
Abstract
Solanum phureja is one of the most important crops for farmer economy in Colombia, and Phytophthora infestans is considered its main limiting disease. This pathogen is managed using chemical products, generating high production costs as well as environmental pollution. The use of resistant varieties is a strategy that is currently well positioned for the control of this disease, leaving the task of identifying superior parentals to plant breeders. Moreover, the analysis of general and specific combinatorial aptitudes of potential parental genotypes gains importance. The aim of this study was to establish the general and specific combinatorial aptitudes of five elite parental cultivars. For this, 979 clones were assessed. General combinatorial aptitude results show statistically significant differences for the Paola, Primavera and Ocarina parentals. On the other hand, the specific combinatorial analysis allowed concluding that the combinations Ocarina×Violeta and Primavera×Galeras contain non-additive effects involved in inheritance.
Resumen
Solanum phureja es uno de los cultivos más importantes en la economía campesina colombiana. Phytophthora infestans es considerada la principal enfermedad del cultivo. Se controla usando productos químicos, generando altos costos de producción y contaminación ambiental. El uso de variedades resistentes es una estrategia bien posicionada para el control de la enfermedad, dejando la tarea de identificar progenitores superiores a los fitomejoradores, recobrando importancia el análisis de la aptitud combinatoria general y específica de los genotipos potencialmente parentales. El objetivo de este estudio fue determinar la aptitud combinatoria general y específica de cinco parentales élite de S. phureja. Para esto, se evaluaron 979 clones. Los resultados de la aptitud combinatoria general muestran diferencias estadísticamente significativas para los progenitores Paola, Primavera y Ocarina. Por otro lado, el análisis de la aptitud combinatoria específica permitió concluir que las combinaciones Ocarina×Violeta y Primavera×Galeras contienen efectos no aditivos involucrados en la herencia.
Similar content being viewed by others


Introduction
Potato is one of the most important crops for Colombian economy because it is a food security pillar included in the basic family basket. This tuber crop is cultivated by ca. 90,000 families, generating in addition to direct employment, an economy mobility due to the labor required in production, transport, commercialization and industrial transformation. The sector pays about 15×106 wages.year−1. This represents economic income and better life quality for those who are linked directly or indirectly to these activities (Jaramillo 2003; Espinal et al. 2005; Lagoeyte and Arroyabe 2014). The production of this tuber crop in the country in year 2013 was around 138,143 t cultivated in 10,091 ha, with an average yield of 13.1 t.ha−1 (Agronet 2016).
Among all the potato diseases, late blight or potato blight is reported as the most important one, and its causative agent is the oomycete Phytophthora infestans (Mont.) De Bary (Torres 2002; Piñeros 2009; Forbes et al. 2014). This pathogen is found in almost all areas of the world where potato crops have been established, being more destructive in higher humidity areas. It is a polycyclic disease, whose expression depends on the host, on the climatic conditions of the area, on crop management and on the pathogen itself. It requires a temperature between 12 and 18 °C and a relative humidity between 95 and 100% for its development (Agrios 1997; Torres 2002). However, its importance lies on the damage it causes on leaves and stems affecting directly crop yield (Cotes and Ñústez 2014).
To deal with the epidemics of P. infestans, integrated disease management has been used, thus, timely and well-developed cultural practices favor pathogen management. Integrated pathogen management strategies include chemical, cultural, biological and genetic controls. Chemical control involves the use of products seeking to prevent infection. Cultural control involves agronomic management activities on the crop in order to alter the specific conditions in which the pathogen is expressed. Further, biological control that is based on the reduction of the pathogenic microorganism expression by interaction with other living organisms. Finally, genetic control consists in using cultivars that prevent the development of the disease through biochemical reactions (Fry and Smart 1999; Torres 2002; Jaramillo 2003; Forbes and Pérez 2008). In Colombia, late blight is managed mostly by applying chemical molecules such as: Metalaxyl, Cymoxanil, Propamocarb, Dimetomorf, Mandipropamide, Chlorothalonil, Mancozeb, Propineb, Fosetil-aluminum, and Propamocarb+Fluopicolide, which represents up to 10% of the production costs. Apart from this, the continuous use of chemical products implies high management costs and currently these are not always effective (Restrepo and Ñústez 2014; Vargas et al. 2009).
