
Abstract
Tidal marshes provide numerous ecosystem services, but are threatened by recent increases in global sea level rise (SLR). Marsh restoration and creation are important strategies for mitigating marsh loss, restoring ecosystem services, increasing coastal community resilience, and providing much needed habitat for threatened species. Dredged material resulting from navigation channel maintenance can provide a substrate for these restoration projects. Few studies, however, have addressed the sustainability of these marshes. The Paul S. Sarbanes Ecosystem Restoration Project at Poplar Island, where fine-grained, nutrient-rich dredged material from upper Chesapeake Bay is being used to create > 302 ha of tidal marshes, provides a case study. The low supply of inorganic sediment is counteracted by abundant nutrients, stimulating high rates of organic matter production and elevation change. Using > 10 years of data from 39 surface elevation tables, we found that the mean low marsh rate of elevation change (7.7 ± 3.21 mm year−1) was double the mean high marsh rate (3.6 ± 0.47 mm year−1) and exceeded the natural reference marsh (3.0 ± 2.28 mm year−1) and relative SLR (5.7 mm year−1). By stimulating organic matter production, the high nutrient substrate appears to offset the low inorganic sediment inputs in mid-Chesapeake Bay. Spatial variability was correlated with initial elevation, but was also influenced by local factors that may affect sediment redistribution within the marshes.
Similar content being viewed by others
Introduction
Tidal marshes provide important ecosystem services in coastal areas, but are under threat globally from a combination of direct anthropogenic activities and sea level rise (SLR) (Fagherazzi et al. 2020).While there is potential for tidal marsh expansion in some areas as SLR promotes landward marsh migration, net losses are expected to occur in the long term if the rate of SLR continues to increase (FitzGerald and Hughes 2019). The importance of tidal marshes in protecting coastal land from flooding and storm damage (Gedan et al. 2011), in addition to their value for habitat, carbon sequestration, and provision of other ecosystem services, has recently focused attention on tidal marsh conservation, restoration, and creation (Waltham et al. 2021).
Marsh restoration and creation projects using dredged material from navigation channel maintenance are increasingly being considered to reduce or offset the loss of natural habitat and ecosystem services associated with net marsh loss (Suedel et al. 2022; Welch et al. 2016; Yozzo et al. 2004). A range of approaches to marsh creation have been employed, including unconfined, semi-confined, and confined placement of dredged material, with minimally to highly engineered marsh designs. The ability of these new marshes to keep pace with SLR through elevation change, however, and the design factors which promote it are not yet well understood.
Elevation change is the result of complex feedbacks between vegetation and abiotic factors such as relative SLR (RSLR), the combined effect of local SLR and vertical land motion (VLM) (Wöppelmann and Marcos 2016), and the local supply of inorganic sediment, resulting in the accumulation of organic and inorganic matter on the marsh surface and within the root zone (Cahoon et al. 2021; Kirwan and Megonigal 2013; Morris et al. 2002). In many coastal areas, inorganic sediment inputs are low, and elevation change relies primarily on the deposition of organic matter, with elevation change proportional to rates of macrophyte biomass production (Morris and Sundberg 2024). Greenhouse experiments (Darby and Turner 2008; Mendelssohn and Morris 2002) and fertilization studies conducted in natural marshes (Deegan et al. 2012; Morris et al. 2013) demonstrate that biomass production by Spartina alterniflora, a dominant species in many tidal marshes, is regulated largely by nutrient availability, especially nitrogen (N), and marsh platform elevation (Morris et al. 2002). While nutrient availability determines the maximum potential biomass production, there is an optimal elevation, above or below which biomass production is limited by other factors, such as competition at higher elevations (Bertness 1991) and hypoxic soil conditions and high soil sulfide concentrations at lower elevations (Mendelssohn and Morris 2002), in addition to other anthropogenic toxins found in coastal lands and seascapes. Soil fertility levels which enhance biomass production can also enhance elevation gain in tidal marshes through the increased deposition of organic matter (Anisfeld and Hill 2012; Davis et al. 2017).
In addition to biomass production, other potential factors which may influence rates of elevation change include geomorphic dynamics such as exogenous sediment inputs and internal sediment redistribution between marsh platforms, tidal channels, and mudflats (Fagherazzi et al. 2004). In created marshes, many of these factors can be influenced through the choice of substrate (e.g., grain size), initial elevation, and channel, pond, and inlet geometry. Channel morphology, such as cross-section width and depth, affects the ebb and flood dominance of a tidal marsh and sediment dynamics (Fleri et al. 2019). Restored tidal marshes evolve toward a steady state by changing the initial geometry of channels and ponds, affecting both sediment transport and delivery to the adjacent marsh. The tidal channel network plays a key role in marsh hydrology and sediment distribution in response to seasonal and climatic changes (Nardin et al. 2021; Taddia et al. 2021). Ecological changes in the Poplar Island marshes observed by aerial monitoring include low marsh migration into higher elevations (Nardin et al. 2021; Windle et al. 2023), likely as a consequence of changing water levels and marsh hydrology, or localized marsh elevation deficits at construction. While studies such as these are providing feedback for design of created marshes, performance gauged by elevation change has not been well documented.
There are now examples of tidal marshes created with dredged material in many coastal areas of the USA (Berkowitz et al. 2017; Marcus 2000; Suedel et al. 2022; Yozzo et al. 2004) and other countries (Baptist et al. 2019), but these often lack long-term monitoring programs to assess performance and resilience to RSLR. An example of this type of project with a long-term monitoring program is the Paul S. Sarbanes Ecosystem Restoration Project at Poplar Island (hereafter, Poplar Island) located in mid-Chesapeake Bay, MD. It was conceived as a beneficial use of dredged material to restore an historic island which had nearly completely eroded due to natural and anthropogenic causes. The project includes the creation of 302 ha of tidal marsh habitat and 335 ha of upland habitat, which is being developed incrementally over a period of approximately 30 years. The substrate is fine-grained, N-rich dredged sediment from upper Chesapeake Bay (Cornwell et al. 2020). An extensive, long-term monitoring program includes elevation change monitoring using a network of surface elevation tables (SETs) (Cahoon et al. 2002). Installation of SETs commenced in 2008 (Table S1).
