
Abstract
The effects of elevated CO2 on the early life stages of coral were investigated by culturing the pelagic larvae and new recruits of Acropora gemmifera at three concentrations of CO2 (corresponding to pH = 8.1, 7.8 and 7.5, respectively). Acidified seawater resulted in fewer A. gemmifera larvae settling, and led to the production of smaller new recruits by slowing the development of the skeleton. The delayed development of new recruits due to elevated CO2 was consistent with the downregulation of calcification related genes. Several genes related to HCO3− and Ca2+ transporters were downregulated by elevated CO2, with solute carriers (SLC) (membrane transport proteins) possibly playing an important role. The downregulation of these membrane transport proteins might suppress the transport of calcium, bicarbonate and organic matter, resulting in the delayed development of A. gemmifera.
Similar content being viewed by others


Introduction
Coral calcification is believed to be largely controlled by the degree of aragonite saturation (ΩA), and is significantly decreased by elevated atmospheric partial pressure CO2 (pCO2)1,2,3 The early life stages of many marine organisms seem to be particularly vulnerable to acidified seawater. Consequently, there is an increasing concern about the potential impacts of increasing pCO2 on corals during their early life stages4.
There are both pelagic and benthic phases in the coral life-cycle5. The planktonic larvae of stony corals are non-calcifying until they settle, with a previous study documenting their morphological changes6 (Fig. 1). After coral larvae settle, new recruits develop a calcifying base, and form a synapticular ring and septa to establish the structure of the coral skeleton. The skeletal morphology changes rapidly during the early life stage, suggesting that the expression of calcification related genes also changes7. Coral growth and calcification are closely related to various protein transporters of Ca2+, H+ and HCO3− 8,9,10,11, as well as coral skeleton organic matrix proteins (SOMPs)7,12,13. There has been detailed information on the settlement and metamorphosis of early life stages of coral14,15. In addition, previous studies reported that high CO2 affected the early life stage of coral, by delaying calcification and settlement16,17,18,19,20, modulating metabolism21,22, changing skeleton morphology19,23, and reducing survival and algal infection rates24. Also, some studies showed that elevated CO2 exhibited no significant effects on coral survival and corallite area14,25.

Embryonic development and larval development of coral A. gemmifera (From zoom stereo microscope). (A) 2 cleavage (2 hr), (B) larvae (prawn chip, 6 hr), (C) rodlike planula (47 hr), (D) settled new recruits (5 d).
Gene expression has been analysed to understand the post-settlement success and skeleton formation of coral during the entire period of early development26,27. Moya, et al.28 used the Illumina RNAseq approach to study how acute exposure to elevated CO2 affected the gene expression of the early life of Acropora millepora. The authors reported that the expression of most ion transport proteins was not affected by elevated CO2, while many membrane-associated or carbonic anhydrases were downregulated. Although corals lacked a strong response to elevated pCO2 in the study of Rocker, et al.29, new recruits might have the capacity to acclimate rapidly to elevated pCO2 by upregulating specific heat shock proteins (HSPs) and a suite of anti-apoptotic proteins (e.g. Bcl-2 family members)30. Kaniewska, et al.31 also demonstrated the upregulation of membrane transporters under high CO2 concentrations, as well as the regulation of genes involved in the cytoskeletal interactions and cytoskeletal remodelling of membranes. The expression of heat shock proteins, carbonic anhydrase, and rubisco protein has also been studied in coral larvae32,33,34, supporting the assumption that elevated temperature and CO2 are impacting coral development.
Previous studies primarily used A. millepora as a model species in the analysis of gene expression. However, studies of other species might provide a more complete understanding of how coral communities respond to high CO2 concentrations. The rapidly growing branching coral, A. gemmifera, is widely distributed, and is an ecologically important genus in the South China Sea. In the current study, the pelagic larvae and new recruits of A. gemmifera were exposed to three pCO2 concentrations to investigate the transcriptome of the coral during the early life stages, with a focus on the adhesion and structural proteins involved in calcification. In previous studies, sequencing reads were usually mapped onto a transcriptome assembly of an A. millepora. The present study conducts a transcriptome analysis using de novo RNA-seq and gene expression analysis to gain deep insight into the responses of calcification related genes to climate changes.
