
Abstract
Purpose
Amylase being one of the most important industrial enzymes requires large-scale production. When producing an enzyme, high productivity, high purity and low production costs need to be considered. This study focuses on comparing various agro-industrial waste substrates, for production of alpha-amylase using Bacillus amyloliquefaciens. Moreover, it studies the stability and activity of amylase–gold nanoparticles composite.
Methods
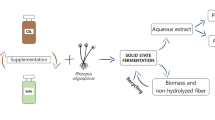
This study is divided into two parts, in the first part various agro-industrial waste substrates, such as wheat bran, rice bran and potato peel were used to produce alpha-amylase using solid-state fermentation (SSF). The production of the enzyme was quantified and compared in specific enzyme activity units. In the second part, change in the stability and activity of amylase in enzyme–gold nanoparticles (AuNPs) composite has been discussed.
Results
Highest enzyme production was observed in wheat bran and potato peel substrate with specific enzyme activity of almost 1.2 U/ug and 1.1 U/ug. Among combination substrates, wheat bran with potato peel showed a high enzyme production of 1.3 U/ug. On the other hand, the optimum temperature for amylase activity shifted to 55 °C in the composite compared to 37 °C for free enzyme.
Conclusions
Comparison of specific enzyme activity of extracts from various substrates showed that wheat bran alone, and in combination with potato peel, produces active and pure amylases. To stress on various catalytic activities of alpha-amylase, the capability of the enzyme to synthesize gold nanoparticles and the effect of conjugation of the nanoparticle on its optimum catalytic activity are also discussed in this paper.
Similar content being viewed by others


Introduction
With growing population, demands of agro-industrial products are increasing exponentially. To meet this high demand, an increased quantity of food products is being produced, leaving an elevated level of agro-industrial wastes. It has been reported that approximately 998 million tons of agricultural wastes are produced yearly all over the world (Agamuthu 2009). Considering the need of managing the huge amount of waste produced, the Agricultural Waste Management System has been formulated under which a 3R approach is used (Hai and Tuyet 2010). The 3R approach stands for reduce, reuse and recycle. Due to high carbon content in agro-industrial wastes, they can be used as substrates in producing hydrolytic enzymes that hydrolyse glycosidic bonds and convert polysaccharides into simple sugars. Among many such enzymes, α-amylase is one of the most commercially exploited enzymes.
Amylases are found in all life forms ranging from microorganisms, plants to animals (Sidkey et al. 2010). Microbes have two types of amylases—α-amylase and glucoamylase. α-Amylase is an extracellular enzyme that cleaves glucosidic linkages between adjacent glucose units in a linear amylose chain (Pandey et al. 1999). Among various extracellular enzymes, α-amylase has wide applications in many sectors such as clinical, medicinal and analytical chemistry. Beside its use in starch saccharification, it also finds application in baking, brewing, detergent, textile, and paper industries as well as distilleries (Ramachandran et al. 2004b).
The cost of enzyme production is high, which necessitates reduction of production cost by alternative methods such as utilizing solid-state fermentation (SSF) (Castilho et al. 2000; de Castro et al. 2010). The contents of synthetic media are very expensive and these contents might be replaced with more economically available agro-industrial wastes (Ikram-ul-Haq et al. 2003; Ramachandran et al. 2007; Queiroz et al. 2016). The use of agro-industrial waste also makes SSF an attractive method (Pandey et al. 1999). Several studies have reported amylase production from agricultural wastes such as wheat bran, rice husk, coconut oil cake, sugarcane bagasse and corn cob using SSF (Baysal et al. 2003; Ramachandran et al. 2004a; Anto et al. 2006; Khan and Yadav 2011; Kumar et al. 2013; Aliyah et al. 2017; Al-awsy et al. 2017).
Wheat, rice and potato are famous staple foods found all over the world, making their wastes (wheat bran, rice bran and potato peel) a concern for us. This work proposes a use of these agro-industrial wastes in α-amylase production using Bacillus amyloliquefaciens in SSF. A comparison of these wastes as substrate for amylase production showed that wheat bran and potato peel produce a high amount of α-amylase.