Among the most important challenges that potato breeding programs have is to achieve that the new cultivars obtained combine flavor, starch content, shape and size characteristics of the tuber, with sources of resistance to sanitary problems. Namely, promoting the search for genotypes, that in addition to their agroindustrial characteristics also include lasting resistance to the attack of a disease. In this case, this will lead to a deeper understanding of the genetics of interactions and resistance in potato genotypes to the attack of P. infestans (Abreu et al. 2008; Forbes 2012; Pérez et al. 2014; Forbes and Huarte 2014). In order to understand the gene action of characteristics of interest such as disease resistance and yield components, techniques such as diallelic crosses formed by hybrid populations obtained from n progenitor lines have been used in genetic breeding. These have been designed to estimate genetic effects of the study population, but especially to evaluate the general and specific combinatorial aptitude of parentals and identify materials with higher potential to transmit characteristics to their descendants. The fundamental purpose of diallel crosses is to estimate the variations of the genetic components and to determine which parent has the ability to transfer desirable traits to their progeny. Furthermore, also to identify the possible presence of heterosis or expression of the character of the superior progeny in the expression manifested by the parentals. This condition, according to several authors, is due to genetic effects mainly regarding dominance (Falconer and Macray 1996; Cruz and Regazzi 2001; Gabriel et al. 2008; Guillen et al. 2009; May et al. 2010; Tinjacá 2010; Gabriel et al. 2011). In order to understand the gene action of characteristics of interest such as disease resistance and yield components, in genetic breeding, techniques such as diallelic crosses formed by hybrid populations obtained from n progenitor lines, have been used. These have been designed to estimate genetic effects of the study population, and especially to evaluate the general and specific combinatorial aptitude of parentals. Furthermore, also to identify materials with higher potential to transmit characteristics to their descendants. The fundamental purpose of diallelic crosses is to estimate genetic component variations, to establish which parental cultivar has the ability to transfer desirable traits to their progeny, and identify the possible presence of heterosis or character expression in the progeny character that is higher to the expression expressed by the parentals. According to different authors, this condition is due to genetic effects, but mainly dominance (Falconer and Macray 1996; Cruz and Regazzi 2001; Gabriel et al. 2008; Guillen et al. 2009; May et al. 2010; Tinjacá 2010; Gabriel et al. 2011).
The general combinatorial aptitude evaluated in potato breeding programs for resistance to P. infestans is used to identify the potential that parental cultivars have to transfer characteristic of interest to their descendants. However, in breeding programs sufficient information must be provided as a basis for the selection of parents (Gabriel et al. 2008; Tinjacá 2010; Jaramillo 2015). Genetically, the general combinatorial aptitude is associated with the additive effect of genes, and their low absolute values express that the parentals do not show differences compared to the general average in the crosses carried out. On the contrary, high absolute values indicate differences in favor of the superior value parental (Martínez 1983; Garófalo et al. 2006; Gabriel et al. 2011; Jaramillo 2015). On the other hand, the specific combinatorial aptitude corresponds to the deviation of some of the crosses beyond what is expected, allowing the identification of specific combinations that generate differential responses, whether superior or inferior for the desired characteristic. Hence, this gene component is related to non-additive effects, i.e. dominance and epistasis, and therefore, it is effective to identify useful germplasm sources in potato breeding programs for resistance to late blight (Gabriel et al. 2008; Gabriel et al. 2011; Jaramillo 2015).
According to the aforementioned, the aim of this study is to evaluate the general and specific combinatorial aptitudes in a diallel cross obtained from elite materials selected by the potato breeding program of Universidad Nacional de Colombia, as a basis for the identification of superior and hybrid parentals to be used in future works.
Materials and Methods
Plant Material and Study Area
The diallel cross evaluated includes a total of 979 individuals distributed in 14 F1 families obtained by crossbreeding five cultivars. Three cultivars have high resistance to P. infestans (Paola, Violeta and Primavera) and are adapted to the natural conditions of the mountainous area of the department of Antioquia. In addition, two cultivars with medium resistance to the disease (Ocarina and Galeras) are adapted to the natural conditions of Nudo de los Pastos (department of Nariño) provided by the potato breeding program of Universidad Nacional de Colombia. The plant material was multiplied from sexual seed in mesh houses, and the crop was established in the field under the climatic conditions of the village of Santa Elena, in the municipality of Medellín, department of Antioquia, Colombia. It is located in a very humid montane forest life zone, at an altitude of 2671 m a.s.l.The population corresponds to the set of p (p − 1)/2 hybrids, excluding reciprocals or parentals (Table 1). The crop was subjected to regular and required agronomic practices such as fertilization, earthing up, and weed control.
