
Abstract
Apelin, a member of the adipokine family, is a novel endogenous peptide which regulates the male reproductive system of mammals by interacting with a specific receptor. Recent studies have highlighted that apelin may play a role in the regulation of reproduction by reducing testosterone production and inhibiting LH secretion. To the best of our knowledge, there is no available data on the presence of the apelin and its receptor in canine testes. Therefore, the aim of this study was to reveal the presence of apelin and evaluate its distribution in the canine testes using immunohistochemical and RT-PCR techniques. For this purpose, five fertile and healthy male dogs were subjected to elective orchiectomy. The immunohistochemical reaction revealed the presence of apelin and its receptor in the canine testes. Apelin was localized in spermatids and spermatozoa with a positive signal in the “acrosomal bodies”. As regards the apelin receptor, a positive immunoreaction was detected in the cytoplasm of the cells localized near to the basal membrane of the seminiferous tubules and in the cytoplasm of Leydig cells. The RT-PCR analysis showed the presence of transcripts for apelin and apelin receptor in all of the samples under study. A 35kDa band confirmed apelin receptor protein expression in all of the samples analysed. In conclusion, the paracrine and endocrine role of apelin and its cognate receptor on male reproduction reported in humans and laboratory animals could also be hypothesized in dogs.
Similar content being viewed by others

Introduction
Adipose tissue is an active metabolic organ that requires intrinsic systems to maintain its function and health (Ferhat et al. 2019). It is widely known that it produces specific factors called adipokines such as chemerin, visfatin, adiponectin, leptin, resistin and apelin. These molecules can engage in complex communication between adipose tissue and other organs producing autocrine, paracrine or endocrine effects (Vázquez-Vela et al. 2008). The correlation between adipokines and reproduction has been reported in humans and several other species (Shokrollahi et al. 2021). Apelin (APLN) is an endogenous peptide that was originally isolated from bovine stomach extracts (Tatemoto et al. 2001). This pleiotropic molecule fulfills its function by binding to a G protein-coupled receptor (GPCR) called APJ (or APLNR) which is structurally similar to an angiotensin II receptor type 1 (Tatemoto et al. 2001). The peptide precursor of apelin is composed of a chain of 77 amino acids. Through enzymatic catalysis many molecular forms with different biological functions can be obtained from this apelin precursor: apelin-13, apelin-17, apelin-36, and pyroglutamate-apelin-13 (Tatemoto et al. 2001). The shorter forms of apelin have proved to possess greater biological potency than longer forms (Tatemoto et al. 2001). APLN and its receptor have been detected in many tissues of numerous mammalian species. In fact, transcript and protein for both molecules are expressed in specific hypothalamus and cerebroventricular regions of the brain, pituitary gland, skeletal muscle, kidney, spinal cord, thyroid gland, lungs, heart, adipose tissue and reproductive tract (Medhurst et al. 2003; Carpéné et al. 2007; Falcao-Pires et al. 2010; Mercati et al. 2018; Shokrollahi et al. 2022). In veterinary medicine, apelin was isolated from cow and sow ovaries (Roche et al. 2017) and from canine placenta (Troisi et al. 2020). It appears to play an important role in biological events such as folliculogenesis, cellular proliferation or apoptosis and the release of steroid hormones (Estienne et al. 2019). It is also involved in angiogenesis during pregnancy (Troisi et al. 2020). APLN and its receptor have recently been detected in the male gonads of human and laboratory animals (Medhurst et al. 2003; Estienne et al. 2019; Kawamata et al. 2001; Pope et al. 2012). More specifically, it has been demonstrated that the intracerebroventricular infusion of apelin in male rats reduces serum testosterone levels thus causing a drastic reduction in the number of testosterone-producing Leydig cells (Sandal et al. 2015). Therefore, it has been hypothesized that they may have an important role in male reproductive system function. To the best of our knowledge, there is no available data on the presence of apelin and its receptor in domestic animals. Therefore, the aim of this study was to highlight the presence and distribution of apelin and its receptor in canine testes using immunohistochemical and RT-PCR techniques.
