
Abstract
The Early Jurassic and Cretaceous deposits of India are known for their diverse sauropod fauna, while little is known from the Middle and Late Jurassic. Here we report the first ever remains of a dicraeosaurid sauropod from India, Tharosaurus indicus gen. et sp. nov., from the Middle Jurassic (early–middle Bathonian) strata of Jaisalmer Basin, western India. Known from elements of the axial skeleton, the new taxon is phylogenetically among the earlier-diverging dicraeosaurids, and its stratigraphic age makes it the earliest known diplodocoid globally. Palaeobiogeographic considerations of Tharosaurus, seen in conjunction with the other Indian Jurassic sauropods, suggest that the new Indian taxon is a relic of a lineage that originated in India and underwent rapid dispersal across the rest of Pangaea. Here we emphasize the importance of Gondwanan India in tracing the origin and early evolutionary history of neosauropod dinosaurs.
Similar content being viewed by others


Introduction
Sauropods, a speciose group of saurischian dinosaurs that dominated the terrestrial landscapes until the end-Cretaceous1,2, are characterized by a small skull, elongated neck and tail, columnar limbs, and a quadrupedal gait3. Within Sauropoda, Dicraeosauridae represents a clade of small-bodied diplodocoids that are known for their distinctive vertebral morphology with long paired neural spines4,5,6,7. Dicraeosaurids range in age from the Middle Jurassic–Early Cretaceous and are mostly known from the Gondwanan landmasses of Africa and South America, besides a few Laurasian occurrences in the USA and China3,8. Amongst the well-known dicraeosaurids are Dicraeosaurus from East Africa and Brachytrachelopan, Amargasaurus, Bajadasaurus, Pilmatueia and Amargatitanis from Argentina4,5,8,9,10,11, Suuwassea, Kaatedocus and Smitanosaurs from USA12, and Lingwulong from China13.
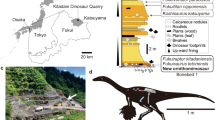
In India, early-diverging sauropods Barapasaurus and Kotasaurus are known from the Early Jurassic Kota Formation of Pranhita–Godavari Basin, whereas putative Middle Jurassic camarasauromorph remains occur in Kutch2,14,15,16,17,18. No diplodocoid sauropods are yet known from India18,19. Here we report on the discovery of a new dicraeosaurid from the Jaisalmer Formation, Rajasthan, western India (Fig. 1, Supplementary Note 1). Fossils were collected from a shale unit situated at the base of the early–middle Bathonian Fort Member20,21 and include disarticulated, but associated, specimens of the axial skeleton spread over an area of ~ 25 m2 (L1–L3, Fig. 1). This discovery provides new insights into sauropod diversity of the Indian Gondwana, with important implications for the origin and dispersal of Neosauropoda.

Geological map of Jaisalmer Basin showing (a) the fossil locality; (b) stratigraphic column showing the position of the dinosaur fossil yielding horizon; (c) photograph of the fossil site. The map and stratigraphic column were drawn by K.K. using CorelDRAW 2019 (Version number: 21.0.0.593, URL link: http://www.corel.com/en/).
Results
Systematic palaeontology
Sauropoda Marsh, 1878
Neosauropoda Bonaparte, 1986
Diplodocoidea Marsh, 1884
Dicraeosauridae Janensch, 1929
Tharosaurus indicus gen. et sp. nov.
Etymology
Generic name is a combination of Tharo, referring to the ‘Thar desert’ of western India where the type specimen was found, and saurus, which is derived from the Greek word ‘sauros’ meaning lizard; specific name is for the country of origin i.e., India.
Holotype
RWR-241A–K (Palaeontology Division, Geological Survey of India, Western Region, Jaipur, Rajasthan, India); partial middle/posterior cervical vertebra; middle/posterior cervical anterior condyle and right prezygapophyses; partial anterior dorsal neural arch; middle/posterior neural spines; anterior dorsal rib; partial anterior and middle caudal vertebrae (Figs. 2, 3, 4, 5; Supplementary Table 1).

Cervical vertebrae (CV6/8) of Tharosaurus indicus. (a) RWR-241-A, anterior cotyle in anterior view. (b–k) RWR-241-B, partial vertebra, photographs and line drawings in (b,c) right lateral view, red line indicates U-shaped ridge demarcating anterior and posterior halves of lateral pneumatic fossa; (d,e) left lateral view; (f,g) ventral view, red line indicates posteriorly bifurcated midline keel and arrow indicates accessory ridge; (h,i) posterior view, arrows and red arrowheads indicate deep bifurcation of neural arch and triangular facets below cotyle, respectively. (j,k) dorsal view, arrowhead indicates passage enclosed by bifid neural arch and ligament scars and striations marked in red and purple, respectively. Broken areas and artefacts in grey and pink, respectively. c centrum, cpof centropostzygapophyseal fossa, cpol centropostzygapophyseal lamina, lf lateral fossa, lvf lateroventral flange, mk midline keel, na neural arch, nc neural canal, pvf posteroventral fossa, tpol intrapostzygapophyseal lamina. Scale bars represent 50 mm.

Right cervical prezygapophysis (CV6/8) of Tharosaurus indicus. RWR-241-C, photographs and line drawings in (a,b) lateral view; (c,d); medial view; (e,f) anterior view; (g) dorsal view. Arrowheads indicate transverse sulcus beneath dorsal articular surface of prezygapophysis. cprf centroprezygapophyseal fossa, epi pre-epipophysis, lcprl lateral branch of centroprezygapophyseal lamina, mcprl medial branch of centroprezygapophyseal lamina, nc neural canal, tprl intraprezygapophyseal lamina. Scale bars represent 50 mm.

Dorsal vertebrae of Tharosaurus indicus. RWR-241-F, partial anterior dorsal neural arch, photographs and line drawings in (a,b) anterior view; (c,d) posterior view; (e,f) anterior view. RWR-241-G, partial middle/posterior dorsal neural arch-spine complex, photographs and line drawings in (g,h) anterior view; (i,j) posterior view; (k,l) lateral view. RWR-241-I, nearly complete anterior dorsal rib in (m) anterior view; (n) posterior view. acdl anterior centrodiapophyseal lamina, ca capitulum, cdf centrodiapophyseal fossa, da diapophysis, ns neural spine, pcdl posterior centrodiapophyseal lamina, podl postzygodiapophyseal lamina, posdf postzygapophyseal spinodiapophyseal fossa, prcdf prezygapophyseal centrodiapophyseal fossa, prsl prespinal lamina, spdl spinodiapophyseal lamina, spol spinopostzygapophyseal lamina, sprl spinoprezygapophyseal lamina, sprf spinoprezygaposphyseal fossa, tp transverse process, tu tuberculum. Scale bars represent 50 mm.

Caudal vertebrae of Tharosaurus indicus. RWR-241-J, partial anterior caudal vertebra in (a) anterior view; (b) right lateral view; (c) ventral view. RWR-241-K, middle caudal centrum in (d) anterior view; (e) posterior view; (f) left lateral view; (g) ventral view. c centrum, chf chevron facet, lf lateral fossa, lpfo lateral pneumatic foramen, vf ventral fossa, vr ventrolateral ridge.
Horizon and locality
Fort Member of the Jaisalmer Formation (early–middle Bathonian), Jethwai village, District Jaisalmer, Rajasthan state, western India.
