
Abstract
This study investigated the anaerobic digestion of an algal–bacterial biofilm grown in artificial wastewater in an Algal Turf Scrubber (ATS). The ATS system was located in a greenhouse (50°54′19ʺN, 6°24′55ʺE, Germany) and was exposed to seasonal conditions during the experiment period. The methane (CH4) potential of untreated algal–bacterial biofilm (UAB) and thermally pretreated biofilm (PAB) using different microbial inocula was determined by anaerobic batch fermentation. Methane productivity of UAB differed significantly between microbial inocula of digested wastepaper, a mixture of manure and maize silage, anaerobic sewage sludge, and percolated green waste. UAB using sewage sludge as inoculum showed the highest methane productivity. The share of methane in biogas was dependent on inoculum. Using PAB, a strong positive impact on methane productivity was identified for the digested wastepaper (116.4%) and a mixture of manure and maize silage (107.4%) inocula. By contrast, the methane yield was significantly reduced for the digested anaerobic sewage sludge (50.6%) and percolated green waste (43.5%) inocula. To further evaluate the potential of algal–bacterial biofilm for biogas production in wastewater treatment and biogas plants in a circular bioeconomy, scale-up calculations were conducted. It was found that a 0.116 km2 ATS would be required in an average municipal wastewater treatment plant which can be viewed as problematic in terms of space consumption. However, a substantial amount of energy surplus (4.7–12.5 MWh a−1) can be gained through the addition of algal–bacterial biomass to the anaerobic digester of a municipal wastewater treatment plant. Wastewater treatment and subsequent energy production through algae show dominancy over conventional technologies.
Graphical abstract

Similar content being viewed by others


1 Introduction
Renewable natural gas is a clean-burning energy carrier that can be fully interchanged with conventional natural gas (NG). The primary component of renewable NG is methane (CH4). Biogas can be upgraded to higher purity standards for the use as pipeline-quality fuel, for example, in the form of compressed natural gas (CNG) or liquefied natural gas (LNG) [1]. Biogas is produced from various biomass sources through anaerobic digestion. With minor clean-up, it can be burned to generate electricity and heat. Therefore, it has been gaining attention as a replacement for conventional NG.
Compared to European Union (EU) fossil fuels, biogas production can save up to 240% of greenhouse gas emissions (GHG) and biomethane up to 202% [2]. The production of biogas from renewable raw materials can be one interim solution for lessening global climate change. The EU is aiming at a 55% reduction in GHG emissions and a 32% share of renewable energy by 2030 [3]. By 2050, the EU plans to become carbon neutral [4]. Moreover, Germany aims at an 80% share of total renewable energy by 2050 [5]. However, in 2021 renewable energy sources overtook fossil fuels and provided 41.1% of Germany’s electricity demand. Germany has been striving to increase the share of renewable energy in other sectors such as heating and cooling (16.5% as of 2021) and transport (6.8% as of 2021) [6]. To reach these ambitious targets, the use of renewable energies must be intensified by increasing the share of renewable NG in energy-consuming sectors.
Microalgae can be one feasible source of alternative biomass for renewable energy production [7,8,9]. They can exhibit high growth rates under a wide range of environmental conditions. Biogas production by anaerobic digestion of algal biomass was first described in the 1950s. In the last decades, the focus has been partly on utilizing total microalgal biomass or even using the extracted oil/lipids from algae for biogas production [10]. However, most applications of microalgae for energy production remain underdeveloped mainly because of cost-intensive cultivation and harvest processes at an industrial scale [11]. Moreover, the investigation of the cultivation of microalgal biofilms for biofuel feedstock production has been limited to traditional open ponds and closed photobioreactors [12]. Algal biofilms are gaining attention from an economical perspective because of their high biomass density and relatively easier harvest. The Algal Turf Scrubber (ATS) is an innovative commercial system consisting of an inclined flow-way with an attachment substrate for the biofilm and a circulating culture medium [13]. The biofilm is a mesocosm of benthic bacteria, pro- and eukaryotic microalgae, and fungi embedded in their extracellular matrix (Fig. 1). Through complex mutual interactions and synergistic effects among the different species, the algal–bacterial biofilm can sequester a wide range of nutrients, for example, N, P, and K [14]. Thus, ATS systems are studied as alternative biological wastewater treatment technologies [15, 16], but so far, they have not been systematically evaluated as substrates for biogas production by anaerobic digestion. Previous studies focused more on defining microalgal biomass of ATS as fertilizer rather than wild cultures such as those found in wastewater treatment systems for energy production [17,18,19]. Microalgae usually have a high protein content and low C/N ratio, which is unfavorable for the anaerobic digestion [20]. Therefore, the inoculum and the biomass composition play an important role for methane productivity. Several authors have studied the methane productivity of defined microalgal biomass using different inocula such as sewage sludge, poultry manure, or food waste and showed that the inoculum must be tailored to the specific algae biomass outlining the need for ATS specific studies [20,21,22].

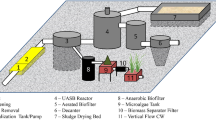
Illustration to the anaerobic digestion of algal–bacterial biomass of the Algal Turf Scrubber (ATS) system. 1: Algal Turf Scrubber (ATS, conditions: 6–30 °C, 5000–220,000 Lux, 0.0165 kg influent PO4-P m.−3); 2: Algal–bacterial biofilm (mesocosm, high N, P, C); 3: Anaerobic digester (operational conditions: 40 °C, 30 d digestion period)
Anaerobic digestion of microalgal biomass often suffers from low methane yields due to their recalcitrant cell walls [20] that need to be disintegrated using either thermal, mechanical, chemical, or biological (enzymatic) pretreatment methods for improved methane yields [23]. Many of these technologies have been evaluated in the last few decades to improve these processes. In particular, mechanical and thermal pretreatment technologies have been shown to effectively disrupt algal cells for the anaerobic digestion [24]. So far, studies on the impact of a pretreatment on the methane productivity of microalgae has been limited only to defined cultures. Therefore, its effect on complex substrates such as algal–bacterial biomass to increase methane production is still unknown [25].
To expand the renewable energy share in as many sectors as possible, the utilization of a mixed algal–bacterial consortium can be a promising approach. Therefore, this study focuses on the investigation of the methane potential of algal–bacterial biofilms using various industrial microbial inocula for their anaerobic digestion. In addition, the effects on the final methane yield caused by additional pretreatment steps of algal–bacterial biomass were evaluated. Based on previous findings and results of methane potential tests, different biorefinery concepts using algal–bacterial biomass of ATS system for methane production as well as for wastewater treatment at municipal wastewater and industrial biogas facilities were devised in the present study (Fig. 1). To assess the electric power potential of the methane production from algal–bacterial biomass of ATS systems at municipal wastewater facilities, scale-up calculations were conducted in this study.
2 Methods
2.1 Algal Turf Scrubber (ATS) system and biofilm
The ATS system (2.5 × 0.5 m) was photoautotrophically operated under local environmental conditions since 2019 (Fig. 2). It was kept in a greenhouse at the Forschungszentrum Jülich (FZJ, 50°54′19ʺN, 6°24′55ʺO), and growth was monitored throughout. In weekly intervals, the culture medium was renewed, and the biofilm was harvested by scraping it off the substrate.

a Wastewater remediation and alternative biomass production by algal–bacterial biofilm in an Algal Turf Scrubber system (ATS), b Algal–bacterial biofilm on the substrate
2.2 Methane potential tests of algal–bacterial biofilm
Anaerobic batch fermentation tests were conducted according to DIN38414 using an automatic methane potential test system (AMPTS II, Bioprocess Control, Sweden). The equipment consisted of a water bath with temperature control, 15 × 0.0005 m3 glass digestion bottles, stirrers, and 15 carbon dioxide (CO2)-absorption bottles. Each absorption bottle was connected to a digestion bottle and a tipping cup volumetric gas-measuring device. The batch tests were performed at 40 °C for up to 35 days. The real-time methane production from the digestion bottle was monitored using the AMPTS web-based software. The cultures were stirred at 30 rpm for 15 min per hour.
