
Abstract
Background
Particulate matter (PM)2.5 is a concern for public health nowadays. Although few studies have reported the skin diseases associated with PM2.5, its effects on keratinocytes have yet to be elucidated.
Objective
The goal of this experiment was to analyze and identify the changes of gene expression in PM2.5-treated keratinocytes using RNA-sequencing (RNA-Seq) data.
Results
PM2.5-treated keratinocytes exhibited changes in cell cycle-related genes as well as genes involved in DNA replication, endoplasmic reticulum (ER) stress, intrinsic apoptosis, and immune response. A total of 669 genes showed changes in gene expression in PM2.5-treated keratinocytes, including 304 upregulated and 365 downregulated genes.
Conclusion
Unlike other studies investigating skin disorders associated with PM2.5, our study found the mechanism of apoptosis suppression in keratinocytes. The findings may provide a novel insight into the management of chronic skin diseases in relation to PM2.5.
Similar content being viewed by others

Introduction
Particulate matter (PM) is a mixture of particles and droplets of atmosphere. It is classified according to size. PM10 has a diameter less than 10 μm and PM2.5 is less than 2.5 μm in diameter. It contains polycyclic aromatic hydrocarbon (PAH), organic or inorganic materials and heavy metals. It comes from nature and exhaust gas of cars and factories. In many countries, increasing PM concentration in the air is the cause of serious health concerns. Several studies demonstrated that PM induced an inflammatory response in respiratory and cardiovascular disease (Kim et al. 2015). According to a study analyzing the effects of air pollutants on hospital admissions for respiratory, dermatological and ophthalmic diseases in children, increased PM10 and SO2 were associated with increased rates of hospitalization (Zhang et al. 2018). The studies demonstrated that air pollution threatens public health, especially involving children.
One of the functions of skin is to protect the body against exogenous materials that stimulate the immune system (Proksch et al. 2008). Because of its small size, PM2.5 penetrates into deeper layers of skin than other particles (Magnani et al. 2016). Although the mechanisms underlying PM-induced gene expression have been detected, the precise mechanisms remain unclear (Kim et al. 2015). Thus, the purpose of the present study was to elucidate the cytotoxicity and changes of gene expression in keratinocytes after PM2.5 exposure.
Proteins for cell formation are translated by RNA and released after processing in endoplasmic reticulum (ER). When ER function is perturbed, unfolded protein response (UPR) is activated to remove unfolded protein and to protect ER. Aberrant function of ER stress-induced UPR can cause many diseases. Therefore, these mechanisms are important for keeping normal health conditions (Sano and Reed 2013; D’Erme et al. 2015).
Recently, a bioinformatics analysis of microarray data was conducted for various diseases (Li et al. 2018). However, the effects of PM2.5 on skin using bioinformatics approaches have yet to be reported fully. We analyzed and identified the changes of gene expression in PM2.5-treated keratinocytes using RNA-sequencing (RNA-Seq) data.
Materials and methods
Cell culture and reagents
Neonatal epidermal keratinocytes (NHEK) were purchased from Gibco BRL, Life Technologies (Grand Island, NY, USA) and cultured in EpiLife (Gibco BRL, Grand Island, NY, YSA) with 60 μM calcium media, which included 0.2% bovine pituitary extract (BPE), 1 μg/mL recombinant human insulin-like growth factor-I, 0.18 μg/mL hydrocortisone, 5 μg/mL bovine transferrin, and 0.2 ng/mL human epidermal growth factor. These keratinocytes were used for experiments in early passages (up to passage 5). All cells were cultured at 37 °C with 5% CO2 and 95% relative humidity. NHEK showing 70% confluence were treated with 1 mg/mL of PM2.5 for 6 h.