Diagnosis
Tharosaurus exhibits a unique combination of the following characters: middle/posterior cervical centroprezygapophyseal lamina divided into lateral and medial branches, the latter connecting with the intraprezygapophyseal lamina (shared with all dicraeosaurids); middle/posterior cervical centropostzygapophyseal fossa elliptical and bordered laterally by pillar-like centropostzygapophyseal lamina (shared with the dicraeosaurids Lingwulong, Brachytrachelopan, and Pilmatueia); deep bifurcation of cervical neural arch extending up to the dorsal margin of the neural canal (shared with the dicraeosaurids Amargasaurus and Pilmatueia); paired fossae on the ventral surface of the middle/posterior cervical centrum separated by a mid-line keel (shared with all dicraeosaurids, barring Smitanosaurus, Kaatedocus and Suuwassea, and the diplodocids—Barosaurus and Dinheirosaurus); paired ventral fossae extending up to the posterior margin of centrum; anterior condyle of middle/posterior cervical more rugose compared to the rest of the centrum (shared with the dicraeosaurid Kaatedocus); bifid middle/posterior cervical neural spine (shared with Flagellicaudata); divided lateral fossa/pleuroceol on cervical centra (shared with Flagellicaudata); lateral fossa divided into deep posterior and shallow anterior half by weak ridge on the ventral surface of the fossa; prominent lateroventral flanges on middle/posterior cervical centrum (shared with Flagellicaudata); middle/posterior cervical vertebrae with pre-epipophysis (shared with Flagellicaudata); cordiform cross-section of anterior caudal centra (shared with Flagellicaudata); middle/posterior dorsal transverse process laterally directed (shared with Diplodocidae); middle caudal centra articular surface with flat ventral margin (shared with Diplodocoidea); ventral surface of mid-caudal centrum with deep median fossa.
Autapomorphies: triangular ventrolateral projections lying below posterior cotyle of middle/posterior cervical centrum visible in posterior view; ventral mid-line keel on middle/posterior cervical centrum bifurcating posteriorly but not meeting the lateroventral flanges; lateral pleuroceol on anterior caudal centra.
Description
The cervical vertebrae are represented by five partial specimens found in association (Supplementary Table 1), including the anterior condyle, posterior half of a vertebra and three right prezygapophyses (Figs. 2, 3). The anterior condyle is U-shaped with an opisthocoelous condition as in other eusauropods22,23 (Fig. 2A), and is mediolaterally wider than dorsoventrally tall [RWR-241-A, centrum articular surface height (cH)/centrum articular surface width (cW) = 0.7; Supplementary Table 2, Supplementary Fig. 2]. In this respect, Tharosaurus is similar to the middle/posterior cervicals of several flagellicaudatans: Suuwassea24 (cH/cW = 0.7), Lingwulong13 (cH/cW = 0.7), Pilmatueia8 (CV7/8, cH/cW = 0.9), Amargasaurus5 (CV8, cH/cW = 0.8), Apatosaurus louisae25 (CV9, cH/cW = 0.7) and Diplodocus carnegii26 (CV11, cH/cW = 0.9). The anterior condyle is markedly more rugose than the other vertebral elements, as in Kaatedocus27 (CV14).
The centrum shows strong anterior constriction with the lateral surfaces excavated by large fossae, similar to many flagellicaudatans8,13,24,26 (Fig. 2B–E). The fossa is partially preserved with much of the dorsal rim and medial wall broken off. The preserved portion suggests the fossa was elliptical, being elongated and dorsoventrally compressed (Fig. 2B–C). The dorsal margin of the fossa is rod-like and extends laterally beyond the ventral margin. The latter is robust, rounded and thickest near the posterior end of the centrum, accentuating the depth of the fossa. Anteriorly it becomes flush with the lateral surface of the centrum (Fig. 2B–C). Beneath the ventral margin of the fossa, the lateral surface of the centrum is markedly concave and flares posteriorly. The fossa is restricted to the posterior half of the centrum and includes two distinct halves: a short and shallow anterior half and a longer and deeper posterior half. The two halves are demarcated by a low U-shaped ridge on the ventral surface of the fossa, medial to the ventral margin and extending slightly onto the medial wall of the fossa. This contrasts with the dicraeosaurids Amargasaurus, Dicraeosaurus, Bajadasaurus, and Pilmatueia where the fossa is undivided and shallow5,7,8. In the middle cervicals of Lingwulong and Suuwassea, however, the lateral fossa is deep and extends nearly along the entire length of the centrum13,24. Furthermore, the lateral fossa in Tharosaurus is succeeded by a posteroventral fossa (Fig. 2D,E), as seen in the mid-cervicals of Amargasaurus, Apatosaurus, Bajadasaurus, and Pilmatueia26,28.
The ventral surface of the centrum (Fig. 2F–G) accommodates paired longitudinal depressions/fossae. Such depressions are common in diplodocids and most dicraeosaurids, barring Bajadasaurus which only bears a longitudinal keel7,8,13,26. The fossae are asymmetrical, with the right fossa mediolaterally wider, and extend up to the posterior margin of the centrum where they flare out. However, in Dicraeosaurus, Lingwulong, and Pilmatueia the fossae are symmetrical and strongly expressed anteriorly, but do not reach the posterior margin of the centrum8,13,26. The fossae in Tharosaurus are flanked laterally by prominent lateroventral flanges and separated by a sharp midline keel (Fig. 2F,G). The keel shows left lateral convexity and remains prominent throughout its preserved length. It extends nearly up to the posterior margin of the fossae and bifurcates into two short ridges. The latter, however, do not reach up to the lateroventral flanges. An accessory ridge, present only on the right ventral surface, extends posterolaterally from the midline keel. The presence of a ventral keel is shared with a diverse array of flagellicaudatans including most dicraeosaurids (Lingwulong, Dicraeosaurus, Pilmatueia, Bajadasaurus and Brachytrachelopan)7,8,9,13 and some later-diverging diplodocids (Barosaurus lentus, Diplodocus carnegii, Dinheirosaurus)23,26. It is, however, absent in some putative plesiomorphic dicraeosaurids (Suuwassea and Smitanosaurus)24,29, barring Kaatedocus27 which is the earliest-diverging dicraeosaurid. The disposition and morphology of this midline keel differentiate Tharosaurus from Lingwulong, Dicraeosaurus, Brachytrachelopan and Barosaurus where the keel is prominent anteriorly4,8,9,26. In Dinheirosaurus the keel is restricted to the posterior part of the centrum and does not extend anteriorly as in Tharosaurus, whereas in Pilmatueia the keel forks anteriorly and posteriorly with the bifurcation point placed slightly anterior to the mid-length of the centrum.
The posterior cotyle of the centrum (RWR-241-B, cH/cW = 0.6, Fig. 2H,I, Supplementary Table 2) is strongly concave and mediolaterally wider than dorsoventrally tall, a feature seen in the middle cervicals of many sauropods including Lingwulong13 (cH/cW = 0.7), Apatosaurus louisae25 (cH/cW = 0.7) and Gleamopus30 (cH/cW = 0.9), and the posterior cervicals of Pilmatueia8 (cH/cW = 0.6) and Suuwassea24 (cH/cW = 0.7). The posterior cotyle is cordiform in outline with a weakly concave dorsal margin immediately ventral to the neural canal as in Lingwulong and Pilmatueia8,13. Tharosaurus, however, can be distinguished by the presence of posteriorly facing triangular facets along the ventrolateral margin of the posterior articular surface.
