
Abstract
The common genetic variation of the serotonin transporter-linked polymorphic region (5-HTTLPR) has been related to depressive symptoms, in particular after stressful life events. Although it has been investigated in the past, results suggesting that the 5-HTTLPR genotype also affects hippocampal volume are often inconsistent and it remains unclear to what extent reduced hippocampal volume is influenced by the effect of stressful life events and 5-HTTLPR genotype. Moreover, sex, which is known to affect the prevalence of depression substantially, has not been taken into account when trying to disentangle the interactive effect of common genetic variation and environmental stressors on the hippocampus. We investigated this potentially relevant three-way interaction using an automatic magnetic resonance imaging (MRI)-based segmentation of the hippocampus in 357 healthy individuals. We determined the 5-HTTLPR genotype as a biallelic locus and childhood adversity (CA) using a standard questionnaire. An interaction for hippocampal volume was found between the factors sex, genotype, and severe CA (p=0.010) as well as an interaction between genotype and severe CA (p=0.007) in men only. Post hoc tests revealed that only male S’-allele carriers with severe CA had smaller hippocampi (p=0.002). Interestingly, there was no main effect of genotype in men, while female S’-allele carriers had smaller hippocampi than L’L’ carriers (p=0.023). Our results indicate that sex modulates the interactive effect of the 5-HTTLPR genotype and CA on hippocampal volume. While the S’-allele is associated with hippocampal volume independent of CA in women, men only have smaller hippocampi if they carry the risk allele and experienced severe CA.
Similar content being viewed by others


INTRODUCTION
Depression is a highly disabling and recurrent disease. Despite intensive research, optimal treatment is still lacking and hence, primary prevention of the disorder is a major goal. Therefore, recent research has focused on understanding vulnerability factors for depression investigating in particular the effect of common genetic variations and environmental factors on brain regions involved in the pathophysiology of depression. Understanding the effect of genetic variation in serotonin neurotransmission is one important focus of this line of research after an initial study has shown that short (S)-allele carriers of the serotonin transporter-linked polymorphic region (5-HTTLPR) exhibit more depressive symptoms, when they had also experienced stressful life events and childhood adversity (CA) (Caspi et al, 2003). After two meta-analyses failing to replicate this result (Munafò et al, 2009; Risch et al, 2009), a recent and more complete meta-analysis (Karg et al, 2011) has confirmed that 5-HTTLPR moderates indeed the relationship between stress and depression. Moreover, a strong association between the S-allele and increased stress sensitivity was found in particular when the authors included only those studies that used childhood maltreatment as their stress measure (Karg et al, 2011). Childhood maltreatment is known to cause HPA axis dysregulation and extensive neurobiological alterations (Heim and Nemeroff, 2001) and can therefore considered to be a form of severe CA.
It is important to identify brain regions that are sensitive to gene-by-environment interactions (Brown and Hariri, 2006). Here, the hippocampus has gained attention because reduced hippocampal volume is related to both childhood psychosocial stress (Tottenham and Sheridan, 2010) and depression (Sapolsky, 2000; Videbech and Ravnkilde, 2004). Moreover, small hippocampal volumes are associated with poor clinical outcome, a mechanism through which potential gene-by-environment interactions further impact on the course of the disease (MacQueen and Frodl, 2011). The hippocampus has a dense serotonergic innervation (Jacobs and Azmitia, 1992) so that common genetic variation of serotonergic neurotransmission could affect hippocampal morphology. Interestingly, developmental imaging studies using spectroscopy found a significantly lower hippocampal N-acetylaspartate concentration indexing altered neurogenesis, in S-allele as compared with homozygous L carriers (Gallinat et al, 2005).
While Frodl et al (2008) showed that healthy subjects homozygous for the S’-allele exhibit smaller hippocampi compared with L/L carriers, Canli et al (2006) only found evidence for a greater hippocampus in L/L carriers related to greater life stress (see also O’Hara et al, 2007; Cole et al, 2011; Dutt et al, 2009). Along the continuum to psychopathology, depressed patients carrying the S’-allele and a history for emotional childhood neglect had smaller hippocampal volumes compared with patients carrying the risk S’-allele without CA as well as compared with patients with CA, but homozygous for the non-risk allele (Frodl et al, 2010).
