
Abstract
The floristic composition, diversity, and conservation status of woody plant species in Ethiopia are determined in this study. Data on vegetation are collected using a systematic random sampling method from 45 20 m × 20 m (400m2) plots laid every 100 m along four parallel transect lines running from south to north. The Flora of Ethiopia and Eritrea identification key (Volume I–VIII) is used to identify the scientific names of all woody plant species. The Shannon–Wiener diversity index is used to calculate species diversity, whereas Sorensen's similarity coefficient is used to assess similarities and differences among plant communities. The studied forest contains 44 plant species from 29 families and 40 genera. The Fabaceae is the most dominant family, followed by the Rubiaceae and Combretaceae. In terms of habit, the collected woody species consisted of 30 (68.2%) trees and 14 (31.8%) shrubs. The two most frequent species are Syzygium guineense (51.11%) and Maytenus gracilipes (46.66%). Three plant community types are identified using Agglomerative Hierarchical Cluster Analysis. Most species (Calpurnia aurea-Combretum molle) are observed in community type two. The study is generally limited to the floristic and diversity of woody plant species. Therefore, a thorough investigation of all species' floristic composition, population structure, regeneration status, and ecological aspects is recommended.
Article Highlights
-
The study is focused on the Floristic Composition and Diversity of Woody Plant Species in the Hereje Natural Forest.
-
The forest contains forty-four woody species classified into 28 families with the dominant families being Fabaceae, Rubiaceae, and Combretaceae.
-
The Agglomerative Hierarchical Cluster Analysis shows three types of plant community types, with community type two containing the greatest number of species.
Similar content being viewed by others

1 Introduction
Human-caused activities are widely acknowledged as the leading cause of forest damage in many tropical countries [1]. Conversion of forest land to agricultural land, land degradation, habitat destruction and fragmentation, poaching, forest fires, invasive species, wetland over-harvesting, and illegitimate trafficking of wild and domestic animals, destruction and, climate change are the major threats to Ethiopian biodiversity [2].
Ethiopia has a diverse ecosystem, ranging from humid forests and extensive wetlands in the west and south west to the desert of Afar depression in the northeast [3]. Because of its tremendous geological history, broad latitudinal spread, and vast altitudinal range, it is known as a biodiversity hotspot and center of origin and diversity of plants and animals [4, 5]. Ethiopia owns 59.7 million ha of woody vegetation, of which 3.56% are high forest (about 4.07 million ha), 49% woodland (29.24 million ha), 44.2% shrub land or bush land (26.4 million ha), and 955,705 ha are plantations [6]. The country's flora contains approximately 6000–7000 species of higher plants, of which approximately 780–840 (12–13%) are endemic [7].
Despite being recognized as a center of floral and faunal species diversity and endemism, Ethiopia's forest cover declined from 15.11 million hectares in 1990 to 12.9 million hectares in 2010, with 18.66% of the forest cover deforested in those 20 years [8]. Furthermore, deforestation continues at an alarming rate, with nearly 141,000 ha (0.93%) of forest cover destroyed each year [5, 9]. This led to biodiversity loss, drought, ecological imbalance, and environmental degradation [8, 9]. Several factors have been identified as the causes of forest reduction in Ethiopia, including a growing population with increased demand for farmland and wood for construction and fuel, resettlement programs, an unstable land-tenure system, property rights over forest, migration, political unrest, poverty, forest fires, inappropriate conservation methods, and lack of awareness [10]. Climate change also affected the frequency and severity of forest disturbances by increasing the severity of wildfires, storms, insect outbreaks, and the occurrence of invasive species [11]. It also contributed to exacerbating the negative trend in water quality and forest coverage by interfering with the frequency and regime of aquifer recharge associated with land use, soil characteristics, and rainfall [12].
The majority of Ethiopia's remaining forests are concentrated in the country's south and south-western regions, where they are less accessible and/or populated [5]. A significant proportion of these forests exist in fragmented patches and represent dry evergreen Afromontane forest, grassland complex, and riverine vegetation, but they are disappearing at an alarming rate as agricultural activities expand [5, 13]. Deforestation patterns will soon decimate the remaining forests unless appropriate interventions are designed [9].