2.3 Substrates for methane potential tests
Freshly harvested biofilm (overall 2200 ml) was used as a substrate for the methane potential batch tests, and 50% of the untreated biofilm was used directly for digesting tests (Fig. 3a). The remaining biofilm was thermally pretreated (cooked) in sealed serum bottles at 105 °C for 2 h using an oven (Thermo Fisher Scientific). Four industrial digestates were tested as microbial inocula: (1) recycled waste paper of a laboratory biogas fermenter at NOWUM-Energy Institute (inoculum A); (2) mixture of manure-maize silage digestate of a full-scale biogas plant, Biogas Wassenberg GmbH & Co. KG, (inoculum B); (3) anaerobic sewage sludge of an anaerobic digester at the local municipal wastewater treatment plant (inoculum C); and (4) percolated green waste of a composting and digestion plant, Kompostierungs- und Vergärungsanlage Würselen (inoculum D) (Fig. 3b–e). The inocula were stored at room temperature and activated at 40 °C for 24 h at the start of each batch test.

Tested substrates for methane production a Algal–bacterial biomass grown in ATS and the microbial inocula, b digestate of recycled wastepaper of laboratory biogas fermenters (inoculum A), c mixture of manure and maize silage of a full-scale biogas plant (inoculum B), d anaerobic sewage sludge of the biogas fermenter of a municipal wastewater treatment plant (inoculum C), and e percolated green waste sludge of a full-scale biogas plant (inoculum D)
The inocula A and B were tested with algal–bacterial biomass harvested in July 2021. While the inocula C and D were tested with algal–bacterial biomass harvested in August 2021. The composition of respective algal–bacterial biomass at the time of harvest is listed in Table 1. A digestion bottle was filled with inocula A–D and untreated algal–bacterial biomass (UAB) or pretreated (PAB) (Table 2). The methane potential tests of algal–bacterial biomass were conducted in triplicate using inocula A and B and in single determination using inocula C and D as preliminary trials. The residual gas potential of each inoculum was measured as a control. The term methane potential of algal–bacterial biomass in the results represents the sole yield of algal–bacterial biomass after deducting the yield from the used inocula.
2.4 Analytical methods
The pH value of the biomass mixture in the digestion bottles was measured at the start and end of each batch test using GE 117 pH electrode (Greisinger® 600,770). The dissolved chemical oxygen demand (COD) of both the untreated and pretreated algal–bacterial biomasses was measured spectrophotometric using cuvette tests LCK 114, DIN 38,409-H41-H44, (Hach®, Germany). The samples were centrifuged at 13,000 rpm for 10 min before COD analysis. Dry matter (DM) and total organic or volatile solids (VS) of the algal–bacterial biomass were determined before methane potential tests according to DIN EN 12,880 and DIN EN 12,879. The elemental composition (technical duplicates) and the caloric value of the algal–bacterial biomass (technical triplicates) were measured using an ICP-OES spectrometer iCAP 6500, DIN EN ISO 11885 and DIN 51,732, (Thermoscientific®) and an isoperibol oxygen bomb calorimeter 6200, DIN 51,900, (Parr®), respectively.
3 Results and discussion
3.1 Methane potential batch tests
The methane potential of the four inocula (blank values) varied largely. Inocula A, B, and C yielded methane potentials of 0.016 ± 0.003, 0.03 ± 0.001, and 0.224 Nm3 CH4 kgVS−1, respectively. Inoculum D showed no methane potential in the batch tests. The substrate for inoculum D was extracted from a local composting and digestion facility. This sampled substrate contained only 2.1% DM, resulting in low organic carbon loading of 0.83% VS, which is not preferable for fermentation. In addition, inoculum D was sampled at a very late stage of substrate residence time during the fermentation at the local plant. Moreover, the initial pH of inoculum D in the digester bottle was 8.5, which was outside the suitable range (6.9–7.5) for the anaerobic digestion [26, 27]. These conditions together may have resulted in inoculum D being unable to generate methane potential in the batch tests.
3.2 Effect of microbial inocula on the specific methane potential of algal–bacterial biomass
Methane production of algal–bacterial biomass is a very sensitive process comprising mainly four consecutive stages of anaerobic digestion. Namely: (i) hydrolysis, (ii) acidogenesis, (iii) acetogenesis/dehydrogenation, and (iv) methanation. Complex molecules and compounds like carbohydrates, lipids, and proteins of the algal–bacterial biomass are converted into simple sugars, long-chain fatty acids, and amino acids by hydrolytic bacteria during the (i) hydrolysis. Then, fermentative bacteria convert these into volatile fatty acids, acetic acid, CO2, and hydrogen (H2) during the (ii) acidogenesis. The volatile fatty acids are further converted into acetic acid, CO2, and H2 by syntrophic bacteria and homoacetogenic bacteria during the (iii) acetogenesis. Yet, some acetic acid is converted back into CO2 and H2 by acetate oxidizing syntrophic bacteria. Finally, during the (iv) methanation stage either hydrogenotrophic or acetoclastic methanogenic archaea convert CO2 and H2 or acetic acid, into methane respectively [28]. These four stages explain the level of complexity in the involvement of required bacteria and archaea during the anaerobic digestion of algal–bacterial biomass for methane production.
Methane yield of algal–bacterial biomass mainly depends on the biomass composition (lipids, carbohydrates, and proteins) and (ii) the microbial inoculum [29]. The biomass composition changes according to the growing conditions in the ATS system. In our study, the ratio of ash: organic substance varied approximately by 35% between two harvests (July 2.11%, August 1.36%), due to environmental factors, Table 1. The detailed temperature, pH, and solar radiation profiles are given in the Supplementary information 1. The availability of organic substances during anaerobic digestion impacts methane productivity. One approach to increase organic substance in algal–bacterial biomass is the supply of nitrogen-rich wastewater to stimulate growth. Yet, nitrogen limitation increases the synthesis of lipids, fatty acids, or carbohydrates in algae, which can increase methane production [10]. However, in this study, the inoculum had a much greater effect than the substrate, Fig. 4.

Specific methane production of untreated algal–bacterial biomass (UAB) from the Algal Turf Scrubber system using microbial inoculum A (
 ) wastepaper digestate (n = 3); B (
) wastepaper digestate (n = 3); B (
 ) mixture of manure and maize silage (n = 3); C (
) mixture of manure and maize silage (n = 3); C (
 ) anaerobic sewage sludge (n = 1); and D (
) anaerobic sewage sludge (n = 1); and D (
 ) percolated green waste (n = 1). Norm cubic meter methane per kilogram volatile solids of algal–bacterial biomass
) percolated green waste (n = 1). Norm cubic meter methane per kilogram volatile solids of algal–bacterial biomass
We found the lowest specific methane productivity (0.120 ± 0.008 Nm3 CH4 kgVS−1) for inoculum A (wastepaper digestate) with untreated algal–bacterial biomass, Fig. 4. 513% increase in productivity (0.154 ± 0.008 Nm3 CH4 kgVS−1) was observed for inoculum B (mixture of manure and maize silage) comparatively to the inoculum itself. Similar productivity (0.15–0.117 Nm3 CH4 kgVS−1) was reported for dairy manure inoculum [19]. Methane productivity (< 0.006 Nm3 CH4 kgVS−1) was reduced for poultry manure compared to sewage sludge inoculum when using the three microalgal Chlorella sp., Nannochloropsis sp., and Scenedesmus sp. [20]. However, most available studies focused on microalgal biomass as a co-substrate rather than as a sole substrate. Yet, the studies show that the addition of microalgal biomass to cattle manure and maize silage affects biogas productivity and quality. The addition of Arthrospira platensis as co-substrate to cattle manure and maize silage increased methane yield (0.390 ± 0.011; at 10% of microalgal biomass to 0.581 ± 0.024 Nm3 CH4 kgVS−1; at 40% of microalgal biomass). The addition of Platymonas subcordiformis biomass to the maize silage and cattle slurry only slightly increased the methane yield from 0.487 ± 0.019 Nm3 CH4 kgVS−1 (at 10% of microalgal biomass) to 0.577 ± 0.024 Nm3 CH4 kgVS−1 (at 80% of microalgal biomass) [30].