The Poplar Island SET network was designed to monitor elevation change with the goal of evaluating the sustainability of marshes constructed with upper Chesapeake Bay dredged material under current and future RSLR. An additional goal was to help identify patterns and drivers of spatial variability in rates of elevation change, to inform future marsh designs at Poplar Island and similar beneficial use projects.
In this study, we examined trends in elevation change in seven marshes at Poplar Island, ranging in age from 5 to 20 years, using a subset of data from the SET network. We compared rates in the Poplar marshes with the local rate of RSLR and with a nearby natural reference marsh. We further analyzed the accretion data for indications of important factors which may be manipulated in marsh design, including initial elevations, soil characteristics, and proximity to sediment sources (external and internal). Here we report on the results of these analyses and provide insights to inform future marsh restoration efforts.
Methods
Site Description
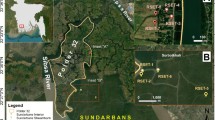
Poplar Island is located in mid-Chesapeake Bay, 3.2 km northwest of Tilghman, MD, USA (Fig. 1). The site experiences a mean diurnal tide range of 0.47 m (NOAA Tides and Currents, station ID 8572271) and a mean surface salinity range (1985–2018) of 10.1–12.5 ppt (U. S. Environmental Protection Agency 2019). Mean annual total suspended solid (TSS) concentrations at Poplar Island are low, < 20 mg L−1 (Staver et al. 2020), typical for the mid-Chesapeake Bay (Yunus et al. 2021). The local rate of RSLR is 5.7 mm year−1 (1995–2022, Morris and Staver, in review), based on water levels in Annapolis, MD (NOAA station ID 8575512), located approximately 25 km northwest of Poplar Island. Water levels at Poplar Island and Annapolis are similar and highly correlated (Kent 2015).

Location of Poplar Island (top right) and Tar Island reference marsh (bottom right) within Chesapeake Bay, MD, USA. Yellow points represent surface elevation tables. Map credit (left), Tracey Saxby and Kate Boicourt, Integration and Application Network (ian.umces.edu/media-library); photo credit (middle and right), Landsat
Data for the study came from seven tidal marsh units (cells) developed between 2003 and 2018 (Fig. 1). The substrate in six cells is upper Chesapeake Bay, fine-grained dredged material. In the seventh cell (Cell 4D), the substrate is locally dredged sand, a low nutrient substrate. The marsh platforms were designed with elevations to provide 80% low marsh (LM) and 20% high marsh (HM), planted with S. alterniflora and Spartina patens, respectively (USACE 1996, p. 6–1). The marsh substrate in the dredged material cells (to a depth of approximately 2 m, after consolidation) is fine-grained sediment resulting from maintenance dredging of the upper Chesapeake Bay navigation channels (Cornwell and Owens 2011), providing very high fertility (Cornwell et al. 2020). The dredged material was de-watered over a period of 2–6 years, after which tidal creeks were incised into the consolidated material using heavy equipment and a tidal inlet structure was constructed to permit vehicle passage along the perimeter dike. Tidal exchange was established approximately 6 months prior to planting, providing an opportunity for neutralization of soil acidity resulting from soil oxidation during the drying process (Demas et al. 2004). The graded elevation ranges (Supplementary Table S1) in the older marshes (e.g., Cell 3D) were 0.01–0.19 m North American Vertical Datum of 1988 (NAVD88) (0.05–0.23 m mean sea level (MSL)) and 0.19–0.40 m NAVD88 (0.23–0.45 m MSL) in the LM and HM, respectively (personal communication, Catherine Perkins and Seth Keller, U. S. Army Corps of Engineers, Baltimore District). In the more recently constructed marshes, elevation ranges were higher, with the LM lower limit at 0.04 m NAVD88 (0.08 m MSL), the LM–HM break line at 0.22–0.34 m NAVD88 (0.26–0.39 m MSL), and the HM upper limit at 0.40–0.55 m NAVD88 (0.45–0.60 m MSL). Vegetation was established by manually planting nursery-raised S. alterniflora and S. patens stock.
Each marsh is surrounded by sand dikes extending approximately 3 m above MSL, and tidal exchange occurs through inlets with a variety of designs. Three of the cells (Cells 1A, 1B, and 1C) are connected via shallow lateral channels designed to facilitate fish passage, but exchange through these channels is less than 5% of tidal exchange at the inlet (Fleri et al. 2019). Upland drainage into the marsh cells is minimal, limited to the area of the surrounding dikes.
SET Installation and Monitoring
SET Experimental Design
The SET data analyzed in this study were from a subset (n = 39) of the Poplar Island SET network (n = 51) and included deep-rod SETs (Cahoon et al. 2002) installed in LM areas dominated by S. alterniflora (n = 35), in a HM area of Cell 1A dominated by S. patens (n = 3), and on a mudflat in Cell 1B (n = 1, Fig. 1). Three of the 35 LM SETs were located in a nearby natural marsh which served as a reference (Tar Island, Fig. 1). In Cells 3D, 1A, and 5AB, triplicate SETs were located in different areas of the cell to evaluate factors which could potentially affect elevation change, including elevation, proximity to the tidal inlet (source of external sediment), and proximity to the nearest tidal creek (source of internal sediment). In Cell 5AB, two groups of three were located in the low marsh interior in areas of the cell which had different initial platform elevations, 0.22 m and 0.32 m NAVD88, specifically to test the effect of initial elevation on the rate of elevation change. In each group, the SETs were located within 5 m of the edge of the nearest tidal creek. All other cells had three SETs, all of which were located in the low marsh interior, with the exception of the one on an unvegetated mudflat in Cell 1B.
SET Installation
Deep-rod SETs were installed in seven marshes beginning in 2008, with stainless steel rods driven to refusal (Table 1). In Cells 1A, 1C, 3A, and 5AB, SETs were installed at the time of vegetation establishment. In Cells 1B, 3D, and 4D, installation occurred 3–5 years after vegetation was planted, and SETs were installed in the reference marsh in 2013. Each SET was surrounded by a permanent wooden frame to protect the measurement surface from foot traffic. The 1.8 m × 1.8 m square frame, consisting of four corner posts connected by 0.05 m × 0.10 m × 1.8 m boards elevated approximately 0.4 m above the marsh surface, also provides a structure to support the platform used to perform measurements. Each SET consists of a receiver connected to the anchoring rods. The elevation of each SET receiver was measured with real-time kinematic (RTK) GPS surveys conducted in 2010 and again in 2021.