Preparations of PMs
SRM 2786 (NIST, USA) (https://www-s.nist.gov/) (Gonzalez et al. 2016) was collected on Teflon® membrane filters using an ultrahigh-volume sampler. SRM 2786 were issued in 2011 as the first certified reference materials (CRMs) for fine particulate matter (Schantz et al. 2016). The PM was then brushed from filters and collected in a clean amber bottle with a Teflon®-lined cap. It was suspended in sterilized water. The PM comprised polycyclic aromatic hydrocarbons (PAHs), nitro-substituted PAHs (nitro-PAHs), polybrominated diphenyl ether (PBDE) congeners, hexabromocyclododecane (HBCD) isomers, sugars, polychlorinated dibenzo-q-dioxin (PCDD), dibenzofuran (PCDF) congeners, and inorganic constituents. Commercial PM2.5 were suspended in distilled water (DW) after measuring the weight and stored in 4 °C without light.
RNA extraction
Total RNA was isolated using TRIzol reagent (Invitrogen). RNA quality was assessed with Agilent 2100 bioanalyzer using the RNA 6000 Nano Chip (Agilent Technologies, Amstelveen, the Netherlands). RNA quantification was performed using the ND-2000 Spectrophotometer (Thermo Inc., DE, USA).
RNA-Seq analysis
QuantSeq 3’ mRNA-Seq reads were aligned using Bowtie2 (Banno et al. 2004). Bowtie2 indices were either generated from the genome assembly sequence or the representative transcript sequences for alignment with the genome and transcriptome. The alignment file was used to assemble the transcripts, estimate their abundances, and detect the differential expression of genes. Differentially expressed genes were determined based on counts derived from unique and multiple alignments covering BED files (Quinlan and Hall 2010). The RT (read count) data were processed via quantile normalization method based on EdgeR within R using Bioconductor (Gentleman et al. 2004). Gene classification was based on searches conducted using DAVID (http://david.abcc.ncifcrf.gov/) and Medline databases (http://www.ncbi.nlm.nih.gov/).
Gene enrichment analysis and clustering
We sorted out the list of significant changes in gene expression (> twofold, p value < 0.05). Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was executed using DAVID via biological process annotation (Huang et al. 2007; Kanehisa et al. 2016). Clustering was carried out using MeV 4.9 for the analysis of gene expression data.
Protein–protein interaction (PPI) network
The Search Tool for the Retrieval of Interacting Genes (STRING) database (http://string-db.org/) and Cytoscape software for determination of protein interactions were used to map PPI network (Szklarczyk et al. 2015). The cutoff criteria were a score of > 0.9 for a PPI network.
Statistical analysis
All experiments in this study were conducted in duplicate and independent experiments were repeated twice. The statistical significance of the data was determined via paired t test using GraphPad PRISM version 4.02 for Windows (GraphPad Software).
Result
PM2.5 altered changes of mRNA expression in keratinocytes
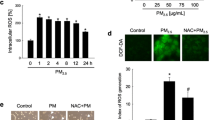
To investigate the altered gene expression induced by PM2.5, the keratinocytes were treated with PM2.5 at 1 mg/ml concentration for 6 h. RNA-Seq was performed in groups treated with and without PM2.5. Keratin-associated protein 2-3 (KRTAP2-3), heme oxygenase 1 (HMOX1), cytochrome P450 family 1 subfamily A member 1 (CYP1A1), and keratin 34 (KRT34) were upregulated more than tenfold while growth differentiation factor 15 (GDF15) was downregulated more than tenfold (Supplementary Figs. 1 and 2). IL-36γ, one of the psoriasis markers, was increased at 6 h than 0 h in PM2.5-treated keratinocytes6 (Supplement Fig. 3).
Profiling gene and clustering molecular responses to PM2.5
Clustered heat maps of 304 upregulated and 365 downregulated genes were generated (Fig. 1). To investigate the signals regulated by these changes, the GO enrichment analysis was performed via DAVID annotation. The top signals increased were related to RNA splicing, ER stress and cell proliferation. ‘Angiogenesis’ was also upregulated by PM2.5, although it was not detected in the top five signals. Downregulated genes were associated with the regulation of transcription from RNA polymerase 2 promoter and the response to ER stress.