Severity of P. infestans
Severity assessment of the pathogen was carried out based on symptom descriptions made by Torres (2002) and Forbes et al. (2014), and established visually based on the percentage of foliar area affected by the disease. Additionally, the key proposed by Henfling (1987) was taken as a reference. Weekly evaluations were carried out starting 62 days after sowing (DAS) and were extended until completing 103 DAS, for a total of seven (7) evaluation points, according to Forbes et al. (2014), and Forbes and Pérez (2008) for the environmental conditions in which this study was carried out.
Susceptibility Assessment
Plant susceptibility was evaluated using the relative area under the disease progress curve (AUDPCr), obtained as the ratio between the area under the disease progress curve (AUDPC), which corresponds to the sum of the sick leaf area percentages recorded in each of the evaluations, and the area under the maximum disease progress curve. The latter was obtained assuming that there is a cultivar with 100% of its leaf area affected with the disease during the whole evaluation cycle. When the AUDPCr value is higher, the clone or cultivar is more susceptible, i.e. in an evaluation, a 100% of the sick leaf area due to late blight during the entire evaluation cycle would have a value of one (1). On the other hand, cultivars with low values correspond to individuals with higher resistance (Zúñiga et al. 2000; Forbes and Pérez 2008; CIP 2009; Forbes et al. 2014).
Experimental Design and Statistical Analysis
In total 979 plots were established corresponding to each of the clones and each plot comprised five plants that represent an experimental unit. Crops were established at a distance of 0.3 m between plants and 0.9 m between rows, planted in a randomized complete block design with three repetitions. Inoculum pressure was guaranteed by establishing plots with plants susceptible to the disease (S. phureja cv. Criolla Colombia regional) arranged randomly in each furrow. Results were subjected to a transformation of the variable to log_e (AUDPCr +0.01). The statistical model used for the calculation of general and specific combinatorial aptitudes corresponds to the fourth model proposed by Cruz and Regazzi (2001). The effect of the Sij factor of the independent crossing of the maternal or paternal effect of the same was evaluated, by means of a balanced diallel that considers the parentals as fixed effects of the analysis. That is, the analysis is performed for these in sensu stricto and it is not possible to extend it to a hypothetical population. The statistical model corresponds to a log _ e (y + 0.01) = μ + X _ 1 β + X _ 2 g + X _ 3 δ + Z _ 1 ε + e, where y is the AUDPCr observations vector, μ is the general average of the model, X_1 β is a fixed effects vector of the block of a mx1 size, and where m is equal to the number of block levels. Further, X_2 g is a fixed effects vector of the general combinatorial aptitude of parental I, X_3 δ is a fixed effects vector of the specific combinatorial aptitude s_ij of parental i, and j. In addition, Z_1 ε is a random effects vector of the experimental error, and e is a residual random effects vector of the sampling error.
Statistical analyzes were performed using the MCMCglmm package (Hadfield 2016) included in the R Core Team software. The estimators used correspond to the median and the mean of the a posteriori distribution of the parameter. Afterwards, its respective a posteriori high-density (HPD) interval of 95% probability was obtained. For this purpose, the CODA package (Plummer 2015) was used.
Results
The results obtained indicate that the disease developed well in the study population, considering that the levels of severity varied between zero (0) and 100% of affectation on the leaf area, and with AUDPCr values between 0.000 and 0.982. However, these show a wide variability between the responses of the cultivars, indicating that the study population has genes that code for resistance to the disease.
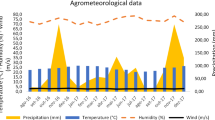
The environmental conditions that governed the study phase corresponded to an average temperature of 15.20 °C in a range between 11.12 and 21.09 °C, an average relative humidity of 76% in a range of 46 to 95% ± 16.84% and an accumulated precipitation during the study time of 165.30 mm.