The neural arch is partially preserved and includes the right prezygapophysis and the region surrounding the neural canal (Figs. 2B–K, 3, Supplementary Table 1). The anterior exit of the canal, although partial, appears oval, but the posterior exit is complete and elliptical, being transversely wider than dorsoventrally high. In Pilmatueia this exit is triangular, whereas in Suuwassea and Lingwulong it is circular–subcircular8,13,24.
The prezygapophysis is anterodorsally directed with the articular facet strongly convex transversely and longitudinally (Fig. 3A–G). The latter feature is shared with middle and posterior cervicals of the flagellicaudatans Diplodocus carnegii (CV8) and Kaatedocus (CV10)26,27,28,29. The prezygapophyseal articular surface is oblong with the posterior border offset from the surrounding dorsal surface of the prezygapophysis by a transverse sulcus (Fig. 3C,D,G), similar to the posterior cervicals of Kaatedocus27. Immediately ventral to the articular surface, the lateral surface of the prezygapophysis bears an anterodorsally-posteroventrally oriented pre-epipophysis (sensu Tschopp et al.26; Fig. 3A,B). The latter is dorsoventrally compressed and succeeded ventrally by a shallow fossa. Such a ridge is also reported in the middle and posterior cervicals of Kaatedocus27 (CV7–10), Apatosaurus louisae26 (CV11) and Suuwassea24,26. The prezygapophysis is supported ventrally by a dorsoventrally high and robust centroprezygapophyseal lamina (cprl, Fig. 3C–F). The lamina is divided into a larger and medially concave lateral branch connecting with the prezygapophysis (lcprl) and a smaller medial branch joining the intraprezygapophyseal lamina (mcprl). The two halves of the cprl accommodate a triangular and deep centroprezygapophyseal fossa (Fig. 3C,D) which borders the neural canal dorsolaterally and is roofed by a sheet-like intraprezygapophyseal lamina. While a divided cprl is common in Flagellicaudata1, in dicraeosaurids the medial branch of the cprl connects with the intraprezygapophyseal lamina and is listed as a synapomorphy of Dicraeosauridae (sensu Whitlock and Wilson Mantilla29). Unlike in Tharosaurus, however, both branches of the cprl join the prezygapophysis in diplodocids1,26,29,31. Nonetheless, the anterior surface of the lcprl immediately ventral to the prezygapophyseal articular surface bears a depression giving the impression of a split cprl, where both branches connect to the prezygapophysis, akin to diplodocids. Further scrutiny shows this depression to be almost indistinct and unlike the morphology of the bifurcated crpl of diplodocids26,32. Furthermore, in diplodocids the apex of this bifurcation is ventrally directed in contrast to the incipient depression in Tharosaurus which is broad and dorsally convex. Therefore, the expression of the divided cprl in the new Indian taxon is closer to that of dicraeosaurids and supports its taxonomic allocation.
The posterior exit of the neural canal is flanked dorsolaterally by two elliptical centropostzygapophyseal fossae [cpof(e)] which are bordered laterally by robust pillar-like centropostzygapophyseal laminae [cpol(e), Fig. 2H,I]. The cpof(e) are roofed by thin intrapostzygapophyseal laminae. The arrangement of the vertebral laminae and fossae in Tharosaurus is comparable with the condition in Lingwulong, Brachytrachelopan, Dicraeosaurus, Amargasaurus and Pilmatueia4,5,8,9,13. However, in the latter two taxa, the cpof(e) are triangular. Furthermore, as in Amargasaurus5,8, the neural arch in Tharosaurus is devoid of a median tubercle.
The preserved neural arch above the neural canal shows deep bifurcation which descends to the roof of the canal and encloses an anteroposteriorly extensive passage of uniform width (Fig. 2H–K; Supplementary Fig. 3). The broken ends of the neural arch dorsal to the cpof(e) suggest the presence of strongly divergent postzygapophyses. These features are reminiscent of the deeply bifurcated neural arch-spine complex of middle to cervicodorsal vertebrae of Amargasaurus and Pilmatueia5,8,28. Moreover, the surface of the neural arch within this passage is finished (as seen in dorsal view, Fig. 2J,K), and bears rugose scars along the mid-line which possibly represent ligament scars. The scars largely occupy the posterior half of the passage and bordered laterally by sub-parallel striations. Rugose tuberosity on the floor of bifurcated neural arch-spine complexes of Apatosaurus ajax and Dicraeosaurus have been interpreted as ligament scars33. Although Tharosaurus lacks the tuberosity, a feature shared with Amargasaurus8, these scars attest to the presence of bifid neural spines in Tharosaurus. Similar bifid neural spines are listed as a synapomorphy of Flagellicaudata in previous studies (e.g., Wilson1).
The specimens (RWR-241-A–E) likely represent the 6th/8th cervical based on comparisons with the middle cervicals of Lingwulong and Bajadasaurus, and the middle–posterior cervicals of Dicraeosaurus, Suuwassea, Amargasaurus, Pilmatueia and Kaatedocus5,8,13,24,27.
Three isolated specimens including a partial neural arch and two partial neural arch-spine complexes are referable to the dorsal vertebral series (Fig. 4A–L, Supplementary Table 1). The neural arch comprises a stout, sub-horizontal transverse process with an anteroposteriorly broad and sigmoidal diapophysis (Fig. 4A–F). Anteriorly, the transverse process bears a broad and moderately deep prezygapophyseal centrodiapophyseal fossa (Fig. 4A,B), whereas its posterior surface bears a thin, mediodorsally-lateroventrally directed lamina, possibly representing the postzygodiapophyseal lamina (Fig. 4C,D). A deep centrodiapophyseal fossa (cdf) positioned ventral to the transverse process is bordered by an anteroventrally directed anterior centrodiapophyseal lamina and a near-vertical posterior centrodiapophyseal lamina (Fig. 4E,F). The specimen is tentatively assigned to the anterior dorsals based on similarity in overall morphology including laminae and fossae configuration with those of the flagellicaudatans Apatosaurus louisae, Dinheirosaurus, Lingwulong and Amargasaurus cazaui5,13,23,25,26.
The neural arch-spine complex is strongly compressed anteroposteriorly and expanded transversely, suggesting a more posterior position in the dorsal vertebral series (sensu McPhee et al.34). The arch comprises a transversely broad and subtriangular spinoprezygapophyseal fossa (Fig. 4G,H). The transverse process is laterally directed, with a vertically oriented distal end, similar to all diplodocids23,35. This laterally oriented transverse process can be considered a local autapomorphy of Tharosaurus unlike a dorsolaterally oriented transverse process in other dicraeosaurids5,9,13,36. This feature may alternatively represent a symplesiomorphy, although Mannion et al.23 listed the dorsolaterally projecting diapophysis in the dorsal vertebrae of the diplodocid Dinheirosaurus as a local autapomorphy. The neural spine is transversely expanded and non-bifid and the disposition of the lateral spinal margins suggests dorsal flaring in anterior/posterior views (Fig. 4G–J). This suggests affinity with the middle/posterior dorsal vertebrae because non-bifurcated neural spines appear from the 6th/7th dorsal of dicraeosaurids, as seen in Lingwulong and Brachytrachelopan9,13. Such flaring of the lateral spinal margins is also known from the middle and posterior dorsals of dicraeosaurids4,13,35. In lateral view, the anterior and posterior borders of the neural spine remain subparallel (Fig. 4K,L), unlike the sub-triangular spinal lateral profile characteristic of many titanosauriforms34,37. The neural spine bears a sharp prespinal lamina placed along the longitudinal midline on the anterior surface (Fig. 4G,H). The prespinal lamina does not extend up to the base of the spine and terminates dorsal to the spinoprezygapophyseal fossa. This lamina, however, extends down to the base of the neural spine in the posterior dorsals of Amargasaurus and Dicraeosaurus5,36. Only a small part of the spinoprezygapophyseal lamina is preserved along the lateral margin of the anterior spinal surface (Fig. 4G,H). The spinodiapophyseal lamina is robust and broad, descending gently from the neural spine to the transverse process (Fig. 4G,H,K,L). The posterior surface of the spine preserves only the right spinopostzygapophyseal lamina (Fig. 4I,J). It is robust and well-rounded, extending medially from the lateral margin of the posterior spinal surface towards the midline. In lateral view, the anterior surface of the spinopostzygapophyseal lamina unites with the posterior surface of the spinodiapophyseal lamina as seen in the posterior dorsals of Brachytrachelopan9 (Fig. 4K,L), and these laminae enclose the postzygapophyseal spinodiapophyseal fossa. Thus, these vertebral specimens likely belong to the middle/posterior dorsals.