SET Monitoring
SETs were monitored 3–6 months after installation and annually thereafter, usually in late winter, except in Cell 5AB where quarterly measurements were made for the first year, followed thereafter by annual measurements. A 2-m-long aluminum platform was placed across the wooden frame to provide access to the SET mark. Measurements were made using a T-shaped detachable arm, with nine pins on each arm, in two orientations. Pins were lowered until they touched, but did not impinge on, the sediment surface. Surface elevation (Z NAVD88) at each pin was determined as \(Z=({\text{Zm}}+B)-{\text{Pl}}+{\text{Ph}}\), where Zm is the height of the mark (m NADV88), B is the height of the SET bar (m), Pl is the total pin length (m), and Ph is the pin length above the bar (m). Surface elevation change was estimated as the mean (n = 9) slope of the regression of annual surface elevation for each subplot.
Marker Horizons and Sediment Characterization
Marker horizons (MH) were installed at all SET sites at various times throughout the study period, primarily to characterize soil accumulating on the marsh surface. The MHs consisted of a bright white, clay material which was surface applied. To estimate surface accretion rates for this study, one MH was sampled at each SET site. The year that the sampled MH was installed varied with site (Table 1), but all sites were sampled in 2021 to estimate surface accretion rates for this study. At each SET, one sediment core was collected from the MH site to a depth below the MH using a knife or a specially fabricated piston corer. The depth to the MH was recorded, and the soil layer above the MH was collected for analysis of soil organic matter (combustion at 450 °C for 4 h). Accretion rates (AR) were estimated as \({\text{AR}}=D/T\) where D is depth and T is time since MH placement.
The change in N availability over time was analyzed using porewater ammonium (NH4+) data collected in the LM of Cell 3D between 2005 and 2014. Porewater NH4+ concentrations were determined for the root (0–12 cm) and deep (13–46 cm) zones of the soil profile using porewater equilibrators (Hesslein 1976), according to Cornwell et al. (2020). Equilibrators were processed immediately upon retrieval in the field and samples were frozen for later colorimetric NH4+ analysis (Parsons et al. 1984).
Statistical Analysis
For trend analysis, rates of elevation change were determined for each SET subplot (through 2021) by fitting a linear model (SAS© 9.4 Proc Reg) to the SET data consisting of the mean elevation of nine pins at each time point. The regression slope was taken as the rate of elevation change. Rates of elevation change reported for individual SETs (Table 1, Fig. 3) represent the mean of the four subplot rates.
Relationships between elevation change rates and potential controlling variables were evaluated using Pearson correlation analysis (SAS© 9.4). In these analyses, each subplot was considered a replicate and it was assumed that the subplots were independent and not replicates of the elevation at a single point. This increased the degrees of freedom and statistical power, but increased the variance and decreased the R2. Variables included the mean elevation of the marsh surface (cm NADV88) at the first measurement (Z0), linear distance of the SET benchmark to the nearest tidal creek (m), distance along the thalweg of the tidal creek from a point adjacent to the SET to the tidal inlet through the perimeter dike (m), tidal creek order (1–3 = smallest to largest), MH accretion rate (mm year−1), and the organic content (%) of the soil layer accumulated above the MH. The difference between mean SET rates of elevation change in two areas of Cell 5AB, differing in starting elevation, was tested with Student’s t test in SigmaPlot 15.0.
Results
Trends in Elevation Change
Rates of elevation change in the Poplar Island marshes ranged from − 8.5 to 15.3 mm year−1 (Table 1, Fig. 2), and mean rates (± standard deviation) of elevation change for all marshes except Cells 4D and 3D exceeded rates of both the natural reference marsh (TI, 3.0 ± 2.28 mm year−1) and the rate of RSLR (5.7 mm year−1). Mean rates of LM elevation change (excluding creek bank and mudflat sites) for each marsh varied by a factor of almost 3, ranging from 5.4 ± 0.15 mm year−1 (Cell 4D, developed in 2003, sand substrate) to 13.5 ± 2.62 mm year−1 (Cell 1B, developed in 2012; Fig. 3a). The mean LM rate for Cell 1A (7.7 ± 1.95 mm year−1, n = 6) was double the mean rate for the HM sites (3.6 ± 0.47 mm year−1, n = 3; Fig. 3b), which showed little variability. The mean rate for the creek bank (− 3.4 ± 4.39 mm year−1, n = 3) and mudflat (− 7.0 mm year−1, n = 1) sites were negative, primarily due to bank and surface erosion, respectively (Fig. 3b). The two groups in Cell 5AB located in low and high areas of the LM zone had mean rates of 13.2 ± 2.52 mm year−1 and 7.8 ± 2.20 mm year−1, respectively, and were significantly different (Student’s t test, df = 4, t = 2.811, P = 0.0482).

Spatial variation in rates of elevation change in the restored Poplar Island marshes. Green and red symbols represent positive and negative rates, respectively, and symbol size indicates scale of rate

a Mean rates of elevation change in the restored Poplar Island marshes (x-axis labels refer to marsh “cell”) and Tar Island (TI) reference marsh and b mean rates for low marsh (LM, n = 23), high marsh (HM, n = 3), creek bank (CB, n = 3), mudflat (MF, n = 1), and Cell 5AB LM low (5AB-L) and high (5AB-H) areas, respectively. Boxes represent 25th to 75th percentile, gray bars represent median, black bars represent mean, whiskers represent 5th and 95th percentiles, and circles represent outliers. Sea level rise is represented by the red line
Similar patterns in cumulative elevation change (Fig. 4a) were observed where LM SETs were installed when the marsh was constructed (Cell 1A, constructed 2009) or 4 years later (Cell 3D, constructed 2005), with a period of higher rate of change for several years followed by slower rates. A much younger marsh (Cell 3A) showed the high initial rate, but had not entered the period of slowing. The mean rate in Cell 3D (5.5 ± 2.84 mm year−1), where initial graded elevations were lower, was less than the mean rates in younger Cell 1A (7.6 ± 1.95) or 3A (10.5 ± 3.19; Fig. 3a), where initial elevations were higher. During a common period of measurement (2013–2021), cumulative elevation change in two Poplar Island marshes (Cells 3D and 1A) exceeded the natural reference marsh (Tar Island, Fig. 4b).

