Technology Development Project of Shandong Province (No.2011GGE27020 and No.BS2013YY037).
Author information
Authors and Affiliations
Contributions
L.H. and Y.Z. designed the study, analyzed the data and wrote the manuscript. Y.Z. performed most of the experiments and prepared the figures. T.L. and X.J. collected the clinical specimen and analyzed the clinical data. L.H. and P.G. did the xenograft tumor model construction and analysis. Q.W. and J.K. contributed to the real-time PCR analysis. W.Z. contributed to the data interpretation and immunoprecipitation analysis. W.H., Y.Q. and L.S. participated in the microRNA isolation from liver tissues and sera of HCC patients. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Online Supplement data
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Zhang, Y., Li, T., Guo, P. et al. MiR-424-5p reversed epithelial-mesenchymal transition of anchorage-independent HCC cells by directly targeting ICAT and suppressed HCC progression. Sci Rep 4, 6248 (2014). https://doi.org/10.1038/srep06248
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06248
This article is cited by
-
Recent insights into the microRNA-dependent modulation of gliomas from pathogenesis to diagnosis and treatment
Cellular & Molecular Biology Letters (2022)
-
The E3 ubiquitin ligase MG53 inhibits hepatocellular carcinoma by targeting RAC1 signaling
Oncogenesis (2022)
-
CircCBFB is a mediator of hepatocellular carcinoma cell autophagy and proliferation through miR-424-5p/ATG14 axis
Immunologic Research (2022)
-
The potential roles and mechanisms of non-coding RNAs in cancer anoikis resistance
Molecular and Cellular Biochemistry (2022)
-
LINC00922 promotes the proliferation, migration, invasion and EMT process of liver cancer cells by regulating miR-424-5p/ARK5
Molecular and Cellular Biochemistry (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