Materials and methods
Animals
Testicular tissues were collected from 5 healthy mixed breed dogs (aged 2 to 5 years) with an average weight of 30kg. The dogs were admitted to the Veterinary Teaching Hospital (VTH) of the University of Perugia for elective orchiectomy with the written consent of their owners.
Surgical procedures
The orchiectomy was carried out under general anaesthesia according to the following protocol: premedication with methadone 0,2 − 0,4mg/kg IM (or IV) and medetomidine 1–5 mcg/kg; induction with preoxygenation and propofol 4–6mg/kg IV; anaesthesia for surgery maintained with isoflurane, adjusting the vaporiser setting according to anaesthetic depth; þ IV Ringer 10 ml/kg/h. A conventional surgical technique was used.
Tissue collection and processing
Following orchiectomy, the testicular tissue samples were promptly removed from all animals and thoroughly washed with saline. After dividing the tissue samples into small pieces under stereoscopic magnification, they were immediately sent for examination. For the RT-PCR, the samples were rinsed with RNase-free water then immediately frozen at -80C and were later evaluated for gene and protein expression. For the immunohistochemical analysis, some small pieces of testicular tissue were fixed by immersion in 4% (w/v) formaldehyde in PBS (pH 7.4) for 24h at room temperature and then processed following routine tissue preparation procedures (Dall’Aglio et al. 2014).
RT-PCR
Reagents
Deoxyribonuclease I (DNAse I Amp. Grade), Superscript III Reverse Transcriptase (Superscript III First-Strand Synthesis System) and DNA ladders were purchased from Life Technologies Italia (Monza, Monza Brianza, Italy). Reagents for the isolation and purification of total RNA (TRIzol), Taq DNA polymerase (Platinum), RNAse free tubes, water and deoxyNTPs, and primers for APLN and APLNR were also purchased from Life Technologies while NucleoSpin Gel and PCR clean up were supplied by Macherey-Nagel Inc (Bethlehem, PA, USA).
APLN and APLNR gene expression qualitative analysis
Total RNA was extracted from the testicular tissues (100mg each) of five male dogs. DNAse treatment was performed following the manufacturer’s instructions. Five micrograms of total RNA were reverse transcribed in 20 µL of Superscript III First-Strand Synthesis System using random hexamer according to the protocol provided by the manufacturer. The polymerase chain reaction (PCR) method without reverse transcriptase was used to test for genomic DNA contamination. The multiplex PCR amplification was performed as previously described (Mercati et al. 2019) using 1.0 µL of complementary DNA as a template for APLN and APLNR primers (Table1). Cycling conditions consisted of an initial denaturalizing cycle at 94C for 75s followed by 35 cycles for each target gene at 94C for 15s, 60C for 30s, 72C for 45s and a final extension step at 72C for 10min. For each experiment the complete set of samples was processed in parallel in a single PCR using aliquots of the same PCR master mix. The amplified PCR-generated products (18 µL of 25 µL total reaction volume) were analysed by electrophoresis on a 2% agarose gel using ethidium bromide staining (Troisi et al. 2020). The amplified products, collected from the agarose gel following electrophoresis, were purified with a Nucleospin Extract II kit and their identity was confirmed by DNA sequencing using the Sanger technique.
Immunohistochemistry and western blot reagents
For the immunohistochemical analysis (IHC), the rabbit polyclonal anti-APLN antibody (NBP2-31176) was purchased from Novus Biologicals (Novus Biologicals, USA); the mouse monoclonal anti-APLNR antibody (sc-517,300) used for IHC and Western Blot analysis was supplied by Santa Cruz Biotechnology (Santa Cruz, CA, USA); the normal goat serum (s-1000), the two secondary biotin-conjugated antibodies, the goat anti-mouse (BA-9200 and goat anti-rabbit (BA-1000) and the ABC Kit and DAB were purchased from Vector Laboratories (Vector Laboratories, Burlingame, CA, USA). Finally, the Eukitt (03989) was supplied by Sigma-Aldrich.
