
Abstract
An oasis is a unique natural landscape in arid and semi-arid areas, significant for regulating regional microclimates and hydrological processes in deserts. However, little is known regarding the response of natural oasis plants communities to various environmental factors. Nineteen sample plots (50 m × 50 m) were selected in the Daliyabuyi Oasis in the Taklimakan Desert hinterland based on the location of groundwater monitoring wells and 76 vegetation quadrats (25 m × 25 m) were established. A two-way indicator species analysis, Mantel test, detrended correspondence analysis, canonical correspondence analysis (CCA), and hierarchical partitioning were used to provide an in-depth analysis of community classification, species composition, and environmental interpretation of the oasis. A generalized linear model was used to verify the results which showed that the current oasis community could be divided into four types according to the dominant species, which is controlled by soil moisture. Measurement of species composition and distribution of communities showed significant differences between species diversity of individual community types. Variations in groundwater depth affects patterns of species diversity which is sensitive to richness, while the degree of surface water disturbance affects the pattern of species evenness. Moreover, the CCA ordination map showed that community distribution and diversity characteristics have their own preferences in habitat gradients. The study concluded that the species dominance of the community and the composition and distribution are not dominated by a single factor. There are differences in the scale and effect of different water resource types in maintaining community characteristics.
Similar content being viewed by others

Introduction
In the context of global climate change, the over-exploitation of limited resources due to human activities or land degradation due to natural causes is becoming increasingly severe (Zhang et al. 2007; Xie et al. 2020). Desertification refers to the phenomenon of land degradation in arid or sub-humid areas with drought under various factors (Yu et al. 2022). In the past few decades, issues such as biodiversity loss caused by intensified desertification have seriously restricted regional development. and has become an urgent ecological issue that needs immediate attention in arid and semi-arid regions. Understanding the vegetation site conditions in these areas and the response patterns to resource conditions will help identify the mechanisms underlying community formation, making it possible to seek preventive means to resist desertification and prevent further degradation of vegetation habitats (Keyimu et al. 2018). Such initiatives have significant practical value in promoting the formulation of vegetation protection strategies in fragile ecosystems.
An oasis is a natural geographical landscape ubiquitous and unique in arid areas and can be divided into artificial oases and natural oases (Qi et al. 2007; Li et al. 2008; Lei et al. 2015). The artificial oasis is an ecological landscape formed by human activities intervening in structuring desert ecosystems (Qiao et al. 2009). It is used as a central place for human survival and development. A natural oasis is an “island”-shaped green space formed by various biological and ecological processes (Pankova 2008). This type of oasis has a specific role in improving the microclimate, water resources, and soil of the surrounding area (Wang et al. 2021). Water is an essential resource for maintaining the sustainable development of oases, playing a decisive role in forming their distribution and size. Water, oases, and the surrounding desert form a system that interacts and restricts each other (Li et al. 2016; Yin et al. 2022). The balance of this system is directly related to the stability of oasis development and significant to biodiversity and ecosystem protection.
A community is a collection of populations of different species distributed in a specific geographical space. Community ecology is an important branch of ecology that explains the interactions between biological organisms and the environment on a local scale (Thammanu et al. 2021). As an important part of an oasis, the coexisting species of desert riparian forest communities have characteristic resistance to high temperatures, salinity, and wind and sand (Nargis et al. 2013; Wang et al. 2020; Zhou et al. 2020), but the characteristics of different communities are affected by the internal habitat heterogeneity of the oasis. Research has shown that changes in water availability can lead to changes in species composition and function of plant communities (Muscarella et al. 2016; Gong et al. 2017; Blanchard et al. 2019). The survival of plant communities in arid areas depends on precipitation, surface runoff, and groundwater. Extreme drought and other factors result in most areas having minimal precipitation and surface runoff (Lamontagne et al. 2005). Therefore, groundwater is considered a key factor affecting community dynamics and the functional characteristics and species distribution in conjunction with soil physicochemical properties (Chen et al. 2021). However, not all desert communities are constrained by groundwater conditions during their formation. Research on the Tarim River in western China has shown that overflow caused by surface water in inland river basins with periods of high water positively affected the restoration of degraded ecosystems (Ling et al. 2019). Studying desert riparian forest communities under both conditions is helpful to identify the role of different ecological processes in community construction and aids in obtaining universal laws.
The Daliyabuyi Oasis, located in China’s Tarim Basin, is an alluvial oasis formed by the Keriya River deep into the hinterland of the Taklimakan Desert. The Oasis is influenced by the flow of surface water during the annual high-water period, forming a desert-wetland ecosystem in an arid area; however, only a few studies highlight the vegetation characteristics in this area and its response to environmental factors. An in-depth study of the plant community and the environment of the Daliyabuyi Oasis was carried out to promote its sustainable development. Our aim was to answer the following questions: (1) How do the community types and species diversity of natural oases respond to environmental factors? (2) How does the differences in water resource supply play a role in perspectives at the community scale. By answering these questions, we hope to provide a scientific basis for the protection and management of the Oasis.