Mean (± standard error) cumulative elevation change in a three Poplar Island marshes, Cells 3D (n = 6), 1A (n = 6), and 3A (n = 3), from the date of surface elevation table (SET) installation to 2021 and in b Poplar Island Cells 3D and 1A and the natural reference marsh, Tar Island (n = 3), over a common period of measurement (2013–2021). The Poplar Island marshes were constructed in 2005 (Cell 3D), 2009 (Cell 1A), and 2015 (Cell 3A). SET installation occurred in the years Cells 1A and 3A were constructed, but 4 years after construction in Cell 3D
Marker Horizons and Sediment Characteristics
Rates of surface accretion at Poplar Island determined using marker horizons ranged from 0.0 to 7.5 mm year−1; TI rates ranged from 1.6 to 11.0 mm year−1 (Table 1). Organic matter content in the sediment layer above the marker horizons ranged from 1.4 to 18.9%.
Porewater NH4+ declined rapidly in both near-surface (< 12 cm) and deeper horizons in Cell 3D (Fig. 5). Concentrations in the near-surface horizons decreased to < 300 µmol NH4+-N L−1 in the first 2 years, but were sustained by both remineralization of organic material and slow upward diffusion from deeper soils. The initial high concentrations in deeper horizons persisted at concentrations > 1000 µmol NH4+-N L−1 for the initial 2 years, but decreased steadily afterward to concentrations similar to near-surface horizons. The initial burst of exceptionally high above-ground biomass (Staver et al. 2020) coincided with the rapid depuration of soil NH4+.

Mean porewater ammonium (NH4+) concentrations in the top 12 cm (“Shallow,” open circles) and 13–46 cm (“Deep,” green triangles) of the soil profile, respectively, in the Cell 3D marsh at Poplar Island, MD. The error bars represent standard errors
Correlations
A correlation matrix (Table 2) shows important correlations between variables, including between the rate of elevation change (slope) and variables with potential management implications. There was a highly significant negative correlation between the rate of elevation change (slope) and MH age (Fig. 6a), and between elevation change and % organic matter in the layer above the MH (marker horizon organic content (MHOC), Fig. 6b). There were significant positive correlations between MHOC and MH age, between MHOC and surface accretion rate (MHAC, Fig. 6c), and between the rate of elevation change and the distance from the nearest creek (Cr Dist, Fig. 6d).

Regressions of important variables related to elevation change in the Poplar Island, MD, restored marshes. a The rate of elevation change versus marker horizon (MH) age. b The rate of elevation change versus percent organic matter above the MH (MH %OM). c The depth of the layer above the MH versus MH %OM. d The rate of elevation change versus the distance to the nearest tidal channel
Other notable significant correlations occurred between MHOC and SET installation date (Pearson correlation coefficient, PCC = − 0.63388, P < 0.0001) and between MHOC and marker horizon age (PCC = 0.63388, P < 0.0001, Fig. 6b). Creek order was negatively correlated with MHOC (PCC = − 0.47550, P < 0.0001). Also of note, the rate of elevation change was not significantly correlated with MH accretion rates (PCC = − 0.0489, P = 0.787, n = 33), and a second-order polynomial regression fitted to the LM slopes was significant (R2 = 0.0313, P = 0.012; Fig. 7).

Rates of elevation change versus the elevation of the marsh surface (Z0) at the beginning of the surface elevation table (SET) data record in the restored tidal marshes at Poplar Island, MD. Red dots represent data from SETs located in the low marsh, fitted with a polynomial regression (R2 = 0.0313, F = 6.504, P = 0.012). High marsh (open circles), mudflat (black downward pointing triangles), and creek bank (blue upward pointing triangles) data are shown for reference, but not included in the regression. Statistical analysis is shown in Supplementary Table S3
Discussion and Conclusions
Trends in Elevation Change
Developing strategies to enhance elevation gain in restored marshes will be essential to their sustainability in the face of accelerating RSLR, and to justify future investment in marsh restoration. The slow trajectory of some restored marshes toward maturity and maximum biomass production, which can take years to decades depending on the substrate (Craft et al. 1999, 2003), presents a challenge where rates of RSLR are high and accelerating rapidly. In the Chesapeake Bay, sea level is likely to rise at 0.6–1.1 m by the end of this century (Boesch et al. 2023), a rate of 7.7–14.2 mm year−1 for the remainder of the century, which is substantially higher than the current rate of 5.7 mm year−1.
This study has shown that rapid development of the restored marshes at Poplar Island, where rates of biomass production are high within a year of construction (Staver et al. 2020), has resulted in high rates of elevation gain, with most LM rates (Table 1) exceeding both the mean rate in the natural reference marsh (3.0 ± 2.28 mm year−1) and the current rate of RSLR (5.7 mm year−1). The LM rates also compare favorably with rates reported for other restored marshes, for example marshes restored by tide gate removal (Anisfeld et al. 2016) or created by excavation of upland (Kamrath et al. 2019; Table 3). In the former, plant roots and rhizomes were not colonizing unvegetated substrate, as is the case in newly created marshes. This rapid growth of roots and rhizomes adds to the sediment volume, driving positive elevation change (Morris 2007). In the case of excavation, exposed subsoil may be unfavorable for plant growth due to low nutrient content or other biogeochemical factors (Broome and Craft 2019). Higher rates reported by Poppe and Rybczyk (2021) resulted from dike lowering and breaching in a site located at the mouth of the Stillaguamish River, WA, where there was an ample supply of riverine sediment and a ~ 1-m elevation deficit, resulting in high rates of accretion and elevation change. Rates were expected to decline as elevation in the restored marshes approached equilibrium with sea level.
From this study, it appears that with appropriate management to avoid problems associated with acid sulfate soils (Cornwell et al. 2020), and appropriate marsh design, the use of fine-grained dredged material as a substrate for tidal marsh restoration results in marshes that are resilient to RSLR in the first one to two decades of development, supporting the use of this type of substrate for future projects. Differences between and within marshes provide some insights which may help guide the design of future projects.