Immunohistochemistry
Following microwave oven antigen retrieval and using 10 mM citric acid, pH 6.0, (three 5-minute cycles at 750W), five-µm-thick serial sections were placed on poly-lysine-coated glass slides and processed for immunohistochemical reaction,. All subsequent steps were carried out in a moist chamber at room temperature to prevent evaporation of the reagents. After proper cooling the sections were pre-incubated for 30min with specific normal goat serum (1:10) in order to avoid non-specific binding of the primary antibodies. The serial sections were then incubated overnight with anti-APLN rabbit polyclonal (1:100) and anti-APLNR mouse monoclonal (1:100) primary antibodies. The next day, after washing in PBS, the sections were incubated with specific secondary biotin-conjugated antibodies (a goat anti-rabbit and a goat anti-mouse respectively (both 1:200) for 30min). Then, after another washing in PBS using the ABC KIT, the sections were incubated again for another 30min. Finally, the tissue sections were rinsed in PBS and the reaction was developed using diaminobenzidine (DAB) as the chromogen. At the end of the immunoreaction, the sections were rinsed in PBS, counterstained with haematoxylin, dehydrated and mounted in Eukitt. The sections in which the primary antibodies were omitted were used as negative controls of unspecific staining. A canine placenta was used as a positive control for the apelinergic system (Troisi et al. 2020). In order to further confirm the specificity of the anti-apelin receptor antibody, a western blot was performed using the same antibody on testicular tissue. All tissue analyses were carried out on coded slides using a light microscope (Nikon Eclipse E800) connected to a digital camera (Dxm 1200 Nikon digital camera). An image analysis system was used to process the images. The settings for image capture were standardized by subtracting the background signals obtained from the matched tissue sections which had not reacted with the primary antibodies and which were used as immunohistochemical controls (Dall’Aglio et al. 2012). However, these controls were not quantified given the predominantly qualitative nature of the immunohistochemical technique.
Western blot
The protein expression of APLNR was analysed by WB in all of the testicular samples. In short, total testicular proteins were extracted from the testicular tissue of each dog as previously described (Maranesi et al. 2021). The testicular tissues were homogenized in 300 ml of ice-cold RIPA buffer (PBS containing 1% Igepal CA-630, 0.5% sodium deoxycholate and 0.1% SDS) containing a protease inhibitor (Roche Complete™ Protease Inhibitor Cocktail) under agitation for 2 hat 4°C. Following incubation at 4°C for 20min, the homogenates were centrifuged at 12,000g for 60min at 4°C. The protein concentrations of the supernatants were measured using the bovine serum albumin (BSA) protein assay as reference standard. Equivalent amounts of protein (50mg) were separated by discontinuous 12% SDS-PAGE with 4% stacking gel for 40min at 200V and 500 mA. The proteins were then transferred onto nitrocellulose membrane using the Trans-Blot Turbo System (Bio-Rad). The membrane was then blocked by incubating it in Tris-buffered saline (TBS) containing 0.05% Tween 20 and 3% BSA. Immunoblotting was performed via overnight anti-APLNR monoclonal antibody (1:1000) incubation at 4°C. The membrane was then probed with HRP-labeled anti-mouse IgG antibody (1:10,000) for 60min at room temperature respectively under gentle agitation. All antibody incubations were performed in TBS containing 5% non-fat dried milk and 0.05% Tween-20 (Maranesi et al. 2019). The immunocomplexes were detected using an enhanced chemiluminescence method according to the manufacturer’s protocol (Clarity Western ECL Substrate, Bio-Rad). The blot images were scanned and acquired.
Results
RT-PCR
APLN and APLNR transcripts were detected in all samples. The APLN and APLNR PCR products were of the expected size (171 and 97bp, Fig.1A and B, respectively).

