
Metabolomic studies attempt to identify and profile unique metabolic differences among test populations, which may be correlated with a specific biological stress or pathophysiology. Due to the ease of collection and the metabolite-rich nature of urine, it is frequently used as a bio-fluid for human and animal metabolic studies. High-resolution 1H-NMR is an analytical tool used to qualitatively and quantitatively identify metabolites in urine. Urine samples were collected from healthy male and female subjects and prepared: raw, following centrifugation, filtration, or the addition of the bacteriostatic preservative sodium azide and analyzed by NMR. In addition, these samples were stored at room temperature (22 °C), in a refrigerator (4 °C), or in a deep-freeze (−80 °C). Samples were analyzed by NMR every week for a month and changes in concentrations of 55 easily identifiable metabolites were followed. The degree of change in metabolite concentrations following storage over a 4-week period were influenced by the different methods of sample preparation and storage. Significant changes in urine metabolites are likely due to bacterial contamination of the urine. Our study demonstrates that bacterial contamination of urine in normal individuals significantly alters the metabolic profile of urine over time and proper preparation and storage procedures must be followed to reduce these changes. By identifying appropriate methods of urine preparation and storage investigators will preserve the fidelity of the urine samples in order to better reflect the original metabolic state.
Similar content being viewed by others
Introduction
Metabolomic investigations attempt to detect and profile changes in metabolites, which reflect changes in metabolic pathways and may provide information concerning a disease state or the biological stress of an organism (Lindon et al., 2004; Weckwerth and Morgenthal, 2005). In order to follow human metabolic pathways, biological tissues or fluids are sampled from the subject or patient and analyzed for any changes in metabolite concentrations from baseline levels. Currently the metabolic analysis of a whole human subject is not possible so a sample of tissue or biofuild is used. Studies have analyzed many different biological fluids, such as urine, sputum, bronchoalveolar lavage, breath condensate, whole blood, serum, feces, and cerebral spinal fluid (Moolenaar et al., 2003; Bollard et al., 2005). Urine is a very popular biofluid for metabolomic investigations due to non-invasive collection, the complex metabolic nature of the fluid, and the ability to collect multiple samples over a period of time.
The detection of metabolites and the information gained by tracking metabolic flux provides information regarding an animals’ physiological response following an environmental insult or the pathophysiology of a disease. Many different methods of detection have been used for metabolomic studies, such as high performance liquid chromatography, high-resolution nuclear magnetic resonance (NMR), and mass spectrometry (Chadha et al., 2001; Lindon et al., 2004; Dunn et al., 2005). Recent technological advancements and studies have enhanced and defined the ability of 1H-NMR to accurately profile the metabolic makeup of a biofluid qualitatively and quantitatively (Griffiths and Irving, 1998).
The correlation of metabolite changes with a particular physiology is the basis for all metabolomic investigations. Many studies have found correlations of key metabolites with particular diseases, metabolites that may be used for diagnosis, or changes that may arise due to laboratory conditions of the study (Brindle et al., 2002; Coen et al., 2005; Sabatine et al., 2005). Human urine studies have found changes in metabolite concentrations as a result of age and diet, variation in urine metabolite concentrations within large populations, and variation of particular urine metabolites in individuals over time (Guneral and Bachmann, 1994; Dyer et al., 1997; Zuppi et al., 1997; Symanski and Greeson, 2002; Lenz et al., 2003, 2004; Bollard et al., 2005). Studies of animal urine have found metabolic differences within genetically similar animal models (Holmes et al., 2000; Tate et al., 2001). In addition, studies have found metabolic changes following the movement of animals from a sterile environment to one that is open to ambient pathogens (Nicholls et al., 2003). To ensure the fidelity of the biological sample it is clear that attention must be given to how a sample is prepared, and stored prior to analysis.
Our study documents the effect sample handling has upon the metabolic make-up of urine. Different methods of sample preparation (centrifugation, filtration, or addition of the preservative sodium azide) as well as sample storage (room temperature (22°°C), refrigerator (4 °C), or the deep-freeze (−80 °C)) have significant effect upon the changes of urine metabolites over time, as detected by 1H-NMR. By outlining proper sample handling and storage techniques for urine samples further metabolomic studies may ensure the sample reflects the original metabolic state of the subject.