An isolated, nearly complete rib (Fig. 4M,N) is identified as the right anterior dorsal rib, based on similarity with that of Galeamopus pabsti30. The specimen is Y-shaped, flares out proximally, and becomes dorsoventrally skewed distally. The rib lacks pneumatic openings as in diplodocoids but contrasts with those of titanosauriforms38. The capitulum is elliptical, anteroposteriorly compressed, and directed anterodorsally. It is longer than the tuberculum and bears prominent striations on the anterior surface (Fig. 4M). The tuberculum is short and curved towards the capitulum. The tuberculum and capitulum enclose a broad U-shaped space. A robust and rounded ridge occupies the anterior surface of the rib distal to the capitulum and tuberculum (Fig. 4M). The ridge extends distally and gradually becomes flush with the surface. The ventral margin is sigmoidal with the portion immediately distal to the tuberculum strongly convex. The dorsal margin is concave, and its posterior surface is largely flat and featureless, except for the strongly rugose tuberculum.
The collection includes two caudal vertebrae (Supplementary Table 1). One of these, a partial centrum of an anterior caudal vertebra, preserves the anterior cotyle and the proximal-most part of the neural arch (Fig. 5A–C). The centrum is robust with a moderately concave anterior cotyle. The latter is dorsoventrally taller than mediolaterally wide (RWR-241-J, cH/cW = 1.2, Supplementary Table 2) and comparable in proportion to the anterior caudals of other flagellicaudatans such as Dicraeosaurus39,40 (cH/cW = c. 1), Suuwassea24 (ANS 21122, cH/cW = 1.1), Lingwulong13 (cH/cW = 1.1) and Amazonsaurus41 (cH/cW = 1.1). It is, however, dorsoventrally taller compared to Diplodocus40 (cH/cW = 0.9), Amargatitanis11 (MACN PV N53, cH/cW = 0.7) and Brontosaurus excelsus26,42 (YPM 1980, cH/cW = 0.9). The ventral margin of the anterior cotyle, although partial, preserves a small chevron facet (Fig. 5A). The preserved lateral surface of the centrum is convex and bears a lateral foramen immediately ventral to the neural arch, a feature present in anterior caudals of most diplodocids23,40,43 (Fig. 5B). The lateral surface, however, does not bear any ridges, similar to Amargatitanis11. In lateral view, the ventral margin of the centrum is strongly concave, akin to Lingwulong13. Moreover, the curvature of the ventral margin suggests the ventral rim of the posterior central articular surface to be at a lower level than the anterior, a feature shared with Dicraeosaurus39 and Suuwassea24. The neural arch is poorly preserved (Fig. 5A,B) and placed anteriorly on the centrum as in Dicraeosaurus39, Lingwulong13 and Suuwassea24. The ventral surface is weakly convex transversely with an incipient midline keel (Fig. 5C). This ventral keel is also present in other dicraeosaurids, including Suuwassea and Dicraeosaurus8,24,39.
The second caudal (RWR-241-K) vertebra is a nearly complete centrum but without the neural arch (Fig. 5D–G). The centrum is anteroposteriorly elongated, suggesting a more posterior position in the caudal series compared to RWR-241-I (sensu Coria et al.8). The centrum is 1.6 times as long as high, with a deep fossa extending along the entire length of the centrum, identifying it to be a middle caudal (sensu McPhee et al.34). This is corroborated by similarities with middle caudal centra of Suuwassea24 (cL/cH = 1.5) and Amargatitanis11 (cL/cH = 1.3). A prominent lateral fossa is also reported in the dicraeosaurid Amargatitanis11. Furthermore, judging from the broken dorsal surface of the centrum, the neural arch was possibly more anteriorly placed in RWR-241-J as in the afore-mentioned dicraeosaurids. The centrum is platyceolous with the anterior and posterior cotyles dorsoventrally taller than mediolaterally wide (Fig. 5D,E), although there is some evidence of deformation. The ventral surface is strongly concave in lateral view with the ventral margin of the posterior cotyle descending below the level of the anterior cotyle. The fossa on the ventral surface is deep, constricted at midlength and bordered by robust ventrolateral ridges (Fig. 5G), similar to the derived diplodocids such as Diplodocus44, Seismosaurus45, and Barosaurus46.
Phylogenetic analysis
The inter-relationship of Tharosaurus within Sauropodomorpha was tested in a reduced version of the data matrix used by Gallina et al.7 (see “Methods” and Supplementary Note 2). The analysis recovered 20 most parsimonious trees with a tree length of 760, consistency index (CI) of 0.483 and retention index (RI) of 0.647. The topology of the strict consensus tree (Supplementary Fig. 4) shows a well resolved clade Sauropoda with diplodocoids and macronarians showing distinct clustering within Neosauropoda, consistent with previous studies7,13. Tharosaurus is recovered as a dicraeosaurid flagellicaudatan, although the clade Dicraeosauridae is poorly resolved. In the 50% majority rule tree (Fig. 6), Dicraeosauridae is better resolved with Tharosaurus being a sister taxon to ((Pilmatueia + Amargatitanis) + (Brachytrachelopan + (Dicraeosaurus + Amargasaurus))).

Phylogenetic position of Tharosaurus indicus gen. et sp. nov. (RWR-241) in 50% majority-rule tree. Clade Dicraeosauridae shaded in pink. Numbers above nodes indicate Bremer support values.
Discussion
Phylogenetic implications
Tharosaurus shares five synapomorphies with Flagellicaudata [divided lateral pleuroceols on cervical centra (ch. 115); bifurcated presacral neural spines (ch. 376); lateroventral flanges on middle/posterior cervical centra (ch. 382); heart-shaped anterior caudal centra cross-section (389); pre-epipophysis on middle/posterior cervicals (ch. 393)] and one unambiguous synapomorphy supports its recovery as a dicraeosaurid [divided cprl in cervicals with the medial lamina connecting with the intraprezygapophyseal lamina (127)]. Tharosaurus also shares two synapomorphies with other diplodocoids [transversely concave ventral surface of cervical centra (ch. 112); quadrangular middle caudal centra (ch. 208)]. Furthermore, three autapomorphies characterize Tharosaurus—smooth and narrow prespinal lamina on non-bifid dorsal neural spine (ch 143), pleuroceol on anterior caudal (ch 194), ventral mid-line keel in middle/posterior cervicals bifurcated posteriorly but not meeting the lateroventral flanges (ch 380). Barring the last autapomorphy, the remaining are local, being also present in a few diplodocids.