On the other hand, immobilization of enzymes is an emerging field of research which may address various stability issues of otherwise less stable enzymes in their free form. Additionally, immobilized enzymes can be recovered and reused, hence lowering the cost (Cowan and Fernandez-Lafuente 2011; Liese and Hilterhaus 2013). Previous studies have shown that immobilizing enzymes, such as lipases and redox enzymes, to nanoparticles increases their stability while retaining their activity (Dyal et al. 2003; Xiao et al. 2003; Willner et al. 2006). The process of immobilization can be achieved by functionalizing the nanoparticles and enzymes under physiological condition forming enzyme–nanoparticle composite (Rangnekar et al. 2007).
In this work, alpha-amylase–gold nanoparticle composites were synthesized, and the activity of these composites was estimated at various temperatures. It was observed that the optimum temperature of the enzyme shifted in the composites as compared to that of free enzyme, in addition to the change in the rate of activity. The activity of the enzyme in the composite was highest at 55 °C. This property change in the enzyme can be harnessed positively as per requirements.
Materials and methods
Test organism
The microorganism used in this study is Bacillus amyloliquefaciens, obtained from Institute of Microbial Technology (IMTECH), Chandigarh, India. The culture was grown and maintained in Spizizen potato agar (SPA) slants.
Fermentation
1 ml of overnight culture was inoculated in the production medium containing 100 gm of starch substrate with 4 ml of mineral solution (12.5% NH2NO3, 5% KH2PO4, 2.5% MgSO4.7H2O and 2.5% NaCl) in an Erlenmeyer flask, which was incubated at 37 °C for 4 days at 100 rpm.
Extraction of enzyme
The enzyme was extracted at the end of 4 days using 4-time v/v of 0.05 M phosphate buffer at pH 7.0. The buffer was added to the culture flasks and agitated at 150 rpm for 1 h. The mixture was filtered using a muslin cloth and the filtrate was centrifuged at 5000 rpm for 30 min. The supernatant was collected and used as the crude enzyme source.
Enzyme activity assay
Amylase activity was determined by incubating a mixture of 1 ml of enzyme source and 1% soluble starch at 40 °C for 30 min. The reaction was stopped by adding 2 ml of DNS (3,5-dinitrosalicylic acid) followed by boiling for 10 min. The final volume was increased to 20 ml with sterilized water and the sugar was determined by measuring the absorbance at 540 nm wavelength. The values were obtained and compared with the standard Maltose graph. One unit of enzyme activity is the amount of enzyme that releases 1 micro-mol of glucose per minute and is expressed in U/ml.
Estimation of total soluble protein
The total soluble protein was estimated using Folin–Ciocalteu reagent method and the absorbance was measured at 660 nm. The values were compared with the standard BSA graph to obtain the protein content in mg/ml of sample.
Western blot
Crude extracts after fermentation from various substrates were loaded equally on SDS–PAGE gel. The proteins were transferred on nitrocellulose membrane and blocked and probed for alpha-amylase antibody. Phosphoglycerate kinase (PGK) was used as a loading control. The bands obtained from the western were then quantified using ImageJ and normalized to PGK.
Synthesis of AuNP–alpha-amylase composite
As reported previously (Rangnekar et al. 2007), a solution of alpha-amylase in phosphate buffer (10 mM phosphate buffer pH 7.0) was prepared by adding 10 mg of the enzyme to 10 ml buffer. To this solution, 262.0 μL of HAuCl4 (1.6272 × 10−3 M) was added so that the final concentration was 4.5 × 10−4 M. The mixture was incubated at 37 °C for 48 h. The formation of gold nanoparticles (AuNPs) was monitored by recording UV–vis spectra of the solution at regular intervals, in the range of 400–800 nm, using a PerkinElmer Lambda 50 UV–vis spectrophotometer.