Materials and methods
Urine collection
One healthy female and male volunteer provided urine samples in accordance with guidelines established by the University of Alberta Health Research Ethics Board. Mid-stream urine samples were collected in the morning and immediately prepared as outlined below.
Sample preparation
Sample preparation was duplicated for both male and female urine samples and in triplicate for the three different methods of storage. In a biosafety fume hood fresh urine samples were prepared by transferring a 630 μL aliquot of urine to a 1.5 mL Eppendorff tube followed by the addition of 70 μL of a standard solution (4.9 mM DSS (disodium-2,2–dimethyl-2-silapentane-5-sulphonate), and 100.0 mM imidazole in D2O, Sigma-Aldrich, Mississauga, ON). Filtered urine samples were prepared by filtering 1 mL of urine through a 0.22 μm syringe filter (Millipore, Cambridge, Ont. Canada), a 630 μL aliquot of the filtered urine was transferred to a new Eppendorff tube and 70 μL of the standard solution was added (see above). Spun urine samples were prepared by the centrifugation of 1 mL of urine in an Eppendorff tube at 10,000 rpm for 10 min (MSE Microcentaur, Sanyo-Gallenkamp), a 630 μL aliquot of the spun urine was placed in a new Eppendorff followed by the addition of 70 μL of the standard solution (see above). For urine samples with the preservative sodium azide, 1 mL of raw urine was transferred to a 1.5 mL Eppendorff tube followed by the addition of a stock solution of sodium azide (Sigma Aldrich) in order to reach a final sample concentration of 0.1, 1.0, and 10 mM sodium azide. A 630 μL aliquot of the urine and sodium azide solution was transferred to a 1.5 mL Eppendorff tube followed by the addition of 70 μL of the standard solution (see above). For each of the urine samples (fresh, spun, filtered, and each of the samples with sodium azide) a final aliquot of 600 μL was transferred to a standard 5 mm glass NMR tube (Wilmad, NJ, USA).
Sample storage
Urine samples for each of the preparatory methods above (in triplicate) were stored in the 5 mm NMR tubes at room temperature (22 °C), a refrigerator (4 °C), or a deep-freeze (−80 °C) for the 4-week duration of the study. Once a week samples were removed from storage and allowed to equilibrate to room temperature (roughly 1 h) prior to NMR data acquisition.
In order to investigate the effects of freeze–thaw cycles on metabolites found in the urine additional samples were prepared from the male and female subjects. Male and female urine samples were prepared as raw urine and another with 10 mM azide (see sample preparation outlined above); both were stored at −80 °C and thawed twice a week (left at room temperature for 1 h, then returned to freezer).
NMR Analysis
All 1H-NMR spectra were acquired on a 500 MHz Inova (Varian Inc, Palo Alto, CA) spectrometer equipped with a 5 mm triple resonance probe with z-axis pulsed field gradients. One-dimensional 1H-NMR spectra were collected at 25 °C using a standard pre-saturation pulse sequence (one-dimensional, transmitter pre-saturation delay of 1 s for water suppression, followed by a 8.4 μs 90o read pulse), and a spectral width of 8000 Hz (Hoult, 1976). The time-domain data points were 64 k, acquisition time was 4 s, repetition time was 5 s, four steady state scans, and the number of acquired scans was 128. The FID was apodized with an exponential window function corresponding to a line broadening of 0.5 Hz and Fourier transformed.
NMR quantification
The methyl singlet of the buffer constituent DSS served as internal standard for chemical shifts (set to 0 ppm), and for quantification. Spectral quantification of 55 clearly identifiable metabolites were chosen for quantification using the Chenomx NMR Suite Professional software package version 3.1 (database available at pH 7.0, Chenomx Inc., Edmonton, AB, Canada). For a list of the metabolites identified by NMR refer to table 1. For qualitative and quantitative analysis Chenomx NMR Suite software takes a database of pure compounds (metabolites) and compares the spectral signatures to those found in the urine spectrum. Reference spectra stored in the software are fit to the urine spectra in order to quantify selected metabolites. The internal DSS signal is utilized as the concentration reference (0.49 mM). Work in our laboratory has demonstrated that this procedure provides absolute concentration accuracies of 90% or better.