Besides the relatively small sample sizes of previous genetic neuroimaging studies investigating this interaction, gender differences may have potentially confounded the results. We hypothesize that sex has a modulatory effect on this gene-by-environment interaction on hippocampal volume. Sex steroids are potent modulators of brain development and the hippocampus contains sex steroid receptors (MacLusky et al, 1987). Moreover, there is evidence that stress hormone responses are different between men and women (Kudielka and Kirschbaum, 2005). The hippocampus is a core structure regulating stress hormone release and its morphology itself is also affected by stress, suggesting that sex-specific mechanisms could moderate the gene-by-environment interaction with respect to hippocampal volume (Lupien et al, 2009).
Most relevant with respect to understanding the vulnerability factors of depression, sexual dimorphism is not only present in the neural development of the hippocampus. Heim et al (2010) proposed that gene-by-environment interactions might occur in a sex-specific manner as an explanation for the higher prevalence of depression in women. Thus far, only studies using clinical outcome measures support the effect gender has on the association of 5-HTTLPR and stressful life events in the prevalence of depression (Brummett et al, 2008; Sjoberg et al, 2006; Eley et al, 2004), bypassing possible modulating actions of the brain.
To test the complex interaction between sex, genotype, and CA in affecting hippocampal volume, one needs a large sample of subjects, most optimally a cohort unaffected by consequences of disease and treatment. Investigating young, healthy subjects avoids potential effects of (chronic) disease and medication and thus, it allows a more direct, unconfounded investigation of potential correlates of vulnerability. Therefore, we set out to investigate 357 young, healthy subjects using automatic volumetry of the hippocampus based on 3 Tesla magnetic resonance imaging (MRI), genotyping, and a questionnaire on CA.
MATERIALS AND METHODS
Participants
A total of 357 (136 male and 221 female) subjects were included in this study. All participants gave written informed consent and the study was approved by the local ethics committee (CMO Region Arnhem-Nijmegen, The Netherlands). The sample consisted of healthy subjects of Caucasian descent, with a mean (SD) age of 24.3 (6.3) years for males and 23.3 (5.1) years for females. They were screened before participation in this study by self-report questionnaires: participants were excluded if they had a history of somatic disease potentially affecting the brain, current or past psychiatric or neurological disorder, medication (except hormonal contraceptives) or illicit drug use during the past 6 months, history of substance abuse, current or past alcohol dependence, pregnancy, lactation, menopause, or MRI contraindications.
Genotyping
Genetic analyses were carried out at the Department of Human Genetics of the Radboud University Nijmegen Medical Centre, in a laboratory that has a quality certification according to CCKL criteria. High molecular weight DNA was isolated from saliva using Oragene containers (DNA Genotek, Ottawa, Ontario, Canada) according to the protocol supplied by the manufacturer. The 5-HTTLPR genotype was the genotype at the single-nucleotide polymorphism (rs25531) in this region, with alleles designated as LG, LA, and S (Parsey et al, 2002). Because it is thought that the LG is comparable to the S-allele with regard to gene transcription and function, we reclassified the alleles on the basis of the resulting expression levels of the SLC6A4/5-HTT gene into L’L’, S’L’, and S’S’ genotypes (Wendland et al, 2006). Testing for Hardy–Weinberg equilibrium did not show deviation from the expected distribution (p>0.05).
MRI Acquisition
Anatomical T1-weighted MRI data were acquired at the Donders Centre for Cognitive Neuroimaging. All scans covered the entire brain and had a voxel size of 1 × 1 × 1 mm3.
To make use of the best possible signal to noise ratio, all images were acquired at 3 Tesla Siemens Trio or TimTrio scanners (Siemens, Erlangen, Germany), using small variations to a standard T1-weighted 3D MPRAGE sequence (TR 2300 ms, TI 1100 ms, TE 3.93 ms, 192 sagittal slices, field of view 256 mm). These variations included TR/TI/TE/slices of 2300/1100/3.03/192, 2300/1100/2.92/192, 2300/1100/2.96/192, 2300/1100/2.99/192, 1940/1100/3.93/176, and 1960/1100/4.58/176, and the use of GRAPPA parallel imaging with an acceleration factor of 2.