Isolation and purification of AuNP–alpha-amylase composite
After 48 h, the solution turned dark red and was centrifuged at 20,000 rpm at 15 oC for 20 min. The supernatant was discarded and the pellet collected at the bottom of the tube was suspended in the same volume of buffer as the initial one. This cycle was continued for twice more to have the composite devoid of unreacted enzyme. Finally, the composite was suspended in 10 ml of buffer solution (which is the same as the initial volume). This solution was used as the stock solution for all subsequent experiments.
UV–visible spectroscopic study
The AuNP–alpha-amylase composite solution, synthesized as above, was characterized by UV–vis spectroscopy. A PerkinElmer Lambda 50 UV–vis spectrophotometer was used for this purpose. The absorption spectra were recorded in the range of 400–800 nm.
Estimation of protein content in the composite
A standard Bradford assay method was used for the estimation of protein in pure amylase as well as in the composite. The dye used for the assay was Coomassie Brilliant Blue. Bradford Stock Solution was prepared by mixing 100.0 mL 95% ethanol, 200 mL 88% phosphoric acid and 350.0 mg of the dye. This stock solution was indefinitely stable at room temperature. Bradford Working Buffer was prepared by mixing 425.0 mL distilled water, 15.0 mL 95% ethanol, 30.0 mL 88% phosphoric acid and 30.0 mL Bradford Stock Solution. This solution was filtered through Whatman No. 1 paper and was stored at room temperature in a brown glass bottle. First, the Bradford Assay standard curve was obtained by performing the assay with BSA (1 mg/mL) taking samples of known protein concentration in the range 0–60 μg and diluting up to 300.0 μL with experimental buffer. 3.0 ml of Bradford working buffer was added to each of them and A595 was observed after 2 min. From that, the standard curve protein content in both pure alpha-amylase (1 mg/ml solution) and AuNP–alpha-amylase composite (stock solution) was found out. The estimation of composite was also ascertained by measuring the unreacted enzyme (alpha-amylase) in the supernatant by the same method.
Starch digestion kinetics studies with pure enzyme and the composite
Time-dependent kinetic studies of pure alpha-amylase and AuNP–alpha-amylase composite digestions of starch were performed using the iodine indicator method, which is based on the formation of starch–iodine complex. An equal protein concentration from pure enzyme and AuNP–alpha-amylase composite solution was added to the starch solution. The mixture was incubated at a fixed temperature until the starch digestion study was completed. An aliquot of 0.15 mL was taken from the mixture at regular time intervals and was diluted with 2.0 ml of phosphate buffer. To this, 0.1 mL of 1:10 diluted I2 solution (Gram’s iodine) was added followed by the measurement of visible spectrum. The starch degradation kinetics was followed by measuring absorbance in the range of 400–800 nm. The rate of starch digestion by pure amylase and AuNP–alpha-amylase composite at different temperatures was determined.
Results and discussion
Estimation of amylase enzyme activity from various waste substrates
Agro-industrial waste is a big concern for us, as they need to be recycled and managed. Therefore, three very common agro-industrial wastes—wheat bran, rice bran and potato peel were used as substrates for alpha-amylase production using Bacillus amyloliquefaciens in SSF. After 4 days of incubation at 37 °C, the enzyme was extracted and the enzyme activity of this crude extract was measured using 1% starch solution and DNS solution (described in Materials and methods). The productivity from the substrates was measured in terms of enzyme activity. One unit of enzyme activity is the amount of enzyme that releases 1 micro-mol of glucose per minute and is expressed in U/ml. Enzyme activity was determined by comparing the observed absorbance with the standard maltose curve, the activity was later normalized to the dilution factor.
A high titer of amylase activity was obtained from the medium containing wheat bran (112 U/ml) as a substrate followed by potato peel (89 U/ml) and rice bran (77 U/ml) (Fig. 1a). These substrates were also evaluated in their respective mixtures with equal percentage amount (Fig. 1b). The combination of wheat bran and potato peel also produced a high amount of amylase (99 U/ml) followed by other combinations.