Heatmap of gene expression patterns
In the KEGG pathway, genes involved in inflammatory response, metabolism and RNA processing were enriched. Downregulated genes associated with signal transduction and amino acid metabolism were also enriched (Fig. 2).

GO terms and KEGG pathway analysis of PM2.5-treated keratinocytes. Upregulated and downregulated GO terms and KEGG pathways
These data suggest that PM2.5 regulate genes associated with cell proliferation, inflammatory response, and decreased transcription activity in keratinocytes.
ER-induced apoptosis was inhibited via suppression of UPR in PM2.5-treated keratinocytes
RNA-Seq analysis showed that PM2.5 suppressed the genes related to UPR such as activating transcription factor 4 (ATF4) and DNA damage-inducible transcript 3 (DDIT3) also known as CHOP (Fig. 3). Increased GO terms of ‘regulation of cellular response to heat’ and decreased GO terms ‘response to endoplasmic reticulum stress’, ‘intrinsic apoptotic signaling pathway in response to endoplasmic reticulum stress’, and ‘PERK-mediated unfolded protein response’ suggest that PM2.5 was related to changes in cellular response after ER stress.

PM2.5 acts as an ER stress trigger (a) and suppressor of intrinsic apoptosis (b) in keratinocytes
Although PM2.5 induced ER stress, PM2.5 decreased HSPA5 known as binding immunoglobulin protein (BiP) that acted as an intrinsic apoptosis factor to regulate UPR. The levels of ATF4 and CHOP related to apoptosis after ER stress were decreased. Pro-apoptotic genes such as BCL-2-associated X (BaX) and B-cell lymphoma 2 (BCL-2) were not activated. The anti-apoptotic gene BBC3, known as p53-upregulated modulator of apoptosis (PUMA), was decreased after PM2.5 treatment (Sano and Reed 2013).
Tumor necrosis factor superfamily member 10 (TNFSF10), known as TNF-related apoptosis-inducing ligand (TRAIL), was upregulated by PM2.5 (Abdulghani and El-Deiry 2010). However, central proteins of apoptosis such as caspase-3 or caspase-8 were not activated after PM2.5 treatment. These results suggest that PM2.5 does not induce apoptosis in keratinocytes.
Altered gene expression induced immune response and inflammatory genes
To investigate the inflammatory response in keratinocytes, genes associated with inflammatory and immune response were analyzed. The levels of ‘positive regulation of innate immune response’ genes and ‘positive regulation of type 1 interferon production’ genes were increased by PM2.5. These results suggest that PM2.5 induces cytokines underlying immune response in keratinocytes.
PM2.5 changed proliferation, differentiation, and cell cycle-related factors in keratinocytes
PM2.5 increased keratinocyte proliferation and cell cycle, but decreased the expression of genes associated with differentiation. The levels of transforming growth factor (TGF)-α acting as keratinocyte proliferation factors were increased. Additionally, the level of differentiation factor GDF15 was decreased after PM2.5 treatment. Proliferation terms of GO analysis showed that ‘positive regulation of smooth muscle cell proliferation’, ‘angiogenesis’, ‘positive regulation of angiogenesis’, ‘cell proliferation’, and ‘positive regulation of cell proliferation’ were upregulated. In contrast, genes related to differentiation such as ‘cellular response to insulin stimulus’ and ‘cell maturation’ were decreased by PM2.5. These data suggest that PM2.5 induces cell proliferation by stimulating genes for growth factor secretion and cell cycle regulation.