The specific methane potential of algal–bacterial biomass using sewage sludge inoculum showed the highest yield, 0.350 Nm3 CH4 kgVS−1. Similarly, microalgae showed a higher methane yield using sewage sludge as inoculum compared to poultry manure [20]. Further, Du et al. [31] reported the methane potential of Spirulina platensis to be 0.343 Nm3 CH4 kgVS−1 using sewage sludge as inoculum. Passos et al. [32] discovered that the methane productivity of wastewater-grown microalgal biomass of a high-rate algal pond accounted for 0.117 Nm3 CH4 kgVS−1 using sewage sludge. Algal–bacterial biomass of ATS in this study attained higher values using sewage sludge (inoculum C) than in earlier studies (Fig. 4). The better adaption of inoculum C’s microbiome to the digestion of algal–bacterial biomass in the present study can be one reason for achieving higher methane yields. The lipid content and composition of algal–bacterial biomass itself could be another influential factor in attaining higher methane yields.
Algal–bacterial biomass of the ATS system generated the second-highest overall specific methane yield of 0.311 Nm3 CH4 kgVS−1 using inoculum D. Unexpectedly, no more methane productivity was observed from the digestion bottle after day 16 of anaerobic digestion (Fig. 4). This observation could be due to the unfavorable properties of inoculum D, namely that it contains very low organic loads. This property could result in low digesting potential. According to the plant operator who delivered inoculum D, the biogas potential of green waste at the Würselen plant ranges from 0.075 to 0.1 Nm3 kgfresh matter−1.
Typically, organic degradation is directly proportional to biogas production but not methane production, since biogas consists of methane, CO2 and other contaminants. According to Table 3, higher organic degradation took place during the digestion of algal–bacterial biomass using inocula A and B than inocula C and D. On the contrary, higher methane was produced using inocula C and D than inocula A and B (Fig. 4). Several factors could lead to lower biogas quality, which means lower methane productivity. One of the factors could be a higher conversion of acetic acid to CO2 and H2 by acetate oxidizing syntrophic bacteria in inocula A and B during acetogenesis, leading to a lower share of methane but higher CO2 in biogas. Since different species of microbiome are involved during the biogas production, their activity and proportions in the starting microbial culture impact organic degradation and eventually share of methane in biogas. A similar observation was found by Torres et al. [20]. The biogas from algal biomass was highly rich in CO2 (91%) using poultry manure, while biogas using sewage sludge was rich in CH4 (65%) in his study. A linear correlation between organic degradation and methane production of algal–bacterial biomass did not always exist and was found to be dependent on the inoculum used. This observation can be further explained by the daily flow rate of methane production in Table 3. Despite higher organic degradation during anaerobic digestion using inocula A and B, lower flowrates of methane production were identified compared to inocula C and D. It is assumed that the share of CO2 was higher in biogas using inocula A and B. Another factor could be total solids (TS) concentration. The average TS in the digesters of algal–bacterial biomass was observed to be 7.77%, 5.53%, 0.93%, and 2.26% using inocula A, B, C, and D, respectively. High TS concentration could negatively affect methane production.
Further, the nutrient concentrations influence the methane yields too. Mass fraction of C was lower in algal–bacterial biomass using inocula A and B than biomass using inocula C and D. By contrast, mass fraction of N and P were higher in algal–bacterial biomass using inocula A and B than biomass using inocula C and D. In the anaerobic digestion of mixed organic substrates, algal biomass acts as a source of micronutrients and macronutrients like N and P for the growth of other microorganisms [33]. However, high N concentrations lead to the rapid accumulation of volatile fatty acids (from 0.05 kg m−3) and total ammonia nitrogen (from 0.733 kg m−3) in the anaerobic digester resulting in digestion mortifications [34].
Furthermore, one must also consider the dominant microalgal species presented in the algal–bacterial biomass used for anaerobic digestion. Especially regarding the mixed algal–bacterial biomass, microalgal species vary in their biomass composition and subsequently exhibit different potentials for yielding biogas. Chlorella kessleri and Scenedesmus obliquus are regarded as species of low biogas potential, attaining only 0.33 ± 0.007 and 0.287 ± 0.001 Nm3 kgVS−1, respectively [35]. Singh et al. [36] and Parmar et al. [37] emphasized the impact of algal species on biogas production efficiency. The growth of microalgal species in ATS changed over time. Parallel investigations of algal–bacterial biomass through quantitative polymerase chain reactions were conducted [38]. They showed that the abundance of general microalgal groups and cyanobacterium in ATS harvested for inocula C and D was six times (1.93E + 14 copies microalgae m−3 sample) and 2.2 times (1.72E + 15 copies cyanobacteria m−3 sample) higher than in algal–bacterial biomass harvested for inocula A and B. Since each microalgal species has a different biogas production capacity, the population of each microalgal species in ATS is very important.
Water content is crucial for the mobility and nutrient supply of bacteria [20]. The average water content in the digesters of algal–bacterial biomass was observed to be 92.23%, 94.47%, 99.07%, and 97.74% using inocula A, B, C, and D, respectively. The low gas production in the digestion of algal–bacterial biomass using inocula A and B may be due to a lack of water [20]. Given all the above, the wide range of algal and bacterial species used in the current study limits the possibility of systematically assigning specific influential factors. More research is needed to further elucidate the kinetics of the digestion process and the relationship between ATS-derived algal–bacterial biomass and biogas production by anaerobic digestion.
3.3 Impact of thermal pretreatment of algal–bacterial biomass on its methane yield
Prior research on pretreatment methods determined that 3% higher algal cell degradation can be achieved at a higher temperature without increasing the exposure time (105 °C for 2 h) compared to a lower temperature with increased exposure time (80 °C for 2.5 h) [38, 39]. The procedure for evaluating the cell degradation rate of a pretreatment method that was inspired by previous experiments by Bischof et al. [40] was slightly modified in our prior research. The thermal pretreatment of algal–bacterial biomass led to an increase in dissolved COD from 0.468 to 4.379 kg m−3 in the biomass (harvested for inocula A and B) and from 0.487 to 4.907 kg m−3 (harvested for inocula C and D). The treatment resulted in cell degradation rates of 25% and 33% (based on COD concentrations) in the biomass, respectively. The increased COD concentrations in algal–bacterial biomass could be due to organelle disruptions and the release of organic matter during pretreatment. The increased COD in the biomass expands the availability of organic matter in algal–bacterial biomass for anaerobic digestion. The thermally pretreated algal–bacterial biomass (PAB) influenced methane productivity of algal–bacterial biomass for each inoculum, as shown in Fig. 5.