Materials and methods
Study area
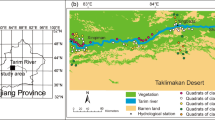
The Daliyabuyi Oasis is located in the Taklimakan Desert of China’s hinterland at 38° 16′ N/38° 37′ N, 81° 41′ E/82° 20′ E, marking the last oasis of the Kriya River, the second largest river on the southern margin of the Tarim Basin. The Oasis has an area of approximately 342 km2 and altitudes of 1100–1300 m, with a typical warm temperate, arid climate (Wan et al. 2022) and significant diurnal temperature differences. Average annual precipitation is less than 20 mm and potential evaporation is more than 2000 mm. The interior of the oasis has the characteristics of reticular fluvial deposition under the scouring of the Keriya River. The river course is intricate, with yearly high and low water periods (Shi et al. 2021). In the high-water period, the increase in volume leads to surface water overflow of varying degrees in some areas. The Oasis plant community consists of sparse desert vegetation, mainly Populus euphratica, Tamarix chinensis, and Phragmites australis. At the end of 2018, several vegetation monitoring transects were established in the oasis from south to north. Nineteen groundwater monitoring wells were built in the transects according to the shape of the oasis to record the groundwater depth, providing an ideal platform for community ecology research (Fig. 1).

Overview map of the study area
Vegetation survey
The research group entered the Daliyabuyi Oasis in the summer of 2021 and established 19 vegetation 50 m × 50 m sample plots based on the locations of the groundwater monitoring wells. Each sample plot was divided into four 25 m × 25 m quadrats, totaling 76. Tree and shrub surveys were carried out within each quadrat, and the species name, number, height, crown width, and other indicators were recorded. For sample plots with high herbaceous cover, five 1 m × 1 m quadrats were randomly set up in each larger quadrat, and the recordings of the small quadrat were converted to the area of the larger to extrapolate the overall characteristics of herbaceous plants in the community. For areas with sparse herbaceous plants, species-specific survey methods were similar as with trees and shrubs. A total of 10 species were recorded (Table 1). After the quadrat surveys, the sample plots were geolocated using a handheld GPS and latitude and longitude recorded.
Acquisition and quantification of environmental factors
Three 1-m deep soil profiles were randomly established within each sample plot, and each was divided into six layers (0–5, 5–20, 20–40, 40–60, 60–80, and 80–100 cm) for soil collection. The same soil layer from the three profiles was thoroughly mixed during the collection and placed in aluminum boxes and plastic bags and brought to the laboratory to determine soil water content, pH, total dissolved solids, organic matter, total nitrogen, and total phosphorus. Groundwater depth data were obtained from the average of the monitoring data from the 2019–2021 growing season (April–October). The monitoring well of sample plot 16 was damaged by flooding and caused missing data; therefore, its groundwater depth was obtained from the monitoring data of 2019 and 2020.
Landsat-8/OLI remote-sensing images from 2017 to 2021 were used in this study. The characteristics of surface water distribution in these five years were obtained by extracting the water index and threshold segmentation technique (Fig. 2) (Li et al. 2021). The following formula was used to quantify the effect of surface water disturbance (SWD) on the community:

Oasis 2017–2021 surface water distribution frequency map
To consider any errors in positioning the handheld GPS at the sample site, S represents the number of surface water pixels within 1 ha of the sample site, and wi the frequency of the ith pixel in the monitoring time frame. SWD represents the degree of surface water disturbance at the local scale; the higher the value, the greater the degree of surface water disturbance at the community level.
Community type classification and Mantel test
Species importance value (IV) is a comprehensive quantitative index in community ecology that reflects the status and role of a population (Sun et al. 2020). The calculation is:
where Ar denotes the relative abundance, Hr the relative height, Cr the relative cover, and Fr the relative frequency. IVsample plot and IVquadrat represent species importance values at two sampling scales: sample plot and quadrat, respectively. The characterization of community types as sets renders them as basic taxonomic unit subsets of these sets. Therefore, while focusing on overall species composition and dominant species characteristics within a certain range, the classification of the relevant community types should be calculated using the species importance values of the sample plots at larger scales, that is, IVsample plot. Species diversity, as an indicator to quantify the community characteristics using the quadrat as the study unit, helps to understand the specific structural details of the community, increases the reproducibility of ecological studies in environmentally homogeneous lots, and increases significance statistically; therefore, IVquadrat was chosen for the correlation analysis of species diversity.