Table 2, shows that in families 13, 14, 23, 43, 45, 52 and 53 more than 50% of the cultivars exhibit low leaf area affected by the disease (less than 5%). On the other hand, in families 15 and 42 most of the cultivars show average resistance to the disease attack. In these families, ca. 95% of the cultivars evaluated show a maximum of 30% of the leaf area affected during the evaluations. Family 12 showed 75% of its population with medium resistance to the disease (with values ranging between 5 and 30% of the leaf area affected) and its remaining population (25%) presented a non-resistance response. The family that showed a higher percentage of individuals with medium resistance to the disease (99% of the individuals) corresponds to family 45, meanwhile family 14 had the highest percentage of individuals who did not show any leaf area affected by the disease (3%). Nonetheless, it must be noted that families 23, 43 and 53 are the product of crosses with the Paola cultivar, reported as highly resistant. As expected, these have a higher average number of individuals in the highly resistant category (AUDPCr equal to zero).
Li: Lower value of the interval, Ls: Upper value of the interval.
General and Specific Combinatorial Aptitude
The mean AUDPCr value for the study population is −1.8168, the variance of the experimental error was 0.0093, and that of the sampling error was 0.0795.
General Combinatorial Aptitude
Regarding general combinatorial aptitude values, in Table 3 statistically significant differences are observed for the cultivars Ocarina, Primavera and Paola, meanwhile cultivars Violeta and Galeras did not show statistically significant differences. Regarding average population values, parentals Violeta (−0.0603), Paola (−0.1311) and Primavera (−0.1470) showed the lowest average values, meanwhile Galeras and Ocarina showed the highest average values (0.1649 and 0.1736, respectively).
Specific Combinatorial Aptitude
The results of this study for specific combinatorial aptitude are shown in Table 4. Results show that the hybrid combinations of cultivars Ocarina x Paola, Ocarina x Primavera, Violeta x Paola, Violeta x Primavera, Violeta x Galeras, Paola x Primavera, Paola x Galeras, and Primavera x Galeras, did not show statistically significant differences in their response to P. infestans. On the contrary, the crosses between cultivars Ocarina x Violeta and Ocarina x Galeras, showed statistically significant differences.
Discussion
Description of the Study Population
The severity of late blight is expressed as a direct response to climatic conditions, thus, the pathogen requires for an optimal development relatively low temperatures and an accumulated precipitation higher than 393 mm during the crop development cycle (Haynes and Christ 1999; Costanzo et al. 2004). This allows to infer that the parental has genes that code for resistance. Nonetheless, notice should be taken in the fact that precipitation during the study period did not correspond to the optimum value. However, the development of the disease was completed, which implies that it is possible that the cultivars behave slightly more susceptible under other conditions. Nevertheless, in relative terms, families are expected to show a similar behavior. The results allow observing that it is possible to select genotypes that show resistance to P. infestans in the study population, so these can be included in potato breeding programs with resistance to the pathogen. In addition, these can also be focused on yield increase or concentration of other valued features such as tuber quality, similar to what has been reported by Muhinyuza et al. (2016). These authors selected materials for resistance to late blight with high yields, reporting a direct positive relationship between the reduction of late blight levels in the crop versus increases in tuber yield.
General Combinatorial Aptitude
When the evaluated AUDPCr variable is considered results show that at more negative values there is a higher resistance of the material to the disease. Furthermore, taking into account parentals with statistically significant differences, Table 3 shows that those who showed this combination of characteristics, i.e. the best overall combinatorial aptitude, corresponds to the Paola and Primavera parentals (−0.1311 and − 0.1470, respectively). Meanwhile, Ocarina with statistically significant differences but with a positive mean value (0.1736) corresponds to the parental that transmits less resistance to the descending population. Since resistance implies low AUDPC values, progenitors that reach the lowest general combinatorial aptitude value with statistically significant differences correspond to parentals that should be included in genetic breeding programs for resistance to P. infestans. The results obtained with the cultivars Paola and Primavera indicate that variability in responses of the parentals is regulated by the gene action of non-additive effects, allowing to identify these cultivars as the parentals that have better general combinatorial aptitude to transmit resistance characteristic in the study population (Garofalo et al. 2006; Gabriel et al. 2011; Jaramillo 2015).