Protein–protein interaction (PPI) network
Analyzed genes that constituted 632 nodes and 618 edges were mapped to the PPI network of STRING (Fig. 4). Screening of the PPI showed that the three highest-scoring edges were ‘STIP1-HSP90AA1’, ‘POLR3F-POLR2F’, and ‘CDC6-MCM3’. Among them, ‘STIP1-HSP90AA1’ and ‘POLR3F-POLR2F’ were modulated by HSPA8, while ‘CDC6-MCM3’ was regulated by POLR2F via PCNA. All genes in the module were upregulated by PM2.5.

PPI network and module analysis. a STRING was used to express 632 nodes and 618 edges. Upregulated genes are displayed in red, while downregulated genes are shown in green. b PPI showing the following three highest-scoring edges
Discussion
Since PM2.5 is known to be associated with respiratory disease generally, several studies have focused on the respiratory system (Hetland et al. 2004).
With increasing air pollution, we now know that PM2.5 not only affects the respiratory system, but also the skin and eyes. It is known that PM2.5 induces intrinsic apoptosis via ER and mitochondria. Contrary to our expectation, the results of RNA-Seq analysis showed that PM2.5 blocked the activation of ER stress-induced apoptotic genes such as ATF4 and DDIT3. When unfolding proteins are synthesized in ER, UPR is activated. Under normal conditions, transmembrane proteins (TP) such as protein kinase R (PKR)-like ER kinase (PERK), activating transcription factor 6 (ATF6), and inositol-requiring enzyme (IRE1) bind to BiP to suppress its activation. If unfolding polypeptides are increased, BiP stops binding with those proteins and the unfolding protein is eliminated via TP-mediated phosphorylation (Sano and Reed 2013; Wang et al. 2017).
UPR is known to regulate ER stress, and is decreased in proliferating cells. Suppression of apoptosis occurs in skin diseases such as cutaneous squamous cell carcinoma (cSCC) and psoriasis. Our data showed that PM2.5 suppressed apoptosis by decreasing the expression of ATF4 and DDIT3 genes. It has been demonstrated that UPR is significantly involved in the normal differentiation of keratinocytes. If UPR is not normally activated, keratinocytes would not normally differentiate. Downregulation of those genes is very similar to psoriatic gene expressions, such as UPR and apoptosis inhibition (Sugiura et al. 2009). These gene changes will result in keratinocyte proliferation without differentiation that is seen in the psoriasis epidermis.
PM2.5 consists of many types of PAH, nitro-PAH, and metal components (Kim et al. 2015). In addition to particles, their chemical composition can affect the inflammatory response and cell cycle (Abdel-Shafy and Mansour 2016). Exposure to nitro-PAH is known to induce mutagenesis and carcinogenesis (Rosenkranz and Mermelstein 1983). It increases cytokine and chemokine genes (Ovrevik et al. 2010). Especially, the exposure to 1-nitropyrene (1-NP) and 3-nitrofluoranthene (3-NF) increases the expression of CXCL8 and TNF-α. PAH is an organic compound with classical mutagenic effect on cellular homeostasis (Ewa and Danuta 2017).
Benzo[a]pyrene (B[a]P) is a well-known cellular irritant (Joo et al. 2015). It induces cyclooxygenase-2 (COX-2), CYP1A1, and CYP1B1 that are upregulated in response to the activation of aryl hydrocarbon receptor (AhR) (Piipari et al. 2000). The present study also showed an increased expression of COX-2, CYP1A1, and CYP1B1 in keratinocytes after treatment with PM2.5.
Psoriasis is a chronic skin inflammatory disease characterized by hyperkeratosis angiogenesis, and immune cell infiltration. It is believed that psoriasis is triggered by both genetic and environmental factors. Known environmental risk factors of psoriasis include UV exposure, medications interfering with individual immune system, smoking, obesity, alcohol intake, infections, and stress. Although the precise mechanism has yet to be elucidated, PM2.5 acts as an environmental risk factor that triggers or aggravates psoriasis at specific concentrations (Zeng et al. 2017). In the present study, the levels of IL-36γ and chemokine ligand (CXC)-8 were increased along with apoptosis suppression and increased angiogenesis. Another study demonstrated the upregulation of biomarkers for psoriasis including IL-36γ, S100 calcium-binding protein A (S100) 7, and S100A8, using PM2.5-treated NHEKs (Kim et al. 2017). Although the study suggested that PM2.5 elicited psoriasis-like epidermal changes, it did not elucidate the underlying mechanism.