Image Data Processing
Whole brain segmentation of gray matter, white matter, and cerebrospinal fluid was performed using the VBM 5.1 toolbox version 1.19 (dbm.neuro.uni-jena.de/vbm/) in SPM5 (http://www.fil.ion.ucl.ac.uk/spm/) using priors (default settings). Total volume of gray matter and white matter was calculated by adding the resulting tissue probabilities. Total brain volume (TBV) was defined as the sum of white matter and gray matter volume. For the automatic segmentation of the hippocampus, we used FIRST v1.2 (http://www.fmrib.ox.ac.uk/fsl/first/index.html) in FSL 4.1.4 (http://www.fmrib.ox.ac.uk/fsl; Patenaude, 2007). This method is based on Bayesian statistical models of shape and appearance for 17 structures from 317 manually labeled T1-weighted MR images. To fit the models, the probability of the shape given the observed intensities is used (Patenaude, 2007). In addition, to model intensity at the structural boundary, automatic boundary correction was applied. After segmentation, the volume of the left and right hippocampus was calculated by multiplying the number of voxels in a specific structure with the voxel volume (1 mm3). To detect obvious segmentation errors (like brain structures located outside the brain), visual inspection of the segmented structures projected onto the T1-weighted MRI scans was done using the software MRIcroN Version Beta 7 (http://www.mricro.com/mricron). Total hippocampal volume was defined as the sum of left and right hippocampal volumes.
Assessment of CA
CA was assessed using an adapted version of the List of Threatening Life Events developed by Brugha et al (1985) with the aim of assessing the stress of life events. This inventory encompasses life events that are likely to occur relatively frequently and score relatively high on long-term threat. Participants were asked whether they had experienced a list of predefined events before the age of 16 years. This list comprised events like death of a parent, severe illness of self or spouse, moving house and abuse. Presence of CA was defined as having experienced at least one CA before the age of 16 years.
Negative Mood
The Positive Affect Negative Affect Schedule (PANAS) consists of two 10-item scales designed to provide brief measures of positive and negative affect (Watson et al, 1988). The two scales index two distinct, but moderately negatively correlated, factors. PANAS was used as a covariate in our analysis to correct for possible recollection bias.
Recollection bias is not only found in depressed subjects, but also in never-depressed individuals that are for example neurotic (Chan et al, 2007) or family members of MDD patients (Jaenicke et al, 1987). In the light of these findings, it is therefore possible that those of our healthy subjects with more risk factors for depression are also more likely to report negative life events due to recollection bias, resulting in an overestimation of the effect of life events when not correcting for PANAS scores.
Statistical Analysis
Statistical analysis was performed using PASW Statistics 18. Significance level was p=0.05. An interaction effect of sex, genotype, and CA was tested by adding a product term (sex*5-HTTLPR*CA) to the fully adjusted models. When significant, these interaction effects would be further explored using two-way interaction models and simple post hoc T-tests with Bonferroni correction. With Bonferroni correction, our post hoc significance level will be p=0.05/(2 × 2 × 2)=0.006. Covariates in all analyses were age, level of education, MRI protocol, and negative mood state measured with PANAS. Moreover, we used TBV as a covariate to control for gender-specific differences in brain size that could affect the outcome of our specific volumes.
The main effect of 5-HTTLPR on hippocampal volumes was examined by conducting analyses of covariance (ANCOVA). The main effect of CA on hippocampus was examined in an equivalent analysis. Individual differences between the different genotypes were explored using simple post hoc T-tests. When comparing 5-HTTLPR genotypes, we also compared S’/S’ and S’/L’ together to L’/L’ (thus S’-allele carriers vs non-S’-allele carriers, respectively) to investigate whether potential effects are more generally related to the presence of the S’-allele.
RESULTS
There were no significant differences in covariates between the two sexes, except for TBV (larger in males, p<0. 001) and educational level (higher in females, p=0.037) (Table 1). Note that all analyses were stratified for these aspects.
To make sure that the variation in MRI protocols did not systematically influence our automatic segmentation of the hippocampus, we conducted an ANOVA with MRI protocol as independent and total hippocampus volume as dependent variable. This analysis showed no significant effect of MRI protocol on hippocampus volume (F(16 340)=20.4, p=0.204). Moreover, the different MRI protocols were equally distributed between the genotypes (χ2=30.5, df=32, p=0.542), CA groups (χ2=11.9, df=16, p=0.750), and sexes (χ2=23.7, df=16, p=0.096).
