
Abstract
Enterotoxigenic Escherichia coli (ETEC) cause more than 500,000 deaths each year in the developing world and are characterized on a molecular level by the presence of genes that encode the heat-stable (ST) and/or heat-labile (LT) enterotoxins, as well as surface structures, known as colonization factors (CFs). Genome sequencing and comparative genomic analyses of 94 previously uncharacterized ETEC isolates demonstrated remarkable genomic diversity, with 28 distinct sequence types identified in three phylogenomic groups. Interestingly, there is a correlation between the genomic sequence type and virulence factor profiles based on prevalence of the isolate, suggesting that there is an optimal combination of genetic factors required for survival, virulence and transmission in the most successful clones. A large-scale BLAST score ratio (LS-BSR) analysis was further applied to identify ETEC-specific genomic regions when compared to non-ETEC genomes, as well as genes that are more associated with clinical presentations or other genotypic markers. Of the strains examined, 21 of 94 ETEC isolates lacked any previously identified CF. Homology searches with the structural subunits of known CFs identified 6 new putative CF variants. These studies provide a roadmap to exploit genomic analyses by directing investigations of pathogenesis, virulence regulation and vaccine development.
Similar content being viewed by others

Introduction
The enterotoxigenic Escherichia coli (ETEC) pathogenic variant (pathovar) has been implicated in 1 billion cases of diarrhea annually1,2,3. These pathogens are especially problematic in ETEC endemic areas, such as Bangladesh4, 5. ETEC are characterized on a molecular basis by the presence of genes that encode the heat-stable (ST) and/or heat-labile (LT) enterotoxin6, 7. Both toxins activate the cystic fibrosis trans-membrane regulator (CFTR) that results in ion secretion followed by water and diarrhea in infected individuals8. In addition to the enterotoxins, ETEC possess fimbrial appendages that attach to intestinal epithelium known as colonization factors (CFs). Most ETEC-specific virulence factors including the CFs are plasmid-encoded, with greater than 30 CFs described in the literature9, 10.
In addition to known ETEC virulence factors, other putative virulence factors have been identified, primarily in the prototypical ETEC isolate, H1040711. These factors include the adhesin autotransporter TibA12, the invasion locus tia 13, and leoA 14, which has been associated with maximum LT secretion. Additionally, the serine protease autotransporter EatA15, which has only been identified in ETEC isolates16, accelerates delivery of LT by degrading MUC2, the major mucin secreted by gastrointestinal goblet cells17, and by modulating adhesion mediated by EtpA18. EtpA is a glycoprotein that appears to act as a bridge between FliC and host surface structures19, 20. In addition to these previously identified ETEC-associated factors, a study by Vidal et al.21 suggested that an iron acquisition system (irp2, fyuA) most similar to a system in Yersinia species22, 23 may play a role in ETEC virulence based on its variable presence in clinical ETEC isolates24.
The majority of the functional work in ETEC has focused on the prototype isolate H1040725, 26. However, detailed transcriptional studies using RNA-seq have demonstrated that the response to environmental and host signals, such as bile and carbohydrates, can vary widely between ETEC isolates27. Additionally, it had been suggested that some putative virulence factors identified in H10407 are not widely distributed among diverse ETEC isolates28. This suggests that the genomic diversity within ETEC isolates is significant and a reference-independent global approach is required to comprehensively characterize the genomic diversity.
The advent of large-scale sequencing has increased our understanding of the evolution of the members of the ETEC pathovar. Until 2014, there were relatively few sequenced and assembled human-associated ETEC isolates, all from symptomatic patients, available in Genbank11, 16, 29, as well as, four porcine ETEC isolates that had also been sequenced30. A recent study by von Mentzer et al. in 2014 utilized a genomic mapping approach of unassembled genomes for the examination of genome similarity in a collection of 362 global E. coli isolates31. The isolates sequenced in the von Mentzer et al. study were selected for the greatest variability of colonization factor and enterotoxin profiles from a historical collection ranging from 1980 and 2011 of ETEC isolates maintained at the University of Gothenburg. The current study examines a collection of 94 ETEC isolates, 89 of which represent circulating isolates within Bangladesh between 2002 and 2011. Additionally, 84 isolates were obtained from individuals with diarrhea (symptomatic ETEC) and 10 isolates obtained from asymptomatic ETEC colonization. Comparative genomics of this wealth of information is providing novel insights into the evolution and distribution of ETEC virulence factors.
Although large-scale sequencing projects can now rapidly generate a data from large numbers of isolates, informatics pipelines and comparative analyses to take advantage of these large-scale genomic data have languished. These types of genomic epidemiology studies have been recently completed with other E. coli pathovars32,33,34, but this study provides a further example of the application of this comparative analysis paradigm to isolates from the ETEC pathovar.
Results
Core genome single nucleotide polymorphism (SNP) phylogeny of ETEC
To examine the phylogenetic relationship of the sequenced ETEC isolates in the broader context of diverse E. coli and Shigella spp., a SNP-based phylogeny was inferred from ~220,000 SNPs from 136 E. coli/Shigella genomes (Supplementary Data File 1). The genomes from this study include the 94 ETEC genomes sequenced in this study, eight previously sequenced ETEC reference genomes, 34 reference E. coli and Shigella genomes representing prototype members of each of the diarrheagenic pathogenic variants (Metadata and GenBank Accession numbers are included in Table S1). The results demonstrate that the majority of ETEC genomes fall into the E. coli phylogroups A or B1 (Fig. 1), with one genome, isolate 2845650, falling into phylogroup E; ETEC isolates from this phylogroup have been previously described31. The retention index (RI) value of 0.82 for this tree was determined using Phangorn, suggesting significant homoplasy, likely resulting from homologous recombination.