Specific methane production of thermally pretreated algal–bacterial biomass (PAB) from Algal Turf Scrubber system using microbial inoculum A (
 ) wastepaper digestate (n = 3); B (
) wastepaper digestate (n = 3); B (
 ) mixture of manure and maize silage (n = 3); C (
) mixture of manure and maize silage (n = 3); C (
 ) anaerobic sewage sludge (n = 1); and D (
) anaerobic sewage sludge (n = 1); and D (
 ) percolated green waste (n = 1). Norm cubic meter methane per kilogram volatile solids of algal–bacterial biomass
) percolated green waste (n = 1). Norm cubic meter methane per kilogram volatile solids of algal–bacterial biomass
The specific methane potential of PAB using inocula A and B increased to 0.140 ± 0.008 and 0.165 ± 0.014 Nm3 CH4 kgVS−1, respectively, following pretreatment. The methane potentials accounted for 116.4% and 107.4% higher using inocula A and B, respectively, when compared to the UAB. The specific methane production of PAB using inocula C and D was reduced to 50.6% and 43.5%, respectively, in comparison with UAB.
The variability of the methane yield can be attributed to pretreatment temperature, exposure time, inoculum acclimation, and the type of algal species. The solubilization of microalgal biomass is typically higher when higher temperatures (75–95 °C versus 55 °C) are applied, whereas longer exposure times do not have a significant effect (15 h versus 10 h) [10]. Many studies underline the positive impact of the thermal pretreatment of algal biomass on its methane yield. Marsolek et al. [41] found a 41% increase in methane yield using thermal pretreatment (at 90 °C PAB for 4 h) before algal anaerobic digestion relative to the control. Wang et al. [24] observed increases in methane productivity of 37% and 48% using thermal pretreatments (of 70 °C and 90 °C PAB for 0.5 h) in comparison with UAB. Other authors reported that thermal pretreatment (121 °C for 0.3 h) resulted in 0.322 Nm3 CH4 kgVS−1, which was 8% higher than UAB [24]. Table 4 shows further comparative studies on the methane yield of thermally pretreated algae. Methane yields of the 100 °C thermally PAB of the present work are in line with the literature. Table 4 shows that lower exposure times are more effective.
Methane yields obtained after algal thermal pretreatment (< 100 °C) could vary between a decrease of 13% to an increase of 12% in comparison with untreated algae for inocula such as anaerobic sewage sludge [43,44,45,46]. Our study corroborates these observations. The specific methane production of PAB for inocula C and D decreased in comparison with UAB. Similar results were found by Saleem et al. [47] who pretreated microalgae (Nannochloropsis oculata) through individual treatments and combinations of both ultrasonication and thermal methods (121 °C for 2 h) using cow manure as a co-substrate. Pretreatment may not always increase the biogas yield, and lower specific methane potentials were achieved (inocula C and D). This variability can be attributed to several factors, including inoculum acclimation and the type of algal species. One possibility for the reduced biogas yield is inhibitory effects that occur through a change in the physical properties of biomass fed into the anaerobic digester (like hydrophobicity), or inhibitory components are formed during fermentation. It should be further noted that a single pretreatment technology might not be suitable for all substrates and inocula. More research is required to understand which combinations of substrates are favorable for which pretreatment method.
3.4 Biorefinery and circular bioeconomy potential of algal–bacterial biofilm
3.4.1 Biorefinery pathways of algal–bacterial biofilm
By combining the results of this work and the literature study, we present possible biorefinery pathways for the nutrient-rich algal–bacterial biofilm of ATS systems (Fig. 6). Thanks to the mesocosmic species-assembly in the algal–bacterial biofilm, a wide range of high- and low-value compounds can be found, such as antioxidants (pigments) and vitamins, proteins (amino acids), minerals, sugars (biobutanol, bioethanol), carbohydrates (biogas, biochar), oils (bio-oil, chemicals, omega fatty acids) and lipids (biodiesel) [48,49,50,51]. Potential applications could be animal feed, soil ameliorate, plant stimulant and fertilizer, as well as bio-energy.

Schematic diagram of a biorefinery concept for nutrient-rich algal–bacterial biomass of an Algal Turf Scrubber (ATS) system. Orange: wastewater treatment by algal–bacterial biomass; blue: applicable extraction processes for harvested biomass; violet: respective fractions available from each extraction process; green: bioproducts resulting from each extracted fraction. The byproducts (heat, exhaust gas, and liquid effluents) resulting from the green channel are reused in the orange channel to fulfill the demands of wastewater treatment and algal cultivation
However, the valorization of algal–bacterial biomass derived from wastewater using systems such as open ponds or ATS has a limited range of applications. The biomass can potentially accumulate heavy metals, pharmaceutical residues, and other anthropogenic contaminations [52]. However, this algal–bacterial biomass can be converted into various biofuels for energy applications. Today, the transformation innovations of such fuels can be separated into four classifications: specific conversion through thermochemical reactions, conversion through biochemical reactions, conversion through transesterification, and photosynthetic conversion in microbial fuel cells [50, 53, 54]. Examples of thermochemical transformation include gasification (production of syngas), thermal liquefaction (production of bio-oil), pyrolysis (production of bio-oil, syngas, charcoal, etc.), and direct burning or combustion (generation of heat and electricity). Biochemical transformation processes mainly include anaerobic digestion (production of biogas), alcoholic fermentation (production of bioethanol), and photobiological hydrogen generation (production of biohydrogen) [50, 51]. Biodiesel can be obtained through the transesterification of algal fatty acids. During the production of these bio-based energy carriers, byproducts such as unused heat, wastewater, exhaust gases, and nutrient-rich wastes (such as digestate through anaerobic fermentation) are generated. These residual materials can be further utilized in various processes such as producing additional biogas. Unused heat from the system can be fed to the ATS system to maintain the system at an optimal temperature for efficient nutrient removal in water. Nutrients in wastewater or other waste streams can further serve as feed for the growth of an algal–bacterial consortium. To be able to reuse the waste heat and gas, the ATS should be established in a closed system in which unused heat and CO2 from exhaust gases can be fed into it. Research indicates that up to 50–60% of CO2 produced during the upgrading process of raw biogas after anaerobic digestion can be used by algae such as Chlorella vulgaris (in the case of a photobioreactor) [55]. However, it is challenging to integrate CO2 in an open system like an ATS, and therefore, assembling ideas for unused heat and CO2 must be designed. Nevertheless, these closed systems represent a promising way to mitigate CO2 emissions, remediate wastewater through algae, and establish an alternate bioenergy pathway for the sustainable production of bioenergy and bioproducts.
3.4.2 Circular bioeconomy in municipal wastewater treatment plants through algal–bacterial biomass
Municipal wastewater treatment plants often adopt advanced chemical treatment processes for P-removal, as the enhanced biological phosphate removal process alone cannot fulfill the required P discharge limits (0.002 kg m−3 according to EU regulation) [56]. In many municipal sewage wastewaters, there is an insufficient concentration of dissolved organics to allow bacteria to consume all the nutrients [57]. Furthermore, the conventional nitrogen and carbon removal processes (nitrification/denitrification) are very energy-intensive and require 60–75% of the electricity demand of a municipal wastewater treatment plant [58]. Although the de-ammonification process might improve the energy efficiency of nitrogen removal in the future, its process stability is still not well understood. Nutrient removal by algae could play an important role in making wastewater less energy-consuming. The utilization of microalgae for wastewater treatment was first reported in 1960 [59]. However, there are still no plants operating at an industrial scale. Despite the many advantages, microalgal harvest from photobioreactors and high-rated algal ponds remains still cost-intensive. Immobilizing algal cells on a surface or in a matrix can simplify the harvesting process and production costs [60]. In ATS systems algal–bacterial consortia grow on a stationary mesh net that is frequently flushed with nutrient-rich wastewater and can be easily harvested by scraping off the biofilm.