Amylase enzyme activity from agro-industrial waste substrates. a Enzyme activity (U/ml) of crude extracts from substrates containing wheat bran, potato peel or rice bran. b Enzyme activity (U/ml) of crude extracts from substrates containing a combination of equal ratio of various starchy waste substrates—wheat bran + potato peel (1:1), wheat bran + rice bran (1:1), potato peel + rice bran (1:1) and a mixture of the three (1:1:1)
Estimation of specific enzyme activity from various waste substrates
High productivity and high purity are requisites of large-scale production of enzymes. To estimate the purity of enzyme, we need to determine the protein concentration in the extracts. This has been achieved using Folin–Ciocalteu reagent and measuring absorbance at 660 nm and extrapolating the protein concentration using standard BSA curve. The protein concentration in the crude extract is higher than the substrates containing wheat bran (Table 1). Similar results were also observed from Western blot and its quantification (Fig. 2a).

Specific enzyme activity of amylase from agro-industrial waste substrates. a Western blot of alpha-amylase and phosphoglycerate kinase (PGK) as a loading control (upper panel) from extracts of respective waste substrates. Quantification of Western blot bands of alpha-amylase normalized to that of PGK (lower panel), to estimate the amount of amylase. b Specific enzyme activity (U/ug) of extracts from substrates containing wheat bran, potato peel or rice bran. c Specific enzyme activity (U/ug) of extracts from substrates containing a combination of equal ratio of various starchy waste substrates—wheat bran + potato peel (1:1), wheat bran + rice bran (1:1), potato peel + rice bran (1:1) and a mixture of the three (1:1:1)
Specific enzyme activity is the number of enzyme units per ml divided by the protein concentration; it is an important measure of enzyme purity and quality. Figure 2b, c shows amylase activity from various starchy substrates. Media containing starchy substrates from wheat bran and potato peel showed a high specific amylase activity titer.
Amylases have been produced by several bacterial species, one of the most commonly used being Bacillus amyloliquefaciens. We produced alpha-amylase using Bacillus amyloliquefaciens from substrates containing starch from agro-industrial wastes—wheat bran, potato peel and rice bran. The substrates containing wheat bran shows high alpha-amylase production with a higher specific enzyme activity, this is consistent with previous reports that show high yield of amylase from wheat bran substrate (Babu and Satyanarayana 1995; Baysal et al. 2003). Another report suggests wheat bran as the best source for amylase production using Bacillus species from agricultural wastes (Ikram-ul-Haq et al. 2003). Previously, substrates with rice bran waste proved to be promising substitute for wheat bran in amylase production using fungal culture (Anto et al. 2006), however, we see a reduced production of alpha-amylase from rice bran substrate using Bacillus species. Recent studies have reported low levels of amylase production from rice husk substrate (Aliyah et al. 2017); however, they report high amylase production from potato peel substrate (Al-awsy et al. 2017).
Synthesis of AuNP–amylase composites
To characterize the enzyme activity, we produced a stable amylase–gold nanoparticle (AuNP) composite as previously reported, by the reaction of alpha-amylase with AuCl-4 (Rangnekar et al. 2007). Commercially available pure alpha-amylase was used for this purpose. The composite can be isolated and purified and the catalytic activity of the enzyme is retained in the composite. Following the same protocol, when native alpha-amylase solution (at pH 7.0) was incubated with AuCl4−, the solution turned light pink in 6 h and dark purple in 48 h, indicating the formation of AuNP composite in the solution. The formation of AuNP composite was further monitored by recording the characteristic surface plasmon resonance absorption peak of AuNP at 530 nm. The control sample which did not contain alpha-amylase showed no color in the visible region confirming that alpha-amylase is responsible for synthesizing AuNPs from HAuCl4 (Fig. 3a). The synthesized composite was centrifuged at 20,000 rpm at 15 °C for 20 min, producing a dark purple-colored pellet. The pellet was washed with phosphate buffer to remove all of the unreacted enzymes. The pellet so obtained was dispersed in the same initial volume of buffer and the UV–vis spectrum of the composite solution at 530 nm showed a single peak (Fig. 3b), suggesting that spherical NPs were present in the medium. This observation is consistent with earlier observations with respect to the same composite and hence the process described above resulted in the production of well-dispersed AuNPs–alpha-amylase composite (Rangnekar et al. 2007).