Classification of communities was performed using two-way indicator species analysis (TWINSPAN) in PC-ORD 5.0, detrended correspondence analysis (DCA) in CANOCO 5.0 (Khan et al. 2013; Moradi and Vacik 2018; Ma et al. 2020), and the Mantel test to assess the correlation between plant composition and environmental factors for different community types (Xu et al. 2022). Plant composition distance of the community was measured using the Bray–Curtis distance matrix transformed by the importance value of species at the sample plot scale, and the environmental factors were transformed into the European distance matrix after standardization to calculate the correlation coefficient between the two vectors and the significance after 999 permutation tests. The process is analyzed with the ‘vegan’ package in R 4.1.3.
Species diversity calculation and CCA
In this study, five single-species samples were found during data collection and were excluded because they did not meet the data requirements for species diversity calculations. The remaining 71 quadrats were included in the diversity analysis, and the species richness, Shannon–Wiener, Simpson, and Pielou evenness indices were selected to calculate species diversity (Lu et al. 2020). The specific formulae for these four equations are as follows:
where Pi is the importance value of the ith species in the corresponding community quadrat, IVquadrat, and S is the total number of species.
Canonical Correspondence Analysis (CCA) is a multiple regression ranking method based on correspondence analysis (Palmer et al. 2008). In this study, the environmental factors were first standardized. The variance inflation factor (VIF > 10) was then removed by collinearity diagnostics. The remaining environmental factors were included in the matrix of explanatory variables required for multiple regression to analyze them with the matrix of response variables. The CCA and permutation tests for explanatory variables were implemented using the ‘vegan’ package in R 4.1.3, and hierarchical partitioning performed using the ‘rdacca.hp’ package to obtain the individual contributions of each environmental factor (Lai et al. 2022).
Generalized linear model regression and validation
Generalized linear models were developed using the datasets identified. Logistic regression based on multi-category data, quasi-Poisson regression fixed on count data, gamma regression using continuous variables, and beta regression based on continuous ratio data were used as the main models. Several regression models were developed using community and water resources data. Combining the results of hierarchical partitioning and significance analysis, the driving factors of each response variable were identified, thereby verifying the differences in the roles of explanatory variables in maintaining community characteristics. The hierarchical partitioning of the generalized linear model was implemented using the ‘hier.part’ package in R 4.1.3 (Olea et al. 2010).
Results
Classification of plant communities and their diversity
The outcomes of the TWINSPAN and community DCA ranking showed general agreement on the classification of community types in the Daliyabuyi Oasis (Fig. 3). The communities could be classified into four types using the second level of the TWINSPAN results as the basis for classification:
-
Type 1: Phragmites australis + Tamarix chinensis. This community type included five sample plots: 1, 2, 3, 4, and 16. The average coverage was 43.4%, and companion species included P. euphratica, Glycyrrhiza uralensis, Alhagi sparsifolia, Hexinia polydichotoma, and Onopordum acanthium.
-
Type 2: Tamarix chinensis + Karelinia caspia. This community type included four sample plots: 8, 13, 15, and 18. The average coverage was 27.6%, and the companion species included P. euphratica, G. uralensis, A. sparsifolia, and H. polydichotoma.
-
Type 3: Populus euphratica + Tamarix chinensis. This community type included eight sample plots as 6, 9, 10, 11, 12, 14, 17, and 19. The average coverage was 32.2%, and both were co-dominant communities with only two species of P. euphratica and T. chinensis.
-
Type 4: Populus euphratica + Tamarix chinensis + Sophora alopecuroides. This community type included two sample plots 5 and 7. The average coverage was 41.5%, and the companion species was Inula salsoloides.

Community classification results of 19 sample plots based on TWINSPAN (a) and DCA (b)
Alpha diversity calculation of community types showed significant differences between some community types (Fig. 4). Species richness and Shannon–Wiener index rank the community types from largest to smallest as Type 1 > Type 2 > Type 4 > Type 3; the Simpson index ranks as Type 2 > Type 1 > Type 4 > Type 3, and the Pielou index ranks as Type 3 > Type 4 > Type 2 > Type 1.

Species diversity index of different plant community types (mean ± SE). Note: different letters represent significant differences between paired samples: P < 0.05
Responses of different community types to environmental factors
The Spearman correlation coefficient indicated a correlation between some environmental variables (Fig. 5). The disturbance degree of surface water had a negative correlation with groundwater depth and a positive correlation with soil moisture and pH. Groundwater depth was negatively correlated with soil moisture and pH. There was a positive correlation between soil moisture and pH. In terms of soil chemical properties, there was a positive correlation between organic matter and total nitrogen. Moreover, there was a positive correlation between total nitrogen and total phosphorus.