The algal–bacterial biofilm can remediate up to 1.83 kg CO2 kg−1algal biomass [61] and 0.01 kg Peliminated m−3 per week (as shown in parallel research to the current study [38]. Further, biogas can be produced through the anaerobic digestion of the algal–bacterial biomass. The biogas can be refined to methane or directly burned for electricity generation. The released CO2 could be directly led back to the ATS for bioremediation during the wastewater treatment in a circular process chain [62]. However, in regions with low temperatures or few light hours, the use of an ATS could be limited to optimal growth seasons. Additionally, large-scale ATS applications are currently limited by the large footprint of ATS flow-ways. These factors were in-depth analyzed in a scale-up calculation for nutrient-rich lake water (38°59′22ʺN, 76°56′44ʺW) by Witarsa et al. [19]. The calculations highlighted that up to 8.73 × 10−4 kg N∙km−2 d−1, 1.18 × 10−4 kg P∙km−2 d−1, and 52.5 × 10−4 kg C km−2 d−1 could be remediated by ATS algal-biofilm of 10,000 km2 size. The produced algal–bacterial biomass could be used in anaerobic digestion for biogas production and generate direct heating (2.8∙108 J a−1) or power a 1.13-kW generator [19].
Our scale-up calculations used data collected from the pilot-scale ATS system at the FZJ and the methane potential tests in our lab. In this study, nutrient-enriched water (0.3 m3, 0.00165 kg PO4-P m−3) was treated at weekly intervals on an ATS (1.25 × 10−6 km2) under greenhouse conditions. Approximately 1.48 million m3 of wastewater is treated yearly (28,000 m3 weekly) at an average municipal wastewater treatment plant according to the German Association for Water Management [63]. Based on our scale-up calculations, a 0.116-km2 ATS would be enough to treat a weekly average of 28,000 m3 of wastewater.
In a parallel study, the biomass productivity ranged from 0.0044 to 0.0115 kg m−2 d−1 according to the water temperatures of 14 °C and 25 °C, respectively (personal communication by Xinyu Gan, FZJ).
Additional scale-up calculations to the electric potential of algal–bacterial biomass used yearly mean biomass productivity of an ATS of 1.606 and 4.197 kg m−2 a−1 at 14 and 25 °C, respectively. With the 2.11% VS level of algal–bacterial biomass found in this study, the maximum production of yearly biomass corresponds to 0.0338 kgVS m−2 a−1 and 0.088 kgVS m−2 a−1 at the lowest and highest biomass productivity conditions, respectively. The methane potential in this biomass further accounts for a minimum of 0.011 Nm3 CH4 m−2 a−1 and a maximum of 0.030 Nm3 CH4 m−2 a−1 through the anaerobic digestion of ATS-derived biomass at the lowest and highest biomass productivity conditions, respectively. With an electrical conversion efficiency of 35% (standard: 3.48 kWh m−3 CH4) and specific methane potential of 0.350 Nm3 CH4 kgVS−1 (found in this study), the algal–bacterial biomass of the ATS could alone contribute minimum and maximum electric potentials of 0.041 and 0.107 kWh m−2ATS a−1, respectively. This would result in electric potentials of 4.7 to 12.5 MWh a−1 for the 0.116-km2 ATS system.
The ATS system is relatively easy to operate compared to other systems, and parameters such as hydraulic loading, flow-way length, and harvest period enable easy process control and optimization of the system for water treatment [64]. This optimization might reduce the size of ATS in the future. The specific power consumption of state-of-the-art municipal wastewater treatment plants ranges between 20 and 45 kWh PE−1 a−1 depending on the class of the plant [65]. By implementing algae-bacterial consortia in ATS systems for wastewater treatment, energy expenditure (300,000 kWh a−1) for the conventional nitrification–denitrification process at the municipal wastewater treatment plant (20,000 PE) is saved. Additionally, the 4.7–12.5 MWh a−1 of energy, estimated in the scale-up calculations, gained from algae shows dominancy over the conventional nitrification–denitrification process.
3.4.3 Circular bioeconomy in biogas plants through algal–bacterial biomass
In 2019, approximately 9500 biogas plants were operated in Germany [66]. The average biogas plant (output: 500 kWel) produced approximately 10,000 t of digestate per year [67]. Digestate is mainly enriched with nutrients like N and P. In general, N and P may serve as fertilizer, but the N and P are partly lost through soil erosion and leaching, leading to the eutrophication of water bodies and increased nitrate concentrations in the groundwater. Therefore, the Fertilizer Ordinance of 2020 has set a limit for digestate exploitation on fields to 170 kg N h−1 a−1 [68]. An area of approximately 412 ha is required to apply the untreated digestate of an average-sized biogas plant in Germany under the fertilizer ordinance. This figure exceeds the size of an average farm. To apply the digestate to a smaller area, the nutrients from the digestate must be recovered or eliminated.
Digestate treatment methods such as membrane separation and evaporation can efficiently gather organic matter, but these methods are energy-intensive [69, 70]. For most nutrient recovery techniques, solids must be separated [71]. Digestate solids can also be used as substrates in compost, providing sources of carbon and nutrients as fertilizer. Moreover, medium-density fiberboard, wood, or plastic composite material can be created using digestate solids without sacrificing mechanical or aesthetic properties [71]. Another opportunity for a circular bioeconomy would be to utilize the digestate (after nutrient recovery) as fertilizer for food and energy crops such as maize. The cultivated maize could be used as a feed and substrate alike.
The nutrients (e.g., N and P) of the liquid fraction of digestate need to be recovered. Besides light and CO2, microalgae require also other nutrients to grow. The algal–bacterial consortium of the ATS system in this work could assimilate and recover N and PO4-P from water. The nutrient-rich algal–bacterial biomass acted as a co-substrate in anaerobic digesters. Due to the rapid growth of biogas upgrading, there is significant potential for capturing the upgrading byproduct CO2. CO2 from biogas plants or biogas power plants could be directed back to the ATS system in which the photoautotroph organisms fixate CO2. With this process, GHG emissions can be reduced, nutrients recycled, and energy recovered through anaerobic digestion of algal–bacterial biomass. Thus, the digestate treatment via the algal–bacterial consortia of the ATS system is very promising for the circular bioeconomy at biogas plants, as shown in Fig. 6.
The calorific value of the algal–bacterial biomass must be further evaluated to properly estimate the potential application in biogas plants. The calorific value ranged between 15.3 MJ kgDM−1 and 19.35 MJ kgDM−1 in the current work. Some of the factors influencing the calorific value of biomass are weather conditions that affect the growth of algae and its harvesting frequency. With the harvesting frequency of 1 week used in this study, the calorific value ranged between 15.3 kgDM−1 and 18.8 MJ kgDM−1 during summer (06.-08.2021). By contrast, during the autumn and winter (09.-12.2021), where no harvest took place, the productivity ranged between 16.72 MJ kgDM−1 and 19.35 MJ kgDM−1.
The structure of the flow layer also appears to play a significant role in maintaining algal species diversity and density. By increasing the available surface area for turf colonization and by retaining a greater number of filamentous algae under the screen after harvesting, adding a biomass retention screen to the flow layer surface could improve both calorific value and nutrient removal performance [72]. The composition of algal biomass could vary among strains and vary due to the prevailing growth conditions, including nutrient composition and concentration, the duration of cultivation, light intensity, temperature, pH, and salinity [73, 74]. Insofar, the properties of the provided water or medium, as well as the selected strain, can influence the metabolism of the microalgae To date, there are only a few studies that have determined the calorific value of an algal–bacterial consortium of an ATS system. Most previous experiments were limited to indoor lab-scale cultivation studies using a limited number of strains to determine the nutrient removal efficiencies of targeted compounds [62]. Scragg et al. [75] found that the calorific value of mixed cultured Chlorella vulgaris and C. emersonii in a tubular reactor was about 28 MJ kgDM−1.