To further test the position of Tharosaurus within Sauropodomorpha, two additional analyses were conducted (A1 and A2; Supplementary Notes 3 and 4) using an expanded version of the data matrix of Gallina et al.7 which comprises a spatiotemporally and phylogenetically diverse array of sauropodomorphs, thereby allowing Tharosaurus to be placed anywhere within Sauropodomorpha. Furthermore, in analysis A2, characters involving cervical spines were scored as ‘?’, as these are not preserved in Tharosaurus, although the neural arch morphology strongly suggests the presence of bifurcated spines. Both analyses produce the same results where Tharosaurus is recovered as a dicraeosaurid (Supplementary Figs. 5–8). Furthermore, the phylogenetic bracketing of Tharosaurus by Bajadasaurus and ((Pilmatueia + Amargatitanis) + (Brachytrachelopan + (Dicraeosaurus + Amargasaurus))) in the majority rule tree of analysis A2 (Fig. 8), supports our original inference suggesting bifurcated cervical neural spines in Tharosaurus, as the bracketing taxa also possess the same feature.
Palaeobiogeography
Sauropods are considered to have originated in the Late Triassic/Early Jurassic38,47 but the origin and radiation of Neosauropoda and its major clades (Diplodocoidea and Macronaria) are still amongst the most contentious issues48. Non-neosauropods were restricted to eastern Gondwana (India and Zimbabwe) and parts of Laurasia (Thailand, Germany and China) during Late Triassic–Early Jurassic, suggesting possible physiological constraints to their dispersal to the Americas and Australia47, although sampling biases cannot be ruled out. Neosauropods possibly appeared and radiated during the late Early/ early Middle Jurassic, with Asia and North–South America being some of the areas occupied by their most recent common ancestors (MRCAs)13,49,50. Palaeogeographic reconstructions support this hypothesis since Gondwana and Laurasia remained united for much of the early Mesozoic as the supercontinent Pangaea51. Although the Tethys Ocean was a barrier between Europe + Asia and Gondwana, land connections between North America and South America + Africa during the Triassic and Early Jurassic would have allowed sauropod dispersal51,52. Similarly, dispersal between North America and Europe53 possibly occurred during the Middle Jurassic, although these continents were likely separated by a narrow epicontinental sea (the Viking Corridor) during the Early Jurassic54. However, by the early Bajocian (~ 175 Ma), sea-floor spreading started in the western Tethys, the Central Atlantic region and the Gulf of Mexico55,56,57, and global transgressions flooded most continental shelves during the Middle and Late Jurassic, separating North America from South America and Africa56,57. Consequently, major neosauropod clades in these Laurasian and Gondwanan continents must have originated and radiated, prior to the Bajocian rifting57.
Until recently, the East Asian Jurassic dinosaurs were regarded as endemic fauna, characterized by mamenchisaurids and tetanurans13,58. Dispersal of neosauropods into East Asia apparently occurred only during the Early Cretaceous with the appearance of titanosauriforms, whereas the absence of diplodocoids was thought to be a result of reduced diversity and geographical range due to an end-Jurassic extinction event13. Faunal differences between East Asia and the rest of Laurasia were explained by the presence of an epicontinental seaway isolating Central and East Asia from the rest of Laurasia during the Middle and Late Jurassic49,59. Nonetheless, discoveries of the Chinese Middle and Late Jurassic neosauropods (Dashanpusaurus, Lingwulong, Bellusaurus) tend to weaken the isolation hypothesis13,50 and support a circum-Pangaean distribution of the major neosauropod clades by Middle Jurassic13,50. The age of the Chinese macronarian Dashanpusaurus is controversial but possibly Bajocian50, whereas the age and stratigraphic horizon in which the dicraeosaurid Lingwulong was found has recently been revised from the late Toarcian–Bajocian Yanan Formation to the Bathonian-Callovian Zhiluo Formation50,60. In their palaeobiogeographic analysis of sauropods, Ren et al.50 (p. 6, Fig. 3) assigned a Callovian age to Lingwulong. Thus, the discovery of Tharosaurus assumes considerable significance owing to its older, Early–Middle Bathonian age.
To assess the palaeobiogeographic significance of Tharosaurus, a time-calibrated phylogenetic tree was constructed (Figs. 7, 8). The resultant topology is consistent with the current consensus on rebbachisaurid origins29,61,62 as it suggests a largely Gondwanan origin since most of the taxa are from South America and Africa. Although the oldest rebbachisaurid in this study, Histriasaurus, is from Croatia, it is regarded as a Gondwanan taxon because Croatia was part of the Adriatic-Dinaric Carbonate Platform during the Early Cretaceous, sharing biotic affinities with Africa61,63. Based on the Late Jurassic Maraapunisaurus64, a North American origin of rebbachisaurids and their dispersal into South America through Europe and Africa during the latest Jurassic–earliest Cretaceous has been suggested. However, the rebbachisaurid affinity of this sauropod is questionable29. Early Cretaceous rebbachisaurids from England including Xenoposeidon, claimed to be the oldest member of this clade29,62, also add uncertainty to the Gondwanan origin of Rebbachisauridae. The lower-level phylogenetic affinity of these sauropods remains unresolved and further work is necessary to better understand rebbachisaurid origins. Our study recovers a long rebbachisaurid ghost lineage extending into the Middle Jurassic, corroborating recent work on the timing of origin and dispersal of this clade13,50,65. This work also suggests that differentiation within Rebbachisauridae started during the Late Jurassic, consistent with previous studies53,64.

Time-calibrated phylogenetic tree, based on the 50% majority-rule tree of Supplementary Fig. 6. Macronarians have been combined into a single lineage to enhance clarity. Red star indicates position of Tharosaurus indicus.

Palaeogeographic distribution of diplodocoids with taxa of different ages plotted together in a simplified Middle Jurassic (170 Ma) map to show their spatio-temporal distribution across Pangea. Silhouettes indicate the type of diplodocoid and fossil occurrences. Numbers adjoining sauropod silhouettes indicate age of the fossils as follows: 1—Middle Jurassic (early–middle Bathonian); 2—Late Jurassic; 3—Cretaceous; 4—Middle Jurassic (Callovian). Palaeogeographic map after Scotese67 and sourced from https://www.earthbyte.org/paleomap-paleoatlas-for-gplates/ [This work is licensed under the Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/]. Source of information on sauropod distribution from the Paleobiology database (https://www.paleobiodb.org/) and Ren et al.50.
The diplodocids included in our time-calibrated tree are from the Upper Jurassic horizons of North America (Figs. 7, 8) and suggest a North American origin for this clade. Current consensus on diplodocid taxonomy supports this inference since most of the other valid taxa (Brontosaurus, Galeamopus, Supersaurus and Amphicoelias) are also from North America29,30,37. Furthermore, the tree topology supports a pre-Bajocian divergence and dispersal of diplodocids from dicraeosaurids, consistent with most previous studies on flagellicaudatan dispersal and palaeobiogeography29,57,65. Diplodocids were, however, not restricted to North America, but are also known from the Late Jurassic of Europe (Dinheirosaurus66) and Africa (Tornieria56), and the Early Cretaceous of South America (Leinkupal57).