A core genome single nucleotide polymorphism (SNP) phylogeny of ETEC genomes sequenced in this study as well as reference E. coli genomes. SNPs were identified by NUCmer59 alignments of query genomes against the reference genome, K-12 W3110; these methods were wrapped by the NASP pipeline62. A phylogeny was inferred on the concatenated SNP alignment using RAxML v860 including 100 bootstrap replicates. ETEC genomes sequenced in this study were assigned to disease groups based on clinical observations at site of isolation or presented in literature.
The phylogenetic diversity of sequenced ETEC isolates was remarkable (Fig. 1). The majority of the diarrheagenic ETEC (symptomatic ETEC) sequenced in this study were isolated from Dhaka, Bangladesh between 2002 and 2011, yet based on the relatedness displayed in the phylogeny, the isolates are broadly distributed across the known diversity in E. coli. This highlights that the genetic background of E. coli is generally amenable to the uptake and maintenance of ETEC plasmids and virulence factors. However, no currently sequenced ETEC genomes, of the ~450 ETEC isolates, are present within Phylogroup B2, which suggests that this particular genetic background is not amenable for the uptake or retention of ETEC plasmids. The study by von Mentzer et al.31 suggested that three B2 ETEC genomes were identified however, one of the reported B2 genomes (E1642) was not B2 by our phylogenomic analysis (Figure S1), and the other two (E523, E2439) were negative for all queried ETEC enterotoxins (0% toxin coverage at a minimum depth of 2x), indicating that they were not ETEC based on the sequence data that is publically available.
Despite this overall phylogenetic diversity, strains isolated from geographically and temporally dispersed cases of cholera-like illness were phylogenetically similar. These results taken as a whole indicate that particular combinations of pathovar-specific genes and genomic backgrounds may be optimal for survival, virulence and transmission as determined by the prevalence of the combinations in these and other studies31; however, detailed transcriptional studies are required for this to be elucidated.
Bioinformatic analysis of symptomatic ETEC and asymptomatic ETEC for known and putative virulence factors
The ETEC isolates sequenced in this study were identified as members of this pathotype, as they contained one or more of the enterotoxin genes (LT, STh and/or STp) (Fig. 2, Table S3). The presence of additional known and predicted virulence factor genes was determined using LS-BSR35. The results demonstrate that the previously identified virulence factors of ETEC do not cluster among either the asymptomatic ETEC or symptomatic ETEC isolates (Fig. 2). Many of the virulence factors (e.g. leoA) identified in the prototype isolate H1040711 were sparsely distributed among the 94 isolates sequenced in this study (Fig. 2). The etpA gene was included in the initial analyses, but the five repeat regions in the 3′ end of the gene, each ~600 nucleotides, confounded proper assembly of this genomic region and precluded an accurate estimation of etpA conservation. Proteomic analysis36 of the strains corresponding to the genomes analyzed here found that EtpA was produced by more than 60% of the isolates, and that the sizes of the secreted peptides were similar to that reported for H10407 (~170 kD). Thus, etpA was removed from downstream in silico analyses.

A core genome single nucleotide polymorphism (SNP) phylogeny of ETEC genomes sequenced in this study as well as reference E. coli genomes associated with a heatmap of BSR values of previously-characterized virulence and colonization factors (Table S3). Disease categories were assigned based on clinical observations. Orange brackets around genomes indicate lineages (Groups 1–3) compared to identify coding regions associated with the observed clinical presentations. The heatmap was associated with the phylogeny using the interactive tree of life (47).
In examining correlations between individual known and predicted virulence factors and the corresponding clinical presentation associated with the respective isolate, no single virulence factor or gene region segregated exclusively with symptomatology or phylogenomic group (Fig. 2). Thus, there is not a single proposed virulence factor that can conclusively and consistently distinguish ETEC isolates that are diarrhea-associated or only from an asymptomatic colonization.
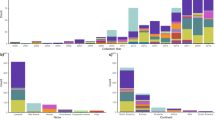
The data analysis did reveal correlations between certain combinations of virulence factors, CF types, and phylogenomic location suggesting that some strains may possess a suite of features that are more optimal for human infection. The boxes highlighted in Fig. 2 demonstrate phylogenetic groups of isolates that have similar genomic content as defined by the phylogeny, and similar virulence factor profiles. As highlighted in Fig. 2, there are three monophyletic groups with a significant number of isolates that have similar virulence and colonization factor profiles, but are variable for the clinical presentations. These groups are labeled groups 1–3 in this study and can be correlated, but not perfectly matched to the previously identified dominant lineages 5, 1/2 and 3, respectively from von Mentzer et al.31. The isolates in Group 1 are dominated by symptomatic ETEC isolates, the majority of which contain CS5 and CS6 (Figure S2). In contrast, the asymptomatic isolates from this study in this phylogenomic group are only positive for a novel CF identified in the current study (see below). The distinction in this phylogenomic group based on clinical presentation is extended to the virulence factors, as all the symptomatic isolates are generally LT and ST positive (22/29 isolates), whereas the asymptomatic isolates are only LT positive (Figure S2). Interestingly, the symptomatic isolates in this phylogenomic cluster are also positive for the regulator known as peaR 27, which has homology to the rns regulator which is involved in colonization factor regulation37. Group 2 isolates possess genes for CS1, CS3 and CS21 (Figure S2). Group 2 isolates contain both symptomatic ETEC and asymptomatic ETEC isolates (13 versus 7, respectively). One of the largest phylogenetic groups, Group 3 in Figs 1 and 2, consisted primarily of isolates from symptomatic infections, with only one isolate obtained from asymptomatic colonization. This group contains primarily colonization factors CFA/I or CS7, and encoded LT and ST genes. Overall, these examples demonstrate that there is significant genomic and virulence factor diversity among these isolates, but there are also common combinations of virulence factors and genomic backgrounds that may create a more optimal pathogen or allow the isolate to survive in the environment. These more common clones will need to be examined functionally, via detailed mutagenesis, transcriptional and virulence assays to test these hypotheses.