Three-Way Interaction between CA, Genotype, and Sex
We did not find an interaction effect of CA with genotype and sex. Since a previous meta-analysis suggests a particularly strong association between the S’-allele and more severe CA like maltreatment (Karg et al, 2011), we selected the following items from the List of Threatening Life Events: death of a close relative or friend and victim of abuse, war or disaster (items 3, 6, 7, 8, 9, 17, and 18; see Table 2). These events were marked as severe CA. The selection of these specific events was motivated by evidence that death of a loved one and any violent event or sexual trauma in childhood is highly associated with subsequent PTSD symptoms (Breslau et al, 1999; Copeland et al, 2007). Most importantly, with respect to our research question we found that there was a three-way interaction between severe CA, genotype, and sex on total hippocampus volume (p=0.01), even when not corrected for the PANAS (p=0.016).
Two-Way Interaction between Severe CA and Genotype
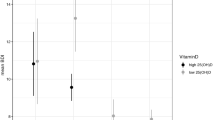
When further exploring this interaction by computing a two-way analysis with the factors genotype × severe CA for males and females, separately, we found a significant interaction in males (p=0.007/p=0.01), but not in females (p=0.638/p=0.740). When reanalyzing these data for S’-allele carriers vs L’/L’ carriers, the three-way interaction sex × genotype × severe CA (p=0.045/p=0.062) and the two-way interactions genotype × severe CA in males (p=0.003/p=0.005) and in females (p=0.344/p=0.299) confirmed our results (Table 2), indicating that we identified an effect that is more generally related to the presence of the S’-allele. Finally, post hoc T-tests performed in the male subgroup only revealed that severe CA is associated with smaller hippocampi in male S’-allele carriers (p=0.002/p=0.002, also significant after Bonferroni correction), but not in males with the L’/L’ genotype (p=0.600/p=0.686) (Figure 1). We also followed up the two-way interaction between genotype and severe CA by separately investigating the effects of severe CA and genotype on hippocampal volume. We found no main effect of either of these factors in men (maximum p=0.995/p=0.991). While females also showed no main effect of severe CA (p=0.611/p=0.579), female L’/L’ carriers showed larger hippocampi compared with S’-allele carriers (p=0.023/p=0.024) (Figure 2). This last effect, however, is not significant after Bonferonni correction (p>0.006) (Table 3).

Differences in the effect of severe childhood adversity (CA) on total hippocampus volumes between subjects with different genotypes. Results are indicated per sex and corrected for age, level of education, TBV (total brain volume), MRI protocol, and PANAS score. This graph shows a significant negative effect of a history of severe CA in male S’-allele carriers only. Error bars represent standard errors. **Indicates p<0.01.

Differences in total hippocampus volumes of S’-allele compared with L’/L’ carriers per sex when corrected for age, level of education, TBV (total brain volume), MRI protocol, and PANAS score. This graph shows that female S’-allele carriers have a significantly smaller hippocampal volume than female L’/L’ carriers. Error bars represent standard errors. *Indicates p<0.05.
DISCUSSION
In this study, we show for the first time that sex modulates the interactive effect of the 5-HTTLPR genotype and CA on hippocampal volume. We found a significant three-way interaction, which was driven by the fact that only male S’-allele carriers exhibited an association between severe CA and smaller hippocampal volume (Figure 1). Contrary, in females differences in hippocampal volume were only found as a function of genotype (independent of CA) whereby S’-allele carriers showed smaller hippocampal volumes. Interestingly, we neither found a main effect of genotype nor CA in men. Taken together, our results imply a sex-specific dissociation in genetic and environmental effects on hippocampal volume in a healthy sample that is suggestive of a sex-specific vulnerability to develop depression. A smaller hippocampus that reflects a risk factor to develop depression (Amico et al, 2011) can already be found in females with the more pathogenic allele of 5-HTTLPR, whereas in males it may be only present if both genetic and environmental risk factors co-occur.