Typically urine metabolites are reported as ratios with the metabolite creatinine. As a result of the changing concentration of creatinine over long-term storage in this study all metabolites are reported as absolute concentrations (mM). The metabolite urea is not listed in table 1. Caution must be used when drawing any metabolic conclusions based upon changes in the concentration of urea. Solvent suppression by pre-saturation (pulse sequence) lends to the likelihood of resonant suppression of the urea peak due to proton exchange with water. As a result urea is not included as a metabolite, but allowing for this caveat, it will be discussed later as a potential metabolite of interest.
Statistical analysis
Differences between variables were completed using t-tests and analysis of variance (ANOVA). A p-value of <0.05 was considered significant.
Results and discussion
Urine has been studied for many years as a fluid through which researchers may gain insight into the metabolic processes of an organism. Studies have looked at different properties of urine; bactericidal factors (Kaye, 1968; Griffith et al., 1976), precipitation potentials (Udert et al., 2003b), the array of metabolites present (Armstrong et al., 1956), even the effect of different preservatives in urine collection tubes (Nickander et al., 1982; Allen et al., 1987). Such studies have provided a basic understanding of urine as a biofluid that may be utilized in the laboratory for a number of investigative studies concerning changes in an organisms’ metabolism. However, before any differences in urine metabolites are correlated to a biological response it is important to address the cause of the changing metabolite concentrations. Our study investigates the influence laboratory sample preparation and storage may have on the profile of metabolites in a normal urine sample.
Gender differences
Differences were noted in the metabolites that changed for male and female urine samples over the 4-week period. Of the 55 metabolites we chose to follow in female urine acetate, benzoate, creatine, glycine, lactate, malonate, succinate, trimethylamine, and formate increased, while creatinine, urea, guanidinoacetate, hippurate and citrate decreased over the four weeks. Table 2 lists changes in female urine metabolite concentrations after a 4-week period of storage at room temperature (22 °C).
In the male urine creatinine, creatine, and phenylacetylglycine were the only metabolites to change significantly following storage at room temperature (22 °C) over 4 weeks. Table 3 lists the metabolites in the male urine that changed significantly.
Urine preparation
This study was concerned with maintaining the composition of metabolites in a normal urine sample over a period of time. The efficacy of each preparatory step was determined according to the fidelity of the metabolite profile over the 4-week period. Table 2 shows the effect urine preparation has on the degree of metabolite change of female urine over time. Spinning the urine in a counter top centrifuge reduced the degree of change of metabolites over the 4 weeks, but filtering the urine produced a greater reduction in the degree of metabolite change. Addition of increasing concentrations of the preservative sodium azide (0.1, 1, and 10 mM) had a correlated rise in efficacy of inhibition of metabolite changes over a four-week period. Both male and female urine samples with 10 mM of sodium azide demonstrated the greatest degree of metabolite stability (Tables 2 and 3). Across all methods of urine preparation the male urine had a smaller degree of change in metabolites when compared to the female urine.
Figure 1 shows the change in absolute concentrations of three random metabolites (benzoate, creatine, and lactate) in female urine that was prepared raw, or following filtration and stored at room temperature (22 °C) for four weeks. Benzoate and lactate demonstrated a dramatic increase in concentration within the first week of storage, followed by only a slight increase over the next three weeks for raw urine. However, following filtration benzoate and lactate did not change significantly over the 4 weeks. Both raw and filtered female urine showed a steady increase in creatine concentration over four weeks of storage at room temperature.

Absolute concentrations of three random metabolites; benzoate, creatine, and lactate, in fresh (solid line) and filtered (dotted line) normal female urine stored at room temperature (22 °C) over a 4-week period.