Genomic epidemiology comparison of symptomatic ETEC and asymptomatic ETEC genome content
To identify any genomic regions unique to symptomatic ETEC isolates when compared to asymptomatic ETEC isolates, a whole-genome large-scale blast score ratio (LS-BSR)35 comparison was conducted on the ETEC genomes sequenced in this study as well as a set of previously sequenced ETEC genomes (Table S1). The results demonstrate that no coding regions (CDSs) were exclusive to either asymptomatic ETEC (n = 23) or symptomatic ETEC (n = 262).
While no features could be identified that were exclusive in each of the clinical presentations, a statistical approach identified genomic regions that were associated with either the asymptomatic ETEC or symptomatic ETEC. The LS-BSR data set generated here (Supplementary Data File 2) was examined for the prevalence of gene features with either the clinical symptomology or phylogenetic distribution. When the genomes were examined for features that were associated with symptomatic ETEC or asymptomatic ETEC, we identified 28 features that were statistically (FDR p value < 0.05) associated with symptomatic ETEC (Table S4) and 28 features statistically associated with the asymptomatic ETEC (Table S4). These features highlight the use of genomic epidemiology for the identification of features associated with virulence. The genes associated with symptomatic ETEC isolates included a number of hypothetical proteins, phage related genes, as well as transcriptional regulators (Table S4). Coding regions that were associated with the asymptomatic isolates primarily included hypothetical proteins (Table S4). While none of these features stand out as obvious virulence factors, and do not demonstrate a difference in prevalence between ETEC and other pathovar isolates, they do provide additional evidence of features that may play a role in the interactions of the host and pathogen. These in silico studies provide a starting point for the functional analysis of these genes and the potential role they play in virulence.
Our group has previously identified that there is a component of the ETEC genomes is phylogenomically linked16, 27. While these earlier calculations were based on far fewer genomes, the general trends of those studies are confirmed in this study, as the majority of ETEC isolates were identified in phylogroups A or B1 (Fig. 1), with only one ETEC genome in this study identified in phylogroup E. As with the clinical presentation comparisons, we do not observe canonical virulence factors in these groups, but rather a number of genes that could serve as accessory functions to virulence.
In addition to a global comparison of all symptomatic ETEC and asymptomatic ETEC, we also attempted to identify specific regions unique to symptomatic ETEC in two lineages (group 1 and 2) of the inferred phylogeny (Figures 1 and 2), chosen due to the substantial ETEC representation on these lineages (Figure S2). This analysis was performed to identify genomic regions that could explain differences in clinical presentation among patients colonized with phylogenetically-related isolates. Genomic regions were identified in these two clades that could largely differentiate the asymptomatic ETEC and symptomatic ETEC isolates (Table 1). The asymptomatic ETEC and symptomatic ETEC in group 2 contained 16 and 9 coding regions, respectively, that could be used to distinguish isolates based on clinical presentation on this branch (Supplementary Data File 3). Many of these features are associated with plasmids (Table S4), suggesting that divergent plasmids, or parts of plasmids, may have been acquired by the isolates in these two different groups. However, because the isolates were not collected contemporaneously, (Table S1), we cannot exclude temporal effects on genome content and it remains unclear whether these regions are explicitly associated with pathogenesis.
No single gene feature distinguished ETEC associated with symptomatic illness or asymptomatic colonization. However comparative analyses within phylogenetic groups did identify genes that statistically segregate with the clinical presentation, thus providing stable genomic encoded targets for virulence studies, functional characterization and/or additional phylogenomic features for use as diagnostic markers, as we have previously done in other E. coli pathovars38, 39.
Comparison of ETEC and non-ETEC genomes
A previous study of E. coli comparative genomics using far fewer genomes, demonstrated pathovar-specific genome conservation among ETEC isolates16. The earlier study utilized a reference-based method that included only seven symptomatic ETEC genomes, a relatively small number of genomes compared to those analyzed in this study and the von Mentzer study31. Using a global approach, the genomic content of ETEC and non-ETEC isolates was examined in E. coli phylogroups A and B1, where the majority of ETEC isolates are located. A total of 506 genomes from phylogroups A and B1 (253 ETEC and 253 non-ETEC; Table S2) were compared in a LS-BSR analysis. While most coding regions were distributed between groups, outliers (n = 118) were identified (Table 2, Table S4; Fig. 3). Several of the ETEC-specific regions were associated with the ETEC toxins and putative plasmid components, as expected (Table S4). However, the non-ETEC genomes contained coding regions with various functional annotations, including features of central metabolism and type III secretion genes, suggesting that a specific genomic background and selective pressures are involved in the acquisition and retention of ETEC plasmids that harbor enterotoxins, as well as the non-ETEC virulence factors.

In silico colonization factor identification
Functional screening for common CF types (Table S3, Fig. 2) identified a significant proportion of the isolates (n = 21 of 94) sequenced in this study did not encode a recognizable CF (peptide BSR value < 0.90). However, several sequences from these isolates did share homology (peptide BSR > 0.50) with the structural subunits from known CFs (Table S3)9. Further analysis of contigs from genomes encoding potential CF structural subunits identified extended homology to previously characterized CF gene clusters. Phylogenetic (Fig. 4A) and direct amino acid comparisons to previously annotated CFs identified six new putative colonization factors (Table 3, Fig. 4B). Based on divergence in the peptide sequences of structural subunits these putative colonization factors were named pcf (putative colonization factor), b (Bangladesh), and a number [1–6]. When the complete CF gene cluster structure of each new CF was examined, a similar gene order and cluster structure was observed (Fig. 4B). The contigs in the draft assembly that contained these gene clusters often only contained the CF cluster structure; in the case of pcf_b03, pcf_b04, and pcf_b05, homologs were seen for aalR and aalA (part of the CS23 CF operon), but were present on different contigs. This suggests that these CF-containing regions are flanked by regions that were not resolved during the genome assembly, and are possibly repetitive elements or insertion sequences, as has been highlighted as a common genomic feature in previous studies11, 29. Of the six novel CF clusters, three putative CF clusters showed limited homology ( < 50% AA identity) to the CS23 CF40.