A statistically insignificant value in the general combinatorial aptitude implies that the parents behave the same as their offspring, and that the breeding value would not represent importance in the inheritance of the resistance (Cruz y Regazzi 2001; Garofalo et al. 2006; Jaramillo 2015). In this sense, it can be concluded that the Galeras and Violeta cultivars do not show statistically significant differences in the character of interest, i.e. in resistance to P. infestans, are not recommended to be included in future breeding programs for resistance to potato late blight.
Specific Combinatorial Aptitude
Values with statistically significant differences establish that the breeding value is important, and that it is involved in the inheritance of the character. Additionally, this indicates that populations with significant results behave better compared to others (Cruz and Regazzi 2001; Jaramillo 2015; Tinjacá 2010). Then, it is possible to conclude that within the study population these combinations contain non-additive effects involved in the inheritance of the character, finding that they have a superior behavior compared to the others. Moreover, crossing between cultivars Ocarina x Violeta with negative average values generate more resistance to P. infestans than what was expected for this crossing, taking into account that the Ocarina cultivar, according to the results obtained for the general combinatorial aptitude, does not transmit, or transmits little resistance. On the contrary, the crossing between cultivars Ocarina x Galeras with statistically significant differences and a higher average AUDPCr value generates more susceptibility than expected. Further, the ocarina cultivar generates less resistance when it is crossed with a moderately resistant material and higher resistance when it is crossed with the cultivar Violeta, with high resistance to the disease. However, this parental cultivar does not show a good general combinatorial aptitude.
Nevertheless, results indicate that in the study population, the general combinatorial aptitude contributes more to the resistance compared to the specific combinatorial aptitude. This is contrary to what was reported by Muhinyuza et al. (2016), who found in their study population, both general as well as specific combinatorial aptitudes were equally important when selecting parentals. Moreover, in the study population and dissimilar from other studies, a predominance of non-additive effects over the additive gene action is observed in the inheritance of the quantitative resistance to potato late blight. Hence, results indicate that promissory materials to be considered in future potato-breeding works for resistance to P. infestans that contain the parentals used in this study, should be selected focusing on their general combinatorial aptitude. Thus, in this study, these correspond to the cultivars Paola and Primavera. In this sense, Preciado et al. (2005) as well as May et al. (2010), indicated that in case of detecting higher effects of the general combinatorial aptitude, these contribute more to the genetic variation of the resistance than the effects of the specific combinatorial aptitude. This means that the evaluated characters are associated with high breeding values, rather than non-additive effects, making it possible to exploit the breeding value of the genetic variance available through any variation of the recurrent selection.
Conclusions
The parental cultivars Ocarina, Primavera and Paola showed statistically significant differences in the general combinatorial aptitude.
The parental cultivars Paola and Primavera showed negative mean values and statistically significant differences for the evaluation of resistance to P. infestans, which indicates that of the entire study population, the recommended parentals to be included in new breeding programs for the variable assessed, are Primavera and Paola.
The cultivars Galeras and Violeta do not show statistically significant differences in terms of general combinatorial aptitude. Therefore, these are not recommended to be included in future breeding programs for resistance to P. infestans.
Within the study population, the hybrid combinations Ocarina x Violeta and Ocarina x Galeras have the best specific combinatorial aptitude, which indicates that in these parental cultivars, there are non-additive effects involved in the inheritance of resistance to P. infestans, and are considered the best hybrid combination for the character assessed.
Regarding specific combinatorial aptitude, in hybrid combinations that include the parental cultivar Ocarina, the best combination is obtained with the parental cultivar Violeta, in which the highest resistance capacity is transmitted to its descendant population.
References
Abreu, F., D. Silva, C. Cruz, and E. Gomide. 2008. Inheritance of resistance to Phytophthora infestans (Peronosporales, Pythiaceae) in a new source of resistance in tomato (Solanum sp. (formerly Lycopersicon sp.), Solanales, Solanaceae). Genetics and Molecular Biology 31: 493–497.
Agrios, G. 1997. Plant pathology. Fourth ed. California: Academic Press 635 p.
Agronet. 2016. Reportes estadísticos. In Cifras agropecuarias. Colombia: Bogotá.