Urine storage
Human urine collection for research studies sometimes occurs at the residence of the volunteer, but particularly in clinical practice sample collection takes place at a clinic or hospital setting so it is important to determine proper urine storage techniques to ensure accurate metabolite analysis at a later time. Urine samples were stored at room temperature (22 °C), in a refrigerator (4 °C), and a deep-freeze (−80 °C) for a 4-week period. Table 4 presents the change in raw female urine metabolite concentrations relative to the method of storage over a 4-week period. Both female and male urine samples stored at room temperature for 4 weeks had a significant change in a number of metabolites. Storage of the urine in a refrigerator (4 °C) produced a slight reduction in the degree of metabolite change, but storage in the deep-freeze (−80 °C) provided urine with a metabolite profile that best reflected the original metabolite concentrations (comparing raw urine only, table 4). The raw urine samples that underwent repeated cycles of freeze/thaw over the 4-weeks had an intermediate degree of metabolite change when compared with raw urine stored at room temperature and in the deep-freeze (table 4).
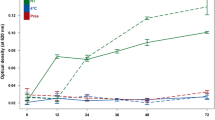
Figure 2 shows changes in absolute concentrations of three randomly selected metabolites in raw female urine over a 4-week period of storage at either room temperature or in a deep-freeze. The metabolites citrate and glycine had a slight decrease in concentration when stored in the deep-freeze, but when stored at room temperature citrate showed a significant and linear decrease in concentration, while glycine had a significant increase in concentration over the 4 weeks. The metabolite hippurate remained fairly constant when stored in the deep-freeze, but when raw female urine was stored at room temperature hippurate dropped immediately within 1 week and then remained unchanged.

Concentration of citrate, hippurate, and glycine in fresh female urine stored at room temperature (solid line, 22 °C) and in a deep-freeze (dotted line, −80 °C) over a 4-week period.
Similar differences in the rate and degree of metabolite change were present in female urine that was spun on the countertop centrifuge. Figure 3 shows absolute concentration changes for three randomly chosen metabolites as identified in spun female urine and stored at either room temperature or in a deep-freeze. Benzoate and lactate showed significant increases in concentration during the first 2 weeks of storage at room temperature, with a reduction in the rate of change during the last 2 weeks of analysis. Creatine demonstrated a linear increase in concentration throughout the 4 weeks of storage at room temperature. For all three metabolites there was no significant change in absolute concentration when spun female urine was stored in the deep-freeze over 4 weeks.

Absolute concentrations for three random metabolites; benzoate, creatine, and lactate, measured in centrifuged female urine stored at room temperature (solid line, 22 °C) and in a deep-freeze (dotted line, −80 °C) over a four-week period.
In our study female urine benzoate concentrations increased significantly over time (table 2 and figure 3). Studies have found that when laboratory animals are moved from sterile to ambient-air environments there is an associated increase in the presence of benzoate in the urine (Hansen et al., 1972; Goodwin et al., 1994; Nicholls et al., 2003). This increase has been correlated with increased bacterial activity in the animals. In our study the increase in the formation of benzoate in female urine, and the reduction of benzoate formation in filtered urine, or urine with sodium azide suggests that bacteria in the urine may be the cause of such a change. In addition, it is known that some bacterial species are able to metabolize hippurate to benzoic acid (Hansen et al., 1972). Our study found increased concentrations of benzoic acid with a decrease in hippurate in the female urine, again, this is suggestive of bacterial activity in the sample (table 2, figures 2 and 3).
Over the 4-week period raw female urine demonstrated a significant increase in the concentration of the metabolite trimethylamine (tables 2 and 4). Previous studies have shown that trimethylamine is produced by the bacterial breakdown of dietary choline (Brewster and Schedewie, 1983; Zeisel et al., 1985, 1989). As well, bacteria are known to produce formic acid from the substrate citrate (Schiwara et al., 1992). In our present study, raw female urine showed a large increase in formic acid with a reduction in the metabolite citrate. The correlated rise of formate and fall of citrate concentrations in the female urine stored over 4 weeks suggests bacterial activity in the female urine. The limiting of these absolute concentration changes by particular sample preparation (i.e. filtering) and storage (i.e. deep-freeze) techniques leads to the conclusion that such measures should be taken by others using urine as a biofluid for metabolomic studies.