Drivers of Spatial Variability
Although rates of elevation change compare favorably with RSLR, spatial variability is high (Fig. 2, Table 1), and mean rates for the two oldest marshes, Cells 3D (5.5 ± 2.84 mm year−1) and 4D (5.4 ± 0.15 mm year−1), are near or below the rate of RSLR. The rate of elevation change decreased with MH age (Table 2, Fig. 6a) and with increasing organic matter content above the marker horizon (Fig. 6b). Based on linear regression, a MH age of 4 gains 13.4 mm year−1 while a MH age of 15 gains 5.3 mm year−1. Similarly, at 2% MHOC, the marsh gains 9.4 mm year−1, while at 15% MHOC, it gains only 5.8 mm year−1. Older marshes (with older MHs) have accumulated more organic matter. As the buildup of sediment organic matter progresses, the sediment organic matter content rises, but the rate of both surface accretion and elevation gain slows down.
Several factors may contribute to these observed relationships. First, marsh platform elevation specifications in the oldest marshes (Cells 3D and 4D) were slightly lower than those in the younger marshes. The specified LM ranges were 0.01 to 0.19 m NAVD88 in Cell 3D (developed in 2005) and 0.04 to 0.22, 0.31, or 0.34 m NAVD88 in subsequent marshes (Supplemental Table S1). These small differences in elevation capital (Cahoon and Guntenspergen 2010) appear to be important in microtidal marshes in locations such as mid-Chesapeake Bay. The two older marshes appear to be suffering inundation stress, reflected in a distinct microbial community in Cell 3D compared with other Poplar Island marshes (Kim et al. 2023), which may be limiting macrophyte production and resulting in lower rates of elevation change. In addition, our analysis of data from two zones in Cell 5AB showing significantly different rates of elevation change (Fig. 3b) indicates that initial elevation plays a key role, consistent with marsh equilibrium theory (Morris et al. 2002).
Secondly, the initially high soil N concentrations have declined as the marshes have matured (Fig. 5), and may also contribute to the limitation of primary production after the first few years and lower rates of elevation change in the older marshes. In addition, the lowest rate of elevation gain occurred in Cell 4D, the marsh with a sand substrate.
Fertilization studies in natural marshes have demonstrated that high soil fertility can translate into enhanced rates of elevation gain (Anisfeld and Hill 2012; Morris and Sundberg 2024). This suggests that a high nutrient substrate may also enhance elevation gain in restored marshes, especially autochthonous marshes, helping to compensate for a low supply of inorganic sediment. The newly constructed tidal marshes at Poplar Island support this hypothesis. While initial porewater NH4+ concentrations were not collected at each SET location, in general, the Poplar Island marshes have very high initial levels of N availability (Cornwell et al. 2020), stimulating rapid colonization and high levels of primary production by the dominant vegetation, S. alterniflora and S. patens, within 2 years (Staver et al. 2020). In future studies of newly constructed marshes, it is recommended that initial porewater nutrient concentrations are measured at each SET location, at a consistent point in the growing season (to avoid seasonal variability), as part of the adaptive management monitoring program.
Thirdly, this is consistent with the marsh development model that marsh elevation gain is proportional to the expansion of belowground biomass, which proceeds in time much like an old field succession (Morris and Staver, in review). Newly planted marshes fill the unvegetated sediment with new roots and rhizomes, expanding the sediment volume and increasing surface elevation (Nyman et al. 2006; Cahoon et al. 2021). It may take several years before root and rhizome production, turnover, and diagenesis reach an equilibrium, at which point organic matter will continue to accumulate, but at a slower rate (Morris et al. 2002; Krauss et al. 2017). As a result, rates of elevation change may be expected to slow as these marshes mature, as has been observed in restored mangrove habitats (Castillo et al. 2022; Krauss et al. 2017). Lower rates of elevation gain in older marshes in this study, as well as lower rates in marshes where SETs were not installed at the time the marsh was developed, suggest that the initial phase of marsh vegetation growth is an important factor in the high rates of elevation gain observed at Poplar Island. Furthermore, a comparison of elevation gain and surface accretion rates (Table 1) suggests that belowground biomass development is a more important factor in elevation gain than surface accretion, in contrast to many natural marshes (Saintilan et al. 2022), including the reference marsh presented here.
Since the organic matter contribution dominates elevation change where inorganic sediment inputs are low (Turner et al. 2002; Morris et al. 2016), the rate of elevation change should be responsive to macrophyte biomass production, which has a parabolic relationship with elevation. The LM rates of elevation change in the Poplar Island marshes show a similar response to elevation, with an optimal elevation at ~ 20 cm NAVD88, slightly lower than the optimal elevation for biomass production in these marshes, ~ 30 cm NAVD88 (Morris and Staver, in review).
Although these appear to be the dominant processes in the Poplar Island marshes, sediment redistribution is likely a factor on smaller scales. For example, the two groups of SETs in Cell 5AB had different Z0 values, but are also located in separate tidal creek systems. The group in the lower elevation zone includes a marsh pond, which the other does not. Surface accretion rates were higher, and organic content was lower in the lower elevation zone (SETs 37–39, Supplementary Table S2), consistent with models showing higher levels of inorganic deposition at lower elevations (Kirwan and Megonigal 2013), but also suggesting that internal redistribution of sediment resulting from creek and pond bank erosion may be important in discreet locations within marshes.
Morphological changes in channels (Williams et al. 2002) and ponds (Mariotti et al. 2020) are often observed as marshes mature. At Poplar Island, channels have widened and become shallower compared with initial configurations (Taddia et al. 2021), resulting in loss of elevation of the marsh platform immediately adjacent to channels (Fig. 6d), but the eroded sediment may contribute to surface elevation in adjacent areas. It appears that there may be an optimal distance from the channel, suggesting that channel density may be a design element which could be manipulated for resilience. Additionally, the evolution of channel cross sections may switch the tidal system from flood to ebb dominated, with implications for sediment availability in the wetland (Fleri et al. 2019).