Gene expression of APLN (panel A) and APLNR mRNA (panel B) in the testicular tissue (T) of five dogs. Representative agarose gel electrophoresis stained with ethidium bromide was used to verify matches between expected and obtained PCR products. Lane CTR– represents a negative control of non-reverse-transcribed RNA submitted to PCR amplification, LD = 100bp DNA ladder
Immunohistochemistry
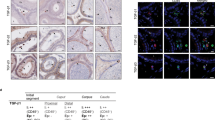
The immunohistochemical reaction showed the presence of apelin and its receptor in the canine testes. Apelin proved to be localized in the apical portion of the seminiferous tubules with a positive signal in the “residual bodies” (arrows, Fig.2a). As regards the apelin receptor, a positive immunoreaction was observed in the cytoplasm of the cells localized in the basal portion of the seminiferous tubules (arrow) and in the cytoplasm of Leydig cells (asterisk). (Fig. 2b)

Immunostaining for apelin (a) and apelin receptor (b) in the canine testes. Apelin (a) stained the residual bodies in the apical portion of the seminiferous portions (arrows). The apelin receptor (b) stained the basal line of spermatogenic epithelium (arrow) and Leydig cells in the connective tissue (asterisk). The image insets show the results of the negative controls performed to validate the IHC.
Western blot
The APLNR protein expression was evaluated with western blot in all of the samples. The immunoblot showed a strong band at approximately 35 Da (Fig.3).

Protein expression in the testicular tissue (T) of five dogs. MW = molecular weight marker
Discussion
In humans adipokines regulate numerous metabolic processes (Bertrand et al. 2015), including female and male reproductive systems, (Dupont et al. 2015) by controlling the activity of hypothalamic-pituitary gonadal axis under both normal and pathological conditions (Campos et al. 2008). In particular, in mammals, numerous other adipokines directly are able to influence male spermatogenic function (Milon et al. 2019; Kotula-Balak et al. 2022) reported that in landrace boar, immunocastration results in altered adipokine and leptin system. Moreover, some authors report that the change in adipokine expression and their receptor in testis are correlated with senescence and reproductive pathology (cryptorchidism and testis tumors) (Duliban et al. 2020) 0; Ramisz et al. 2021).
The role of apelin and its receptor in animal reproduction has been widely reported (Dupont et al. 2015; Estienne et al. 2019; Roche et al. 2017; Shokrollahi et al. 2021; Troisi et al. 2020). However, to the best of our knowledge, to date few studies have investigated the role that apelin and apelin receptors play in the male reproductive system (Bertrand et al. 2015; Dupont et al. 2015; Campos et al. 2008). Our results revealed, for the first time, the presence of apelin and its receptor in dog testes which are in agreement with those obtained for humans and mice (Medhurst et al. 2003). In particular, this study concerned the immunohistochemical localization of apelin in the residual bodies, in the apical portion of the seminiferous tubules, and of apelin receptor in the cytoplasm of cells localized in the basal portion of the seminiferous tubules. These observations suggest that apelin is involved in spermatogenesis of dog, as well as other adipokines reported in mammalian, via paracrine or endocrine manner mechanism (Thomas et al. 2013; Kurowska et al. 2018). Recently it was observed that the apelin signaling pathway was the most dominant in the seminal plasma of boar (Fraser et al. 2021). This observation could allow us to hypothesize that, if its presence in seminal plasma is confirmed also in the dog, apelin produced by the testes could represent one of the components of the seminal plasma with potential roles in key reproductive events (Martinez et al. 2020).
Despite the study conducted on rats (Brzoskwinia et al. 2020) where both apelin and its receptor were detected in the Leydig cells, in our study only APLNR was found in the cytoplasm of Leydig cells, allowing us to hypothesize the action of the molecule on the control of steroidogenesis with only an endocrine mechanism in dogs.
In conclusion, our results obtained using RT-PCR and immunohistochemical techniques to analyze the dog testis reveal, for the first time, the presence of the APLN-APLNR system in the dog testes under study as previously reported in the testes of adult rats (Brzoskwinia et al. 2020).
These results could provide valuable insights to study the possible role of the apelin/APJ system under normal and pathological conditions in dogs, which could represent an alternative animal model for comparative pathology in addition to the rats and mice commonly used in studies for comparative pathology.