Another reason for inconsistencies with previous studies could be found in the use of different segmentation methods for the hippocampus. Many different segmentation protocols for the hippocampus exist and discrepancies are known to occur, most frequently at specific anatomical boundaries such as the anterior hippocampal-amygdala border (Konrad et al, 2009). The deformable model of FSL FIRST has been shown to give accurate and robust results for the segmentation of 15 subcortical structures. For the hippocampus, the median quantitative Dice overlap measures are in the range of 0.8–0.844, which is comparable to or better than other automated methods (Patenaude et al, 2011).
Our results support the data of Frodl et al (2008) showing that in healthy subjects the S’-allele is the risk allele for smaller hippocampal volumes. In the light of missing additional evidence on the hippocampus, in particular Selvaraj et al (2011) support a reduction of grey matter in healthy S’-allele carriers in different brain regions. Moreover, our findings support the meta-analysis of Karg et al (2011) in that we only found an effect of CA when severe CA was entered in the analysis, suggesting that at least in healthy male S’-allele carriers there was no strong impact of milder stressors on hippocampal volume.
The modulatory effect of sex on the interaction between 5-HTTLPR and environmental stressors has thus far only been investigated in clinical studies on depressive symptoms that are not directly comparable to our results. In fact, at first sight those data seem counterintuitive to the data presented here. For example, Eley et al (2004) showed that an increase in the number of S-alleles increased the likelihood of being in the high depression group only for female adolescents within a high environmental risk group, suggesting that the interactive effect may also play a role for females. Sjoberg et al (2006) and Brummett et al (2008) found also that in women the S-allele, combined with a stressor, was associated with higher depression scores. In males, however, it was the L-allele, combined with a stressor, that was related to more depressive symptoms. It should be noted that all three studies did not reclassify the LG-allele as S’-allele, which could lead to an unbalanced underestimation of the effect per sex (eg, an underestimation of S’-allele effects in males only if the LG-allele would have been more prevalent in males than in females). In addition, two of the three studies investigated adolescents. In adolescence, however, neural sex differences show dynamic changes during development, with a 1–2 years earlier peak in cortical and subcortical gray matter trajectories in females than in males (Lenroot and Giedd, 2010). More specifically, the hippocampus, containing sex steroid receptors, also shows sexual dimorphism in growth rate during this period (MacLusky et al, 1987; Suzuki et al, 2005). Thus, evaluating sex differences in this age can lead to different results than those found in an adult population. Moreover, the interaction between 5-HTTLPR and environmental stressors during development may be related to different psychopathology in the two sexes. The polymorphism has not only been associated with depressive symptoms, but also with other behavioral outcome measures such as social cognition and behavior (Canli and Lesch, 2007). In conclusion, it is difficult to compare the effect of the interaction between 5-HTTLPR and environmental stressors on clinical outcome measures with those on brain correlates that may represent a neurobiological vulnerability in healthy subjects.
The interaction of sex with CA, possibly due to the neuroprotective effects of estrogen (Sherwin, 1998), may be an important bias when investigating genotype effects on the hippocampus. Estrogen positively modulates serotonergic effects of the brain (Kugaya et al, 2003) and may act as a ‘brake’ on the HPA axis (Young and Korszun, 2010). In the light of these findings, it is not surprising that salivary cortisol increases in response to stress in men are up to twice as high as in women (Kudielka et al, 2009). Hence, neurotoxic effects of cortisol, caused by childhood stress, could have a larger impact in males than in females before menopause (Sapolsky, 2000). This has already been demonstrated in children with PTSD, where boys show more adverse effects of childhood maltreatment on brain development than girls (De Bellis and Keshavan, 2003). Estrogen can also directly influence hippocampal development by blocking the neurodegenerative effects of glucocorticoids (Pruessner et al, 2010).
Given that the S-allele seems to be associated with an impaired serotonin reuptake activity in the brain (Heils et al, 1996), this may also dampen the modulatory effects of estrogen. Such a direct association between female-specific effects of the S-allele and brain structure is also supported by the main effect of the S-allele on hippocampal volume in females in our data.
A major strength of our study is the large sample size of healthy men and women. One limitation is the use of questionnaires in the assessment of CA. However, by making the distinction between CA and severe CA, we expect to have created a more specific assessment of CA. Finally, by using the PANAS as a covariate in our analysis, we have corrected for recollection bias, in which negative mood would influence memory by making it more sensitive to recall negative events.