Forest resource data is a prerequisite for forest management and planning in the context of sustainable development [14]. Floristic composition, species diversity, and community analysis, in particular, are critical for providing needed information on forest species richness and diversity, and knowledge of vegetation types is useful for forest management, sustainable utilisation, and understanding their ecology [13]. Knowledge of forest floristic composition and structure is also beneficial in identifying ecologically and economically important plants and their diversity, as well as protecting threatened and economically important plant species [5, 15]. A study of the Ethiopian forest's fragmented patches is thus required to ensure the long-term use of the vegetation, ecological management, and conservation practices [1].
Several researchers have investigated the vegetation coverage and species composition of southern Ethiopia [5, 7, 9, 16]. Despite these studies, the area is still facing significant challenges in conserving its vegetation and is becoming one of Ethiopia's most threatened ecosystems. Furthermore, due to the Southern part's unique physical setting, climatic conditions, and biogeographical position, the area received little attention from researchers compared to other corners of Ethiopian vegetation [16]. Thus, it would be critical to conduct additional floristic studies for each patch of the Southern forest, particularly in areas that had not previously been studied [4].
Hereje Forest is one of these priority areas with potential remnant vegetation in the Wolaita Zone. The forest is thought to be one of the few tropical forest forests in the country, with forest land covering the majority of the forest. However, information on the status of woody species, floristic composition and community types in some of the remnant patches in general and the Hereje Natural Forest in particular, are still lacking. Hence, this study aimed to assess the woody floristic composition, species diversity, and community types as a baseline for assessing the nature and status of plant life in the study area, which is critical for future management of the forest.
The next section (Sect. 2) details the materials and methods of the study. In Sect. 3, the findings of the study are presented along with associated figures and tables. Furthermore, the discussions are presented in the same section, with an emphasis on the observations and potential interpretations. Finally, Sect. 4 reiterates the conclusion and future work.
2 Materials and methods
2.1 Description of study area
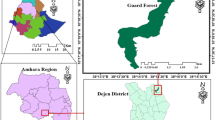
The study was conducted in Boloso Bombe, which is located in the Wolaita zone of Ethiopia's Southern Nations, Nationalities, and Peoples National Regional State (Fig. 1). The area is located along Ajora Falls in the Wolaita Zone, 325 km from Addis Abeba and 55 km from Wolaita Sodo town on the way to the Hossana exit [17]. The neighboring districts are Kambata Tambaro in the North, Boloso Sore District in the East, Sodo Zuria and Kindo Koysha District in the South, and Damot Sore District in the West.

Map of the Boloso Bombe district and Hereje forest, Southern Ethiopia
The study area is located at 37°33′0″ to 37°36′0″E longitude and 7°43′30″ to 7°73′30″N latitude, with elevations ranging from 1542 to 2375 m.a.s.l. There are three agro-climatic regions in the study area: the midland (45%), the lowland (30%), and the highland (25%). The area has two rainy seasons, with an average annual rainfall of 1427 mm, with the main rainy season lasting from July to September. The study area has a minimum temperature of 13.6 °C and a maximum temperature of 28.8 °C, with an annual average temperature of 21.2 °C (EMSA 2019). The soil in the area is mostly made up of Vertisol and Nitisol.
A subsistence mixed farming system that includes crop and livestock production is a common economic activity in the Boloso Bombe District. Because of the presence of various agro-ecologic regions, various crops such as cereals, legumes, vegetables, root crops, perennials, fruits, and so on are widely cultivated in the area. Teff, maize, coffee; root crops such as sweet potato, yam, and taro; haricot bean, enset, and fruits are among the most commonly produced crops in the area (Boloso Bombe Woreda District Agricultural and Rural Development Office, 2019 unpublished).
2.2 Sampling design
A cross-sectional sampling design with qualitative and quantitative data collection methods was used in this study. The vegetation data for woody species were collected using systematic random sampling techniques with random starting points [18]. In the sampling sites, four transect lines were laid at 200 m intervals from south to north using a compass. Along each transect, sample plots measuring 20 m × 20 m were taken at 100 m intervals. Data on woody vegetation were collected from a total of 45 plots from April 2019 to May 2019.