In our study, the inhibition of ER stress-induced apoptosis was prominent, and was related to hyperkeratosis without differentiation into psoriasis. Gene expression results showed that PM2.5 downregulated the genes underlying intrinsic apoptosis such as ATF4, DDIT3, and caspase, which interfered with ER stress-induced UPR resulting in keratinocyte proliferation. Proliferation factors such as TGF-α and CXCL8 were increased and differentiation growth factors such as GRB7, GDF15 were decreased in keratinocytes. These changes reflect hyperkeratosis associated with decreased differentiation. Simply put, PM2.5 induce ER stress, although UPR related genes or apoptosis-related genes are downregulated. Also, PM2.5 is presumed to make similar patterns as psoriatic gene expression by increasing proliferation factors and decreasing differentiation factors in keratinocytes.
The increased expression of TNFSF10 and COX-2 indicates inflammation and formation of reactive oxygen species (ROS) by PM2.5 inducing ER stress linked to apoptosis. However, the suppression of genes associated with intrinsic apoptosis was blocked in this pathway. Along with increase in the expression of IL-36γ, the upregulation of inflammatory mediators such as CYP1A1 and CYP1B1 in response to the activation of AhR by PM2.5 induced hyperkeratosis.
In conclusion, the presenting study used RNA-seq analysis to determine the changes of gene expression in keratinocytes induced by exposure to PM2.5. PM2.5 suppressed UPR of keratinocytes which is related to differentiation of keratinocytes. Additionally, it induced angiogenesis and cytokines such as IL36γ and CXCL-8 known to be key markers of psoriasis. In contrast to proliferation-related factors, factors associated with differentiation were downregulated. The study findings suggest that PM2.5 might be an aggravator or inducer of inflammatory skin diseases such as psoriasis.
References
Abdel-Shafy HI, Mansour MS (2016) A review on polycyclic aromatic hydrocarbons: source, environmental impact, effect on human health and remediation. Egypt J Pet 25:107–123
Abdulghani J, El-Deiry WS (2010) TRAIL receptor signaling and therapeutics. Expert Opin Ther Targets 14:1091–1108
Banno T, Gazel A, Blumenberg M (2004) Effects of tumor necrosis factor-alpha (TNF alpha) in epidermal keratinocytes revealed using global transcriptional profiling. J Biol Chem 279:32633–32642
D’Erme AM et al (2015) IL-36gamma (IL-1F9) is a biomarker for psoriasis skin lesions. J Invest Dermatol 135:1025–1032
Ewa B, Danuta MS (2017) Polycyclic aromatic hydrocarbons and PAH-related DNA adducts. J Appl Genet 58:321–330
Gentleman RC et al (2004) Bioconductor: open software development for computational biology and bioinformatics. Genome Biol 5:R80
Gonzalez C, Choquette S, NIST (2016) Certificate of Analysis SRM2786 - Fine Atmospheric Particulate Matter (Mean Particle Diameter < 4 μm). National Institution of Standard Technology (NIST). https://wwws.nist.gov/srmors/certificates/2786.pdf
Hetland RB et al (2004) Release of inflammatory cytokines, cell toxicity and apoptosis in epithelial lung cells after exposure to ambient air particles of different size fractions. Toxicol Vitro 18:203–212
Huang DW et al (2007) DAVID bioinformatics resources: expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res 35:W169–W175
Joo DH et al (2015) Benzo (a) pyrene represses melanogenesis in B16F10 mouse melanoma cells. Mol Cell Toxicol 11:349–355
Kanehisa M, Sato Y, Kawashima M, Furumichi M, Tanabe M (2016) KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res 44:D457–D462