The concentration of creatinine fell for both the male and female urine samples, while the concentration of creatine increased. Studies have shown that creatinine in the urine can be used by bacteria and will be converted by the enzyme creatininase to creatine (Cattell et al., 1969; Jones and Burnett, 1972). The coordinated reduction of creatinine with an increase in creatine concentration is suggestive of bacterial activity in both male and female urine samples. Figure 1 shows that filtering female urine had a limited reduction in the degree of change in creatine concentration over 4 weeks (stored at room temperature). It is known that creatinine is formed non-enzymatically from creatine over a period of time. However, our data shows that creatinine concentrations fell while creatine concentrations grew. This may provide further evidence of the predominantly bacterial influence on metabolite levels over time as opposed to non-enzymatic reactions.
Similar to creatinine, urea is a metabolite found in urine that is rapidly broken-down by bacteria. Numerous investigations have looked at the breakdown of urea by bacteria; the formation of urinary stones from urease-producing bacteria (Kaya et al., 2003), the blockage of urine-collecting systems (Udert et al., 2003a), and breath tests for H. pylori detection in gastric ulcer patients (Pathak et al., 2004a, b). Our study showed a reduction in the concentration of urea in normal urine over time. This reduction in urea concentration was ameliorated somewhat by centrifugation, filtration, and the addition of the preservative sodium azide when compared to raw urine. Storage in a refrigerator or a deep-freeze (−80 °C) limited the loss of urea somewhat, but no storage method was able to halt the significant reduction in urea.
The absolute concentrations for succinate and malonate increased in female urine over the 4-week period (tables 2 and 4). Although sample preparation steps reduced the rise in the metabolites, there remained a significant increase in these two metabolites over long-term storage. Other studies have shown that these metabolites increase over time as a result of the activity of bacteria in the urine (Brenner et al., 1986; Nicholls et al., 2003; Stenina et al., 2003).
The metabolite lactate also increased over time. Lactate increased in the female urine over time and once again may be the result of metabolic activity of bacteria in the urine. Processing of the urine and storage in the refrigerator or deep-freeze helped to reduce the rise in lactate when compared to raw urine at room temperature (table 4). An increase in lactate has been shown to correlate with bacterial activity in a number of pathophysiologies; such as bacteraemia (Bar-Meir et al., 2005), urinary tract infections (Tal et al., 2005), pneumonia (Utine et al., 2005), and meningitis (Lutsar et al., 1995; Nazifi et al., 1997).
Another metabolite that increased significantly in raw female urine was acetate. Storage in a refrigerator or a deep-freeze, or the filtration and the addition of sodium azide reduced the increase in acetate significantly (tables 2 and 4). Acetate, formate, lactate, and succinate are known to significantly rise during bacterial metabolism in media where carbohydrate substrates are present (D’Argenio and Mazzacca, 1999; Macfarlane and Macfarlane, 2003). Our study has shown that these metabolites increase over time and different methods of sample preparation and storage has an effect of reducing the amount of change in the urine metabolites. When considering the changing metabolites as a whole, the data suggests that bacterial activity in the urine is changing the metabolite profile of the original sample.
The presence of bacteria in human urine is known to the scientific and medical communities, for example urinary tract infections (Funfstuck et al., 1987; Peddie et al., 1996; Chambers et al., 1999), and the influence of gut microflora on metabolites found in urine (Bain et al., 1988). The metabolites that changed from the original urine samples over the 4-week period of this study are highly suggestive of bacterial contamination of the urine. Our study suggests that the presence of bacteria in normal urine samples can dramatically affect the metabolic profile of the urine and appropriate measures must be taken to preserve the sample.
Differences in the degree of metabolite change between male and female urine samples over time, as well as the number of changing metabolites are possibly due to the differences in the degree of bacterial contamination. It is possible that the urological differences between male and female urinary tracts allows for greater bacterial culture and urine contamination in female patients.
Our study focused primarily upon sample preparation and storage of normal urine samples. Urine samples were collected from a male and female volunteer. Due to biological variance observed in urine another extended study would be beneficial where a number of individuals are sampled and the effect of biological variability on the degree of metabolite change in stored urine over time may be determined.