2.3 Data collection
All woody plant species encountered in each of the established 20 m × 20 m plots were recorded using their local names in the Wolaitigna language, pressed, and dried using standard herbarium procedures. To obtain the population structure, all tree/shrub species with a Diameter at Breast Height (DBH) greater than 2.5 cm were measured.
The scientific names of the species were determined with the help of experts and the Flora of Ethiopia and Eritrea identification key (Volume I–VIII). All woody species in the study forest were counted to analyze the floristic composition of the forest. The abundance data for all woody species were visually estimated and later converted to the modified 1–9 Braun-Blanquette scale [19].
2.4 Data analysis
2.4.1 Plant community classification
The classification of vegetation is an extremely useful tool for data reduction and synthesis, interpretation, effective communication, and land management and planning. The synoptic abundance cover value determined the dominant plant species in the community [5, 14]. Each community type was named after two or three dominant species in the group [20]. The plant community types in the study forest were identified using agglomerative hierarchical cluster analysis in SPSS (version 20.0). To determine the types of communities, the species cover-abundance data were used in Agglomerative hierarchical cluster analysis. According to the findings, vegetation samples with similar woody species compositions were grouped. The identified plant community types were given names based on the relative magnitude of their mean cover-abundance values, which were chosen from a list of two or three dominant species [20].
2.4.2 Plant diversity analysis
The Shannon–Wiener Diversity Index (H′) [21] was used to analyze species diversity, whereas the Shannon Evenness Index (J) [22] was used to calculate species Evenness (a measure of species balance). The H′ and J indices were calculated using the species' mean cover-abundance value [23].
The H′ for species diversity and evenness was calculated as follows:
where H′ = Shannon diversity index, Pi = proportion of individuals in the ith species, ln Pi = log proportion of individual species.
where J = Shannon-Weiner evenness, H′ = Shannon–Wiener diversity index and Hmax = is the maximum Shannon-Weiner diversity index, lnS = the natural logarithm of the total number of species and S = total number of species in the sample.
To compare similarity among plant community types, Sorensen's similarity coefficient (SC) was used. This was used to assess the composition of woody species (tree or shrub) and species distribution within plant communities [24]. SC was calculated as:
where Sc = Sorensen’s similarity coefficient, a = number of species common to both communities, b = number of species present in the first community only, and c = number of species present in the second community only.
3 Results and discussion
3.1 Species composition
In this study, a total of 44 woody plant species categorized into 40 genera and 29 families were recorded from the Hereje natural forest. The species richness was higher than that of Yemrehane Kristos Church forest (39 woody species) [21]. It is, however, smaller than that of the Wotagisho forest, which is also located in the same zone (51 woody species) [22]. Furthermore, the Hereje forest has a smaller number of woody plant species (44) compared to other forest patches elsewhere in Ethiopia [1, 25, 26].
In terms of habit, the woody species composition is 30 (68.2%) trees and 14 (31.8%) shrubs. The number of tree species is less than that of the Chato forest (50 species) [27], the Wotagisho forest (35% species) [28], and the Kenech forest (35 species) [5].
Fabaceae was the most dominant family, with five genera and six species, followed by Combretaceae, which had two genera and three species. Other seven families, Myrsinaceae, Myrtaceae, Oleaceae, Rhamnaceae, Rubiaceae, and Tiliaceae were represented by two genera and two species each. In contrast, each of the other 20 plant families was represented by a single species and genus (Fig. 2).

Families of plants with highest number f plant species
Fabaceae were the dominant family in the Hereje forest. Similar findings have also been reported in the Yemrehane Kristos Church forest [29]. This, however, contradicts the finding of Gojamme and Tanto [28], who reported the dominance of Solanaceae in the Wotagisho Forest. Because the current study was limited to woody plant species, the floristic composition of the study area may not have revealed all of the information.
Syzygium guineense (Willd.) DC. (23 (51.1%)), Rytigynia neglecta (Hiern) Robyns (22(48.9%)), Maytenu sgracilipes (Welw.exOliv) Exell(21 (46.7%)) and Pterolobium stellatum (Forssk.) Brenan (20(44.4%)) are the four most common woody species in the Hereje Natural forest.