Mantel test results for different community types based on species importance value matrix and environmental factor matrix (SWD: degree of surface water disturbance; GWD: groundwater depth; SWC: soil water content; TDS: total dissolved solids; SOM: soil organic matter; STN: soil total nitrogen; STP: soil total phosphorus)
There was a positive correlation between the importance value of community species and soil water by the Mantel test (r = 0.373, P = 0.018), and the difference in soil water was the main factor influencing the classification of oasis community types. The Mantel test results revealed that the main factors affecting species composition and dominance of the various communities within the community types of Phragmites australis + Tamarix chinensis (Type 1) were the degree of surface water disturbance (r = 0.267, P = 0.049) and soil moisture content (r = 0.394, P = 0.013). The main environmental factor affecting the differences in species composition and dominance within the other three community types was soil water (Type 2: r = 0.312, P = 0.047; Type 3: r = 0.357, P = 0.023; Type 4: r = 0.350, P = 0.020).
Response of community species diversity to environmental factors
The CCA ranking model established for the species diversity of the Daliyabuyi Oasis had a high overall significance (P = 0.001), which explains better the analytical effect of environmental factors on species diversity information. The distribution of this information of each quadrat on the two-dimensional map was consistent with community classification. The differences in the species diversity of each community can be clearly distinguished from the two-dimensional map (Fig. 6).

CCA of environmental factors and species diversity of oasis community. Note: the definitions of the environmental variable abbreviations are consistent with those in Fig. 5
From the sample distribution information, community (Type 1) of Phragmites australis + Tamarix chinensis tended to be distributed in areas with higher surface water disturbance and higher soil moisture content. The community type of Tamarix chinensis + Karelinia caspia (Type 2) had a strong ability to adapt to the environment, and there was no clear tendency in habitat selection. Type 3 community of Populus euphratica + Tamarix chinensis was distributed in areas with deep groundwater and high soil organic matter; The Type 4 community of Populus euphratica + Tamarix chinensis + Sophora alopecuroides also tended to be in habitats with deeper groundwater but the degree was weaker than that of Type 3.
As indicated in Fig. 6 and Table 1, information on the response of community species diversity to environmental factors is mainly concentrated on the first axis, which is negatively correlated with the degree of surface water disturbance, soil moisture content and conductivity, and positively associated with other environmental factors. Among them, this was significant in the degree of surface water disturbance, depth of groundwater, and soil moisture and organic matter. Moving from left to right along the first axis, the degree of surface water disturbance and soil moisture in the explanatory variables gradually decreased. In contrast, the depth of groundwater and organic matter steadily increased. Changes in diversity characteristics also accompany changes in community type along the axis. According to the species diversity characteristics of different community types in Fig. 4, the species richness and Shannon–Wiener diversity index in this moving mode tended to decrease, and the Simpson diversity index to increase and then decrease gradually. The Pielou evenness index showed a gradual increase.
The hierarchical partitioning results of CCA showed that the depth to groundwater and surface water disturbance were the two variables with the highest individual contribution among all environmental variables, accounting for 13.1% (P = 0.005) and 5.8% (P = 0.050), respectively, key factors for species diversity pattern of oasis communities (Table 2).
Effect test of different water resources based on generalized linear model
In the multiclass logistic regression model of community types and different water resources, 12 formulas can be constructed according to different reference variables. The regression coefficients of soil water content were significant in eight of these equations, the regression coefficients of groundwater depth significant in four equations, while those of degree of surface water disturbance were significant in two equations (Table S1). Among all the significant regression coefficients, soil water content accounted for 54.1%, groundwater depth for 28.6%, and surface water disturbance for 14.3% (Fig. 7). Therefore, it is verified that soil water is the dominant factor in the division of community types. The overall final prediction accuracy of the model is high at 71.8%.

Logistic regression model between community types and different water resources (McFadden R2 = 0.433, P < 0.05). Note: numbers in the matrix represent the regression coefficients
In the generalized linear regression models developed to examine the relationship between species diversity and different water resources (Table S2, Fig. 8), groundwater depth was significant affecting both species richness and Shannon–Wiener diversity index; surface water disturbance was a significant factor affecting Pielou evenness index. These results validate the driving role of these two environmental variables in the diversity patterns. However, different water resources did not have a significant effect on the Simpson index (P > 0.05).

Generalized linear regression model between species diversity and different water resources (*: P < 0.05), showing the relative contribution of different water resources after hierarchical partitioning; abbreviations of the environmental variables are shown in Fig. 5
Discussion
Relationship between community type and species diversity
Community ecology has consistently prioritized identifying the mechanisms of species coexistence and biodiversity maintenance within a certain spatial range (Kraft et al. 2015; Hart et al. 2017). In desert oases, different community types reflect differences in species composition and dominant species status and ultimately lead to distinct species diversity patterns due to biological attributes (Chesson 2000a; Uriarte and Menge 2018) or to abiotic factors (Zeng et al. 2020). The results using different indices for individual community types were inconsistent with the comparison results of diversity. For example, community Type 1, Phragmites australis + Tamarix chinensis, had the highest Shannon–Wiener index but it was still lower than that of Type 2, Tamarix chinensis + Karelinia caspia type in the Simpson index, primarily because the Shannon–Wiener index was more sensitive to richness, while the Simpson index was more sensitive to evenness of species distribution (Nagendra 2002). In this study area, P. australis was highly tolerant to flooding and to high salinity. Most communities using the Pielou evenness to identify the dominant species tend to be extremely uneven in species distribution, resulting in significantly lower Pielou evenness than other community types.