Concluding remarks
Urine is an extremely attractive biofluid for metabolomic and metabonomic investigations due to the ease of sample collection and the metabolite rich nature of the fluid. However, as investigators identify and follow changes in urine metabolite concentrations it becomes increasingly important to define the origin of the observed differences. Our study has shown that appropriate storage and preparatory measures must be taken immediately following urine collection to ensure the metabolite composition of the sample is maintained throughout the collection and analytical process. The bacteriostatic preservative sodium azide should be added to all urine samples, the samples should be stored in a deep-freeze (−80 °C), and cycles of sample freeze-thaw should be avoided whenever possible. The filtration of urine provided additional preservation of original metabolite concentrations, likely due to the removal of bacterial contamination, but caution is advised in some cases due to the possibility of losing larger metabolites, such as some proteins, during the filtration process. Investigators must take appropriate measures to remove or inhibit bacterial influence upon the metabolic profile of the urine sample. Standard procedures for urine preparation and storage and the continued monitoring of any metabolite changes will ensure sample fidelity for future metabolomic investigations.
References
Allen T.A., Jones R.L., Purvance J., (1987) Microbiologic evaluation of canine urine: direct microscopic examination and preservation of specimen quality for culture J. Am. Vet. Med. Assoc. 190: 1289–1291
Armstrong M.D., Shaw K.N., Wall P.E., (1956) The phenolic acids of human urine; paper chromatography of phenolic acids J. Biol. Chem. 218: 293–303
Bain M.D., Jones M., Borriello S.P., Reed P.J., Tracey B.M., Chalmers R.A., Stacey T.E., (1988) Contribution of gut bacterial metabolism to human metabolic disease Lancet 1: 1078–1079
Bar-Meir M., Raveh D., Yinnon A.M., Benenson S., Rudensky B., Schlesinger Y., (2005) Non-Typhi Salmonella gastroenteritis in children presenting to the emergency department: characteristics of patients with associated bacteraemia Clin. Microbiol. Infect. 11: 651–655
Bollard M.E., Stanley E.G., Lindon J.C., Nicholson J.K., Holmes E., (2005) NMR-based metabonomic approaches for evaluating physiological influences on biofluid composition NMR Biomed. 18: 143–162
Brenner D.J., Mcwhorter A.C., Kai A., Steigerwalt A.G., Farmer J.J. 3rd (1986) Enterobacter asburiae sp. nov., a new species found in clinical specimens, and reassignment of Erwinia dissolvens and Erwinia nimipressuralis to the genus Enterobacter as Enterobacter dissolvens comb. nov. and Enterobacter nimipressuralis comb. nov J. Clin. Microbiol. 23: 1114–1120
Brewster M.A., Schedewie H., (1983) Trimethylaminuria Ann. Clin. Lab. Sci. 13: 20–24
Brindle J.T., Antti H., Holmes E., Tranter G., Nicholson J.K., Bethell H.W., Clarke S., Schofield P.M., Mckilligin E., Mosedale D.E., Grainger D.J., (2002) Rapid and noninvasive diagnosis of the presence and severity of coronary heart disease using 1H-NMR-based metabolomics Nat. Med. 8: 1439–1444
Cattell W.R., Sardeson J.M., Hutchinson E., O’grady F., (1969) Creatinine content of urine and bacterial growth Br. Med. J. 3: 175–176
Chadha V., Garg U., Alon U.S., (2001) Measurement of urinary concentration: a critical appraisal of methodologies Pediatr. Nephrol. 16: 374–382
Chambers S.T., Peddie B.A., Randall K., Lever M., (1999) Inhibitors of bacterial growth in urine: what is the role of betaines? Int. J. Antimicrob. Agents 11: 293–296
Coen M., O’sullivan M., Bubb W.A., Kuchel P.W., Sorrell T., (2005) Proton nuclear magnetic resonance-based metabonomics for rapid diagnosis of meningitis and ventriculitis Clin Infect Dis 41: 1582–1590