Costanzo, S., B. Christ, and K. Haynes. 2004. Late blight resistance in a diploid full-sib potato family. Plant Breeding 123 (4): 377–381. https://doi.org/10.1111/j.1439-0523.2004.00999.x.
Cotes, J., and C. Ñústez. 2014. Prueba de evaluación agronómica para registro de variedades de papa criolla (Solanum phureja Juz. et Buk.) subregión montaña antioqueña. Universidad Nacional sede Medellín. Medellín. 72 p.
Cruz, C., and A. Regazzi. 2001. Modelos biométricos aplicados ao melhoramento genético. 2da Edición. Editora UFV, 309. Brazil: Vicosa.
Espinal, C., H. Martínez, and Y. Peña. 2005. Ministerio de Agricultura y Desarrollo Rural: Observatorio agrocadenas Colombia documento de trabajo No. 58. http://www.agronet.gov.co/www/docs_agronet/2005112145659_caracterizacion_cacao.pdf. Accessed Noviember 2016.
Falconer, D., and T. Macray. 1996. Introduction to quantitative genetics. 4th ed. England: Longman 464 p.
Forbes, G. 2012. Using host resistance to manage potato late blight with particular reference to developing countries. Potato Research 55 (3–4): 205–216. https://doi.org/10.1007/s11540-012-9222-9.
Forbes, G., and M. Huarte. 2014. La resistencia al tizón tardío como herramienta de control en los países en desarrollo. Revista Latinoamericana de La Papa 18: 36–58.
Forbes, G., and W. Pérez 2008. Manual técnico: El tizón tardío de la papa. Centro Internacional de la Papa (CIP). Lima, Peru. 39 p.
Forbes, G., W. Pérez, and J. Andrade 2014. Evaluación de la resistencia en genotipos de papa a Phytophthora infestans bajo condiciones de campo: Guía para colaboradores internacionales. Centro Internacional de la Papa (CIP). Lima, Peru. 50 p. https://doi.org/10.4160/9789290604501
Fry, W., and C. Smart. 1999. The return of Phytophthora infestans, a potato pathogen that just won’t quit. Potato Research 42: 279–282.
Gabriel, J., N. Estrada, W. García, E. Carrasco, H. Equise, J. Herbas, G. Plata, J. Vallejos, and A. Gandarillas. 2008. Estrategia y perspectiva del mejoramiento genetico de papa (Solanum tuberosum L.) en Bolivia. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Cochabamba, Bolivia. 36 p.
Gabriel, J., L. Orellana, G. Plata, and M. Siles. 2011. Aptitud combinatoria de la resistencia al tizon tardio (Phytopthora infestans) en cultivares nativos de papa. Revista Latinoamerica de La Papa 16: 85–98.
Garófalo, J., H. Andrade, and X. Cuesta. 2006. Evaluación de la aptitud combinatoria general y específica en 21 progenies de papa Solanum phureja para resistencia a tizón tardío Phytophthora infestans. En: II Congreso Ecuatoriano de la Papa. Ambato, Ecuador.
Guillen, P., E. Cruz, G. Castañon, R. Osorio, N. Brito, A. Lozano, and U. López. 2009. Aptitud combinatoria general y específica de germoplasma tropical de maíz. Tropical and Subtropical Agroecosystems 10 (1): 101–107.
Hadfield, J. 2016. MCMC generalised linear mixed models. The MCMCglmm R Package. Version 2.24. 13 p.
Haynes, K., and B. Christ. 1999. Heritability of resistance to foliar late blight in a diploid hybrid potato population of Solanum phureja x Solanum stenotomum. Plant Breeding 118 (5): 431–434. https://doi.org/10.1046/j.1439-0523.1999.00394.x.
Henfling, J. 1987. El tizón tardío de la papa: Phytophthora infestans. Boletín de Información Técnica 4. Centro Internacional de la Papa (CIP). Lima, Peru. 25 p.
Jaramillo, S. 2003. Monografía sobre Phytophthora infestans (Mont) De Bary. Universidad Nacional de Colombia sede Medellín. Colombia: Medellín 120 p.
Jaramillo, P. 2015. Estudio de los componentes genéticos de la resistencia al tizón tardío (Phytophthora infestans) (Mont.) de Bary en papa (Solanum phureja). Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP). Cutuglahua, Pichincha, Ecuador. 21 p.