Community types 3 and 4 with P. euphratica and T. chinensis as dominant species showed lower diversity and higher evenness. Studies have shown that P. euphratica has a higher efficiency for the utilization of shallow soil water than T. chinensis, and that T. chinensis has a higher utilization efficiency of deep soil water and groundwater than P. euphratica (Chen et al. 2017). There are differences in niche and average fitness between the two in mixed forests (Chesson 2000b). The stabilization and equalizing mechanisms jointly determine the coexistence of the two and can maintain a specific dominant position under multiple environmental conditions. In Types 3 and 4 communities, neither P. euphratica nor T. chinensis had were densely distributed, thereby averaging the survival weights of the dominant species to a some extent. This is the main reason for the higher evenness of these two community types.
Significance of soil moisture content on the formation and distribution of community types
The Mantel test and logistic regression between community types and environmental factors showed that soil water contents played an important role in community division, which could be a strong correlation between species dominance and soil water content differences. Tamea et al. (2009) believed that in arid and semi-arid ecosystems, the interaction between groundwater and vegetation is achieved through the soil and that soil water plays a crucial role in the dynamics of ecosystems. The correlation between environmental factors in Fig. 4 indicate that the content of soil water tends to decrease with the groundwater depth and increases with the increase in surface water disturbance, which is consistent with the results of many studies on the Tarim River (Chen et al. 2008; Hao et al. 2010). Additionally, studies have shown a moisture redistribution process in arid soils. This complex mechanism can improve the availability of moisture, beneficial for the growth of herbaceous plants (Yu et al. 2013; Zhang et al. 2018). This process is not only affected by seasonal floods and groundwater levels but is also closely related to environmental conditions such as soil physical properties. By analyzing the floodplain ecosystem in the Tarim Basin, Grashey-Jansen et al. (2014) believed that the spatial difference in soil moisture might be the decisive factor for the zonal distribution difference of non-zonal vegetation. This study verifies this observation but also found that surface water disturbance plays an essential role in creating community differences in the Phragmites australis + Tamarix chinensis (Type 1), and both soil moisture content and surface water have profound effects on this community type.
Significance of groundwater depth to the protection of the oasis community
From the perspective of species diversity in desert oases as an entry point, it was discovered that the dominant factor driving the diversity was not soil water but groundwater, corroborating the results of several studies (Hao et al. 2009; Ye et al. 2010; Zhu et al. 2013). The reason behind such a phenomenon is the division of community types being based on the degree of dominance as the research carrier, and the individual growth of different species and their biological characteristics that occupy the main weight in this research. In contrast, species diversity, as an indicator of community characteristics, holistically incorporates all the species for analysis, and considers the current composition and distribution of species. Although the two studies are on community ecology, the research process was not carried out at the same scale. Therefore, we believe that in desert wetland ecosystems formed at the ends of arid inland rivers, soil water and groundwater play different roles in maintaining the existing communities. Soil water determines the dominance of species in diverse communities by improving the availability of surface water and groundwater, and differences in groundwater depth determine the species composition and distribution of other communities. It is important to note that the contribution of soil water depends on the recharge of surface water and dynamic changes in groundwater depth, and cannot constrain plant growth alone (Ye et al. 2020; Malik et al. 2021). Therefore, considering the evolution of surface processes in arid areas, strengthening the protection of groundwater depth can play a positive role in promoting and restoring vegetation. In the analysis of hierarchical partitioning and the generalized linear model, this study also found that the degree of surface water disturbance has an important influence on the Pielou evenness index. Based on this, further research is required to understand how to combine existing negative correlation between surface water disturbance and groundwater depth to establish an ecological water requirement threshold that considers both diversity and evenness.
Conclusions
Quantification of the environmental factors and community characteristics of oases helped clarify the response of desert-wetland community classification and species composition. The results show that the division of oasis community types is primarily controlled by soil moisture. Based on the dependence on surface water and groundwater, this water redistribution process profoundly impacts the presence or absence of species at the community scale and the formation of dominant species. Moreover, the degree of surface water disturbance also plays an essential role in forming community differences within the Phragmites australis + Tamarix chinensis (Type1). Individual community types showed significant differences in species diversity. In contrast to the degree of dominance, groundwater depth was the key factor driving the pattern of species diversity, followed by surface water disturbance. The generalized linear model found that the depth of groundwater affects patterns of species diversity which are sensitive to richness (species richness and Shannon–Wiener index), while the degree of surface water disturbance affects the pattern of species evenness (Pielou index). This study has shown that different sources of water resources have different roles in desert-wetland ecosystems. A single factor did not dominate the characteristics of the oasis communities at different scales. From our study, it is concluded that rational optimization of water resource allocation plays a positive role in maintaining the sustainability of oases.