Results
Settlement, survivorship, and morphological change
The settling proportion was lower in low pH treatments (p < 0.05), whereas survivorship exhibited no significant difference between the treatments (p > 0.05) (Fig. 2). Tank effects were not significant (p > 0.05). The first synapticular ring and primary septa in new coral recruits were detected after approximately 3 days after settlement. Three synapticular rings with secondary septa and theca were observed after 5 days (Fig. 3A–C). For treatments in which pH was 7.8 and 7.5, the ratios of new recruits with three synapticular rings were always lower than those in the control (Fig. 3D). All settled new recruits had more than three synapticular rings after 17 days at pH 8.1, with skeletal development being visibly suppressed at lower pH (Fig. 3D).

(A) The larval settlement ratios; and (B) survivorship of new recruits after 3-day and 10-day settlement at different pH values. *Denotes significant difference (p < 0.05), compared to the control (pH = 8.1).

Progressive morphological changes in A. gemmifera: (A) first day settlement; (B) formation of first synapticular ring and primary septa on the 2–7th days; (C) three synapticular rings with secondary septa and theca after 5 days; (D) the ratios of new recruits showing three synapticular rings; and (E) average diameter measurement of A. gemmifera (n = 3) at pH = 8.1, 7.8 and 7.5 respectively).
From 5 days after settlement, the diameter of the new recruits remained significantly smaller in the treatment with lower pH than the control (p < 0.05) (Fig. 3E). In addition, crystals were granule-like at pH 8.1, whereas crystals became increasingly distorted and needle-like in treatments with lower pH (Fig. 4A–F).

The skeletal structure on the 40th day after settlement (A) and (B) at pH = 8.1; (C) and (D) at pH = 7.8; and (E) and (F) pH = 7.5 by scanning electron microscopy imaging. In (A), (C) and (E) scale bar represents 10 µm, in (B), (D) and (F) scale represents 1 µm.
Differentially expressed genes
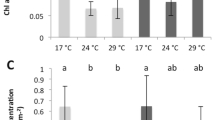
A total of 12,367,906 reads were obtained from the A. gemmifera new coral recruits, with ~60% of reads being mapped to the total de novo unigene assembly (Table S1). As new recruits developed, only ~6% of transcripts were differentially expressed between C1 (3 days after settlement) and C2 (40 days after settlement) (Fig. 5). In addition, ~3% transcripts were differentially expressed in both acute (3 days) and prolonged treatments (40 days) with elevated CO2 (Fig. 5).

The percentage of differentially expressed genes. (A) Control after 40 days vs 3 days (C2 vs C1), (B) high CO2 treatment after 3 days vs control after 3 days (H1 vs C1), (C) high CO2 treatment after 40 days vs control after 40 days (H2 vs C2).
Significant mortality of new recruits was observed in the treatments with elevated CO2 (Fig. 2B). Some apoptosis related genes were downregulated (Table 1) in response to elevated CO2, including apoptosis regulator BCL-2, caspase-3 and caspase-8. Some of these apoptosis-related genes are responsible for the negative regulation of apoptotic processes, while others might positively regulate apoptotic processes (Table 1).
A series of genes related to the HCO3− and Ca2+ transporters were downregulated in response to elevated CO2 (Table 2). Solute carriers (SLC) are membrane transport proteins located in the cell membrane that transport charged and uncharged inorganic and organic molecules. Solute carrier (SLC) genes were mostly upregulated after 40 days compared to 3 days (C2 vs C1), and were mostly downregulated in response to elevated CO2 after 40 days of settlement compared to 3 days of settlement (H1 vs C1 or H2 vs C2). SLC4 member 10 is responsible for transporting HCO3− (Table 2), while SLC8 and SCL24 are Ca2+ transporters. The plasma membrane Ca2+ transporting ATPase was also downregulated in response to elevated CO2 (Table 2). Many other members of various SLC families were also downregulated by the high CO2 treatment. For example, SLC12 and SLC25 were downregulated after 40 days under high CO2 (Table 2). Similarly, some skeleton organic matrix genes were downregulated by the high CO2 treatments, including cadherin, thrombospondin, hemicentin, actin, collagen, and galaxin after 3 days of settlement (Table 3).