The clade Dicraeosauridae is represented by nine taxa ranging from Middle Jurassic to Early Cretaceous (Figs. 7, 8). The tree topology shows a late-diverging clade with a preponderance of South American dicraeosaurids (Pilmatueia, Amargatitanis, Brachytrachelopan and Amargasaurus) along with the African Dicraeosaurus, and argues for a Gondwanan origin for this clade. However, recovery of the Late Jurassic Suuwassea as the earliest-diverging dicraeosaurid raises the possibility of a North American origin for Dicraeosauridae and its subsequent migrations to South America and Africa, as initially proposed by Whitlock and Wilson Mantilla29. Recent work on flagellicaudatan palaeobiogeography based on the dicraeosaurid Lingwulong identified Asia + South America as some of the areas for MRCAs of Dicraeosauridae13.
Although later-diverging than Suuwassea and Lingwulong, the discovery of Tharosaurus in a much older Middle Jurassic horizon calls into question the above two hypotheses. The early–middle Bathonian age of Tharosaurus (see Supplementary Note 1) makes it the oldest diplodocoid globally. Even if Lingwulong is reinstated to its original estimated age of 174 Ma (sensu Xu et al.13), Tharosaurus remains the oldest Gondwanan diplodocoid as the African and South American diplodocoids appear from the Kimmeridgian (157 Ma)13,64 (Fig. 7). The sister taxon relationship between the new Indian taxon and the later-diverging African and South American dicraeosaurids indicates ease of faunal exchanges between India and western Gondwana. Based on the age of Tharosaurus and its phylogenetic position near the base of the clade Dicraeosauridae (Fig. 7), India (or a geographically proximate region of eastern Gondwana) is hypothesized here as a potential centre for the radiation and perhaps origin of dicraeosaurids/diplodocoids. Current palaeogeographic reconstructions67 lend support to this hypothesis since plausible dispersal routes from India to western Gondwana–Laurasia, through Madagascar, still remained in place during the Middle Jurassic. Furthermore, the estimated ancestral ranges depicted by the time-calibrated tree show the origin of Neosauropoda and its major clades—Macronaria + Diplodocoidea—to straddle the late Early Jurassic–early Middle Jurassic interval, consistent with recent studies on neosauropod origins13,50,65.
Our proposal favouring possible diplodocoid radiation/origin in India still needs to be reconciled with the phylogenetically more basal Asian taxon Lingwulong, the only other Middle Jurassic dicraeosaurid apart from Tharosaurs. An explanation for these two geographically disparate occurrences is perhaps a circum-Pangaean dispersal event. A direct dispersal to or from Asia is precluded by the Tethys Ocean which acted as a major barrier to terrestrial fauna during the Mesozoic. Furthermore, there is little support for the introduction of diplodocoids into India from Asia via North America and western Gondwana, since Tharosaurus is geologically older and phylogenetically early-branching relative to nearly all African and South American diplodocoids (Figs. 7, 8). The only exception is the South American Bajadasaurus, which is phylogenetically bracketed by Suuwassea + Lingwulong and Tharosaurus, but known from a much younger stratigraphic horizon (Fig. 7). The phylogenetic position of Bajadasaurus may possibly be explained by a radiation event from North America or from India predating Tharosaurus. However, additional sampling leading to greater anatomical coverage of Tharosaurus in the future may change its phylogenetic position. In any case, the long ghost lineage leading to Bajadasaurus presents the possibility of finding earlier-diverging taxa in western Gondwana.
Thus, a more plausible hypothesis, based on the older stratigraphic age of Tharosaurus, is that migrations from India to Asia could have taken place through western Gondwana and North America via Europe (Fig. 8). However, Lingwulong still poses a biogeographic problem, being older than all western Gondwanan and Laurasian diplodocoids. The biogeographic puzzle arising from the two Middle Jurassic dicraeosaurids in India and China indicates a pre-Middle Jurassic origin of neosauropods followed by widespread dispersal, as inferred in the present work and previous studies13,50,65. The lack of corroborative fossils representing temporally and phylogenetically intermediate taxa appears to be a consequence of sampling and geological biases13, and points to the need for more rigorous collecting efforts in these regions.
Despite the fragmentary material currently available and the possibility that the taxonomic attribution within Flagellicaudata may potentially change with the recovery of additional material/character sampling, Tharosaurus remains the oldest known diplodocoid. Together with the putative Bajocian camarasauromorph from Kutch17, the discovery of Tharosaurus makes India a major centre for not only diplodocoid but neosauropod radiation. While previous authors13,50 have considered Asia and the Americas as regions inhabited by the MRCAs of these clades, this study highlights the importance of pre-Bathonian Indian sauropodomorph record in tracing the origins of Neosauropoda. This suggestion is strongly supported by the well-known early-diverging non-neosauropods, Barapasaurus and Kotasaurus, from the Early Jurassic (Sinemurian–Pliensbachian) Kota Formation of India18,19. To conclude, the discovery of Tharosaurus emphasizes the need for increased sampling of Bathonian and older Jurassic horizons of India in search of ancestral taxa intermediate between the neosauropods and Barapasaurus-like eusauropods.
Methods
Osteological description
The osteological description of the skeletal specimens was carried out following the nomenclature of Bandyopadhyay et al.2, Coria et al.8 and Xu et al.13. Different parameters of the fossil specimens were measured (Supplementary Fig. 2) using Mitutoyo digital callipers with a precision of 0.01 mm. Explanatory line drawings are used wherever necessary. The terminology for vertebral laminae and fossae follows Wilson25 and Wilson et al.32.
Phylogenetic analysis
The phylogenetic affinity of Tharosaurus was determined through an analysis based on a combination of characters from the datasets of Xu et al.13 and Gallina et al.7 with 19 additional characters from Tschopp et al.26 (Supplementary Data 1). The taxa-character matrix included 37 taxa and 394 characters. The phylogenetic analysis was performed in TNT version 1.668 where the software memory was set to retain 10,000 trees with a display buffer of 10 Mb (sensu Coria et al.8). Following Gallina et al.7 the Traditional Search option was used to analyse the dataset. The constraints for the analysis included 5000 replications of Wagner trees, where the swapping algorithm was bisection reconnection with 10 trees saved per replication. To determine the robustness of the nodes, Bremer support values were calculated using the script bremer.run where only trees suboptimal by 20 steps were retained. The outgroup taxon in this analysis was Shunosaurus lii.
Data availability
All data associated with the manuscript are provided in the supplementary file.
Code availability
Nomenclatural acts. This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The LSIDs for this publication are urn:lsid:zoobank.org:act:EB6CB405-6FD6-4EDA-9E49-ED08105AFBD8 (Tharosaurus) and urn:lsid:zoobank.org:act:C43C0866-88BB-475B-9439-86C8837AB833 (T. indicus).
Change history
21 August 2023
A Correction to this paper has been published: https://doi.org/10.1038/s41598-023-40641-4
Abbreviations
- ANS:
-
Academy of Natural Sciences, Philadelphia, USA
- MACN:
-
Museo Argentino de Ciencias Naturales, Buenos Aires, Argentina
- YPM:
-
Yale Peabody Museum, New Haven, USA
References
Wilson, J. A. Sauropod dinosaur phylogeny: Critique and cladistic analysis. Zool. J. Linn. Soc. 136, 217–276 (2002).