References
Blanchard G, Munoz F, Ibanez T, Hequet V, Vandrot H, Girardi J, Birnbaum P (2019) Regional rainfall and local topography jointly drive tree community assembly in lowland tropical forests of New Caledonia. J Veg Sci 30:845–856. https://doi.org/10.1111/jvs.12781
Chen YN, Pang ZH, Chen YP, Li WH, Xu CC, Hao XM, Huang X, Huang TM, Ye ZX (2008) Response of riparian vegetation to water-table changes in the lower reaches of Tarim River, Xinjiang Uygur, China. Hydrogeol J 16:1371–1379. https://doi.org/10.1007/s10040-008-0306-1
Chen YN, Li WH, Zhou HH, Chen YP, Hao XM, Fu AH, Ma JX (2017) Experimental study on water transport observations of desert riparian forests in the lower reaches of the Tarim River in China. Int J Biometeorol 61:1055–1062. https://doi.org/10.1007/s00484-016-1285-x
Chen GG, Yue DX, Zhou YY, Wang D, Wang H, Hui C, Guo JJ (2021) Driving factors of community-level plant functional traits and species distributions in the desert-wetland ecosystem of the Shule River Basin, China. Land Degrad Dev 32:323–337. https://doi.org/10.1002/ldr.3624
Chesson P (2000a) General theory of competitive coexistence in spatially-varying environments. Theor Popul Biol 58:211–237. https://doi.org/10.1006/tpbi.2000.1486
Chesson P (2000b) Mechanisms of maintenance of species diversity. Annu Rev Ecol Evol S 31:343–366. https://doi.org/10.1146/annurev.ecolsys.31.1.343
Gong YM, Lv GH, Guo ZJ, Chen Y, Cao J (2017) Influence of aridity and salinity on plant nutrients scales up from species to community level in a desert ecosystem. Sci Rep 7:6811. https://doi.org/10.1038/s41598-017-07240-6
Grashey-Jansen S, Kuba M, Cyffka B, Halik Ü, Aishan T (2014) Spatio-temporal variability of soil water at three seasonal floodplain sites: A case study in Tarim Basin, Northwest China. Chin Geogr Sci 24:647–657. https://doi.org/10.1007/s11769-014-0717-y
Hao XM, Chen YN, Li WH (2009) Indicating appropriate groundwater tables for desert river-bank forest at the Tarim River, Xinjiang, China. Environ Monit Assess 152:167–177. https://doi.org/10.1007/s10661-008-0305-7
Hao XM, Li WH, Huang X, Zhu CG, Ma JX (2010) Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China. Hydrol Process 24:178–186. https://doi.org/10.1002/hyp.7432
Hart SP, Usinowicz J, Levine JM (2017) The spatial scales of species coexistence. Nat Ecol Evol 1:1066–1073. https://doi.org/10.1038/s41559-017-0230-7
Keyimu M, Halik Ü, Betz F, Dulamsuren C (2018) Vitality variation and population structure of a riparian forest in the lower reaches of the Tarim River, NW China. J Foreatry Res 29:749–760. https://doi.org/10.1007/s11676-017-0478-4
Khan N, Shaukat SS, Ahmed M, Siddiqui MF (2013) Vegetation-environment relationships in the forests of Chitral district Hindukush range of Pakistan. J Foreatry Res 24:205–216. https://doi.org/10.1007/s11676-013-0346-9
Kraft NJ, Godoy O, Levine JM (2015) Plant functional traits and the multidimensional nature of species coexistence. P Natl Acad Sci U S A 112:797–802. https://doi.org/10.1073/pnas.1413650112
Lai JS, Zou Y, Zhang JL, Peres-Neto PR (2022) Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol Evol 00:1–7. https://doi.org/10.1111/2041-210X.13800
Lamontagne S, Cook PG, O’Grady A, Eamus D (2005) Groundwater use by vegetation in a tropical savanna riparian zone (Daly River, Australia). J Hydrol 310:280–293. https://doi.org/10.1016/j.jhydrol.2005.01.009
Lei Y, Li XQ, Ling HB (2015) Model for calculating suitable scales of oases in a continental river basin located in an extremely arid region, China. Environ Earth Sci 73:571–580. https://doi.org/10.1007/s12665-014-3328-0
Li JL, Feng Q, Guo QL (2008) Fractal study of sustainable proportions of natural and artificial oases. Environ Geol 55:1389–1396. https://doi.org/10.1007/s00254-007-1089-8
Li X, Yang K, Zhou YZ (2016) Progress in the study of oasis-desert interactions. Agric for Meteorol 230–231:1–7. https://doi.org/10.1016/j.agrformet.2016.08.022
Li H, Shi QD, Wan YB, Shi HB, Imin B (2021) Influence of surface water on desert vegetation expansion at the landscape scale: a case study of the Daliyabuyi Oasis, Taklamakan Desert. Sustainability 13:9522. https://doi.org/10.3390/su13179522