Discussion
Survivorship and regulators of apoptosis
The survivorship of new recruits decreased after 40 days of exposure to high CO2 concentrations, but not significantly (Fig. 2B). As previous studies showed that the growth rates and colony size of hermatypic corals are inversely proportional to mortality risk35,36, the slow development of skeleton synapticular rings under elevated CO2 may potentially increase the risk of morality. In addition, suppressed growth under elevated CO2 is unfavourable for new recruits that compete for space with algae and other benthic organisms, because it reduces the fitness of new recruits, affecting population dynamics37.
Previous studies showed that the expression of apoptotic genes is generally related to cellular mortality and survivorship, with the expression being either pro- or anti-apoptosis30. Caspase 8 is essential for programmed cell death, and was downregulated in the high CO2 treatments of the current study. In addition, anti-apoptotic proteins (e.g. Bcl-2 family members) were also downregulated (Table 1), whereas heat shock proteins (HSPs) and heat shock factors (HSFs) were not differentially expressed (data not shown). A previous study showed that new recruits of A. millepora could acclimate to elevated pCO2 by upregulating the expression of HSPs, as well as Bcl-2 family members30; however, the current study indicated that this mechanism does not exist in A. gemmifera. Thus, different coral species might trigger different molecular mechanisms in responses to elevated pCO2.
Delayed development and calcification related genes
In this study, the settlement, diameter, skeleton structure, and crystal shape of new recruits were affected by elevated CO2 concentrations. These results were consistent with previous studies where new recruits reared in OA had smaller diameters than did the control38,39,40. The current study showed that, in addition to diameter, the ratio of new recruits with three synapticular rings was lower before 28 days under higher CO2 concentrations. To our knowledge, only a few studies demonstrate the effects of elevated CO2 on the development of skeleton synapticular rings23. The slow development of the coral skeleton prompted us to conduct a more extensive survey of the major genes related to calcification, and to investigate their expression levels. Two major components contribute to the calcification process: (1) transporters of Ca2+, H+ and inorganic carbon, and (2) organic matrix proteins that catalyse the crystals to form macroscopic structures41.
Various transporters of Ca2+, H+ and inorganic carbon have been examined by previous studies9,10,11. Among these transporters, Ca2+ transporting ATPase was presumably a transporter of Ca2+ and H+ 9. In our study, Ca2+ transporting ATPase was only differentially expressed during the first 3 days under high CO2 concentrations, but was downregulated after 40 days of exposure. Carbonic anhydrases contribute to the interconversion of carbon dioxide and bicarbonate11, which were also not differentially expressed under high CO2 in this study.
In addition, solute carriers (SLC) are membrane transport proteins located in the cell membrane that transport charged and uncharged inorganic and organic molecules. Zoccola, et al.42 reported that the SLC4 and SLC26 families are both bicarbonate anion transporters in the coral Stylophora pistillata. SLC4 (anion exchanger) member 2 was neither differentially expressed in response to elevated CO2 in A. millepora28 nor A. gemmifera (this study); however, SLC4 member 10 (bicarbonate anion transporter) was downregulated in under high CO2 concentrations in the current study (Table 2). This result suggests that the bicarbonate transports were suppressed by elevated CO2, with SLC4 member 10 either supplying bicarbonate anion at the site of calcification or aiding in pH regulation42. A previous study showed that SLC26 was differentially expressed by different temperatures43; however, SLC26 member 11 was not differentially expressed by high CO2 in our study (Table 2). Hence, elevated CO2 might reduce bicarbonate anion transport by suppressing the expression of SLC4 member 10 rather than SLC26. In addition to bicarbonate anion transport, the calcium transporters (SLC8 and SLC24) were also downregulated by elevated CO2, including SLC 24 members 2 and 4 (Table 2). The members of the SLC24 gene family encode K+-dependent Na+/Ca2+ exchangers (NCKX) that utilise both inward Na+ and outward K+ gradients to extrude Ca2+ from cells44. However, it remains unclear whether SLC24 is located on the membrane of tissue cells or the calicoblastic cells. Consequently, it is also unclear whether SLC24 is able to regulate coral calcification. Both HCO3− and Ca2+ transporters were downregulated under elevated CO2 concentrations, which probably led to slower calcification rates and the delayed development of new recruits. In addition to HCO3− and Ca2+, the transporters of organic matter (such as oligopeptides, monocarboxylic acid, fatty acids, and amino acids) were also downregulated (Table S2). The presence of fewer organic matter transporters might decrease the energy supply available for the calcification and growth of coral.