Data Availability
not applicable.
Code Availability
not applicable.
Change history
17 February 2023
A Correction to this paper has been published: https://doi.org/10.1007/s11259-023-10085-2
References
Bertrand P, Valet P, Castan-Laurell I (2015) Apelin and energy metabolism. Front Physiol 6:115. doi:https://doi.org/10.3389/fphys.2015.00115
Brzoskwinia M, Pardyak L, Rak A, Kaminska A, Hejmej A, Marek S, Kotula-Balak M, Bilinska B (2020) Flutamide Alters the Expression of Chemerin, Apelin, and Vaspin and Their Respective Receptors in the Testes of Adult Rats. Int J Mol Sci 21:4439. doi:https://doi.org/10.3390/ijms21124439
Campos DB, Palin MF, Bordignon V, Murphy BD (2008) The ’beneficial’ adipokines in reproduction and fertility. Int J Obes (Lond) 32:223–231. doi:https://doi.org/10.1038/sj.ijo.0803719
Carpéné C, Dray C, Attané C, Valet P, Portillo MP, Churruca I, Milagro FI, Castan-Laurell I (2007) Expanding role for the Apelin/APJ System in Physiopathology. J Physiol Biochem 63:359–373
Dall’Aglio C, Polisca A, Boiti C, Ceccarelli P (2012) Immunolocalization of leptin and its receptor in the placenta of cats. Acta Histochem 114:719e22. doi:https://doi.org/10.1016/j.acthis.2011.12.008
Dall’Aglio C, Polisca A, Troisi A, Zelli R, Ceccarelli P (2014) Immunohistochemistry localization of orexin A and orexin type 2 receptor-positive cells in the placenta of dogs. Acta Histochem 116:989–992. doi:https://doi.org/10.1016/j.acthis.2014.03.004
Duliban M, Gorowska-Wojtowicz E, Tworzydlo W, Rak A, Brzoskwinia M, Krakowska I, Wolski JK, Kotula-Balak M, Płachno BJ, Bilinska B (2020) Interstitial Leydig Cell Tumorigenesis-Leptin and Adiponectin Signaling in Relation to Aromatase Expression in the Human Testis. Int J Mol Sci 21(10):3649. doi: https://doi.org/10.3390/ijms21103649
Dupont J, Pollet-Villard X, Reverchon M, Mellouk N, Levy R (2015) Adipokines in human reproduction. Horm Mol Biol Clin Investig 24:11–24. doi:https://doi.org/10.1515/hmbci-2015-0034
Estienne A, Bongrani A, Reverchon M, Ramé C, Ducluzeau PH, Froment P, Dupont J (2019) Involvement of Novel Adipokines, Chemerin, Visfatin, Resistin and Apelin in Reproductive Functions in Normal and Pathological Conditions in Humans and Animal Models. Int J Mol Sci 20:4431. doi:https://doi.org/10.3390/ijms20184431
Falcao-Pires I, Ladeiras-Lopes R, Leite-Moreira AF (2010) The Apelinergic System: A Promising Therapeutic Target. Expert Opin Ther Targets 14:633–645. doi:https://doi.org/10.1517/14728221003752743
Ferhat M, Katsuhiko F, Boudina S (2019) Autophagy in Adipose Tissue Physiology and Pathophysiology. Antioxid Redox Signal 31:487–501. doi:https://doi.org/10.1089/ars.2018.7626
Fraser L, Wasilewska-Sakowska K, Zasiadczyk Ł, Piątkowska E, Karpiesiuk K (2021) Fractionated Seminal Plasma of Boar Ejaculates Analyzed by LC-MS/MS: Its Effects on Post-Thaw Semen Quality. Genes (Basel) 12(10):1574. doi: https://doi.org/10.3390/genes12101574
Kawamata Y, Habata Y, Fukusumi S, Hosoya M, Fujii R, Hinuma S, Nishizawa N, Kitada C, Onda H, Nishimura O, Fujino M (2001) Molecular properties of apelin: Tissue distribution and receptor binding. Biochim Biophys Acta 1538:162–171. doi:https://doi.org/10.1016/s0167-4889(00)00143-9