A clear limitation is that we could not replicate our data in an independent sample, which should be the main goal of future studies. It is possible that we underestimate the interactive effect of the aforementioned factors because we may have excluded those subjects who were more affected by CA and developed neuropsychiatric disorders with hippocampal damage, such as PTSD (Karl et al, 2006). In addition, it is possible that our population is too young to show the full extent of CA effects on the hippocampus and that the effects may have been even larger when using an older population (Tottenham and Sheridan, 2010). Future studies should address these issues ideally comparing subjects of different age groups throughout adulthood. Finally, it is important to mention that our study population does not seem to suffer from less life events than the general population (Cuijpers et al, 2011), but at the same time does not suffer from mental health problems due to the exclusion criteria applied. This implies that our sample consists of subjects belonging to the more resilient part of the population and therefore adequately reflects the healthy segment of a putative health-disease continuum.
In summary, our results show that sex significantly modulates the effects 5-HTTLPR genotype and childhood events have on hippocampal morphology. While in women the S’-allele alone is sufficient to reduce hippocampal volume, in men a history of severe CA modulates the effect within S’-allele carriers. Even though these results require replication, our findings contribute to the understanding of sex differences in the pathophysiology of depression and indicate a mechanistic account of how a specific risk allele and adverse events may increase the vulnerability for depression. Future studies aiming to investigate the effects of the serotonin transporter-linked polymorphic region should consider the sex-specific nature of this gene-by-environment interaction, thereby reducing the chance of false negative results.
References
Amico F, Meisenzahl E, Koutsouleris N, Reiser M, Möller HJ, Frodl T (2011). Structural MRI correlates for vulnerability and resilience to major depressive disorder. J Psychiatry Neurosci 36: 15–22.
Breslau N, Chilcoat HD, Kessler RC, Davis GC (1999). Previous exposure to trauma and PTSD effects of subsequent trauma: results from the Detroit Area Survey of Trauma. Am J Psychiatry 156: 902–907.
Brown SM, Hariri AR (2006). Neuroimaging studies of serotonin gene polymorphisms: exploring the interplay of genes, brain, and behavior. Cogn Affect Behav Neurosci 6: 44–52.
Brugha T, Bebbington P, Tennant C, Hurry J (1985). The list of threatening experiences: a subset of 12 life event categories with considerable long-term contextual threat. Psychol Med 15: 189–194.
Brummett BH, Boyle SH, Siegler IC, Kuhn CM, Ashley-Koch A, Jonassaint CR et al. (2008). Effects of environmental stress and gender on associations among symptoms of depression and the serotonin transporter gene linked polymorphic region (5-HTTLPR). Behav Genet 38: 34–43.
Canli T, Lesch KP (2007). Long story short: the serotonin transporter in emotion regulation and social cognition. Nat Neurosci 10: 1103–1109.
Canli T, Qiu M, Omura K, Congdon E, Haas BW, Amin Z et al. (2006). Neural correlates of epigenesis. Proc Natl Acad Sci USA 103: 16033–16038.
Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H et al. (2003). Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301: 386–389.
Chan SW, Goodwin GM, Harmer CJ (2007). Highly neurotic never-depressed students have negative biases in information processing. Psychol Med 37: 1281–1291.
Cole J, Weinberger DR, Mattay VS, Cheng X, Toga AW, Thompson PM et al. (2011). No effect of 5HTTLPR or BDNF Val66Met polymorphism on hippocampal morphology in major depression. Genes Brain Behav 10: 756–764.
Copeland WE, Keeler G, Angold A, Costello EJ (2007). Traumatic events and posttraumatic stress in childhood. Arch Gen Psychiatry 64: 577–584.
Cuijpers P, Smit F, Unger F, Stikkelbroek Y, Ten Have M, de Graaf R (2011). The disease burden of childhood adversities in adults: a population-based study. Child Abuse Negl 35: 937–945.
De Bellis MD, Keshavan MS (2003). Sex differences in brain maturation in maltreatment-related pediatric posttraumatic stress disorder. Neurosci Biobehav Rev 27: 103–117.
Dutt A, McDonald C, Dempster E, Prata D, Shaikh M, Williams I et al. (2009). The effect of COMT, BDNF, 5-HTT, NRG1 and DTNBP1 genes on hippocampal and lateral ventricular volume in psychosis. Psychol Med 39: 1783–1797.