It has been long hypothesized that the precipitation of aragonite is catalysed by and organized on an extracellular organic matrix. A recent study identified 36 coral of coral skeleton organic matrix proteins (SOMPs) in S. pistillata12. In our study, only five out of these 36 skeletal proteins were differentially expressed, including cadherin, thrombospondin, hemicentin, actin, and collagen (Table 3). However, these proteins were usually downregulated in H1 vs. C1, but not in H2 vs C2. The specific functions of some organic matrix proteins have yet to be determined in coral species, but their functions have been studied in human and other animals. Cadherin plays an important role in cell adhesion, forming junctions to bind cells together within tissues45. Collagens are also important in the regulation of cell-cell adhesion, differentiation, and wound healing46. Coral acid-rich protein (CARPs) members have been reported as putative calcification-related proteins in A. millepora13 and S. pistillata12,47, but were not differentially expressed in response to elevated CO2 in our study (data not shown).
Conclusions
The present study explored the effects of elevated pCO2 on the post-settlement development of new coral recruits of A. gemmifera. Elevated CO2 slowed the formation of the skeleton structure and crystal microstructure of new recruits, slowing settlement and reducing the diameter of new recruits. Many solute carriers (SLC) (membrane transport proteins) were downregulated in response to elevated CO2, as well as the SLC4 family members responsible for supplying HCO3− to the site of calcification. The downregulation of membrane transporters proteins might represent the mechanism underlying the delayed development of new coral recruits.
Materials and Methods
Sampling and incubation
In April 2015, 5 colonies of A. gemmifera (~40 cm diameter) were collected from Luhuitou fringing reef, Sanya Bay, China (18° 15′ N; 109° 25′ E) at ~2–5 m depth. Before spawning, colonies were transferred to outdoor flow-through aquaria in Sanya Bay, Hainan Island, outside the Tropical Marine Biological Research Laboratory. Ten colonies of A. gemmifera (~40 cm diameter) were collected. These colonies were >5 m away from each other, assuming their genotypes were not the same. Gametes were collected from these 10 colonies without separate crossings.
In our CO2 system, seawater chemistry was manipulated by continuous and direct bubbling of pure CO2 (except for the control treatment). The bubbling rates were controlled by high precision pressure gauges and valves (DC01-01, Dici, China) and pH controller (pH2010, WEIPRO, China) to create the desired pCO2 (Table 4). Seawater was pumped from the reef water (~200 m away from our culture tank) at a depth of 5 m and run through a sand- filter. As the seawater was not further filtered, no extra food and zooxanthellae were added into culture aquaria to create. Each aquarium was covered with a screen to provide a light field that was approximately 50% of the surface solar irradiance, a condition similar to that at a depth of 3–5 m. Treated seawater was then pumped to the 90 L experimental tanks (three replicate tanks per treatment) via PVC-pipes. The water-exchange rate was 600 ± 10 mL min−1 (Fig. S1). The tanks were partially immersed in a 2000 L flow-through aquarium.
Larvae were cultivated in flow-through tanks (90 L) at a density of ~1 larvae/ml under ambient conditions (pH NBS 8.16 ± 0.01, 27.5 °C, total alkalinity (TA) 2250.8 ± 5 μM, salinity 33 psu), ~750 ppm CO2 (pH = 7.8) or 1200 ppm CO2 (pH = 7.5) (Table 4), reflecting the control condition, intermediate and high CO2 concentrations based on IPCC predictions for the 21st century (2007). There were four replicates in each treatment, and the turnover time of seawater of each tank was 12 hours. The outflow of each tank was covered with a 180 μm mesh to avoid larvae loss. About 10–15 preconditioned terrocota tiles with a mixed community of crustose coralline algae were placed into each tank to induce larval settlement. After four days, larvae were randomly sampled for settlement assays. Six days later, all swimming larvae were flushed away, and new recruits on tiles were reared in continuous flow-through tanks for observing survivorship, diameter, and skeleton structure (90 L) (Fig. S1). The survivorship of new recruits on tiles was assessed on the 3rd and 10th days after settlement, by observing it using a stereomicroscope (Olympus, SZ51). Death was defined as the time point when live tissue was no longer present. Survivorship was expressed as the proportion surviving within each tank. The number of new recruits was counted, and image of new recruits was taken using a stereomicroscope. The diameter of live tissue was measured with the program ToupView 3.7. The settled recruits were collected at the end of 40-day incubation, and their skeletons was examined by scanning electron microscopy (SEM, ZEISS Supra55).