The energy potential of algal–bacterial biomass (kWh kgVS−1), converted using the results of methane batch tests, was further compared with its actual energy potential from its calorific value, as shown in Table 5. The actual energy potential (kWh kgVS−1) was calculated using the analyzed calorific value and solids (DM and VS).
The energy potential found using methane potential tests must be below or equal to the actual energy potential calculated using the calorific value and solids analysis. The energy potential of the algal–bacterial biomass of both of the analyses is plausible except in the case of the UAB using inoculum C. The energy potential of the UAB using inoculum C (0.3485 kWh kgVS−1) in methane batch tests was found to be 0.051 kWh kgVS−1 higher than its actual energy potential (3.434 kWh kgVS−1) calculated using its calorific value and solids, which is quite improbable. Synergetic effects during the anaerobic digestion of the UAB using inoculum C are, in exceptional cases, possible. However, errors in the analysis using the calorific value and solids are also possible. One should further consider that the batch tests using inoculum C were conducted in a single determination. Further investigations are therefore highly recommended.
The circular bioeconomy concept of biogas production incorporated in a microalgal-biorefinery loop could be more financially and provide surplus energy compared to the contemporary and conventional methodology. To encourage circular bioeconomy concepts, the microalgal production systems need to be further developed to produce systems with favorable energy and nutrient balances. With a prolific microalgal growth rate, the ability of algae to adapt to different wastewaters and uptake nutrients from wastewater, coupled with CO2 assimilation, could make the algal bioremediation of wastewaters and energy production promising. However, there are hurdles to realizing these benefits on a large scale. Energy generation from algal biomass may not be attractive on a commercial scale. This is mainly due to the high prices in the market for other bioproducts (e.g. proteins) that could be extracted from pure algal cultures. The residues from anaerobic digestion must be cleaned and processed for use in an ATS.
Furthermore, there is the challenge of yearly and seasonal variability in algal productivity in an ATS. The impact of winter conditions experienced at outdoor operations on nutrient capture by algal–bacterial consortia is still unknown. These challenges should be considered when determining anaerobic digestion sizing and expected biogas production [19]. However, biogas production from microalgae can be combined with other biomass-based refineries and wastewater treatments. In these integrated approaches, biogas generation is an advanced techno-economical one, especially through the elimination of costly biomass production steps that consume most of the energy used for microalgal cultivation, harvesting, and dewatering.
4 Conclusion
In this study, the methane potential of algal–bacterial biomass of an ATS system was investigated. The utilization of different industrial inocula (digestates) showed significant differences in methane yield rates during the anaerobic digestion of the algal–bacterial biomass with or without additional pretreatment. The correlation between the organic degradation and methane yield of algal–bacterial biomass was largely dependent on inoculum. Algal–bacterial biomass using wastepaper (inoculum A) and a mixture of manure and maize silage (inoculum B) had higher organic degradation but lower methane yields compared to sewage sludge (inoculum C) and percolated green waste (inoculum D). The order of performance of algal–bacterial biomass concerning methane yield begins with inoculum C, followed by inocula D, B, and A. Type of inocula played a significant role in both organic degradation and methane productivity of algal–bacterial biomass. Thermal pretreatment of algal–bacterial biomass produced a positive effect on methane yield for inoculum A (116.4%) and inoculum B (107.4%), whereas the pretreatment reduced productivity for inocula C (50.6%) and D (43.5%) relative to the untreated algal–bacterial biomass.
The biorefinery concepts of algal–bacterial biomass of ATS in this study aimed to mitigate GHG emissions and recover energy from algal–bacterial biomass through anaerobic digestion using nutrients and water recycling to treat wastewater in municipal treatment plants and digestate in biogas plants. Scale-up calculations done for this study showed that the commercial application of ATSs used for nutrient removal at municipal wastewater treatment plants could be limited due to the space requirements of ATSs. However, this limitation could be supplemented or mitigated in the form of energy savings from algae in comparison to the conventional nitrification–denitrification process. Moreover, energy gained from algae through local anaerobic digestion at the plants showed more electric and simultaneous economic potential than the conventional nitrification–denitrification process.
To utilize the algal–bacterial consortia of an ATS on a large scale, the role of individual microorganisms in the consortia needs to be better understood. It could be useful to improve P removal efficiency in wastewater and fulfill the discharge limits (0.002 kg P m−3) of EU regulations. The coupling of wastewater treatment with an ATS and subsequent valorization of produced algal–bacterial biomass into methane could be very promising concerning a circular bioeconomy.
Data availability
The data that support the findings of this study are available from the corresponding author, [D.C.], upon reasonable request.
References
U.S. Department of Energy renewable natural gas production: alternative fuels data center. afdc.energy.gov/fuels/natural_gas_renewable.html. Accessed 05 Mar 2022
Marelli L, Edwards R, Agostini A et al (2017) Solid and gaseous bioenergy pathways: input values and GHG emissions: calculated according to the methodology set in COM(2016) 767. EUR, Scientific and technical research series, vol 27215. Publications Office, Luxembourg
European Commission 2030 targets: Climate and energy framework. EU policy, strategy and legislation for 2030 environmental, energy and climate targets. https://ec.europa.eu/info/energy-climate-change-environment/overall-targets/2030-targets_en. Accessed 5 March 2022
European Environment Agency (2021) Share of energy consumption from renewable sources in Europe. https://www.eea.europa.eu/ims/share-of-energy-consumption-from. Accessed 5 March 2022
European Commission Energy and the Green Deal: a clean energy transition. https://ec.europa.eu/info/strategy/priorities-2019–2024/european-green-deal/energy-and-green-deal_en. Accessed 05 Mar 2022
Federal Environmental Agency Germany (2022) Climate | energy: renewable energies in figures. https://www.umweltbundesamt.de/en/topics/climate-energy/renewable-energies/renewable-energies-in-figures. Accessed 05 Mar 2022
Hussian AEM (2018) The role of microalgae in renewable energy production: challenges and opportunities. In: Türkoğlu M, Önal U, Ismen A (eds) Marine ecology - biotic and abiotic interactions. InTech
Mendonça HV, Assemany P, Abreu M et al (2021) Microalgae in a global world: new solutions for old problems? Renewable Energy 165:842–862. https://doi.org/10.1016/j.renene.2020.11.014
Roy UK, Radu T, Wagner JL (2021) Carbon-negative biomethane fuel production: integrating anaerobic digestion with algae-assisted biogas purification and hydrothermal carbonisation of digestate. Biomass Bioenerg 148:106029. https://doi.org/10.1016/j.biombioe.2021.106029
Passos F, Uggetti E, Carrère H et al (2014) Pretreatment of microalgae to improve biogas production: a review. Bioresour Technol 172:403–412. https://doi.org/10.1016/j.biortech.2014.08.114
Christenson L, Sims R (2011) Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol Adv 29:686–702. https://doi.org/10.1016/j.biotechadv.2011.05.015
Patwardhan SB, Pandit S, Ghosh D et al (2022) A concise review on the cultivation of microalgal biofilms for biofuel feedstock production. Biomass Conv Bioref. https://doi.org/10.1007/s13399-022-02783-9
Adey W (1982) Algae Turf Scrubber(4,333,263.)