D’Argenio G., Mazzacca G., (1999) Short-chain fatty acid in the human colon. Relation to inflammatory bowel diseases and colon cancer Adv. Exp. Med. Biol. 472: 149–158
Dunn W.B., Bailey N.J., Johnson H.E., (2005) Measuring the metabolome: current analytical technologies Analyst 130: 606–625
Dyer A., Elliott P., Chee D., Stamler J., (1997) Urinary biochemical markers of dietary intake in the INTERSALT study Am. J. Clin. Nutr. 65: 1246S–1253S
Funfstuck R., Stein G., Fuchs M., Bergner M., Wessel G., Keil E., Suss J., (1987) The influence of selected urinary constituents on the adhesion process of Escherichia coli to human uroepithelial cells Clin. Nephrol. 28: 244–9
Goodwin B.L., Ruthven C.R., Sandler M., (1994) Gut flora and the origin of some urinary aromatic phenolic compounds Biochem. Pharmacol. 47: 2294–7
Griffith D.P., Musher D.M., Itin C., (1976) Urease. The primary cause of infection-induced urinary stones Invest. Urol. 13: 346–50
Griffiths L., Irwing A.M. (1998) Assay by nuclear magnetic resonance spectroscopy: quantification limits Analyst 123: 1061–1068
Guneral F., Bachmann C., (1994) Age-related reference values for urinary organic acids in a healthy Turkish pediatric population Clin Chem 40: 862–866
Hansen S., Perry T.L., Lesk D., Gibson L., (1972) Urinary bacteria: potential source of some organic acidurias Clin Chim Acta 39: 71–74
Holmes E., Nicholls A.W., Lindon J.C., Connor S.C., Connelly J.C., Haselden J.N., Damment S.J., Spraul M., Neidig P., Nicholson J.K., (2000) Chemometric models for toxicity classification based on NMR spectra of biofluids Chem Res Toxicol 13: 471–478
Hoult D.I., (1976) Solvent peak saturation with single phase and quadrature Fourier transformation J. Magn. Reson. 21: 337–347
Jones J.D., Burnett P.C., (1972) Implication of creatinine and gut flora in the uremic syndrome: induction of “creatininase” in colon contents of the rat by dietary creatinine Clin Chem 18: 280–284
Kaya S., Poyraz O., Gokce G., Kilicarslan H., Kaya K., Ayan S., (2003) Role of genital mycoplasmata and other bacteria in urolithiasis Scand. J. Infect. Dis. 35: 315–317
Kaye D., (1968) Antibacterial activity of human urine J. Clin. Invest. 47: 2374–2390
Lenz E.M., Bright J., Wilson I.D., Hughes A., Morrisson J., Lindberg H., Lockton A., (2004) Metabonomics, dietary influences and cultural differences: a 1H NMR-based study of urine samples obtained from healthy British and Swedish subjects J. Pharm. Biomed. Anal. 36: 841–849
Lenz E.M., Bright J., Wilson I.D., Morgan S.R., Nash A.F., (2003) A 1H NMR-based metabonomic study of urine and plasma samples obtained from healthy human subjects J. Pharm. Biomed. Anal. 33: 1103–1115
Lindon J.C., Holmes E., Bollard M.E., Stanley E.G., Nicholson J.K., (2004) Metabonomics technologies and their applications in physiological monitoring, drug safety assessment and disease diagnosis Biomarkers 9: 1–31
Lutsar I., Gontmacher A., Narska M., Ruutel V., Topman M., Ilves P., Siirde T., Beilmann A., (1995) Five days of antibacterial therapy for bacterial meningitis in children? Infection 23: 113–118
Macfarlane S., Macfarlane G.T., (2003) Regulation of short-chain fatty acid production Proc Nutr Soc 62: 67–72
Moolenaar S.H., Engelke U., Wevers R.A. (2003) Proton nuclear magnetic resonance spectroscopy of body fluids in the field of inborn errors of metabolism Ann Clin Biochem 40: 16–24
Nazifi S., Rezakhani A., Badran M., (1997) Evaluation of hematological, serum biochemical and cerebrospinal fluid parameters in experimental bacterial meningitis in the calf Zentralbl Veterinarmed A 44: 55–63
Nicholls A.W., Mortishire-Smith R.J., Nicholson J.K., (2003) NMR spectroscopic-based metabonomic studies of urinary metabolite variation in acclimatizing germ-free rats Chem. Res. Toxicol. 16: 1395–1404
Nickander K.K., Shanholtzer C.J., Peterson L.R., (1982) Urine culture transport tubes: effect of sample volume on bacterial toxicity of the preservative J. Clin. Microbiol. 15: 593–595