Analysis of novel putative colonization factors (CFs) identified in isolates sequenced in the current study. (A) A phylogenetic tree inferred from an alignment of peptide sequences from previously described CF major structural subunits, shown in black, and sequences from new putative CFs, shown in red. Sequences were aligned with MUSCLE72 and a phylogeny was inferred with RAxML60 with 100 bootstrap replicates. (B) Structural organization of novel putative CFs. Reference CFs were used to organize novel putative CFs. Numbers indicate the percent BLAST identity of protein sequences. The structure of novel putative CFs were identified from Prokka71 annotation.
A LS-BSR screen of novel putative CFs against a collection (n = 223) of confirmed ETEC genomes (this study and31, demonstrated that pcf_b02 was conserved (peptide BSR value > 0.95) in only four genomes and pcf_b06 was conserved in only two genomes (Supplementary Data File 4); all other novel CFs were absent from this genomic dataset. This demonstrates that some of the putative CFs are broadly distributed temporally and geographically, while the other CFs may be unique or specific to Bangladesh.
When the novel putative CFs (Table 3) were combined with known CFs (Table S3), 91 of the 94 isolates sequenced in this study were assigned a CF type based on a peptide BSR value >= 0.98 (Fig. 2, Table S1); however, three isolates were still classified as CF negative, based on an absence of homology to known ETEC CFs. Collectively, these findings reaffirm the critical role of CFs in mediating intestinal colonization by ETEC, while highlighting the potential complexity in targeting CF antigens in vaccines.
Discussion
Outcomes of ETEC infection in humans range from asymptomatic colonization and mild self-limiting diarrhea to severe cholera-like disease41, 42. Despite the prevalence of this pathovar in the developing world, current knowledge of genomic diversity of ETEC derives from screening culture collections for a limited number of virulence markers. The majority of these markers and virulence genes were identified or characterized in a single prototype ETEC isolate, H10407. However, this isolate appears to contain a gene repertoire that is not entirely representative of the majority of ETEC isolates surveyed16, 28, 31. In the current study, a diverse set of ETEC isolates were obtained and sequenced from individuals primarily from Bangladesh. These genomes were then compared in a reference-independent approach to understand the distribution of virulence and colonization factors. With the development of massively parallel genome sequencing, genomic comparisons are no longer reliant on comparisons to limited numbers of prototype reference isolates.
ETEC genomic analyses to date have almost exclusively focused on pathogenic isolates11, 16, although asymptomatic isolates have been described extensively in the literature31, 43,44,45. In the current study, the sequenced isolates include 84 ETEC isolates from individuals with diarrhea (symptomatic ETEC) and 10 ETEC isolates from asymptomatic individuals (asymptomatic ETEC); by including public genomes31, the numbers increased to 262 symptomatic and 23 asymptomatic genomes. Although the relatively low number of asymptomatic ETEC isolates precludes large-scale investigations into genes associated with disease, this study presents a framework of how to analyze large multidimensional datasets to identify genomic features positively associated with a given phenotype, such as disease presentation. These studies highlight the utility of moving beyond the single gene approach and taking a more systems biology approach to the study of pathogenesis.
The majority of the genomes generated in this study were isolated from Bangladesh and are associated with the clinical presentation at the time of isolation. Whole genome analyses attempted to identify genomic features that are conserved in isolates from individuals with diarrhea and absent in isolates from asymptomatic individuals. However, it is possible that an isolate currently identified as asymptomatic is actually a virulent isolate, to which the host has immunity, rather than a truly avirulent isolate. Multiple host factors, including nutrition and gastrointestinal microbiota composition, as well as multiple immune mediators, and blood group antigen presentation on mucosal surfaces may be as important as pathogen virulence factor content in determining clinical presentation. Collectively, host factors that impact the outcome of ETEC infections could confound identification of genomic region(s) exclusive to symptomatic ETEC isolates.
ETEC CFs have previously been associated with ETEC pathogenesis26 and have been the focus of intensive research and multiple vaccine trials46. In this study of 94 ETEC isolates, 23 CFs were identified, including six novel putative CFs. The identification of these six new CFs increases the known CF repertoire by ~25% and highlights the utility of whole genome sequencing in the identification of novel variants of important virulence factors. Additional work is needed to functionally characterize these new CFs and their potential contribution to virulence in those isolates. The sequence diversity observed, even this relatively small collection of isolates, highlights the challenges to the development of a broadly protective ETEC CF-based vaccine47. Interestingly, three symptomatic ETEC isolates were identified that can be characterized as truly CF negative, suggesting that these may possess previously uncharacterized adhesin molecules.
The whole genome phylogeny demonstrated the breadth of phylogenetic diversity of the ETEC pathovar in Bangladesh (Fig. 1, Figure S1). ETEC is one of the most diverse pathotypes5, 31 and have now been identified in all E. coli phylogroups, with the exception of B2 (Fig. 1). Phylogroup B2 is populated mainly with extra-intestinal E. coli and more specifically, urinary tract E. coli, and as such they do contain unique genes compared to the diarrheagenic E. coli phylogroups35, but it is unclear if these unique genes confer greater incompatibility with ETEC plasmids harboring enterotoxins.