Eley TC, Sugden K, Corsico A, Gregory AM, Sham P, McGuffin P et al. (2004). Gene-environment interaction analysis of serotonin system markers with adolescent depression. Mol Psychiatry 9: 908–915.
Frodl T, Koutsouleris N, Bottlender R, Born C, Jäger M, Mörgenthaler M et al. (2008). Reduced gray matter brain volumes are associated with variants of the serotonin transporter gene in major depression. Mol Psychiatry 13: 1093–1101.
Frodl T, Reinhold E, Koutsouleris N, Donohoe G, Bondy B, Reiser M et al. (2010). Childhood stress, serotonin transporter gene and brain structures in major depression. Neuropsychopharmacology 35: 1383–1390.
Gallinat J, Ströhle A, Lang UE, Bajbouj M, Kalus P, Montag C et al. (2005). Association of human hippocampal neurochemistry, serotonin transporter genetic variation, and anxiety. Neuroimage 15: 123–131.
Heils A, Teufel A, Petri S, Stöber G, Riederer P, Bengel D et al. (1996). Allelic variation of human serotonin transporter gene expression. J Neurochem 66: 2621–2624.
Heim C, Nemeroff CB (2001). The role of childhood trauma in the neurobiology of mood and anxiety disorders: preclinical and clinical studies. Biol Psychiatry 49: 1023–1039.
Heim C, Shugart M, Craighead WE, Nemeroff CB (2010). Neurobiological and psychiatric consequences of child abuse and neglect. Dev Psychobiol 52: 671–690.
Jacobs BL, Azmitia EC (1992). Structure and function of the brain serotonin system. Physiol Rev 72: 165–229.
Jaenicke C, Hammen C, Zupan B, Hiroto D, Gordon D, Adrian C et al. (1987). Cognitive vulnerability in children at risk for depression. J Abnorm Child Psychol 15: 559–572.
Karg K, Burmeister M, Shedden K, Sen S (2011). The serotonin transporter promoter variant (5-HTTLPR), stress, and depression meta-analysis revisited. Evidence of genetic moderation. Arch Gen Psychiatry 68: 444–454.
Karl A, Schaefer M, Malta LS, Dörfel D, Rohleder N, Werner A (2006). A meta-analysis of structural brain abnormalities in PTSD. Neurosci Biobehav Rev 30: 1004–1031.
Konrad C, Ukas T, Nebel C, Arolt V, Toga AW, Narr KL (2009). Defining the human hippocampus in cerebral magnetic resonance images--an overview of current segmentation protocols. Neuroimage 47: 1185–1195.
Kudielka BM, Hellhammer DH, Wüst S (2009). Why do we respond so differently? Reviewing determinants of human salivary cortisol responses to challenge. Psychoneuroendocrinology 34: 2–18.
Kudielka BM, Kirschbaum C (2005). Sex differences in HPA axis responses to stress: a review. Biol Psychol 69: 113–132.
Kugaya A, Epperson CN, Zoghbi S, van Dyck CH, Hou Y, Fujita M et al. (2003). Increase in prefrontal cortex serotonin 2A receptors following estrogen treatment in postmenopausal women. Am J Psychiatry 160: 1522–1524.
Lenroot RK, Giedd JN (2010). Sex differences in the adolescent brain. Brain Cogn 72: 46–55.
Lupien SJ, McEwen BS, Gunnar MR, Heim C (2009). Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 10: 434–445.
MacLusky NJ, Clark AS, Naftolin F, Goldman-Rakic PS (1987). Estrogen formation in the mammalian brain: possible role of aromatase in sexual differentiation of the hippocampus and neocortex. Steroids 50: 459–474.
MacQueen G, Frodl T (2011). The hippocampus in major depression: evidence for the convergence of the bench and bedside in psychiatric research? Mol Psychiatry 16: 252–264.
Munafò MR, Durrant C, Lewis G, Flint J (2009). Gene X environment interactions at the serotonin transporter locus. Biol Psychiatry 65: 211–219.
O’Hara R, Schröder CM, Mahadevan R, Schatzberg AF, Lindley S, Fox S et al. (2007). Serotonin transporter polymorphism, memory, and hippocampal volume in the elderly: association and interaction with cortisol. Mol Psychiatry 12: 544–555.