Synthesis and characterization of AuNP–alpha-amylase composites. a Synthesis of gold nanoparticle (AuNP) by α-amylase. Photographs of AuNP solution synthesized by α-amylase (left) and control sample (right). b UV/vis spectra of AuNP–alpha-amylase composite solution. c Starch digestion kinetics of pure alpha-amylase at 37 °C. d Graph of rate constants at various temperatures showing the optimum temperature of pure alpha-amylase solution. e Graph of rate constants at various temperatures showing the optimum temperature of AuNP–alpha-amylase composite solution
The protein content of the composite solution was estimated by Bradford assay. Extrapolating the data from BSA standard curve revealed that a definite volume of AuNP–alpha-amylase stock solution contained 40% of protein as compared to the same volume of 1 mg/mL pure alpha-amylase solution. In other words, 40% of 1 mg/mL of alpha-amylase gets incorporated in the composite.
Temperature-dependent enzymatic activity study
Alpha-amylase obtained from different sources show different optimum temperatures of its catalytic activity. Hence, alpha-amylase used in these experiments was standardized first. Starch digestion activity of alpha-amylase was studied at different temperatures ranging from 30 to 50 °C. Starch and enzyme were mixed at a particular temperature and aliquots were taken from the mixture at regular intervals to measure the UV–vis spectra using the iodine solution. The rate of digestion of starch by the enzyme so obtained was plotted against time at different temperatures. The digestion kinetics followed nearly single exponential decay at all temperatures (representative data shown only for 37 °C in Fig. 3c). From the graphs at different temperatures, rate constants were determined assuming that the enzymatic reaction is of first order. The optimum temperature of amylase activity was determined to be around 37 °C as shown in Fig. 3d.
A similar experiment was performed with AuNP–alpha-amylase composite at a temperature range of 30–60 °C. The optimum temperature of catalytic activity of the composite was found to be at 50 °C, which is significantly different from that of free amylase (Fig. 3e). This increase in optimum temperature of activity could be attributed to enhanced stability of the enzyme in the form of AuNP composite. Although the enzymatic activity was retained in the composite at the temperature reported, it is interesting to observe that the rate of digestion of starch by pure enzyme is higher than that by enzyme in the composite. The decrease in the rate in case of composite can be explained based on the fact that enzyme immobilized on the surface of AuNPs might not be properly oriented to project its catalytic site with the proper orientation for starch digestion as pure enzyme. A possibility of low reaction rate might be the fact that the enzyme binds to the AuNPs through its free thiol groups which can affect the orientation of the enzyme in composites. Furthermore, it is plausible that the exact mechanism of starch digestion by the enzyme and the enzyme in the composite are different. In other words, the binding site and the rate of binding of starch to the enzyme in the two cases could be different. This calls for further supporting studies to find the exact mechanism involved in the starch digestion by the composite, which is out of the framework of this study. Additionally, with the developing techniques in protein engineering, extra amino acids with thiol groups (such as cystines) can be engineered at the C-terminus of the enzyme, which can be used to form highly stable and active AuNP–enzyme composites.
Conclusions
Amylases are one of the most important industrial enzymes that need large-scale production when compared to other enzymes. High productivity, high purity and low production cost are the major necessities of large-scale production.
This work compared agro-industrial wastes as substrates to produce alpha-amylase using Bacillus amyloliquefaciens. In addition to wheat bran, a combination of wheat bran and potato peel showed a high production of amylase. The purity of enzyme was assessed by determining the protein concentration using BSA spectrometric assay and Western blotting. Furthermore, estimation of specific enzyme activity of extracts from various substrates showed that wheat bran alone, and in combination with potato peel, is capable of producing amylases with high activity and high purity.