This observed phylogenetic diversity is mostly likely driven by the significant number of mobile elements including phage and plasmids within the ETEC genomes, as well as the fact that all of the pathovar specific virulence factors are encoded on plasmids11, 29. While a detailed analysis of the complete plasmids is not possible with this dataset due to the sequencing method and the previous studies that indicated that there the ETEC genome contains ~5% repetitive elements and insertion sequences29, 31. However, it must be noted that a number of reference ETEC isolates31 included in this study are from outbreaks that occurred many decades apart and on different continents (Table S1 and S2), and yet the phylogenetic (Fig. 1 and Figure S1) and virulence factor patterns (Fig. 2) are very similar suggesting that there may be an optimal strategy of pathogenesis in these organisms that leads to a successful pathogen that can cause significant outbreaks. One hypothesis could be that there is constant sharing of genetic material among the non-pathogenic or evolving-pathogenic isolates within a host or the environment that only expands significantly when there is the optimal assemblage of the mobile genetic factors, bacterial chromosomal factors, and the opportunity to infect a susceptible population or host.
The results of this study have identified several new putative CFs, as well as a number of genomic regions differentially present in isolates from different clinical presentations among the ETEC in Bangladesh, as well as between ETEC and non-ETEC E. coli isolates. Importantly, this study highlights the difficulty of directly correlating pathogen genomics with clinical outcomes. However, these studies demonstrate the feasibility of large-scale genomic epidemiology as an essential tool for molecular characterization of these globally important pathogens.
Materials and Methods
Strain selection
A total of 94 isolates, 84 associated with moderate to severe diarrheal illness and 10 from asymptomatic colonization were analyzed in this study. The majority of isolates (n = 89) were collected at the International Centre for Diarrhoeal Disease Research (http://www.icddrb.org) in Dhaka, Bangladesh in between 2002 and 2011. Effort was taken to minimize the number of passages of these cultures to prevent plasmid loss or the loss of any other unstable genomic feature. These isolates provide insight into the circulating isolates in Bangladesh between 2002 and 2011. Four additional isolates, Envira 8/11, Envira 10/1, Juruá 18/2, and Juruá 20/10, obtained from outbreaks of severe, cholera-like ETEC diarrhea in the Amazon, were kindly provided by Ana C.P. Vicente48, and a single strain (ThroopD) from the USA, isolated from a patient with severe diarrhea49 was obtained courtesy of Richard Finkelstein. These five isolates represent reference ETEC isolates that have been characterized previously in the literature. Disease severity associated with individual isolates was assigned based on the clinical presentation at the time of isolation. ETEC isolates were confirmed from lactose-fermenting colonies based on assays for heat labile and heat-stable toxin genes, as described previously50. For some of these specimens, multiple isolates were sequenced per patient51; however, for this analysis, only a single non-redundant representative is examined to remove redundancy in the current dataset.
Genomes from a study on ETEC diversity by von Mentzer et al.31 were downloaded from the Sequence Read Archive (Accession: ERP000733). Reads were mapped to the three ETEC enterotoxins (Supplemental Table 3) with BWA-MEM52 and the per base depth of coverage was calculated with the GenomeCoverageBed method in BEDTOOLS53. The breadth of coverage, or percentage of each target that was covered by a minimum number of reads, was then calculated. Isolates having appropriate coverage on one of the toxin genes were considered confirmed ETEC genomes. The breadth of coverage analysis demonstrated that only 309 of these genomes contained one of the three ETEC enterotoxins at a minimum breadth of 80% (2x minimum depth of coverage). The genomes were subsequently assembled with SPAdes v3.6.054. Genomes with an anomalous assembly size (<4.5 mb) or a large number of contigs (>500) were removed from the analysis. Finally, a core genome phylogeny was inferred with all remaining genomes, including genomes sequenced in this study and external references (Figure S1). Genomes with a clade designation that differed from our results were removed (Figure S1). Following this filtering pipeline, only 223 of the original 362 (61.6%) genomes from the study by von Mentzer et al.31 were used in downstream comparative genomic analyses (Table S1), including genomes associated with symptomatic (n = 178) and asymptomatic (n = 34) disease presentations.
DNA extraction, sequencing, assembly
Genomic DNA was extracted with standard methods55 and sequenced on the Illumina HiSeq 2000 platform at the University of Maryland School of Medicine, Institute for Genome Sciences, Genome Resource Center using established SOPs. The resulting 100 bp reads were assembled with the Celera Assembler56; the resulting assemblies and corresponding accession numbers are shown in Table S1. In silico multi-locus sequence type (MLST) profiles were assigned with a public script (https://github.com/Victorian-Bioinformatics-Consortium/mlst) against the pubMLST E. coli database57. As the Celera Assembler will remove contigs with an anomalous coverage, all genomes were also assembled with SPAdes v3.6.
Core genome single nucleotide polymorphism (SNP) phylogeny
SNPs were identified from all genome assemblies compared to K-12 W311058. Each query genome assembly was aligned to the reference with NUCmer59 and a direct mapping of query to reference was constructed. A self-alignment was conducted for the reference with NUCmer and any SNPs falling within duplicated regions were filtered from subsequent analyses. All identified SNPs (n = 220,679) in all genomes (Supplementary Data File 1) compared to the reference were concatenated. A phylogeny was inferred on this concatenated SNP alignment with RAxML v860 using the ASC_GTRGAMMA substitution model (Lewis correction61). The NASP pipeline that wraps all SNP identification methods is publically available (http://github.com/TGenNorth/NASP)62. The Retention Index63, which calculates the consistency of SNPs to a tree topology, was calculated with Phangorn64.
Identification of an ETEC genomic core
Previously we reported the presence of an ETEC pathovar-specific genomic core16 using a reference-based approach. The current study utilizes a greater number of genomes with a reference-independent approach to identify genes differentially present in ETEC genomes. As outlined above, the 94 isolates from this study, as well as additional ETEC genomes from von Mentzer et al.31 were downloaded from public databases, assembled with SPAdes v3.6.054, checked for the presence of one or more ETEC enterotoxins, and the resulting phylogenetic tree was inferred with FastTree265. ETEC genomes were identified as belonging to phylogroups A or B1 based on location in the phylogenetic tree and previously typed historical isolates; corresponding non-ETEC genomes from these same phylogenetic groups were identified from PATRIC66. For phylogenetic clades A and B1, 253 ETEC genomes and 253 reference genomes (Table S2) were tested for common genes using a LS-BSR approach35. Genomic regions with a differential distribution in these phylogroups were identified using an R67 script (https://gist.github.com/jasonsahl/569e502a66dcab5c643f).