Settling proportion
The settling proportion was estimated in a separate experiment. In this separate experiment, about 300 larvae were sampled from each treatment tank and transferred to lidded chambers (10 L) filled with seawater with the corresponding experimental treatments. A preconditioned terrocota tile with a mixed community of crustose coralline algae was placed at the bottom of the chambers. Half of the seawater in each chamber was replaced every 12 hours. Larval settlement was assessed after two days using a stereomicroscope (Olympus, SZ51). Settling proportion was estimated by the number of new recruits/total initial larvae.
TA and pH
Water samples were collected every three days from each tank, and total alkalinity (TA) was measured by an open-cell automatic titrator (Metrohm 877 titrino plus) using 0.1 M HCl for each 70 g seawater sample. Accuracy of the measurements was checked against certified seawater reference material (Batch 118) obtained from Dr A. Dickson. The pH in culture tanks was continuously monitored using a Ross semi-micro glass combination pH electrode (Orion). The other carbonate system parameters (i.e. dissolved inorganic carbon (DIC), Ω and pCO2) were calculated using a spreadsheet version of the CO2SYS program48, based on measurements of temperature, salinity, pH and alkalinity. The calculation used the CO2 equilibrium constants K1 and K249.
De novo transcriptome sequencing
On the 3rd and 40th day after settlement, settled new recruits (~10–50 new recruits per biological sample) of each tank were randomly sampled from ~10–15 tiles with a sterile scalpel, and 1 sample per experimental treatment tank were obtained. The samples of control and high CO2 treatments (1200 μatm CO2) were snap frozen the samples in liquid nitrogen and then stored the samples at −80 °C until further processing. TRI-Reagent (Sigma) was used for RNA extraction according to the manufacturer’s manual. Samples were stored at −80 °C, until transcriptome sequencing was carried out for de novo assembly and analysis by the Huada Gene Company (http://www.genomics.cn/index).
After the mRNAs were fragmented into short fragments as templates, double-stranded cDNA was synthesized. The ligated fragments were then generated by a series of reaction processes that included purification of PCR products, end repair, and ligation of Illumina adapters. AMPure XP system was used for the purification PCR products, and the Agilent Bioanalyzer 2100 system was used to assess the library quality. An Illumina HiSeq™ 2000 sequencing platform was employed to sequence the cDNA library (NEB, USA).
Data filtering, de novo assembly and annotation
We filtered the raw data to generate clean reads by removing adapter sequences, reads with unknown bases (N) >10%, and low-quality sequences (the percentage of low quality bases of quality value <5). After obtaining clean reads, the clean reads were de novo assembled by the Trinity program into contigs50. The read coverage in each sample roughly exhibited a similar distribution. All unigenes were annotated using the BLASTx against the NCBI Nr (http://www.ncbi.nlm.nih.gov/genbank/), and KEGG pathway (http://www.genome.jp/kegg/pathway.html) databases, with an E-value cut-off of 1E-5. To further analyze the annotation results, the GO function of unigenes were classified in terms of molecular function, biological process, as well as cellular component (http://www.geneontology.org/) using Blast2GO51.
Differential expression analysis
Clean reads of each treatment were mapped to reference genes transcriptome generated by total de novo RNA-seq52,53. The gene expression level was calculated by the value of FPKM (Fragments Per Kilobase of transcript per Million). The differentially expressed genes were annotated using GO and KEGG enrichment analyses according to a method similar to that described by52,53. The best BLAST hit was used to assess differential gene expression, which was estimated with log2fold change. The triplicate samples were compared between control 3 days after settlement (C1), control 40 days after settlement (C2), acidified treatment 3 days after settlement (H1), and acidified treatment 40 days after settlement (H2).