Adey WH, Laughinghouse HD, Miller JB et al (2013) Algal turf scrubber (ATS) floways on the Great Wicomico River, Chesapeake Bay: productivity, algal community structure, substrate and chemistry(1). J Phycol 49:489–501. https://doi.org/10.1111/jpy.12056
Delgadillo-Mirquez L, Lopes F, Taidi B et al (2016) Nitrogen and phosphate removal from wastewater with a mixed microalgae and bacteria culture. Biotechnol Rep (Amst) 11:18–26. https://doi.org/10.1016/j.btre.2016.04.003
Liu J, Wu Y, Wu C et al (2017) Advanced nutrient removal from surface water by a consortium of attached microalgae and bacteria: a review. Bioresour Technol 241:1127–1137. https://doi.org/10.1016/j.biortech.2017.06.054
Mulbry W, Westhead EK, Pizarro C et al (2005) Recycling of manure nutrients: use of algal biomass from dairy manure treatment as a slow release fertilizer. Bioresour Technol 96:451–458. https://doi.org/10.1016/j.biortech.2004.05.026
Mulbry W, Kondrad S, Pizarro C et al (2008) Treatment of dairy manure effluent using freshwater algae: algal productivity and recovery of manure nutrients using pilot-scale algal turf scrubbers. Bioresour Technol 99:8137–8142. https://doi.org/10.1016/j.biortech.2008.03.073
Witarsa F, Yarberry A, May P et al (2020) Complementing energy production with nutrient management: anaerobic digestion system for algal turf scrubber biomass. Ecol Eng 143:105618. https://doi.org/10.1016/j.ecoleng.2019.105618
Torres A, Padrino S, Brito A et al (2021) Biogas production from anaerobic digestion of solid microalgae residues generated on different processes of microalgae-to-biofuel production. Biomass Conv Bioref. https://doi.org/10.1007/s13399-021-01898-9
Kumar V, Jaiswal KK, Tomar MS et al (2021) Production of high value-added biomolecules by microalgae cultivation in wastewater from anaerobic digestates of food waste: a review. Biomass Conv Bioref. https://doi.org/10.1007/s13399-021-01906-y
Kumar P, Bhattacharya A, Prajapati SK et al (2022) Anaerobic co-digestion of waste microalgal biomass with cattle dung in a pilot-scale reactor: effect of seasonal variations and long-term stability assessment. Biomass Conv Bioref 12:1203–1215. https://doi.org/10.1007/s13399-020-00778-y
Weber S, Grande PM, Blank LM et al (2022) Insights into cell wall disintegration of Chlorella vulgaris. PLoS ONE 17:e0262500. https://doi.org/10.1371/journal.pone.0262500
Wang M, Lee E, Dilbeck MP et al (2017) Thermal pretreatment of microalgae for biomethane production: experimental studies, kinetics and energy analysis. J Chem Technol Biotechnol 92:399–407. https://doi.org/10.1002/jctb.5018
Kothari R, Ahmad S, Pathak VV et al (2021) Algal-based biofuel generation through flue gas and wastewater utilization: a sustainable prospective approach. Biomass Conv Bioref 11:1419–1442. https://doi.org/10.1007/s13399-019-00533-y
Lee DH, Behera SK, Kim JW et al (2009) Methane production potential of leachate generated from Korean food waste recycling facilities: a lab-scale study. Waste Manag 29:876–882. https://doi.org/10.1016/j.wasman.2008.06.033
Rajput AA, Sheikh Z (2019) Effect of inoculum type and organic loading on biogas production of sunflower meal and wheat straw. Sustain Environ Res 29. https://doi.org/10.1186/s42834-019-0003-x
Hernández-Beltrán. U, J. L, H.-D. O et al. Insight into pretreatment methods of lignocellulosic biomass to increase biogas yield: current state, challenges, and opportunities. Appl Sci 9:3721. https://doi.org/10.3390/app9183721
Klin M, Pniewski F, Latała A (2020) Growth phase-dependent biochemical composition of green microalgae: theoretical considerations for biogas production. Bioresour Technol 303:122875. https://doi.org/10.1016/j.biortech.2020.122875
Dębowski M, Kisielewska M, Kazimierowicz J et al (2020) The effects of microalgae biomass co-substrate on biogas production from the common agricultural biogas plants feedstock. Energies 13:2186. https://doi.org/10.3390/en13092186
Du X, Tao Y, Liu Y et al (2020) Stimulating methane production from microalgae by alkaline pretreatment and co-digestion with sludge. Environ Technol 41:1546–1553. https://doi.org/10.1080/09593330.2018.1540665
Passos F, Solé M, García J et al (2013) Biogas production from microalgae grown in wastewater: effect of microwave pretreatment. Appl Energy 108:168–175. https://doi.org/10.1016/j.apenergy.2013.02.042
Mata-Alvarez J, Macé S, Llabrés P (2000) Anaerobic digestion of organic solid wastes. An overview of research achievements and perspectives. Biores Technol 74:3–16. https://doi.org/10.1016/S0960-8524(00)00023-7
Shahbaz M, Ammar M, Korai RM et al (2020) Impact of C/N ratios and organic loading rates of paper, cardboard and tissue wastes in batch and CSTR anaerobic digestion with food waste on their biogas production and digester stability. SN Appl Sci 2. https://doi.org/10.1007/s42452-020-03232-w
Mussgnug JH, Klassen V, Schlüter A et al (2010) Microalgae as substrates for fermentative biogas production in a combined biorefinery concept. J Biotechnol 150:51–56. https://doi.org/10.1016/j.jbiotec.2010.07.030
Singh J, Gu S (2010) Commercialization potential of microalgae for biofuels production. Renew Sustain Energy Rev 14:2596–2610. https://doi.org/10.1016/j.rser.2010.06.014
Parmar A, Singh NK, Pandey A et al (2011) Cyanobacteria and microalgae: a positive prospect for biofuels. Bioresour Technol 102:10163–10172. https://doi.org/10.1016/j.biortech.2011.08.030
Grömping M, Cheenakula D, Krafft S et al. (2022) Biohybride Systeme zur Verwertung von Phosphor aus Abwasser-BiohyreP. Project final report, Fachhochschule Aachen, Energy technology, Jülich, Germany
Öztürk SS (2021) Entwicklung eines Bioraffineriekonzeptes für phosphathaltige Algenbiomasse und die Bestimmung ihres Biogaspotenzials. Bachelor Thesis, Fachhochschule Aachen
Bischof F, Chwistek M, Schleyer C et al (2012) Biogas aus Algenbiomasse. Amberg, Germany
Marsolek MD, Kendall E, Thompson PL et al (2014) Thermal pretreatment of algae for anaerobic digestion. Bioresour Technol 151:373–377. https://doi.org/10.1016/j.biortech.2013.09.121
Passos F, Ferrer I (2014) Microalgae conversion to biogas: thermal pretreatment contribution on net energy production. Environ Sci Technol 48:7171–7178. https://doi.org/10.1021/es500982v
Alzate ME, Muñoz R, Rogalla F et al (2012) Biochemical methane potential of microalgae: influence of substrate to inoculum ratio, biomass concentration and pretreatment. Bioresour Technol 123:488–494. https://doi.org/10.1016/j.biortech.2012.06.113
González-Fernández C, Sialve B, Bernet N et al (2012) Thermal pretreatment to improve methane production of Scenedesmus biomass. Biomass Bioenerg 40:105–111. https://doi.org/10.1016/j.biombioe.2012.02.008
Cho S, Park S, Seon J et al (2013) Evaluation of thermal, ultrasonic and alkali pretreatments on mixed-microalgal biomass to enhance anaerobic methane production. Bioresour Technol 143:330–336. https://doi.org/10.1016/j.biortech.2013.06.017
Passos F, García J, Ferrer I (2013) Impact of low temperature pretreatment on the anaerobic digestion of microalgal biomass. Bioresour Technol 138:79–86. https://doi.org/10.1016/j.biortech.2013.03.114