3.2 Plant community types
The agglomerative hierarchical cluster analysis used in this study revealed three plant community clusters (Fig. 3). The highest mean cover-abundance value indicates the dominant species in a given community (Table 1).

Dendrogram that represent plant Community types of Hereje forest. Where CI = community I, CII = community II, CIII = community III. Community type I = Plots 31,13,17,22,12,39,34,26,1,44,3,35.30,45,40,18,23,38,32,10 and 43. Community type II = Plots 29,20,11,8,5,41,33,19,15,2,37,21,24,42,7,27 and 4. Community type three = Plots 14, 28, 6, 9, 16, 25 and 36
The first community type noticed was Syzygium guineense-Carissa edulis, which was discovered between the altitudinal ranges of 1575–1705 m.a.s.l. In comparison to the other communities, this was discovered in the highest altitudinal gradient. This was represented by 21 quadrants (46.67%) (Fig. 3).This community had the most species diversity. It included species with medium and maximum mean abundance values, such as Syzygium guineense, Carissa edulis, Rytigynia neglecta, and Calpurnia aurea. Acacia hockii, Triumfitta brachyceras, and Rhamnus prinoides were also discovered to be associated with this community.
The second community type is the Calpurnia aurea –Combretummolle community, which can be found at an altitudinal ranging from1574 to 1626 m.a.s.l. It has 17 plots (37.78%) and is located in the intermediate altitudinal range (Fig. 3). This particular community type had a medium number plot distribution and species diversity. Unlike the other communities, it contained species with medium mean abundance values. Calpurnia aurea, Combretum molle, Maytenus gracilipes, Acacia Senegal and Veronia myriantha had the highest mean abundance values in this community. Grewia bicolor, Pterolobium stellatum, Croton macrostachyus and Brucea antidysenterica were additional species observed in this community.
The third community type is Ziziphus mauritiana–Maytenus gracilipes, which is found at an elevation of 1500–1610 m.a.s.l, and has 7 plots (15.56%). In comparison to other communities, this community was found at the lowest altitude and had the fewest plot distributions and species diversity. Unlike the other communities, it contained species with the lowest mean abundance values. Ziziphus mauritiana, Maytenus gracilipes, Syzygium guineense and Pterolobium stellatumwere identified as the species with the highest mean abundance values. Rhamnus prinoides, Triumfitta brachyceras, Carissa edulis, Maesa lanceolata, Millettia ferruguinea and a few other species were also present (Fig. 3).
3.3 Species diversity and richness
Community type II has the highest number of species (43), whereas community type III has the least number of species (33) (Table 2). Community type II, on the other hand, was the most diverse and had the greatest number of individuals with an even distribution. In other words, community type I had intermediate species evenness, while community III had the least. The difference in the number of species, degree of disturbance of forest, species cover-abundance values, plot slope, and other related factors may have contributed to the variability of each value between each community type.
The Hereje forest’s overall Shannon–Wiener diversity index (H′) and evenness values were 3.5 and 0.95, respectively. The studied forest had higher species diversity than Wotagisho Forest [28], Wurg Forest [30], and Yemrehane Kristos Church Forest [31], which had 3.38, 2.82, and 2.88 values, respectively.
3.4 Similarity among the plant community types
The similarity in species composition varied between community types, ranging from 0.49 to 0.46 (Table 3). The highest similarity ratio was observed between communities I and II (93.18%), followed by I and III (77.27%). Possible explanations for the observed maximum similarity include the close altitudinal similarities among the communities and comparable human activities in the areas. Community II and III, on the other hand, had the lowest similarity ratio (75.0%), indicating that they may have fewer common species. The less similarity could be attributed to differences in altitude, aspect, slope, and degree of disturbance [32].
3.5 Threats to the forest
During the study period, the researchers observed overgrazing, over-browsing by domestic animals, hunting for bush meat, and cutting of trees from the forest for building materials and firewood as the major threats to the vegetation composition of Hereje forest, while hunting hurts the wildlife fauna in the area. This is consistent with the findings of Siraj et al. [10].