Statistical analyses
Two-way analyses of variance (ANOVA) tested the effects of pCO2 and different tanks on coral biological parameters (e.g. settling proportion, survivorship, diameter, and gene expression). Different treatments of pCO2 and different tanks were fixed, whereas each coral biological parameter was a random factor. Post hoc Tukey tests were conducted if significant effects were obtained (p < 0.05). Statistical analyses were performed using SPSS 19 System for Windows. The differential gene expression analysis of the two experimental groups (high CO2 treatment vs control) was determined using Expdiff. A q value (or FDR) of <0.005 & |log2fold change|>1 was set as the threshold for significantly different expressions.
Availability of raw data
The raw data used for assembly, annotated expression data and annotated data were deposited in the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) under accession number GSE96935.
References
Kleypas, J. A. et al. Impacts of ocean acidification on coral reefs and other marine calcifiers. A Guide for Future Research. Report of a workshop sponsored by NSF, NOAA & USGS (2016).
Cohen, A. L. & Holcomb, M. Why corals care about ocean acidification: uncovering the mechanism. Oceanography 22, 118–127 (2009).
Langdon, C. & Atkinson, M. Effect of elevated pCO2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. J Geophys Res 110, C09S07 (2005).
Albright, R. Reviewing the effects of ocean acidification on sexual reproduction and early life history stages of reef-building corals. J Mar Biol 2011, 1–14 (2011).
Grasso, L. C. et al. Microarray analysis identifies candidate genes for key roles in coral development. BMC Genomics 9, 1–18 (2008).
Vandermeulen, J. H. Studies on reef corals. III. Fine structural changes of calicoblast cells in Pocillopora damicornis during settling and calcification. Mar Biol 31, 69–77 (1975).
Mass, T. et al. Temporal and spatial expression patterns of biomineralization proteins during early development in the stony coral Pocillopora damicornis. Proc Roy Soc B-Biol Sci 283, https://doi.org/10.1098/rspb.2016.0322 (2016).
Allemand, D. et al. In Coral reefs: an ecosystem in transition (eds Stambler, N., Dubinsky, Z.) 119–150 (Springer Netherlands, 2011).
Zoccola, D. et al. Molecular cloning and localization of a PMCA P-type calcium ATPase from the coral Stylophora pistillata. Biochimica Et Biophysica Acta-Biomembranes 1663, 117–126 (2004).
Tambutté, É. et al. A compartmental approach to the mechanism of calcification in hermatypic corals. J Exp Biol 199, 1029–1041 (1996).
Bertucci, A. et al. Carbonic anhydrases in anthozoan corals-A review. Bioorgan Med Chem 21, 1437–1450 (2013).
Drake, J. L. et al. Proteomic analysis of skeletal organic matrix from the stony coral Stylophora pistillata (vol 110, pg 3788, 2013). Proc Natl Acad Sci USA 110, 7958–7958 (2013).
Ramos-Silva, P. et al. The skeleton of the staghorn coral Acropora millepora: molecular and structural characterization. PLoS One 9, e97454 (2014).
Edmunds, P. J., Cumbo, V. R. & Fan, T. Y. Metabolic costs of larval settlement and metamorphosis in the coral Seriatopora caliendrum under ambient and elevated pCO2. J Exp Mar Biol Ecol 443, 33–38 (2013).
Chua, C. M., Leggat, W., Moya, A. & Baird, A. H. Temperature affects the early life history stages of corals more than near future ocean acidification. Mar Ecol Prog Ser 475, 85–92 (2013).
Webster, N. S., Uthicke, S., Botté, E. S., Flores, F. & Negri, A. P. Ocean acidification reduces induction of coral settlement by crustose coralline algae. Global Change Biol 19, 303 (2013).
Anlauf, H., D’Croz, L. & O’Dea, A. A corrosive concoction: The combined effects of ocean warming and acidification on the early growth of a stony coral are multiplicative. J Exp Mar Biol Ecol 397, 13–20 (2011).
de Putron, S. J., McCorkle, D. C., Cohen, A. L. & Dillon, A. B. The impact of seawater saturation state and bicarbonate ion concentration on calcification by new recruits of two Atlantic corals. Coral Reefs 30, 321–328 (2011).