Parsey RV, Oquendo MA, Simpson NR, Ogden RT, Van Heertum R, Arango V et al. (2002). Effects of sex, age, and aggressive traits in man on brain serotonin 5-HT1A receptor binding potential measured by PET using [C-11] WAY-100635. Brain Res 954: 173–182.
Patenaude B (2007). Bayesian statistical models of shape and appearance for subcortical brain segmentation. DPhil thesis, University of Oxford.
Patenaude B, Smith SM, Kennedy DN, Jenkinson M (2011). A Bayesian model of shape and appearance for subcortical brain segmentation. Neuroimage 56: 907–922.
Pruessner JC, Dedovic K, Pruessner M, Lord C, Buss C, Collins L et al. (2010). Stress regulation in the central nervous system: evidence from structural and functional neuroimaging studies in human populations-2008 Curt Richter Award Winner. Psychoneuroendocrinology 35: 179–191.
Risch N, Herrell R, Lehner T, Liang KY, Eaves L, Hoh J et al. (2009). Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: a meta-analysis. JAMA 301: 2462–2471.
Sapolsky RM (2000). Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch Gen Psychiatry 57: 925–935.
Selvaraj S, Godlewska BR, Norbury R, Bose S, Turkheimer F, Stokes P et al. (2011). Decreased regional gray matter volume in S′ allele carriers of the 5-HTTLPR triallelic polymorphism. Mol Psychiatry 16: 471, 472-3.
Sherwin BB (1998). Estrogen and cognitive functioning in women. Proc Soc Exp Biol Med 217: 17–22.
Sjoberg RL, Nilsson KW, Nordquist N, Ohrvik J, Leppert J, Lindstrom L et al (2006). Development of depression: sex and the interaction between environment and a promoter polymorphism of the serotonin transporter gene. Int J Neuropsychopharmacol 9: 443–449.
Suzuki M, Hagino H, Nohara S, Zhou SY, Kawasaki Y, Takahashi T et al. (2005). Male-specific volume expansion of the human hippocampus during adolescence. Cereb Cortex 15: 187–193.
Tottenham N, Sheridan MA (2010). A review of adversity, the amygdala and the hippocampus: a consideration of developmental timing. Front Hum Neurosci 8: 68.
Videbech P, Ravnkilde B (2004). Hippocampal volume and depression: a meta-analysis of MRI studies. Am J Psychiatry 161: 1957–1966.
Watson D, Clark LA, Tellegen A (1988). Development and validation of brief measures of positive and negative affect: the PANAS scales. J Pers Soc Psychol 54: 1063–1070.
Wendland JR, Martin BJ, Kruse MR, Lesch KP, Murphy DL (2006). Simultaneous genotyping of four functional loci of human SLC6A4, with a reappraisal of 5-HTTLPR and rs25531. Mol Psychiatry 11: 224–226.
Young E, Korszun A (2010). Sex, trauma, stress hormones and depression. Mol Psychiatry 15: 23–28.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Dr Thomas Frodl received speaker′s fees from Shire and Lilly. All other authors declare no conflict of interest.
PowerPoint slides
Rights and permissions
About this article
Cite this article
Everaerd, D., Gerritsen, L., Rijpkema, M. et al. Sex Modulates the Interactive Effect of the Serotonin Transporter Gene Polymorphism and Childhood Adversity on Hippocampal Volume. Neuropsychopharmacol 37, 1848–1855 (2012). https://doi.org/10.1038/npp.2012.32
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2012.32
Keywords
This article is cited by
-
Additional Insights into the Relationship Between Brain Network Architecture and Susceptibility and Resilience to the Psychiatric Sequelae of Childhood Maltreatment
Adversity and Resilience Science (2020)
-
Sex differences in hippocampal cognition and neurogenesis
Neuropsychopharmacology (2019)
-
Animal models of PTSD: a challenge to be met
Molecular Psychiatry (2019)
-
The effects of childhood maltreatment on brain structure, function and connectivity
Nature Reviews Neuroscience (2016)
-
Overexpression of Forebrain CRH During Early Life Increases Trauma Susceptibility in Adulthood
Neuropsychopharmacology (2016)