We also studied the temperature dependence on the catalytic activity of the enzyme when functionalized with gold nanoparticles as compared to the free enzyme. The thiol groups of the native enzyme were proposed to be responsible for the synthesis and stabilization of the AuNPs (Rangnekar et al. 2007). The composite formed was characterized by UV–vis spectroscopy. Although we observed a slower rate of digestion by the composite (in comparison to the pure enzyme), may be due the immobilization of the enzyme on AuNP surfaces and hence lacking the proper orientation required for catalysis, the enzymatic activity was retained in the composite and the starch degradation kinetics followed similar trends in both the cases. An increased optimum temperature of activity was observed for the composites indicating the possibility of enhanced stability.
References
Agamuthu P (2009) Challenges and opportunities in Agro-waste management: an Asian perspective. Inaugural meeting of first regional 3R Forum in Asia 11–12 Nov., Tokyo, Japan. http://www.uncrd.or.jp/content/documents/Session2_Agamuthu.pdf. Accessed 1 Sept 2019
AL-awsy M, Obiady SA, Obaidi AA (2017) Producing of amylase enzyme from thermophilic bacteria using agricultural wastes as a substrate. Aust J Basic Appl Sci 11(13): 158–164. http://www.ajbasweb.com/old/ajbas/2017/October/158-164.pdf
Aliyah A, Alamsyah G, Ramadhani R, Hermansyah H (2017) Production of α-Amylase and β-glucosidase from Aspergillus niger by solid state fermentation method on biomass waste substrates from rice husk, bagasse and corn cob. Energy Procedia 136:418–423. https://doi.org/10.1016/j.egypro.2017.10.269
Anto H, Trivedi UB, Patel KC (2006) Glucoamylase production by solid-state fermentation using rice flake manufacturing waste products as substrate. Bioresour Technol 97(10):1161–1166. https://doi.org/10.1016/j.biortech.2005.05.007
Babu KR, Satyanarayana T (1995) α-Amylase production by thermophilic Bacillus coagulans in solid state fermentation. Process Biochem 30(4):305–309. https://doi.org/10.1016/0032-9592(95)87038-5
Baysal Z, Uyar F, Aytekin C (2003) Solid-state fermentation for production of a-amylase by a thermotolerant Bacillus subtilis from hot-spring water. Process Biochem 38:1665–1668. https://doi.org/10.1016/S0032-9592(02)00150-4
Castilho LR, Polato CMS, Baruque EA, Sant’Anna GL Jr., Freire DMG (2000) Economic analysis of lipase production by Penicillium restrictum in solid-state and submerged fermentations. Biochem Eng J 4(3):239–247. https://doi.org/10.1016/S1369-703X(99)00052-2
Castro AM, Carvalho DF, Freire DMG, Castilho LR (2010) Economic analysis of the production of amylases and other hydrolases by Aspergillus awamori in solid-state fermentation of babassu cake. Enzyme Res 576872:9. https://doi.org/10.4061/2010/576872
Cowan DA, Fernandez-Lafuente R (2011) Enhancing the functional properties of thermophilic enzymes by chemical modification and immobilization. Enzyme Microb Tech 49:326–346. https://doi.org/10.1016/j.enzmictec.2011.06.023
Dyal A, Loos K, Noto M, Chang SW, Spagnoli C, Shafi KV, Ulman A, Cowman M, Gross RA (2003) Activity of Candida rugosa lipase immobilized on gamma-Fe2O3 magnetic nanoparticles. J Am Chem Soc 125(7):1684–1685. https://doi.org/10.1021/ja021223n
Hai HT, Tuyet NTA (2010) Benefits of the 3R approach for agricultural waste management (AWM) in Vietnam. Under the framework of joint project on Asia resource circulation policy research working paper series. Institute for Global Environmental Strategies supported by the Ministry of Environment, Japan. https://iges.or.jp/en/pub/benefits-3r-approach-agricultural-waste. Accessed 1 Sept 2019