Cohen, A. L., McCorkle, D. C., de Putron, S., Gaetani, G. A. & Rose, K. A. Morphological and compositional changes in the skeletons of new coral recruits reared in acidified seawater: Insights into the biomineralization response to ocean acidification. Geochem Geophy Geosy 10 (2009).
Foster, T., Gilmour, J. P., Chua, C. M., Falter, J. L. & Mcculloch, M. T. Effect of ocean warming and acidification on the early life stages of subtropical Acropora spicifera. Coral Reefs 34, 1217–1226 (2015).
Cumbo, V. R., Edmunds, P. J., Wall, C. B. & Fan, T. Y. Brooded coral larvae differ in their response to high temperature and elevated pCO2 depending on the day of release. Mar Biol 160, 2903–2917 (2013).
Nakamura, M., Ohki, S., Suzuki, A. & Sakai, K. Coral larvae under Ocean acidification: survival, metabolism, and metamorphosis. Plos One 6, e14521 (2011).
Foster, T., Falter, J. L., Mcculloch, M. T. & Clode, P. L. Ocean acidification causes structural deformities in juvenile coral skeletons. Sci Adv 2, e1501130–e1501130 (2016).
Suwa, R. et al. Effects of acidified seawater on early life stages of scleractinian corals (Genus Acropora). Fisheries Sci 76, 93–99 (2010).
Baria, M. V. B., Kurihara, H. & Harii, S. Tolerance to elevated temperature and ocean acidification of the larvae of the solitary corals Fungia fungites (Linnaues, 1758) and Lithophyllon repanda (Dana, 1846). Zool Sci 32, 447 (2007).
Desalvo, M. K. et al. Differential gene expression during thermal stress and bleaching in the Caribbean coral Montastraea faveolata. Mol Ecol 17, 3952–3971, https://doi.org/10.1111/j.1365-294X.2008.03879.x (2008).
Reyes-Bermudez, A., Lin, Z. Y., Hayward, D. C., Miller, D. J. & Ball, E. E. Differential expression of three galaxin-related genes during settlement and metamorphosis in the scleractinian coral Acropora millepora. Bmc Evol Biol 9, doi:Artn 17810.1186/1471-2148-9-178 (2009).
Moya, A. et al. Whole Transcriptome analysis of the coral Acropora millepora reveals complex responses to CO2-driven acidification during the initiation of calcification. Mol Ecol 21, 2440–2454 (2012).
Rocker, M. M. et al. Expression of calcification and metabolism-related genes in response to elevated pCO2 and temperature in the reef-building coral Acropora millepora. Mar Genom 24, 313–318 (2015).
Moya, A. et al. Rapid acclimation of juvenile corals to CO2-mediated acidification by upregulation of heat shock protein and Bcl-2 genes. Mol Ecol 24, 438–452 (2015).
Kaniewska, P. et al. Major cellular and physiological impacts of ocean acidification on a reef building coral. Plos One 7, e34659 (2012).
Masako, N., Masaya, M., Haruko, K. & Satoshi, M. Expression ofhsp70, hsp90 and hsf1 in the reef coral Acropora digitifera under prospective acidified conditions over the next several decades. Biol Open 1, 75–81 (2012).
Putnam, H. M., Mayfield, A. B., Fan, T. Y., Chen, C. S. & Gates, R. D. The physiological and molecular responses of larvae from the reef-building coral Pocillopora damicornis exposed to near-future increases in temperature and pCO2. Mar Biol 160, 2157–2173 (2013).
Ogawa, D., Bobeszko, T., Ainsworth, T. & Leggat, W. The combined effects of temperature and CO2 lead to altered gene expression in Acropora aspera. Coral Reefs 32, 895–907, https://doi.org/10.1007/s00338-013-1046-9 (2013).
Babcock, R. C. Comparative demography of three species of scleractinian corals using age-and size-dependent classifications. Ecol Monogr 61, 225–244 (1991).
Hughes, T. P. & Jackson, J. B. C. Population dynamics and life histories of foliaceous corals. Ecol Monogr 142–166 (1985).
Albright, R., Mason, B. & Langdon, C. Effect of aragonite saturation state on settlement and post-settlement growth of Porites astreoides larvae. Coral Reefs 27, 485–490 (2008).