4 Conclusion and recommendation
The current study revealed that the floristic composition of woody species in Hereje Forest was much lower than in other similar vegetation in Ethiopia. The species diversity, richness, evenness, and dominance also varied across community types. The overall Shannon–Wiener diversity and evenness of woody species in the Hereje forest were 3.5 and 0.95, respectively, indicating that the diversity and evenness of woody species are relatively high in the study forest. The variation among plant communities in terms of woody species diversity and composition is linked to various human influencing activities such as agricultural expansion, firewood collection, charcoaling, commercial logging, overgrazing, and house construction. This necessitates research into the effects of human activities on the species diversity and floristic composition of the forest. A more comprehensive study of all species composition using soil analysis is also very important. In a nutshell, other natural environment components like soil analysis should be part of the future research component of Hereje natural forest for effective conservation and sustainable resource management. The participation of the local communities in the forest management and conservation activities is the only pragmatic decision to ease the forest loss and its diverse resources as complex and valuable natural resource system.
Data availability
The data can be obtained by contacting the correspondent author of the manuscript.
References
Meragiaw M, Woldu Z, Martinsen V, Singh BR (2018) Woody species composition and diversity of Riparian vegetation along the Walga River, Southwestern Ethiopia. PLoS ONE 13(10):e0204733. https://doi.org/10.1371/journal.pone.02047
Wassie SB (2020) Natural resource degradation tendencies in Ethiopia: a review. Environ Syst Res 9:33. https://doi.org/10.1186/s40068-020-00194-1
Tamene Y, Tesfaye A, Sebsebe D (2011) Survey and documentation of the potential and actual invasive alien plant species and other biological threats to biodiversity in Awash National Park, Ethiopia. Manag Biol Invas 2:3–14
Tadesse Z, Kelbessa E, Bekele T (2017) Floristic composition and plant community analysis of vegetation in Ilu Gelan district, West Shewa Zone of Oromia region, Central Ethiopia. Trop Plant Res 4(2):335–350
Sewale B, Mammo S (2022) Analysis of floristic composition and plant community types in Kenech Natural Forest, Kaffa Zone, Ethiopia. Trees For People 7:100170. https://doi.org/10.1016/j.tfp.2021.100170
Dawud S, Chimdessa M, Sasikumar JM (2018) Floristic composition, structural analysis and regeneration status of woody species of Gemechis Natural Forest, West Hararghe zone, Oromia, Ethiopia. J Nat Sci Res 8(24):11–12
Kebebew M, Demissie H (2017) Floristic composition, structure and regeneration status of Riverine Forest at Nech Sar National Park of Ethiopia. J Nat Sci Res 7(7):1–13
Befkadu M, Mesele N, Tesfaye A (2019) Woody species composition, structure and environmental determinants in a moist evergreen Afromontane forest, southern Ethiopia. J For Res 31:1173–1186. https://doi.org/10.1007/s11676-019-00894-0
Tamiru B, Mohammed AS, Mekonnen AB (2021) Floristic composition, structure and regeneration status of woody plants in church forests of Dangila, Northwestern Ethiopia. Cogent Food Agric 7:1. https://doi.org/10.1080/23311932.2021.1911438
Siraj M, Zhang K, Sebsebe D, Zerhiun W (2016) Floristic composition and plant community types in Maze National Park, southwest Ethiopia. Appl Ecol Environ Res 15(1):245–262
Barbieri M, Barberio MD, Banzato F, Billi A, Boschetti T, Franchini S, Gori F, Petitta M (2021) Climate change and its effect on groundwater quality. Environ Geochem Health. https://doi.org/10.1007/s10653-021-01140-5
Barbieri M, Ricolfi L, Vitale S, Muteto PV, Nigro A, Sappa G (2019) Assessment of groundwater quality in the buffer zone of Limpopo National Park, Gaza Province, Southern Mozambique. Environ Sci Pollut Res 26(1):62–77. https://doi.org/10.1007/s11356-018-3474-0
Yemata G, Haregewoien G (2022) Floristic composition, structure and regeneration status of woody plant species in Northwest Ethiopia. Trees For People 9:100291. https://doi.org/10.1016/j.tfp.2022.100291