Ikram-ul-Haq Ashraf H, Iqbal J, Qadeer MA (2003) Production of alpha amylase by Bacillus licheniformis using an economical medium. Bioresour Technol 87(1):57–61. https://doi.org/10.1016/S0960-8524(02)00198-0
Khan JA, Yadav SK (2011) Production of alpha amylases by Aspergillus niger using cheaper substrates employing solid state fermentation. Int J Plant Anim Environ Sci (IJPAES). 5: 100–108. http://www.ijpaes.com/admin/php/uploads/65_pdf.pdf
Kumar MS, Chandana L, Sridevi V (2013) Production and optimization of Glucoamylase from wheat bran by Aspergillus oryzae NCIM 1212 under solid state fermentation. Int J Appl Innovation Eng Manage IJAIEM). 2: 318–323. https://www.ijaiem.org/volume2issue10/IJAIEM-2013-10-31-073.pdf
Liese A, Hilterhaus L (2013) Evaluation of immobilized enzymes for industrial applications. Chem Soc Rev 42:6236–6249. https://doi.org/10.1039/C3CS35511J
Pandey A, Selvakumar P, Soccol CR, Nigam P (1999) Solid state fermentation for production of industrial enzymes. Curr Sci 77: 149–162. https://www.jstor.org/stable/24102923
Queiroz V, Awan A, Tasic L (2016) Low-cost enzymes and their applications in bioenergy sector. Agro-industrial wastes as feedstock for enzyme production: apply and exploit the emerging and valuable use options of waste biomass. Academic Press, Cambridge, pp 111–131. https://doi.org/10.1016/B978-0-12-802392-1.00005-8
Ramachandran S, Patel AK, Nampoothiri KM, Chandran S, Szakács G, Soccol CR, Pandey A (2004a) Alpha amylase from a fungal culture grown on oil cakes and its properties. Braz Arch Biol Technol 47(2):309–317. https://doi.org/10.1590/S1516-89132004000200019
Ramachandran S, Patel AK, Nampoothiri KM, Francis F, Nagy V, Szakacs G, Pandey A (2004b) Coconut oil cake—a potential raw material for the production of alpha-amylase. Bioresour Technol 93(2):169–174. https://doi.org/10.1016/j.biortech.2003.10.021
Ramachandran S, Singh SK, Larroche C, Soccol CR, Pandey A (2007) Oil cakes and their biotechnological applications—a review. Biores Technol 98(10):2000–2009. https://doi.org/10.1016/j.biortech.2006.08.002
Rangnekar A, Sarma TK, Singh AK, Deka J, Ramesh A, Chattopadhyay A (2007) Retention of enzymatic activity of alpha-amylase in the reductive synthesis of gold nanoparticles. Langmuir 23(10):5700–5706. https://doi.org/10.1021/la062749e
Sidkey NM, Abo-Shadi MA, Al-Mutrafy AM, Sefergy F, Al-Reheily N (2010) Screening of microorganisms isolated from some enviro-agro-industrial wastes in Saudi Arabia for amylase production. J Americ Scien. 6(10): 326–339. https://pdfs.semanticscholar.org/2b60/619a8a07e35e3f41129075c95596fdf90631.pdf
Willner I, Baron R, Willner B (2006) Growing metal nanoparticles by enzymes. Adv Mater 18:1109–1120. https://doi.org/10.1002/adma.200501865
Xiao Y, Patolsky F, Katz E, Hainfeld JF, Willner I (2003) Plugging into enzymes: nanowiring of redox enzymes by a gold nanoparticle. Science 299(5614):1877–1881. https://doi.org/10.1126/science.1080664
Acknowledgements
Authors thank Allahabad Agricultural Institute-Deemed University (AAIDU), Allahabad, India and Indian Institute of Technology (IIT) Guwahati, India for providing the lab space and resources to complete this work.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
Authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Mojumdar, A., Deka, J. Recycling agro-industrial waste to produce amylase and characterizing amylase–gold nanoparticle composite. Int J Recycl Org Waste Agricult 8 (Suppl 1), 263–269 (2019). https://doi.org/10.1007/s40093-019-00298-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40093-019-00298-4