Albright, R. & Langdon, C. Ocean acidification impacts multiple early life history processes of the Caribbean coral Porites astreoides. Glob Change Biol 17, 2478–2487 (2011).
Albright, R., Mason, B., Miller, M. & Langdon, C. Ocean acidification compromises recruitment success of the threatened Caribbean coral Acropora palmata. Proc Natl Acad Sci USA 107, 20400–20404 (2010).
Doropoulos, C., Ward, S., Marshell, A., Diaz-Pulido, G. & Mumby, P. J. Interactions among chronic and acute impacts on coral recruits: the importance of size-escape thresholds. Ecology 93, 2131–2138 (2012).
Debashish, B. et al. Comparative genomics explains the evolutionary success of reef-forming corals. Elife 5, e1328 (2016).
Zoccola, D. et al. Bicarbonate transporters in corals point towards a key step in the evolution of cnidarian calcification. Sci Rep 5, 9983 (2015).
Kenkel, C. D., Meyer, E. & Matz, M. V. Gene expression under chronic heat stress in populations of the mustard hill coral (Porites astreoides) from different thermal environments. Mol Ecol 22, 4322–4334 (2013).
Schnetkamp, P. P. The SLC24 gene family of Na+/Ca²+-K+ exchangers: from sight and smell to memory consolidation and skin pigmentation. Mol Aspects Med 34, 455 (2013).
Mbalaviele, G., Chan, S. S. & Civitelli, R. Perspective: Cell–cell adhesion and signaling through cadherins: Connecting bone cells in their microenvironment. J Bone & Mineral Research 21, 1821–1827 (2007).
Heino, J., Huhtala, M., Käpylä, J. & Johnson, M. S. Evolution of collagen-based adhesion systems. Int J Biochem Cell B 41, 341–348 (2009).
Mass, T. et al. Cloning and characterization of four novel coral acid-rich proteins that precipitate carbonates in vitro. Curr Biol 23, 1126–1131 (2013).
Pierrot, D., Lewis, E. & Wallace, D. W. R. MS Excel program developed for CO2 system calculations. ORNL/CDIAC-105, Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy, Oak Ridge, Tennessee (2006).
Mehrbach, C., Culberson, C. H., Hawley, J. E. & Pytkowicz, R. M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol Oceanogr 18, 897–907 (1973).
Grabherr, M. G. et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol 29, 644 (2011).
Conesa, A. et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676 (2005).
Li, T. et al. Comparative Transcriptome analysis of Penicillium citrinum cultured with different carbon sources identifies genes involved in citrinin biosynthesis. Toxins 9, 69 (2017).
Xiang, L. X., Ding, H., Dong, W. R., Zhang, Y. W. & Shao, J. Z. Deep sequencing-based transcriptome profiling analysis of bacteria-challenged Lateolabrax japonicus reveals insight into the immune-relevant genes in marine fish. Bmc Genomics 11, 472 (2010).
Acknowledgements
We acknowledge support from the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA13020403), the Science and Technology Planning Project of Guangdong Province, China (2014B030301064), NSFC Projects (31370499, 41106107), and the National Key Technology R&D Program (2014BAC01B03).
Author information
Authors and Affiliations
Contributions
X.C.Y. wrote the manuscript with all authors commenting. T.Y., H.H., L.J., W.H.Z., S.L. performed the experiment and designed the research.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yuan, X., Yuan, T., Huang, H. et al. Elevated CO2 delays the early development of scleractinian coral Acropora gemmifera. Sci Rep 8, 2787 (2018). https://doi.org/10.1038/s41598-018-21267-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-21267-3
This article is cited by
-
Direct and latent effects of ocean acidification on the transition of a sea urchin from planktonic larva to benthic juvenile
Scientific Reports (2022)
-
The indirect effects of ocean acidification on corals and coral communities
Coral Reefs (2022)
-
Promoting larval settlement of coral Pocillopora damicornis by calcium
Coral Reefs (2022)
-
Larvae of a marine gastropod and a marine bivalve share common gene expression signatures during metamorphic competence
Marine Biology (2022)
-
Developmental series of gene expression clarifies maternal mRNA provisioning and maternal-to-zygotic transition in a reef-building coral
BMC Genomics (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