Identification of ETEC genes associated with clinical presentation
ETEC genomes and reference genomes (Table S1) were tested for gene presence and absence using the LS-BSR approach35. Genomic regions with a differential distribution based on the observed clinical presentation were identified from the outlier R script, with p-values using a Kruskal-Wallis test (https://gist.github.com/jasonsahl/6dd3939d3bb8c83f74f5ec5eac665280#file-kruskal_wallis_v1-py). A False Detection Rate (FDR) adjusted p-value < 0.05 was considered significant.
Identification of phylogenomic specific regions
In addition to a global comparison of all asymptomatic ETEC and symptomatic ETEC, a comparison was also made on selected individual lineages from the whole genome phylogeny that contain a combination of isolates from each clinical presentation (Table S1). Genomes in specific lineages were interrogated with the LS-BSR approach. Genomic regions with a differential distribution based on the observed clinical presentation were identified and the statistical test used was the Kruskal-Wallis test. Any FDR corrected p-value < 0.05 was considered significant.
Colonization factor identification
A set of 21 common/previously identified ETEC colonization factors (Table S3) was collated and compared against the 136 genomes in the phylogeny (Fig. 1) with TBLASTN68 in order to identify the global CF profile. The BSR value69 was identified for each CF in each genome then visualized with the integrated tree of life (iTOL)70; regions with a BSR value of >0.9 were considered to be present. Contigs from draft assemblies that showed more remote homology (<90% peptide ID) to known CFs were annotated with Prokka71 to identify the operon structure of the homologous region.
Structural subunits from novel putative CFs were identified from the predicted coding sequences and annotation of each genome. Peptide sequences associated with previously identified CF structural subunits (Table S3) were aligned with new putative subunits identified in this study using MUSCLE72, and a phylogenetic tree was inferred with RAxML using the BLOSUM62 substitution matrix and 100 bootstrap replicates.
in silico virulence factor screen of ETEC genomes
All verified and predicted ETEC virulence factors were compiled for interrogation of the genomes generated in this study (Table S3). The peptide sequence for each factor was aligned against all sequenced genomes with TBLASTN in conjunction with LS-BSR. The BSR value was calculated, and genes with a BSR ≥ 0.90 were considered to be highly conserved. The proportion of genes that were determined to be highly conserved in each group (clinical presentation, or phylogenomic group etc) was compared with a two-tailed significance test and the p-value was calculated from the resulting z-score.
Accession numbers
The genome sequences generated in this study are deposited in GenBank under the accession numbers listed in Table S1.
References
WHO. Future directions for research on enterotoxigenic Escherichia coli vaccines for developing countries. Wkly Epidemiol Rec 81, 97–104 (2006).
Lozano, R. et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380, 2095–2128 (2012).
Kotloff, K. L. et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 382, 209–222 (2013).
Qadri, F. et al. Disease burden due to enterotoxigenic Escherichia coli in the first 2 years of life in an urban community in Bangladesh. Infect Immun 75, 3961–3968 (2007).
Steinsland, H., Lacher, D. W., Sommerfelt, H. & Whittam, T. S. Ancestral Lineages of Human Enterotoxigenic Escherichia coli. J Clin Microbiol 48, 2916–2924 (2010).
So, M., Boyer, H. W., Betlach, M. & Falkow, S. Molecular cloning of an Escherichia coli plasmid determinant than encodes for the production of heat-stable enterotoxin. J Bacteriol 128, 463–472 (1976).
So, M., Dallas, W. S. & Falkow, S. Characterization of an Escherichia coli plasmid encoding for synthesis of heat-labile toxin: molecular cloning of the toxin determinant. Infect Immun 21, 405–411 (1978).
Sack, R. B. et al. Enterotoxigenic Escherichia coli isolated from patients with severe cholera-like disease. J Infect Dis 123, 378–385 (1971).
Gaastra, W. & Svennerholm, A. M. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol 4, 444–452 (1996).
Nada, R. A. et al. Discovery and phylogenetic analysis of novel members of class b enterotoxigenic Escherichia coli adhesive fimbriae. J Clin Microbiol 49, 1403–1410 (2011).
Crossman, L. C. et al. A commensal gone bad: complete genome sequence of the prototypical enterotoxigenic Escherichia coli strain H10407. J Bacteriol 192, 5822–5831 (2010).
Lindenthal, C. & Elsinghorst, E. A. Enterotoxigenic Escherichia coli TibA glycoprotein adheres to human intestine epithelial cells. Infect Immun 69, 52–57 (2001).
Fleckenstein, J. M., Kopecko, D. J., Warren, R. L. & Elsinghorst, E. A. Molecular characterization of the tia invasion locus from enterotoxigenic Escherichia coli. Infect Immun 64, 2256–2265 (1996).
Fleckenstein, J. M., Lindler, L. E., Elsinghorst, E. A. & Dale, J. B. Identification of a gene within a pathogenicity island of enterotoxigenic Escherichia coli H10407 required for maximal secretion of the heat-labile enterotoxin. Infect Immun 68, 2766–2774 (2000).
Patel, S. K., Dotson, J., Allen, K. P. & Fleckenstein, J. M. Identification and molecular characterization of EatA, an autotransporter protein of enterotoxigenic Escherichia coli. Infect Immun 72, 1786–1794 (2004).
Sahl, J. W. et al. A comparative genomic analysis of diverse clonal types of enterotoxigenic Escherichia coli reveals pathovar-specific conservation. Infect Immun 79, 950–960 (2011).
Kumar, P. et al. EatA, an Immunogenic Protective Antigen of Enterotoxigenic Escherichia coli, Degrades Intestinal Mucin. Infect Immun 82, 500–508 (2014).