Kotula-Balak M, Pawlicki P, Gałuszka A, Pardyak L, Tuz R, Dubniewicz K, Skrzypczak-Wiercioch A, Rak A, Dawid M, Tarasiuk K (2022) Effect of immunocastration using Improvac on the regulation of adiponectin and leptin in the testes of Landrace boars. Med Weter 77:1–6. doi: https://doi.org/10.21521/mw.6685
Kurowska P, Barbe A, Rózycka M, Chmielinska J, Dupont J, Rak A (2018) Apelin in reproductive physiology and pathology of different species: A Critical Review. Int J Endocrinol 9170480. doi: https://doi.org/10.1155/2018/9170480
Maranesi M, Petrucci L, Leonardi L, Bufalari A, Parillo F, Boiti C, Zerani M (2019) Kisspeptin/kisspeptin receptor system in pseudopregnant rabbit corpora lutea: presence and function. Sci Rep 9(1):5044. doi: https://doi.org/10.1038/s41598-019-41623-1
Maranesi M, Dall’Aglio C, Acuti G, Cappelli K, Trabalza Marinucci M, Galarini R, Suvieri C, Zerani M (2021) Effects of Dietary Polyphenols from Olive Mill Waste Waters on Inflammatory and Apoptotic Effectors in Rabbit Ovary. Anim (Basel) 11(6):1727. doi: https://doi.org/10.3390/ani11061727
Mercati F, Scocco P, Maranesi M, Acuti G, Petrucci L, Cocci P, Renzi A, De Felice E, Dall’Aglio C (2019) Apelin system detection in the reproductive apparatus of ewes grazing on semi-natural pasture. Theriogenology 139:156–166. doi: https://doi.org/10.1016/j.theriogenology.2019.08.012
Martinez CA, Cambra JM, Gil MA, Parrilla I, Alvarez-Rodriguez M, Rodriguez-Martinez H, Cuello C, Martinez EA (2020) Seminal plasma induces overexpression of genes associated with embryo development and implantation in Day-6 porcine blastocysts. Int J Mol Sci 21:3662. DOI: https://doi.org/10.3390/ijms21103662
Medhurst AD, Jennings CA, Robbins MJ, Davis RP, Ellis C, Winborn KY, Lawrie KWM, Hervieu G, Riley G, Bolaky JE, Herrity NC, Murdock P, Darker JG (2003) Pharmacological and immunohistochemical characterization of the APJ receptor and its endogenous ligand apelin. J Neurochem 84:1162–1172. doi:https://doi.org/10.1046/j.1471-4159.2003.01587.x
Mercati F, Maranesi M, Dall’Aglio C, Petrucci L, Pasquariello R, Tardella FM, De Felice E, Scocco P (2018) Apelin System in Mammary Gland of Sheep Reared in Semi-Natural Pastures of the Central Apennines. Anim (Basel) 8:223. doi: https://doi.org/10.3390/ani812022
Milon A, Pawlicki P, Rak A, Mlyczynska E, Płachno BJ, Tworzydlo W, Gorowska-Wojtowicz E, Bilinska B, Kotula-Balak M (2019) Telocytes are localized to testis of the bank vole (Myodes glareolus) and are affected by lighting conditions and G-coupled membrane estrogen receptor (GPER) signaling. Gen Comp Endocrinol 271:39–48. doi: https://doi.org/10.1016/j.ygcen.2018.10.021
Pope GR, Roberts EM, Lolait SJ, O’Carroll AM (2012) Central and peripheral apelin receptor distribution in the mouse: species differences with rat. Peptides 33:139–148. doi:https://doi.org/10.1016/j.peptides.2011.12.005
Ramisz G, Turek W, Chmurska-Gasowska M, Rak A, Pietsch-Fulbiszewska A, Galuszka A, Kotula-Balak M, Tarasiuk K (2021) Senescence and adiponectin signaling - Studies in canine testis. Ann Anat 234:151606. doi: https://doi.org/10.1016/j.aanat.2020.151606
Roche J, Ramé C, Reverchon M, Mellouk N, Rak A, Froment P, Dupont J (2017) Apelin (APLN) regulates progesterone secretion and oocyte maturation in bovine ovarian cells. Reproduction 153:589–603. doi:https://doi.org/10.1530/REP-16-0677