Saleem M, Hanif MU, Bahadar A et al (2020) The effects of hot water and ultrasonication pretreatment of microalgae (Nannochloropsis oculata) on biogas production in anaerobic co-digestion with cow manure. Processes 8:1558. https://doi.org/10.3390/pr8121558
Chew KW, Yap JY, Show PL et al (2017) Microalgae biorefinery: high value products perspectives. Bioresour Technol 229:53–62. https://doi.org/10.1016/j.biortech.2017.01.006
Yadav G, Sen R (2018) Sustainability of microalgal biorefinery: scope, challenges, and opportunities. In: De S, Bandyopadhyay S, Assadi M et al. (eds) Sustainable Energy Technology and Policies. Springer Singapore, Singapore, pp 335–351
Bhattacharya M, Goswami S (2020) Microalgae – a green multi-product biorefinery for future industrial prospects. Biocatal Agric Biotechnol 25:101580. https://doi.org/10.1016/j.bcab.2020.101580
Figueroa-Torres GM, Pittman JK, Theodoropoulos C (2021) Optimisation of microalgal cultivation via nutrient-enhanced strategies: the biorefinery paradigm. Biotechnol Biofuels 14:64. https://doi.org/10.1186/s13068-021-01912-2
Wong MT (1984) Sewage sludge for cultivating freshwater algae and the fate of heavy metal at higher trophic organisms. IV. Heavy metal contents in different trophic levels. Arch Hydrobiol 100:423–430
Baicha Z, Salar-García MJ, Ortiz-Martínez VM et al (2016) A critical review on microalgae as an alternative source for bioenergy production: a promising low cost substrate for microbial fuel cells. Fuel Process Technol 154:104–116. https://doi.org/10.1016/j.fuproc.2016.08.017
Mathimani T, Pugazhendhi A (2019) Utilization of algae for biofuel, bio-products and bio-remediation. Biocatal Agric Biotechnol 17:326–330. https://doi.org/10.1016/j.bcab.2018.12.007
Guo P, Zhang Y, Zhao Y (2018) Biocapture of CO2 by different microalgal-based technologies for biogas upgrading and simultaneous biogas slurry purification under various light intensities and photoperiods. Int J Environ Res Public Health 15. https://doi.org/10.3390/ijerph15030528
Su Y, Mennerich A, Urban B (2012) Synergistic cooperation between wastewater-born algae and activated sludge for wastewater treatment: influence of algae and sludge inoculation ratios. Bioresour Technol 105:67–73. https://doi.org/10.1016/j.biortech.2011.11.113
Yu G, Peng H, Fu Y et al (2019) Enhanced nitrogen removal of low C/N wastewater in constructed wetlands with co-immobilizing solid carbon source and denitrifying bacteria. Bioresour Technol 280:337–344. https://doi.org/10.1016/j.biortech.2019.02.043
Fricke K (2009) Energieeffizienz kommunaler Kläranlagen. www.umweltbundesamt.de/sites/default/files/medien/publikation/long/3855.pdf. Accessed 05 Mar 2022
Acién FG, Gómez-Serrano C, Morales-Amaral MM et al (2016) Wastewater treatment using microalgae: how realistic a contribution might it be to significant urban wastewater treatment? Appl Microbiol Biotechnol 100:9013–9022. https://doi.org/10.1007/s00253-016-7835-7
Roostaei J, Zhang Y, Gopalakrishnan K et al (2018) Mixotrophic microalgae biofilm: a novel algae cultivation strategy for improved productivity and cost-efficiency of biofuel feedstock production. Sci Rep 8:12528. https://doi.org/10.1038/s41598-018-31016-1
Xiaogang H, Jalalah M, Jingyuan W et al (2022) Microalgal growth coupled with wastewater treatment in open and closed systems for advanced biofuel generation. Biomass Conv Bioref 12:1939–1958. https://doi.org/10.1007/s13399-020-01061-w
Al-Jabri H, Das P, Khan S et al (2021) Treatment of wastewaters by microalgae and the potential applications of the produced biomass—a review. Water 13:27. https://doi.org/10.3390/w13010027
DWA 33. Leistungsnachweis kommunaler Kläranagen: Entwicklung des Stromverbrauches. https://de.dwa.de/files/_media/content/06_SERVICE/Zahlen%20%7C%20Fakten%20%7C%20Umfragen/leistungsvergleich_2020_final.pdf. Accessed 5 March 2022
Craggs RJ (2001) Wastewater treatment by algal turf scrubbing. Water Sci Technol 44:427–433
HUBER SE Energy consumption of wastewater treatment plants: energy efficiency. www.huber.de/solutions/energy-efficiency/general/wastewater-treatment-plants/. Accessed 05 Mar 2022
FNR Bioenergy: Biogas. https://bioenergie.fnr.de/bioenergie/biogas. Accessed 5 March 2022
Fitzke B (2013) Behandlung von Abwässern und Reststoffen aus der Biogaserzeugung. www.dbu.de/media/0507100926018h6q.pdf. Accessed 05 Mar 2022
Nordrhein-Westfalen L (2021) Vollzugshinweise Nordrhein-Westfahlen für die Umsetzung der Düngeverordnung (DüV)
Fuchs W, Drosg B (2013) Assessment of the state of the art of technologies for the processing of digestate residue from anaerobic digesters. Water Sci Technol 67:1984–1993. https://doi.org/10.2166/wst.2013.075
Tampio E, Marttinen S, Rintala J (2016) Liquid fertilizer products from anaerobic digestion of food waste: mass, nutrient and energy balance of four digestate liquid treatment systems. J Clean Prod 125:22–32. https://doi.org/10.1016/j.jclepro.2016.03.127
Kirk DM, Gould MC (2019) Uses of solids and by-products of anaerobic digestion. farm-energy.extension.org/uses-of-solids-and-by-products-of-anaerobic-digestion/. Accessed 05 Mar 2022
Adey WH, Hackney L (eds) (1989) The composition and production of tropical marine algal turf in laboratory and field experiments, Washington DC
Singh M, Reynolds DL, Das KC (2011) Microalgal system for treatment of effluent from poultry litter anaerobic digestion. Bioresour Technol 102:10841–10848. https://doi.org/10.1016/j.biortech.2011.09.037
Markou G, Nerantzis E (2013) Microalgae for high-value compounds and biofuels production: a review with focus on cultivation under stress conditions. Biotechnol Adv 31:1532–1542. https://doi.org/10.1016/j.biotechadv.2013.07.011
Scragg A, Illman A, Carden A et al (2002) Growth of microalgae with increased calorific values in a tubular bioreactor. Biomass Bioenerg 23:67–73. https://doi.org/10.1016/S0961-9534(02)00028-4
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was part of the research project BiohyreP (Biohybrid systems for recycling of phosphorus from wastewater) and it was funded by the German Aerospace Center (DLR) (grant number 01DJ21007).
Author information
Authors and Affiliations
Contributions
Conceptualization: [Dheeraja Cheenakula, Kevin Hoffstadt, Simone Krafft]; Methodology: [Dheeraja Cheenakula]; Formal analysis and investigation: [Dheeraja Cheenakula]; Writing—original draft preparation: [Dheeraja Cheenakula]; Writing—review and editing: [Dheeraja Cheenakula, Kevin Hoffstadt, Simone Krafft, Diana Reinecke-Levi, Holger Klose, Markus Grömping]; Funding acquisition: [German Aerospace Center (DLR)]; Resources: [FH Aachen, Insitute NOWUM-Energy], Supervision: [Simone Krafft, Isabel Kuperjans, Markus Grömping]. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cheenakula, D., Hoffstadt, K., Krafft, S. et al. Anaerobic digestion of algal–bacterial biomass of an Algal Turf Scrubber system. Biomass Conv. Bioref. 13, 13605–13619 (2023). https://doi.org/10.1007/s13399-022-03236-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13399-022-03236-z