Roy, K. et al. Adhesin degradation accelerates delivery of heat-labile toxin by enterotoxigenic Escherichia coli. J Biol Chem 286, 29771–29779 (2011).
Roy, K., Hamilton, D., Allen, K. P., Randolph, M. P. & Fleckenstein, J. M. The EtpA exoprotein of enterotoxigenic Escherichia coli promotes intestinal colonization and is a protective antigen in an experimental model of murine infection. Infect Immun 76, 2106–2112 (2008).
Roy, K. et al. Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 457, 594–598 (2009).
Del Canto, F. et al. Distribution of classical and nonclassical virulence genes in enterotoxigenic Escherichia coli isolates from Chilean children and tRNA gene screening for putative insertion sites for genomic islands. J Clin Microbiol 49, 3198–3203 (2011).
de Almeida, A. M. et al. Chromosomal irp2 gene in Yersinia: distribution, expression, deletion and impact on virulence. Microb Pathog 14, 9–21 (1993).
Jacobi, C. A., Gregor, S., Rakin, A. & Heesemann, J. Expression analysis of the yersiniabactin receptor gene fyuA and the heme receptor hemR of Yersinia enterocolitica in vitro and in vivo using the reporter genes for green fluorescent protein and luciferase. Infect Immun 69, 7772–7782 (2001).
Schubert, S., Rakin, A., Karch, H., Carniel, E. & Heesemann, J. Prevalence of the “high-pathogenicity island” of Yersinia species among Escherichia coli strains that are pathogenic to humans. Infect Immun 66, 480–485 (1998).
Hernandez, D., Francois, P., Farinelli, L., Osteras, M. & Schrenzel, J. De novo bacterial genome sequencing: millions of very short reads assembled on a desktop computer. Genome Res 18, 802–809 (2008).
Evans, D. G., Evans, D. J. Jr. & Tjoa, W. Hemagglutination of human group A erythrocytes by enterotoxigenic Escherichia coli isolated from adults with diarrhea: correlation with colonization factor. Infect Immun 18, 330–337 (1977).
Sahl, J. W. & Rasko, D. A. Analysis of the global transcriptional profiles of enterotoxigenic Escherichia coli (ETEC) isolate E24377A. Infect Immun (2012).
Turner, S. M. et al. Phylogenetic comparisons reveal multiple acquisitions of the toxin genes by enterotoxigenic Escherichia coli strains of different evolutionary lineages. J Clin Microbiol 44, 4528–4536 (2006).
Rasko, D. A. et al. The pangenome structure of Escherichia coli: comparative genomic analysis of E. coli commensal and pathogenic isolates. J Bacteriol 190, 6881–6893 (2008).
Treangen, T. J., Sommer, D. D., Angly, F. E., Koren, S. & Pop, M. Next generation sequence assembly with AMOS. Curr Protoc Bioinformatics Chapter 11, Unit 11 18 (2011).
von Mentzer, A. et al. Identification of enterotoxigenic Escherichia coli (ETEC) clades with long-term global distribution. Nature Genetics 46, 1321–1326 (2014).
Donnenberg, M. S. et al. Bacterial Factors Associated with Lethal Outcome of Enteropathogenic Escherichia coli Infection: Genomic Case-Control Studies. PLoS Negl Trop Dis 9, e0003791 (2015).
Hazen, T. H. et al. Genomic diversity of EPEC associated with clinical presentations of differing severity. Nature Microbiology 1, 15014, http://www.nature.com/articles/nmicrobiol201514-supplementary-information (2016).
Ingle, D. J. et al. Evolution of atypical enteropathogenic E. coli by repeated acquisition of LEE pathogenicity island variants. Nat Microbiol 1, 15010 (2016).
Sahl, J. W., Caporaso, J. G., Rasko, D. A. & Keim, P. The large-scale blast score ratio (LS-BSR) pipeline: a method to rapidly compare genetic content between bacterial genomes. PeerJ 2, e332 (2014).
Luo, Q. et al. Conservation and immunogenicity of novel antigens in diverse isolates of enterotoxigenic Escherichia coli. PLoS Negl Trop Dis 9, e0003446 (2015).
Munson, G. P., Holcomb, L. G., Alexander, H. L. & Scott, J. R. In vitro identification of Rns-regulated genes. J Bacteriol 184, 1196–1199 (2002).
Hazen, T. H. et al. Refining the pathovar paradigm via phylogenomics of the attaching and effacing Escherichia coli. Proc Natl Acad Sci USA 110, 12810–12815 (2013).
Sahl, J. W. et al. Defining the phylogenomics of Shigella species: a pathway to diagnostics. J Clin Microbiol 53, 951–960 (2015).
Del Canto, F. et al. Identification of Coli Surface Antigen 23, a novel adhesin of enterotoxigenic Escherichia coli. Infect Immun 80, 2791–2801 (2012).
DuPont, H. L. Systematic review: prevention of travellers’ diarrhoea. Aliment Pharmacol Ther 27, 741–751 (2008).
DuPont, H. L. Pathogenesis of traveler’s diarrhea. Chemotherapy 41(Suppl 1), 33–39 (1995).
Lindblom, G. B. et al. Campylobacter jejuni/coli and enterotoxigenic Escherichia coli (ETEC) in faeces from children and adults in Tanzania. Scand J Infect Dis 27, 589–593 (1995).
Bolin, I. et al. Enterotoxigenic Escherichia coli with STh and STp genotypes is associated with diarrhea both in children in areas of endemicity and in travelers. J Clin Microbiol 44, 3872–3877 (2006).
Lasaro, M. A. et al. Genetic diversity of heat-labile toxin expressed by enterotoxigenic Escherichia coli strains isolated from humans. J Bacteriol 190, 2400–2410 (2008).