Kim KH, Kabir E, Kabir SJE (2015) A review on the human health impact of airborne particulate matter. Environ Int 74:136–143
Kim H-J et al (2017) Transcriptome analysis of airborne PM2. 5-induced detrimental effects on human keratinocytes. Toxicol Lett 273:26–35
Li HM, Xiao YJ, Min ZS, Tan C (2018) Identification and interaction analysis of key genes and microRNAs in atopic dermatitis by bioinformatics analysis. Clin Exp Dermatol 44:257–264
Magnani ND et al (2016) Skin damage mechanisms related to airborne particulate matter exposure. Toxicol Sci 149:227–236
Ovrevik J et al (2010) Differential effects of nitro-PAHs and amino-PAHs on cytokine and chemokine responses in human bronchial epithelial BEAS-2B cells. Toxicol Appl Pharmacol 242:270–280
Piipari R et al (2000) Expression of CYP1A1, CYP1B1 and CYP3A, and polycyclic aromatic hydrocarbon-DNA adduct formation in bronchoalveolar macrophages of smokers and non-smokers. Int J Cancer 86:610–616
Proksch E, Brandner JM, Jensen JM (2008) The skin: an indispensable barrier. Exp Dermatol 17:1063–1072
Quinlan AR, Hall IM (2010) BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics (Oxford, England) 26:841–842
Rosenkranz HS, Mermelstein R (1983) Mutagenicity and genotoxicity of nitroarenes: all nitro-containing chemicals were not created equal. Mutat Res 114:217–267
Sano R, Reed JC (2013) ER stress-induced cell death mechanisms. Biochem Biophys Acta 1833:3460–3470
Schantz MM, Cleveland D, Heckert NA, Kucklick JR, Leigh SD et al (2016) Development of two fine particulate matter standard reference materials(< 4 μm and < 10 μm) for the determination of organic and inorganic constituents. Anal Gioanal Chem 408(16):4257–4266
Sugiura K et al (2009) The unfolded protein response is activated in differentiating epidermal keratinocytes. J Invest Dermatol 129:2126–2135
Szklarczyk D et al (2015) STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res 43:D447–D452
Wang J, Lee J, Liem D, Ping P (2017) HSPA5 Gene encoding Hsp70 chaperone BiP in the endoplasmic reticulum. Gene 618:14–23
Zeng J, Luo S, Huang Y, Lu Q (2017) Critical role of environmental factors in the pathogenesis of psoriasis. J Dermatol 44:863–872
Zhang YL, Zhang H, Yi JP, Zhang JJ, Dai XR (2018) Effect of air pollution on hospital admissions of respiratory, dermatological, ophthalmic diseases in a coastal city, China. Glob Environ Health Saf 2:2
Acknowledgements
This work was supported by the National Research Foundation of Korea Grant (NRF), funded by the Korean Government (NRF-2016R1C1B1014180) and Korea University Grants.
Author information
Authors and Affiliations
Contributions
Every author has made substantial contributions to this manuscript by making the conception and design of the study (SJW, KJ, KMG), acquisition of data (KJH, PTJ), and drafting and revision of this article (SSW, RHJ).
Corresponding authors
Ethics declarations
Conflict of interest
All of the authors declare no conflicts of interest.
Ethical approval
All the experimental procedures were performed in accordance with the ethical standards as recommended by the International Committee of Medical Journal Editors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

{kind=link}
Cite this article
Kim, J.H., Son, J.W., Kim, J. et al. Particulate matter (PM)2.5 affects keratinocytes via endoplasmic reticulum (ER) stress-mediated suppression of apoptosis. Mol. Cell. Toxicol. 16, 129–137 (2020). https://doi.org/10.1007/s13273-019-00065-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13273-019-00065-6