Design for Resilience
This study of recent elevation change in the restored Poplar Island tidal marshes suggests that there are three key design factors to consider when creating or restoring tidal marshes using dredged material: elevation, sediment nutrient characteristics, and geomorphology. Nutrients and elevation influence macrophyte production, while channel and pond geometry influence the inorganic sediment supply to the marsh platform. Appropriate elevation within the low marsh should be on the high side of the optimal to promote resilience to RSLR (Fig. 7; Morris 2007; Kirwan and Megonigal 2013). Furthermore, a lower ratio of LM to HM will create more elevation capital and create space for the low marsh to migrate landward in response to RSLR, prolonging the life of tidal marsh habitat. A high fertility substrate promotes rapid marsh establishment and high rates of elevation change in the first decade, and although porewater dissolved NH4+ depletion is fairly rapid, the effect appears to persist for at least a decade (Fig. 5; Anisfeld and Hill 2012; Davis et al. 2017). Finally, channels designed closer to an equilibrium configuration may reduce marsh losses due to edge erosion (Fig. 6d; Williams et al. 2002; Fagherazzi et al. 2012; Fleri et al. 2019), and designs which promote flood dominance will improve sediment retention within the marsh system, also improving marsh resilience.
The future of the existing Poplar Island marshes has been assessed using the Coastal Wetland Equilibrium Model in Morris and Staver (in review), with recommendations for design and adaptive management. Future monitoring will ensure that these designs are performing as expected, provide appropriate data for management and remediation (e.g., thin layer placement), and validate the modeling efforts. Additional studies which would enhance our understanding of how created marsh design influences rates of elevation change should focus on the role of tidal creeks and ponds in promoting sediment deposition on the marsh platform, and the performance of less engineered created marshes with respect to elevation change, e.g. those where creek formation and vegetation colonization are allowed to occur naturally.
Data Availability
Data presented are available from the corresponding author upon request.
References
Anisfeld, S.C., and T.D. Hill. 2012. Fertilization effects on elevation change and belowground carbon balance in a Long Island sound tidal marsh. Estuaries and Coasts 35: 201–211.
Anisfeld, S.C., T.D. Hill, and D.R. Cahoon. 2016. Elevation dynamics in a restored versus a submerging salt marsh in Long Island Sound. Estuarine, Coastal and Shelf Science 170: 145–154.
Baptist, M.J., T. Gerkema, B.C. van Prooijen, D.S. van Maren, M. van Regteren, K. Schulz, I. Colosimo, J. Vroom, T. van Kessel, B. Grasmeijer, P. Willemsen, K. Elschot, A.V. de Groot, J. Cleveringa, E.M.M. van Eekelen, F. Schuurman, H.J. de Lange, and M.E.B. van Puijenbroek. 2019. Beneficial use of dredged sediment to enhance salt marsh development by applying a ‘Mud Motor.’ Ecological Engineering 127: 312–323.
Berkowitz, J., C. Vanzomeren, and C. Piercy. 2017. Marsh restoration using thin layer sediment addition: Initial soil evaluation. Wetland Science and Practice 34: 13–17.
Bertness, M.D. 1991. Zonation of Spartina patens and Spartina alterniflora in New England salt marsh. Ecology 72: 138–148.
Boesch, D.F., G. G. Baecher, W. C. Boicourt, R. I. Cullather, S. Dangendorf, G. R. Henderson, H.H. Kilbourne, M. L. Kirwan, R. E. Kopp, S. Land, M. Li, K. McClure, W. Nardin, W. V. Sweet. 2023. Sea-level rise projections for Maryland 2023. University of Maryland Center for Environmental Science, Cambridge, MD. https://www.umces.edu/sites/default/files/Maryland%20Sea-Level%20Rise%20Projections%202023%20report_0.pdf.
Broome, S.W., C.B. Craft, and M.R. Burchell. 2019. Tidal marsh creation. In Coastal wetlands: An integrated ecosystem approach, ed. G.M.E. Perillo, E. Wolanski, D.R. Cahoon, and C.S. Hopkinson, 789–816. Cambridge, MA: Elsevier.
Cahoon, D.R., and G.R. Guntenspergen. 2010. Climate change, sea-level rise, and coastal wetlands. National Wetlands Newsletter 32: 8–12.
Cahoon, D.R., J.C. Lynch, P. Hensel, R. Boumans, B.C. Perez, B. Segura, and J.W. Day. 2002. High-precision measurements of wetland sediment elevation: I. Recent improvements to the sedimentation-erosion table. Journal of Sedimentary Research 72: 730–733.
Cahoon, D.R., J.C. Lynch, C.T. Roman, J.P. Schmit, and D.E. Skidds. 2019. Evaluating the relationship among wetland vertical development, elevation capital, sea-level rise, and tidal marsh sustainability. Estuaries and Coasts 42: 1–15.
Cahoon, D.R., K.L. McKee, and J.T. Morris. 2021. How plants influence resilience of salt marsh and mangrove wetlands to sea-level rise. Estuaries and Coasts 44: 883–898.
Castillo, J.A., R. MacKenzie, J.R. Manahan, and J. Castillo. 2022. Monitoring the sediment surface elevation change across a chronosequence of restored stands of tropical mangroves and their contemporary carbon sequestration in soil pool. Forests 13: 241.
Cornu, C.E., and S. Sadro. 2002. Physical and functional responses to experimental marsh surface elevation manipulation in Coos Bay’s South Slough. Restoration Ecology 10: 474–486.
Cornwell, J.C., and M.S. Owens. 2011. Quantifying sediment nitrogen releases associated with estuarine dredging. Aquatic Geochemistry 17: 499–517.
Cornwell, J.C., M.S. Owens, L.W. Staver, and J.C. Stevenson. 2020. Tidal marsh restoration at Poplar Island I: Transformation of estuarine sediments into marsh soils. Wetlands 40: 1673–1686.
Craft, C., P. Megonigal, S. Broome, J. Stevenson, R. Freese, J. Cornell, L. Zheng, and J. Sacco. 2003. The pace of ecosystem development of constructed Spartina alterniflora marshes. Ecological Applications 13: 1417–1432.
Craft, C., J. Reader, J.N. Sacco, and S.W. Broome. 1999. Twenty-five years of ecosystem development of constructed Spartina alterniflora (Loisel) marshes. Ecological Applications 9: 1405–1419.
Darby, F.A., and R.E. Turner. 2008. Below- and aboveground biomass of Spartina alterniflora: Response to nutrient addition in a Louisiana salt marsh. Estuaries and Coasts 31: 326–334.
Davis, J., C. Currin, and J.T. Morris. 2017. Impacts of fertilization and tidal inundation on elevation change in microtidal, low relief salt marshes. Estuaries and Coasts 40: 1677–1687.