Ling HB, Xu HL, Guo B, Deng XY, Zhang P, Wang XY (2019) Regulating water disturbance for mitigating drought stress to conserve and restore a desert riparian forest ecosystem. J Hydrol 572:659–670. https://doi.org/10.1016/j.jhydrol.2019.03.049
Lu J, Li ZQ, Gao T, Tang XQ (2020) Seed bank characteristics in a Pinus densata forest and its relationship with vegetation diversity in Southeast Tibet, China. Ecol Evol 10:9214–9222. https://doi.org/10.1002/ece3.6603
Ma W, Lei S, Sun YJ, Grabosky J (2020) Forest succession in post-agricultural Larix olgensis plantations in northeast China. J for Res 31:2495–2505. https://doi.org/10.1007/s11676-019-00960-7
Malik MS, Shukla JP, Mishra S (2021) Effect of groundwater level on soil moisture, soil temperature and surface temperature. J Indian Soc Remote Sens 49:2143–2161. https://doi.org/10.1007/s12524-021-01379-6
Moradi G, Vacik H (2018) Relationship between vegetation types, soil and topography in southern forests of Iran. J Foreatry Res 29:1635–1644. https://doi.org/10.1007/s11676-018-0601-1
Muscarella R, Uriarte M, Erickson DL, Swenson NG, Kress WJ, Zimmerman JK (2016) Variation of tropical forest assembly processes across regional environmental gradients. Perspect Plant Ecol 23:52–62. https://doi.org/10.1016/j.ppees.2016.09.007
Nagendra H (2002) Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Appl Geogr 22:175–186. https://doi.org/10.1016/S0143-6228(02)00002-4
Nargis N, Mansoor H, Tahira N, Sajid AAM, Muhammad A (2013) Soil-plant relationships in the arid saline desert of Cholistan. Arid Land Res Manag 27:140–152. https://doi.org/10.1080/15324982.2012.719576
Olea PP, Mateo-Tomás P, de Frutos Á (2010) Estimating and modelling bias of the hierarchical partitioning public-domain software: Implications in environmental management and conservation. PLoS ONE 5:e11698. https://doi.org/10.1371/journal.pone.0011698
Palmer MW, McGlinn DJ, Westerberg L, Milberg P (2008) Indices for detecting differences in species composition: Some simplifications of RDA and CCA. Ecology 89:1769–1771. https://doi.org/10.1890/07-0842.1
Pankova EI (2008) Environmental conditions and soils of natural oases in the Alashan Gobi Desert, Mongolia. Eurasian Soil Sc 41:827–836. https://doi.org/10.1134/S1064229308080048
Qi SZ, Li XY, Duan HP (2007) Oasis land-use change and its environmental impact in Jinta Oasis, arid northwestern China. Environ Monit Assess 134:313–320. https://doi.org/10.1007/s10661-007-9622-5
Qiao XN, Yang DG, Zhang XH (2009) Evolution stages of oasis economy and its dependence on natural resources in Tarim River Basin. Chin Geogr Sci 19:135–143. https://doi.org/10.1007/s11769-009-0135-8
Shi HB, Shi QD, Zhou XL, Imin B, Li H, Zhang WQ, Kahaer Y (2021) Effect of the competition mechanism of between co-dominant species on the ecological characteristics of Populus euphratica under a water gradient in a desert oasis. Glob Ecol Conserv 27:e01611. https://doi.org/10.1016/j.gecco.2021.e01611
Sun JJ, Guo JH, Shen AH, Xu X, Feng HL, Zhang S, Yuan WG, Jiang B, Wu CP, Wang WF (2020) Composition and environmental interpretation of the communities of Sassafras tzumu, a protected species, at Zhejiang province in eastern China. Glob Ecol Conserv 24:e01218. https://doi.org/10.1016/j.gecco.2020.e01218
Tamea S, Laio F, Ridolfi L, D’Odorico P, Rodriguez-Iturbe I (2009) Ecohydrology of groundwater-dependent ecosystems: 2. Stochastic soil moisture dynamics. Water Resour Res 45:W05420. https://doi.org/10.1029/2008WR007293
Thammanu S, Marod D, Han H, Bhusal N, Asanok L, Ketdee P, Gaewsingha N, Lee S, Chung J (2021) The influence of environmental factors on species composition and distribution in a community forest in Northern Thailand. J Foreatry Res 32:649–662. https://doi.org/10.1007/s11676-020-01239-y
Uriarte M, Menge D (2018) Variation between individuals fosters regional species coexistence. Ecol Lett 21:1496–1504. https://doi.org/10.1111/ele.13130
Wan YB, Shi QD, Dai Y, Marhaba N, Peng LP, Peng L, Shi HB (2022) Water use characteristics of Populus euphratica Oliv. and Tamarix chinensis Lour. at different growth stages in a desert oasis. Forests 13:236. https://doi.org/10.3390/f13020236