Walker, R. I., Steele, D., Aguado, T. & Ad Hoc, E. T. E. C. Analysis of strategies to successfully vaccinate infants in developing countries against enterotoxigenic E. coli (ETEC) disease. Vaccine 25, 2545–2566 (2007).
Fleckenstein, J. M. & Rasko, D. A. Overcoming Enterotoxigenic Escherichia coli Pathogen Diversity: Translational Molecular Approaches to Inform Vaccine Design. Methods Mol Biol 1403, 363–383 (2016).
Vicente, A. C. et al. Outbreaks of cholera-like diarrhoea caused by enterotoxigenic Escherichia coli in the Brazilian Amazon Rainforest. Trans R Soc Trop Med Hyg 99, 669–674 (2005).
Finkelstein, R. A., Vasil, M. L., Jones, J. R., Anderson, R. A. & Barnard, T. Clinical cholera caused by enterotoxigenic Escherichia coli. J Clin Microbiol 3, 382–384 (1976).
Sjoling, A., Wiklund, G., Savarino, S. J., Cohen, D. I. & Svennerholm, A. M. Comparative analyses of phenotypic and genotypic methods for detection of enterotoxigenic Escherichia coli toxins and colonization factors. J Clin Microbiol 45, 3295–3301 (2007).
Sahl, J. W. et al. Examination of the enterotoxigenic Escherichia coli population structure during human infection. MBio 6, e00501–00515 (2015).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv.org (2013).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Bankevich, A. et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. Journal of Computational Biology 19, 455–477 (2012).
Sahl, J. W. et al. Genomic characterization of asymptomatic Escherichia coli isolated from the neobladder. Microbiology 157, 1088–1102 (2011).
Myers, E. W. et al. A whole-genome assembly of Drosophila. Science 287, 2196–2204 (2000).
Wirth, T. et al. Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol 60, 1136–1151 (2006).
Hayashi, K. et al. Highly accurate genome sequences of Escherichia coli K-12 strains MG1655 and W3110. Molecular systems biology 2, 2006 0007 (2006).
Delcher, A. L., Salzberg, S. L. & Phillippy, A. M. Using MUMmer to identify similar regions in large sequence sets. Curr Protoc Bioinformatics Chapter 10, Unit 10 13 (2003).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics (2014).
Leache, A. D., Banbury, B. L., Felsenstein, J., de Oca, A. N. & Stamatakis, A. Short Tree, Long Tree, Right Tree, Wrong Tree: New Acquisition Bias Corrections for Inferring SNP Phylogenies. Syst Biol (2015).
Sahl, J. W. et al. The Northern Arizona SNP Pipeline (NASP): accurate, flexible, and rapid identification of SNPs in WGS datasets. bioRxiv (2016).
Farris, J. S. THE RETENTION INDEX AND THE RESCALED CONSISTENCY INDEX. Cladistics 5, 417–419 (1989).
Schliep, K. P. phangorn: phylogenetic analysis in R. Bioinformatics 27, 592–593 (2011).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS One 5, e9490 (2010).
Wattam, A. R., Gabbard, J. L., Shukla, M. & Sobral, B. W. Comparative genomic analysis at the PATRIC, a bioinformatic resource center. Methods in molecular biology 1197, 287–308 (2014).
R C Team, R. C. T. R: A language and environment for statistical computing, http://www.R-project.org (2013).
Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25, 3389–3402 (1997).
Rasko, D. A., Myers, G. S. & Ravel, J. Visualization of comparative genomic analyses by BLAST score ratio. BMC Bioinformatics 6, 2 (2005).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL): an online tool for phylogenetic tree display and annotation. Bioinformatics 23, 127–128 (2007).
Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069 (2014).
Edgar, R. C. MUSCLE: a multiple sequence alignment with reduced time and space complexity. BMC Bioinformatics 5, 113–113 (2004).
Acknowledgements
The authors wish to thank Dr. Richard Finkelstein, and Dr. Ana Vicente for providing ETEC strains isolated from cases of severe diarrheal illness that were sequenced as part of this analysis. This project was supported in part by the icddr,b and federal funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health, Department of Health and Human Services under contract number HHSN272200900009C, NIH grant numbers 2R01AI089894, and 1U19AI090873; The Department of Veterans Affairs grant number 5I01BX001469-04; and Startup funds from the State of Maryland. Also, icddr,b is thankful to the Governments of Australia, Bangladesh, Canada, Sweden and the UK for providing core/unrestricted support.
Author information
Authors and Affiliations
Contributions
J.W.S., J.R.S., F.Q., J.M.F. and D.A.R. wrote the main manuscript text; N.I.B., Y.B., Q.L., A.S., F.Q. and J.M.F. identified and acquired samples; J.W.S., J.R.S., N.I.B., Y.B., Q.L., A.S. and D.A.R. generated data for figures; J.W.S., J.R.S., F.Q., J.M.F. and D.A.R. analyzed and interpreted data. All authors edited and reviewed the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sahl, J.W., Sistrunk, J.R., Baby, N.I. et al. Insights into enterotoxigenic Escherichia coli diversity in Bangladesh utilizing genomic epidemiology. Sci Rep 7, 3402 (2017). https://doi.org/10.1038/s41598-017-03631-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-03631-x
This article is cited by
-
The population genetics of pathogenic Escherichia coli
Nature Reviews Microbiology (2021)
-
Long-read-sequenced reference genomes of the seven major lineages of enterotoxigenic Escherichia coli (ETEC) circulating in modern time
Scientific Reports (2021)
-
Investigation of the Domestic Reservoirs of Diarrheagenic Escherichia coli in Diarrhea Case Households of Urban Bangladesh
Current Microbiology (2021)
-
Interrogation of a live-attenuated enterotoxigenic Escherichia coli vaccine highlights features unique to wild-type infection
npj Vaccines (2019)
-
Enterotoxigenic Escherichia coli Infections
Current Infectious Disease Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