Davis, J., C. Currin, and N. Mushegian. 2022. Effective use of thin layer sediment application in Spartina alterniflora marshes is guided by elevation-biomass relationship. Ecological Engineering 177: 106566.
Deegan, L.A., D.S. Johnson, R.S. Warren, B.J. Peterson, J.W. Fleeger, S. Fagherazzi, and W.M. Wollheim. 2012. Coastal eutrophication as a driver of salt marsh loss. Nature 490: 388–392.
Demas, S.Y., A.M. Hall, D.S. Fanning, M.C. Rabenhorst, and E.K. Dzantor. 2004. Acid sulfate soils in dredged materials from tidal Pocomoke Sound in Somerset County, MD, USA. Australian Journal of Soil Research 42: 537–545.
Fagherazzi, S., M.L. Kirwan, S.M. Mudd, G.R. Guntenspergen, S. Temmerman, A. D’Alpaos, J. van de Kippel, J.M. Rybczyk, E. Reyes, C. Craft, and J. Clough. 2012. Numerical models of salt marsh evolution: Ecological, geomorphic, and climatic factors. Reviews of Geophysics. https://doi.org/10.1029/2011rg000359.
Fagherazzi, S., M. Marani, and L.K. Blum. 2004. The ecogeomorphology of tidal marshes. Washington: American Geophysical Union.
Fagherazzi, S., G. Mariotti, N. Leonardi, A. Canestrelli, W. Nardin, and W.S. Kearney. 2020. Salt marsh dynamics in a period of accelerated sea level rise. Journal of Geophysical Research: Earth Surface. doi: https://doi.org/10.1029/2019JF005200.
FitzGerald, D.M., and Z. Hughes. 2019. Marsh processes and their response to climate change and sea-level rise. Annual Review of Earth and Planetary Sciences 47: 481–517.
Fleri, J.R., S. Lera, A. Gerevini, L. Staver, and W. Nardin. 2019. Empirical observations and numerical modelling of tides, channel morphology, and vegetative effects on accretion in a restored tidal marsh. Earth Surface Processes and Landforms 44: 2223–2235.
Gedan, K.B., A.H. Altieri, and M.D. Bertness. 2011. Uncertain future of New England salt marshes. Marine Ecology-Progress Series 434: 229–237.
Hesslein, R.H. 1976. An in situ sampler for close interval pore water studies. Limnology and Oceanography 21: 912–914.
Kamrath, B.J., M.R. Burchell, N. Cormier, K.W. Krauss, and D.J. Johnson. 2019. The potential resiliency of a created tidal marsh to sea level rise. Transactions of the ASABE 62: 1567–1577.
Kent, J. 2015. Water level variations at Poplar Island, MD; NOAA technical report NOS CO-OPS. Silver Spring, Center for Operational Oceanographic Products and Services. https://repository.library.noaa.gov/view/noaa/14434.
Kim, C., L.W. Staver, X. Chen, A. Bulseco, J.C. Cornwell, and S.Y. Malkin. 2023. Microbial community succession along a chronosequence in constructed salt marsh soils. Microbial Ecology: 1–20.
Kirwan, M.L., and J.P. Megonigal. 2013. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504: 53–60.
Krauss, K.W., et al. 2017. Created mangrove wetlands store belowground carbon and surface elevation change enables them to adjust to sea-level rise. Scientific Reports 7 (1): 1030.
Marcus, L. 2000. Restoring tidal wetlands at Sonoma Baylands, San Francisco Bay, California. Ecological Engineering 15: 373–383.
Mariotti, G., A.C. Spivak, S.Y. Luk, G. Ceccherini, M. Tyrrell, and M.E. Gonneea. 2020. Modeling the spatial dynamics of marsh ponds in New England salt marshes. Geomorphology 365: 107262.
Mendelssohn, I.A., and J.T. Morris. 2002. Eco-physiological controls on the productivity of Spartina alterniflora Loisel. In Concepts and controversies in tidal marsh ecology, ed. M.P. Weinstein and D.A. Kreeger, 59–80. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Morris, J.T. 2007. Ecological engineering in intertidial saltmarshes. Hydrobiologia 577: 161–168.
Morris, J.T., D.C. Barber, J.C. Callaway, R. Chambers, S.C. Hagen, C.S. Hopkinson, B.J. Johnson, P. Megonigal, S.C. Newbauer, T. Troxler, and C. Wigand. 2016. Contributions of organic and inorganic matter to sediment volume and accretion in tidal wetlands at steady state. Earth’s Future 4 (4): 110–121.
Morris, J.T., P.V. Sundareshwar, C.T. Nietch, B. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83: 2869–2877.
Morris, J.T. and K. Sundberg. 2024. Responses of coastal wetlands to rising sea-level revisited: the importance of organic production. Estuaries and Coasts. https://doi.org/10.1007/s12237-023-01313-8.
Morris, J.T., K. Sundberg, and C.S. Hopkinson. 2013. Salt marsh primary production and its responses to relative sea level and nutrients in estuaries at Plum Island, Massachusetts, and North Inlet, South Carolina, USA. Oceanography 26: 78–84.
Nardin, W., Y. Taddia, M. Quitadamo, I. Vona, C. Corbau, G. Franchi, L.W. Staver, and A. Pellegrinelli. 2021. Seasonality and characterization mapping of restored tidal marsh by NDVI imageries coupling UAVs and multispectral camera. Remote Sensing 13: 4207.
Nyman, J.A., R.J. Walters, R.D. Delaune, and W.H. Patrick. 2006. Marsh vertical accretion via vegetative growth. Estuarine Coastal and Shelf Science 69: 370–380.
Parsons, T.R., Y. Maita, and C.M. Lalli. 1984. A manual of chemical and biological methods for seawater analysis. New York: Pergamon Press.
Poppe, K.L., and J.M. Rybczyk. 2021. Tidal marsh restoration enhances sediment accretion and carbon accumulation in the Stillaguamish River estuary. Washington. Plos ONE 16: e0257244.
Raposa, K.B., M. Bradley, C. Chaffee, N. Ernst, W. Ferguson, T.E. Kutcher, R.A. McKinney, K.M. Miller, S. Rasmussen, and E. Tymkiw. 2022. Laying it on thick: Ecosystem effects of sediment placement on a microtidal Rhode Island salt marsh. Frontiers in Environmental Science 10: 939870.