Lagoeyte, J., and I. Arroyabe 2014. Manual del cultivo de papa bajo buenas prácticas agrícolas. FEDEPAPA, Cadena de la papa Antioquia, Secretaría de Agricultura y Desarrollo Rural, Gobernación de Antioquia. 123 p.
Martínez, A. 1983. Diseños y análisis de experimentos de cruzas dialélicas. Segunda edición. Colegio de posgraduados, Chapingo, Mexico, 252 p.
May, A., G. Nájera, J. Suárez, M. Elos, J. Cortés, A. Pérez y L. Latournerie. 2010. Efectos heteróticos y aptitud combinatoria en poblaciones de chile dulce (Capsicum annuum L.). Revista Fitotecnia Mexicana 33(4): 353–360.
Muhinyuza, J., H. Shimelis, R. Melis, J. Sibiya, and M. Ndambe. 2016. Combining ability analysis of yield and late blight (Phytophthora infestans (Mont.) de Bary) resistance of potato germplasm in Rwanda. Australian journal of crop science. AJCS 10(6):799–807, ISSN:1835-2707. DOI: https://doi.org/10.21475/ajcs.2016.10.06.p7303
Pérez, W., M. Ñahui, D. Ellis, and G. Forbes. 2014. Wide phenotypic diversity for resistance to Phytophthora infestans found in potato landraces from Peru. Plant Disease 98 (11): 1530–1533. https://doi.org/10.1094/PDIS-03-14-0306-RE.
Piñeros, C. 2009. Recopilación de la investigación del sistema productivo papa criolla, Secretaría de Agricultura y Desarrollo Económico. Departamento de Cundinamarca y Fedepapa, 152. Colombia: Bogotá.
Plummer, M. 2015. Output analysis and diagnostics for MCMC. The CODA R package. Version 0.18–11. 45 p.
Preciado, O., I. Terrón, M. Gómez y G. Robledo. 2005. Componentes genéticos en poblaciones heteróticamente contrastantes de maíz de origen tropical y subtropical. Agronomía Mesoamericana 16:145–151. DOI: https://doi.org/10.15517/am.v16i2.11867.
Restrepo, S., and C. Ñústez. 2014. Situación del tizón tardío en Colombia. In: Memorias ALAP, XXVI Congreso de la Asociación Latinoamericana de la Papa. Bogotá Colombia.
Tinjacá, S. 2010. Estudios de heredabilidad de la resistencia horizontal a Phytophthora infestans causante de la gota en la especie diploide de papa Solanum phureja. Magister Thesis in Agricultural Sciences, Faculty of Agronomy, Universidad Nacional de Colombia, 123. Colombia: Bogotá.
Torres, H. 2002. Capítulo 1: Tizón tardío. In pp, vol. 5-14, 68. En: Manual de las enfermedades más importantes de la papa en el Perú. Centro Internacional de la Papa (CIP). Lima, Peru.
Vargas, A., L. Quesada, M. Céspedes, N. Carreño, A. González, A. Rojas, A. Zuluaga, K. Myers, P. Jiménez, J. Bernal, and S. Restrepo. 2009. Characterization of Phytophthora infestans populations in Colombia: First report of the A2 mating type. Phytopathology 99 (1): 82–88. https://doi.org/10.1094/PHYTO-99-1-0082.
Zúñiga, L., G. Molina, H. Cadena, and P. Rivera. 2000. Resistencia al tizón tardío de la papa (Phytophthora infestans) en cruzamientos de cultivares y clones de papa (Solanum tuberosum L.). Revista Mexicana de Fitopatología 18 (1): 1–9.
Acknowledgments
The authors wish to express their gratitude to all those who participated in the development of this study, especially to Universidad Nacional de Colombia, Medellin Campus, through its Postgraduate Course in Agricultural Sciences, and also to the Corporación Colombiana de Investigación Agropecuaria (Agrosavia), research center C.I. La Selva.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Grisales-Vásquez, N.Y., Cotes-Torres, J.M. General and Specific Combinatorial Aptitude in a F1 Population of Solanum phureja with Resistance to Phytophthora infestans. Am. J. Potato Res. 96, 55–61 (2019). https://doi.org/10.1007/s12230-018-9688-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12230-018-9688-3