Bandyopadhayay, S., Gillette, D. D., Ray, S. & Sengupta, D. P. Osteology of Barapasaurus tagorei (Dinosauria: Sauropoda) from the Early Jurassic of India. Palaeontology 53, 533–569 (2010).
Upchurch, P., Barrett, P. M. & Dodson, P. Sauropoda. In The Dinosauria, 2nd ed. (eds Weishampel, D. B., Dodson, P. & Osmolska, H.) (University of California Press, 2004).
Janensch, W. Die wirbelsaule der gatttung dicraeosaurus. Palaeontogr. Suppl. 7, 37–133 (1929).
Salgado, L. & Bonaparte, J. F. Un nuevo saurópodo Dicraeosauridae Amargasaurus cazaui gen. et sp. nov. de la Formación La Amarga, Neocomiano de la provincia del Neuquén, Argentina. Ameghiniana 28, 333–346 (1991).
Carrano, M. The evolution of sauropod locomotion: morphological diversity of a secondarily quadrupedal radiation. In: The Sauropods: Evolution and Paleobiology (eds Curry Rogers, K. A. & Wilson, J. A.) (University of California Press, 2005).
Gallina, P. A., Apesteguía, S. & Canale, J. I. A new long-spined dinosaur from Patagonia sheds light on sauropod defense system. Sci. Rep. 9, 1392 (2019).
Coria, R. A., Windholz, G. J., Ortega, F. & Currie, P. J. A new dicraeosaurid sauropod from the lower cretaceous (Mulichinco formation, Valanginian, Neuquén Basin) of Argentina. Cretac. Res. 93, 33–48 (2019).
Rauhut, O. W. M., Remes, K., Fechner, R., Cladera, G. & Puerta, P. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature 435, 670–672 (2005).
Apesteguía, S. The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). Gondwana Res. 12, 533–546 (2007).
Gallina, P. A. Reappraisal of the Early Cretaceous sauropod dinosaur Amargatitanis macni (Apesteguía, 2007), from northwestern Patagonia, Argentina. Cretac. Res. 64, 79–87 (2016).
Harris, J. D. & Dodson, P. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontol. Pol. 49, 197–210 (2004).
Xu, X. et al. A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs. Nat. Commun. 9, 2700 (2018).
Jain, S. L., Kutty, T. S., Roy-Chowdhury, T. & Chatterjee, S. The sauropod dinosaur from the Lower Jurassic Kota Formation of India. Proc. R. Soc. B Biol. Sci. 188, 221–228 (1975).
Yadagiri, P. A new sauropod Kotasaurus yamanpalliensis from Lower Jurassic Kota Formation of India. Rec. Geol. Surv. India 11, 102–127 (1988).
Yadagiri, P. The osteology of Kotasaurus yamanpalliensis, a sauropod dinosaur from the Early Jurassic Kota Formation. J. Vertebr. Paleontol. 21, 242–252 (2001).
Moser, M., Mathur, U. B., Fürsich, F. T., Pandey, D. K. & Mathur, N. Oldest camarasauromorph sauropod (Dinosauria) discovered in the Middle Jurassic (Bajocian) of the Khadir Island, Kachchh, western India. Paläontol. Z. 80, 34–51 (2006).
Khosla, A. & Bajpai, S. Dinosaur fossil records from India and their palaeobiogeographic implications: An overview. J. Palaeosci. 70, 193–212 (2021).
Bandyopadhyay, S. & Ray, S. Gondwana vertebrate faunas of India: Their diversity and intercontinental relationships. Episodes J. Int. Geosci. 43, 438–460 (2020).
Kumar, K., Bajpai, S., Pandey, P., Ghosh, T. & Bhattacharya, D. Hybodont sharks from the Jurassic of Jaisalmer, western India. Hist. Biol. 34, 953–963 (2022).
Pandey, D. K., Choudhary, S., Bahadur, T., Swami, N. & Sha, J. A review of the Lower—lowermost Upper Jurassic lithostratigraphy of the Jaisalmer Basin, western Rajasthan, India—An implication on biostratigraphy. Vol. Jurass. 10, 61–82 (2012).
Upchurch, P. The evolutionary history of sauropod dinosaurs. Philos. Trans. R. Soc. Lond. B Biol. Sci. 349, 365–390 (1995).
Mannion, P. D., Upchurch, P., Mateus, O., Barnes, R. N. & Jones, M. E. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. J. Syst. Palaeontol. 10, 521–551 (2012).
Harris, J. D. The axial skeleton of the dinosaur Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontology 49, 1091–1121 (2006).
Wilson, J. A. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. J. Vertebr. Paleontol. 19, 639–653 (1999).
Tschopp, E., Mateus, O. & Benson, R. B. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3, e857 (2015).
Tschopp, E. & Mateus, O. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. J. Syst. Palaeontol. 11, 853–888 (2013).
Windholz, G. J., Carballido, J. L., Coria, R. A., Zurriaguz, V. L. & Rauhut, O. W. How pneumatic were the presacral vertebrae of dicraeosaurid (Sauropoda: Diplodocoidea) dinosaurs?. Biol. J. Linn. Soc. 138, 103–120 (2023).
Whitlock, J. A. & Mantilla, J. A. W. The Late Jurassic sauropod dinosaur ‘Morosaurus’ agilis Marsh, 1889 reexamined and reinterpreted as a dicraeosaurid. J. Vertebr. Paleontol. 40, e1780600 (2020).
Tschopp, E. & Mateus, O. Osteology of Galeamopus pabsti sp. nov. (Sauropoda: Diplodocidae), with implications for neurocentral closure timing, and the cervico-dorsal transition in diplodocids. PeerJ 5, e3179 (2017).
Carballido, J. L., Salgado, L., Pol, D., Canudo, J. I. & Garrido, A. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group. Hist. Biol. 24, 631–654 (2012).
Wilson, J. A., D’emic, M. D., Ikejiri, T., Moacdieh, E. M. & Whitlock, J. A. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS ONE 6, e17114 (2011).
Tsuihiji, T. The ligament system in the neck of Rhea americana and its implication for the bifurcated neural spines of sauropod dinosaurs. J. Vertebr. Paleontol. 24, 165–172 (2004).
McPhee, B. W., Mannion, P. D., de Klerk, W. J. & Choiniere, J. N. High diversity in the sauropod dinosaur fauna of the Lower Cretaceous Kirkwood Formation of South Africa: Implications for the Jurassic-Cretaceous transition. Cretac. Res. 59, 228–248 (2016).
Whitlock, J. A. A. phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zool. J. Linn. Soc. 161, 872–915 (2011).
Schwarz-Wings, D. Reconstruction of the thoracic epaxial musculature of diplodocid and dicraeosaurid sauropods. J. Vertebr. Paleontol. 29, 517–534 (2009).
Mannion, P. D., Tschopp, E. & Whitlock, J. A. Anatomy and systematics of the diplodocoid Amphicoelias altus supports high sauropod dinosaur diversity in the Upper Jurassic Morrison Formation of the USA. R. Soc. Open Sci. 8, 210377 (2021).
Wilson, J. A. & Sereno, P. C. Early evolution and higher level phylogeny of sauropod dinosaurs. Soc. Vertebr. Paleontol. Mem. 5, 1–68 (1998).
Janensch, W. Übersicht über die Wirbeltierfauna der Tendaguruschichten, nebsteiner kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv. für Biontologie 3, 81e110 (1914).
Upchurch, P. & Mannion, P. D. The first diplodocid from Asia and its implications for the evolutionary history of sauropod dinosaurs. Palaeontology 52, 1195–1207 (2009).