Siraj M, Zhang K (2018) Structure and natural regeneration of woody species at central highlands of Ethiopia. J Ecol Nat Environ 10(7):147–158
Tesfaye G, Teketay D, Fetene M, Beck E (2010) Regeneration of seven indigenous tree species in a dry Afromontane forest, Southern Ethiopia. Flora-Morphol Distrib Funct Ecol Plants 205(2):135–143. https://doi.org/10.1016/j.flora.2008.12.006
Kefalew A, Soromessa T, Demissew D (2022) Plant diversity and community analysis of Sele-Nono forest, Southwest Ethiopia: implication for conservation planning. Bot Stud 63:23
Honja T, Geta E, Mitiku A (2017) Mango value chain analysis: the case of Boloso Bombe WoredaDistrict, Wolaita Zone, Southern Ethiopia. Dev Country Stud 6(1):103–123
Kent M (2011) Vegetation description and data analysis: a practical approach, 2nd edn. Wiley, Hoboken
Vander-Maarel E (1976) Transformation of cover abundance value in phytosociology and its effect on community similarity. Vegetatio 39(2):97–144
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Shannon CE, Wiener W (1949) The mathematical theory of communication. University of Illinois, Chicago
Jayarman K (2000) A statistical manual for forestry research. FORSPA-FAO Publication, Bangkok
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Willey, Hoboken
Kent M, Coker P (1992) Vegetation description and analysis. A practical approach. Belhaven Press, London
Mekonen T, Ayele B, Ashagrie Y (2015) Woody plant species diversity, structure and regeneration status of Woynwuha Natural Forest, North West Ethiopia. J Agric Environ Sci 1(2):90–113
Meragiaw M, Woldu Z, Martinsen V, Sing BR (2021) Floristic composition and structure of the Kibate Forest along environmental gradients in Wonchi, Southwestern Ethiopia. J For Res 32:2669–2682. https://doi.org/10.1007/s11676-021-01305-z
Abdena F (2010) Floristic composition and structure of vegetation of Chato Natural Forest in Horo GuduruWollega Zone, Oromia National Regional State, west Ethiopia. Doctoral Dissertation, Addis Ababa University, Ethiopia
Gojamme DU, Tanto TT (2016) Floristic composition and diversity of woody plant species of Wotagisho forest, Boloso Sore Woreda, Wolaita Zone, Southwest, Ethiopia. Int J Nat Res Ecol Manag 1(3):63–70
Abunie A, Dalle G (2018) Woody species diversity, structure, and regeneration status of yemrehanekirstos church forest of lastaworeda, north wollo zone, amhara region, Ethiopia. Hindawi Int J For Res 2018:1–8. https://doi.org/10.1155/2018/5302523
Boz G, Maryo M (2020) Woody species diversity and vegetation structure of Wurg Forest, Southwest Ethiopia. Hindawi Int J For Res 1:1–17. https://doi.org/10.1155/2020/8823990
Girma M, Maryo M (2018) The diversity and composition of woody plants in CheberaChurcura National Park (CCNP), Southern. Open J Fores 8(4):439–458
Yirga F, Marie M, Kassa S, Haile M (2019) Impact of altitude and anthropogenic disturbance on plant species composition, diversity, and structure at the Wof-Washa highlands of Ethiopia. Heliyon 5(8):e02284. https://doi.org/10.1016/j.heliyon.2019.e02284
Acknowledgements
The authors are grateful to Ambo University for funding the research work.
Funding
This study was conducted with financial support obtained from Ambo University.
Author information
Authors and Affiliations
Contributions
All authors AA, SM and MT contributed equally to the design, collection, and analysis of data, as well as manuscript preparation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interests.
Ethical approval
This research project was approved by Research Ethics Committee of the College of Natural and Computational Sciences of University (project number NCS99/2018). All methods were carried out in accordance with the guidelines and regulations of the Research Ethics of Ambo University.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Anamo, A., Mammo, S. & Temesgen, M. Floristic composition and community analysis of woody species in Hereje Natural Forest, southwest Ethiopia. SN Appl. Sci. 5, 48 (2023). https://doi.org/10.1007/s42452-022-05265-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42452-022-05265-9