Pathak C.M., Bhasin D.K., Khanduja K.L., (2004a) Urea breath test for Helicobacter pylori detection: present status Trop. Gastroenterol. 25: 156–161
Pathak C.M., Bhasin D.K., Pramod K.A., Khanduja K.L., (2004b) 14C-urea breath test as a ‘gold standard’ for detection of Helicobacter pylori infection Med Sci Monit 10: LE14–LE15
Peddie B.A., Chambers S.T., Lever M., (1996) Is the ability of urinary tract pathogens to accumulate glycine betaine a factor in the virulence of pathogenic strains? J. Lab. Clin. Med. 128: 417–422
Sabatine M.S., Liu E., Morrow D.A., Heller E., Mccarroll R., Wiegand R., Berriz G.F., Roth F.P., Gerszten R.E., (2005) Metabolomic identification of novel biomarkers of myocardial ischemia Circulation 112: 3868–3875
Schiwara H.W., Siegel H., Goebel A., (1992) Increase and decrease in formic acid concentration in urine samples stored at room temperature Eur. J. Clin. Chem. Clin. Biochem. 30: 75–79
Stenina M.A., Voevodin D.A., Stakhanov V.D., Kisilevich O.N., Rozanova G.N., (2003) Tissue hypoxia and intestinal dysbiosis in children with tuberculosis Bull. Exp. Biol. Med. 135: 178–180
Symanski E., Greeson N.M., (2002) Assessment of variability in biomonitoring data using a large database of biological measures of exposure AIHA J (Fairfax Va) 63: 390–401
Tal S., Guller V., Levi S., Bardenstein R., Berger D., Gurevich I., Gurevich A., (2005) Profile and prognosis of febrile elderly patients with bacteremic urinary tract infection J. Infect. 50: 296–305
Tate A.R., Damment S.J., Lindon J.C., (2001) Investigation of the metabolite variation in control rat urine using 1H NMR spectroscopy Anal. Biochem. 291: 17–26
Udert K.M., Larsen T.A., Biebow M., Gujer W., (2003a) Urea hydrolysis and precipitation dynamics in a urine-collecting system Water. Res. 37: 2571–2582
Udert K.M., Larsen T.A., Gujer W., (2003b) Estimating the precipitation potential in urine-collecting systems Water Res. 37: 2667–2677
Utine G.E., Ozcelik U., Yalcin E., Dogru D., Kiper N., Aslan A., Kanra G., (2005) Childhood parapneumonic effusions: biochemical and inflammatory markers Chest 128: 1436–1441
Weckwerth W., Morgenthal K., (2005) Metabolomics: from pattern recognition to biological interpretation Drug Discov. Today 10: 1551–1558
Zeisel S.H., Dacosta K.A., Fox J.G., (1985) Endogenous formation of dimethylamine Biochem. J. 232: 403–408
Zeisel S.H., Dacosta K.A., Youssef M., Hensey S., (1989) Conversion of dietary choline to trimethylamine and dimethylamine in rats: dose–response relationship J. Nutr. 119: 800–804
Zuppi C., Messana I., Forni F., Rossi C., Pennacchietti L., Ferrari F., Giardina B., (1997) 1H NMR spectra of normal urines: reference ranges of the major metabolites Clin Chim Acta 265: 85–97
Acknowledgments
This research was supported by the Canadian Institutes of Health Research, Genome Prairie, Genome Canada, the Natural Science and Engineering Research Council of Canada, and the University of Alberta.
The authors would like to thank Angela Thiessen for her assistance in sample preparation. We would also like to thank Carolyn Slupsky for her helpful discussions. This research was supported by the Canadian Institues of Health Research (CIHR), Genome Prairie, and Genome Canada. We would like to thank the Canadian National High Field NMR Centre (NANUC) for their assistance and use of the facilities. Operation of NANUC is funded by CIHR, the Natural Science and Engineering Research Council of Canada and the University of Alberta.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Saude, E.J., Sykes, B.D. Urine stability for metabolomic studies: effects of preparation and storage. Metabolomics 3, 19–27 (2007). https://doi.org/10.1007/s11306-006-0042-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11306-006-0042-2