Sandal S, Tekin S, Seker FB, Beytur A, Vardi N, Colak C, Tapan T, Yildiz S, Yilmaz B (2015) The effects of intracerebroventricular infusion of apelin-13 on reproductive function in male rats. Neurosci Lett 602:133–138. doi: https://doi.org/10.1016/j.neulet.2015.06.059
Shokrollahi B, Shang J, Saadati N, Ahmad HI, Yang C (2021) Reproductive roles of novel adipokines apelin, visfatin, and irisin in farm animals Theriogenology. 172:178–186. https://doi.org/10.1016/j.theriogenology.2021.06.011
Shokrollahi B, Zheng H, Li L, Tang L, Ma X, Lu X, Duan A, Zhang Y, Tan X, Huang C, Xu Y, Shang J (2022) Apelin and Apelin Receptor in Follicular Granulosa Cells of Buffalo Ovaries: Expression and Regulation of Steroidogenesis. Front Endocrinol 13:844360. doi:https://doi.org/10.3389/fendo.2022.844360
Tatemoto K, Takayama K, Zou MX, Kumaki I, Zhang W, Kumano K, Fujimiya M (2001) The novel peptide apelin lowers blood pressure via a nitricoxide dependent mechanism. Regul Pept 99:87–92. doi: https://doi.org/10.1016/s0167-0115(01)00236-1
Thomas S, Kratzsch D, Schaab M, Scholz M, Grunewald S, Thiery J, Paasch U, Kratzsch J (2013) Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil Steril 99:1256–1263e3. doi: https://doi.org/10.1016/j.fertnstert.2012.12.022
Troisi A, Dall’Aglio C, Maranesi M, Orlandi R, Speranza R, Zerani M, Polisca A (2020) Presence and expression of apelin and apelin receptor in bitch placenta. Theriogenology 147:192–196. doi:https://doi.org/10.1016/j.theriogenology.2019.11.016
Vázquez-Vela MEF, Torres N, Tovar AR (2008) White adipose tissue as endocrine organ and its role in obesity. Arch Med Res 39:715–728. doi:https://doi.org/10.1016/j.arcmed.2008.09.005
Acknowledgements
The authors are grateful to Dr Paola Coliolo for her excellent technical support. The authors also acknowledge the English language editing of the text carried out by Mrs Eleanor Fabri and Mr James Alexander Stevens.
Funding
The authors did not receive any financial support for this study.
Open access funding provided by Università degli Studi di Perugia within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
Polisca, Orlandi, Pastore, Martínez-Barbitta and Troisi examined the animals and collected the samples;
Dall’Aglio carried out the immunohistochemical studies;
Maranesi and Suvieri performed the RT-PCR and WB studies;
Troisi, Dall’Aglio, Maranesi and Polisca prepared the manuscript.
Bazzano analyzed the results.
Polisca designed and supervised study.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval:
The authors declare that Ethics Committee Approval was not required because all the dogs enrolled in this study underwent orchiectomy at the Veterinary Hospital of the Department of Veterinary Medicine of Perugia with the written consent of their owners and did not undergo any other experimental procedures or manipulation.
Consent to participate:
not applicable.
Consent for publication:
not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: This article was originally published with an omitted author (Marcelo Martínez-Barbitta).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Troisi, A., Dall’Aglio, C., Maranesi, M. et al. Presence and localization of apelin and its cognate receptor in canine testes using immunohistochemical and RT-PCR techniques. Vet Res Commun 47, 929–935 (2023). https://doi.org/10.1007/s11259-022-10001-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11259-022-10001-0