De Souza Carvalho, I., dos Santos Avilla, L. & Salgado, L. Amazonsaurus maranhensis gen. et sp. nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) of Brazil. Cretac. Res. 24, 697–713 (2003).
Ostrom, J. O. & McIntosh, J. S. Marsh’s Dinosaurs: The Collection from Como Bluff Vol. 1 (Yale University Press, 1966).
Lovelace, D. M., Hartman, S. A. & Wahl, W. R. Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocoid phylogeny. Arquivos do Museu Nacional, Rio de Janeiro 65, 527–544 (2008).
Hatcher, J. B. Diplodocus (Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Mem. Carn. Mus. 1, 1–63 (1901).
Gillette, D. D. Seismosaurus halli, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA. J. Vertebr. Paleontol. 11, 417–433 (1991).
McIntosh, J. S., Tidwell, V. & Carpenter, K. The genus Barosaurus Marsh (Sauropoda, Diplodocidae). In Thunder-Lizards: The Sauropodomorph Dinosaurs. (Indiana University Press, 2005).
Gillette, D. D. The geographic and phylogenetic position of sauropod dinosaurs from the Kota formation (Early Jurassic) of India. J. Asian Earth Sci. 21, 683–689 (2003).
Averianov, A. O. & Zverkov, N. G. New diplodocoid sauropod dinosaur material from the Middle Jurassic of European Russia. Acta Palaeontol. Pol. 65, 499–509 (2020).
Wilson, J. A. & Upchurch, P. Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Early Cretaceous of China. J. Syst. Palaeontol. 7, 199–239 (2009).
Ren, X. X. et al. Re-examination of Dashanpusaurus dongi (Sauropoda: Macronaria) supports an early Middle Jurassic global distribution of neosauropod dinosaurs. Palaeogeogr. Palaeoclimatol. Palaeoecol. 610, 111318 (2023).
Poropat, S. F. et al. New Australian sauropods shed light on Cretaceous dinosaur palaeobiogeography. Sci. Rep. 6, 34467 (2016).
Blakey, R. C. In Resolving the Late Paleozoic Ice Age in Time and Space vol. 441 Geological Society of America Special Paper (eds Christopher, R. F., Tracy, D. F. & Isbell, J. L.) (Geological Society of America, 2008).
Smith, A. G., Smith, D. G. & Funnell, B. M. Atlas of Mesozoic and Cenozoic Coastlines (Cambridge University Press, 2004).
Bardet, N. et al. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Res. 26, 869–887 (2014).
Golonka, J. et al. In The Continental Jurassic. Museum of Northern Arizona Bulletin 60 (ed. Michael, M.) (Museum of Northern Arizona, 1996).
Remes, K. Revision of the Tendaguru sauropod dinosaur Tornieria africana (Fraas) and its relevance for sauropod paleobiogeography. J. Vertebr. Paleontol. 26, 651–669 (2006).
Gallina, P. A., Apesteguía, S., Haluza, A. & Canale, J. I. A diplodocid sauropod survivor from the Early Cretaceous of South America. PLoS ONE 9, e97128 (2014).
Xing, L. et al. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. J. Vertebr. Paleontol. 35, e889701 (2015).
Russell, D. A. The role of Central Asia in dinosaurian biogeography. Can. J. Earth Sci. 30, 2002–2012 (1993).
You, S. S., Li, Z. J. & Li, Y. B. The stratigraphical characteristics and sedimentary environment of dinosaur fossils in Lingwu, Ningxia. Geol. J. Sichuan 39, 31–35 (2019).
Ibiricu, L. M. et al. New material of Katepensaurus goicoecheai (Sauropoda: Diplodocoidea) and its significance for the morphology and evolution of Rebbachisauridae. Ameghiniana 52, 430–446 (2015).
Salgado, L., Gallina, P. A., Lerzo, L. N. & Canudo, J. I. In South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution (eds Otero, A., Carballido, J. L. & Pol, D.) (Springer Earth System Sciences, 2022).
Dalla Vecchia, F. In Thunder-Lizards: The Sauropodomorph Dinosaurs (eds Tidwell, V. & Carpenter, K.) (Indiana University Press, 2005).
Carpenter, K. Maraapunisaurus fragillimus, n. g. (formerly Amphicoelias fragillimus), a basal rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado. Geol. Intermount. West 5, 227–244 (2018).
Mannion, P. D., Upchurch, P., Schwarz, D. & Wings, O. Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: Phylogenetic and biogeographic implications for eusauropod dinosaur evolution. Zool. J. Linn. Soc. 185, 784–909 (2019).
Bonaparte, J. F. & Mateus, O. A new diplodocid, Dinheirosaurus lourinhanensis gen. et sp. nov., from the Late Jurassic beds of Portugal. Revista del Museo Argentino de Ciencias Naturales 5, 13–29 (1999).
Scotese, C.R. PALEOMAP PaleoAtlas for GPlates and the PaleoData Plotter Program, PALEOMAP Project, http://www.earthbyte.org/paleomap-paleoatlas-for‐gplates/. https://doi.org/10.13140/RG2.2.34367.00166 (2016).
Goloboff, P. A. & Morales, M. E. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39, 144–153 (2023).
Acknowledgements
We acknowledge with thanks the helpful comments, suggestions and a constructive critique of the manuscript by the two reviewers and Editor Scientific Reports. We thank Matthew Carrano, National Museum of Natural History, Smithsonian Institution (Washington DC, USA) for going through the initial draft of the manuscript and offering his valuable comments and suggestions. We also thank Pablo A. Gallina for access to the phylogenetic dataset used in this analysis. Authors from the Geological Survey of India (GSI) thank Director General, and the following officers from the GSI, Western Region, Jaipur for providing departmental facilities, constant support and permission to publish this study: Additional Director General & HOD, Deputy Director General & RMH-IV and Director, Palaeontology Division. Some of the material described in this paper is part of the ongoing doctoral work of one of the co-authors (TG) at IIT Roorkee. SB would like to acknowledge support obtained from IIT Roorkee as part of his Institute Chair Professorship. The Science and Engineering Research Board (SERB), Government of India (Grant no. PDF/2021/00468) and Indian Institute of Technology Roorkee are acknowledged for providing financial and infrastructural facilities to DD.
Author information
Authors and Affiliations
Contributions
K.K., P.P., S.B., and D.B. conceived the problem. K.K., P.P. and T.G. collected the data. D.D. and S.B. analysed and interpreted the data and wrote the manuscript. S.B. and D.D. were involved in further revisions and modifications of the manuscript with contributions from K.K., P.P. and T.G.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this Article was revised: The original version of this Article contained an error in the Results section, under the subheading ‘Systematic palaeontology’, where the expanded form of the specimen number “RWR-241(A–K)” was missing.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bajpai, S., Datta, D., Pandey, P. et al. Fossils of the oldest diplodocoid dinosaur suggest India was a major centre for neosauropod radiation. Sci Rep 13, 12680 (2023). https://doi.org/10.1038/s41598-023-39759-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-39759-2
This article is cited by
-
Largest known madtsoiid snake from warm Eocene period of India suggests intercontinental Gondwana dispersal
Scientific Reports (2024)
-
Marine Jurassic basins along the western and northern margins of the Indian craton: a progress report
Proceedings of the Indian National Science Academy (2024)
-
Fossil find is earliest record of a plant-eating dinosaur group
Nature India (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