Saintilan, N., K.E. Kovalenko, G. Guntenspergen, K. Rogers, J.C. Lynch, D.R. Cahoon, C.E. Lovelock, D.A. Friess, E. Ashe, K.W. Krauss, N. Cormier, T. Spencer, J. Adams, J. Raw, C. Ibanez, F. Scarton, S. Temmerman, P. Meire, T. Maris, K. Thorne, J. Brazner, G.L. Chmura, T. Bowron, V.P. Gamage, K. Cressman, C. Endris, C. Marconi, P. Marcum, K. St. Laurent, W. Reay, K.B. Raposa, J.A. Garwood, and N. Khan. 2022. Constraints on the adjustment of tidal marshes to accelerating sea level rise. Science 377: 523–527.
Staver, L., J. Stevenson, J. Cornwell, N. Nidzieko, K. Staver, M. Owens, L. Logan, C. Kim, and S. Malkin. 2020. Tidal marsh restoration at Poplar Island: II. Elevation trends, vegetation development, and carbon dynamics. Wetlands 40: 1687–1701.
Suedel, B.C., A.D. McQueen, J.L. Wilkens, C.L. Saltus, S.G. Bourne, J.Z. Gailani, J.K. King, and J.M. Corbino. 2022. Beneficial use of dredged sediment as a sustainable practice for restoring coastal marsh habitat. Integrated Environmental Assessment and Management 18: 1162–1173.
Taddia, Y., A. Pellegrinelli, C. Corbau, G. Franchi, L.W. Staver, and W. Nardin. 2021. High-resolution monitoring of tidal systems using UAV: A case study on Poplar Island, MD (USA). Remote Sensing 13: 1364.
Turner, R.E., E.M. Swenson, and C.S. Milan. 2002. Organic and inorganic contributions to vertical accretion in salt marsh sediments. In Concepts and controversies in tidal marsh ecology, ed. M.P. Weinstein and D.A. Kreeger, 583–596. Boston, Kluwer Academic: Publishing.
U. S. Army Corps of Engineers. 1996. Poplar Island, Maryland Environmental Restoration Project Integrated Feasibility Report and Environmental Impact Statement, 654 pp. Baltimore: U. S. Army Corps of Engineers, Baltimore District. https://hdl.handle.net/11681/37457.
U. S. Environmental Protection Agency. 2019. Chesapeake Bay mean surface salinity (1985–2018), map of means CB salinity. Chesapeake Bay Program. https://www.chesapeakebay.net/what/maps/chesapeake-bay-mean-surface-salinity-1985-2018.
Vincent, P. 1989. Geodetic deformation of the Oregon Cascadia margin. M. S. Thesis. University of Oregon.
Waltham, N.J., C. Alcott, M.A. Barbeau, J. Cebrian, R.M. Connolly, L.A. Deegan, K. Dodds, L.A. Goodridge Gaines, B.L. Gilby, C.J. Henderson, C.M. McLuckie, T.J. Minello, G.S. Norris, J. Ollerhead, J. Pahl, J.F. Reinhardt, R.J. Rezek, C.A. Simenstad, J.A.M. Smith, E.L. Sparks, L.W. Staver, S.L. Ziegler, and M.P. Weinstein. 2021. Tidal marsh restoration optimism in a changing climate and urbanizing seascape. Estuaries and Coasts 4: 1681–1690.
Welch, M., E.T. Mogren, and L. Beeney. 2016. A literature review of the beneficial use of dredged material and sediment management plans and strategies. Center for Public Service Publications and Reports. http://archives.pdx.edu/ds/psu/19359.
Williams, P.B., M.K. Orr, and N.J. Garrity. 2002. Hydraulic geometry: A geomorphic design tool for tidal marsh channel evolution in wetland restoration projects. Restoration Ecology 10: 577–590.
Windle, A.E., L.W. Staver, A.J. Elmore, S. Scherer, S. Keller, B. Malmgren, and G.M. Silsbe. 2023. Multi-temporal high-resolution marsh vegetation mapping using unoccupied aircraft system remote sensing and machine learning. Frontiers in Remote Sensing. https://doi.org/10.3389/frsen.2023.1140999.
Wöppelmann, G., and M. Marcos. 2016. Vertical land motion as a key to understanding sea level change and variability. Reviews of Geophysics 54: 64–92.
Yozzo, D.J., P. Wilber, and R.J. Will. 2004. Beneficial use of dredged material for habitat creation, enhancement, and restoration in New York-New Jersey Harbor. Journal of Environmental Management 73: 39–52.
Yunus, A.P., Y. Masago, and Y. Hijioka. 2021. Analysis of long-term (2002–2020) trends and peak events in total suspended solids concentrations in the Chesapeake Bay using MODIS imagery. Journal of Environmental Management 299: 113550.
Acknowledgements
Support for this and ongoing work from the Maryland Port Administration and the Maryland Environmental Service is gratefully acknowledged. The authors thank research assistants Debbie Hinkle and Ashley Hollins, as well as numerous other research assistants and students who helped install, maintain, and monitor the Poplar Island SETs. We also thank reviewer Laura Feher, an anonymous reviewer, and Associate Editor Charles Roman for their many helpful suggestions for improving the manuscript.
Funding
This research was funded by the Maryland Port Administration with project management by the Maryland Environmental Service, grant number 14-07-16.
Author information
Authors and Affiliations
Contributions
Conception and design of the experiment: Staver, Stevenson, Hensel, and Cornwell. Execution of the study: Staver, Cornwell, Stevenson, Owens, Nardin, Hensel, and Schwark. Analyzing and interpreting the data: Staver, Morris, Cornwell, Nardin, Owens, and Hensel. Writing of the manuscript: Staver, Morris, Nardin, Cornwell, and Schwark.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Communicated by Charles T. Roman
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Staver, L.W., Morris, J.T., Cornwell, J.C. et al. Elevation Changes in Restored Marshes at Poplar Island, Chesapeake Bay, MD: I. Trends and Drivers of Spatial Variability. Estuaries and Coasts (2024). https://doi.org/10.1007/s12237-023-01319-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12237-023-01319-2