Wang GH, Munson SM, Yu KL, Chen N, Gou QQ (2020) Ecological effects of establishing a 40-year oasis protection system in a northwestern China desert. CATENA 187:104374. https://doi.org/10.1016/j.catena.2019.104374
Wang J, Xue LQ, Liu YH, Ni T, Wu YB, Yang MJ, Han Q, Bai QY, Li XH (2021) The analytical indicators to explain the distribution of oases in arid zones using the oases integrated analysis model. Ecol Indic 127:107763. https://doi.org/10.1016/j.ecolind.2021.107763
Xie HL, Zhang YW, Wu ZL, Lv TG (2020) A bibliometric analysis on land degradation: current status, development, and future directions. Land 9:28. https://doi.org/10.3390/land9010028
Xu YP, Xiang ZL, Rizo EZ, Naselli-Flores L, Han BP (2022) Combination of linear and nonlinear multivariate approaches effectively uncover responses of phytoplankton communities to environmental changes at regional scale. J Environ Manag 305:114399. https://doi.org/10.1016/j.jenvman.2021.114399
Ye ZX, Chen YN, Li WH (2010) Ecological water demand of natural vegetation in the lower Tarim River. J Geogr Sci 20:261–272. https://doi.org/10.1007/s11442-010-0261-3
Ye C, Butler OM, Chen CG, Liu WZ, Du M, Zhang QF (2020) Shifts in characteristics of the plant-soil system associated with flooding and revegetation in the riparian zone of Three Gorges Reservoir, China. Geoderma 361:114015. https://doi.org/10.1016/j.geoderma.2019.114015
Yin XW, Feng Q, Li Y, Deo RC, Liu W, Zhu M, Zheng XJ, Liu R (2022) An interplay of soil salinization and groundwater degradation threatening coexistence of oasis-desert ecosystems. Sci Total Environ 806:150599. https://doi.org/10.1016/j.scitotenv.2021.150599
Yu TF, Feng Q, Si JH, Xi HY, Li ZX, Chen AF (2013) Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China’s extremely arid region. Plant Soil 372:297–308. https://doi.org/10.1007/s11104-013-1727-8
Yu WZ, Cui JT, Gao Y, Zhu MX, Shao L, Shen YB, Zhang XZ, Guo C, Zhang HXY (2022) Evolution of desertification types on the north shore of qinghai lake. CMC-Comput Mater Contin 71:3635–3646. https://doi.org/10.32604/cmc.2022.023195
Zeng Y, Zhao CY, Shi FZ, Schneider M, Lv GH, Li Y (2020) Impact of groundwater depth and soil salinity on riparian plant diversity and distribution in an arid area of China. Sci Rep 10:7272. https://doi.org/10.1038/s41598-020-64045-w
Zhang K, Yu Z, Li X, Zhou W, Zhang D (2007) Land use change and land degradation in China from 1991 to 2001. Land Degrad Dev 18:209–219. https://doi.org/10.1002/ldr.757
Zhang XL, Guan TY, Zhou JH, Cai WT, Gao NN, Du H, Jiang LH, Lai LM, Zheng YR (2018) Groundwater depth and soil properties are associated with variation in vegetation of a desert riparian ecosystem in an arid area of China. Forests 9:34. https://doi.org/10.3390/f9010034
Zhou HH, Chen YN, Zhu CG, Li Z, Fang GH, Li YP, Fu AH (2020) Climate change may accelerate the decline of desert riparian forest in the lower Tarim River, Northwestern China: Evidence from tree-rings of Populus euphratica. Ecol Indic 111:105997. https://doi.org/10.1016/j.ecolind.2019.105997
Zhu JT, Yu JJ, Wang P, Yu Q, Eamus D (2013) Distribution patterns of groundwater-dependent vegetation species diversity and their relationship to groundwater attributes in northwestern China. Ecohydrol 6:191–200. https://doi.org/10.1002/eco.1258
Acknowledgements
We thank all team members who collected the data and conducted the experimental analysis. We also thank the government of DaliYabuyi Township, Xinjiang, China, for their strong support for scientific research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This study was supported by grants from the National Natural Science Foundation of the China Joint Key Program (No. U1703237), Postgraduate Research Innovation Project in the Autonomous Region (No. XJ2022G017), and the National Natural Science Foundation of China Regional Program (No. 32160260).
The online version available at http://www.springerlink.com.
Corresponding editor: Tao Xu.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shi, H., Zhang, F., Shi, Q. et al. Responses of arid plant species diversity and composition to environmental factors. J. For. Res. 34, 1723–1734 (2023). https://doi.org/10.1007/s11676-023-01618-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-023-01618-1