
Abstract
In fresh waters, the origins of dissolved organic matter (DOM) have been found to exert a fundamental control on its reactivity, and ultimately, its ecosystem functional role. A detailed understanding of landscape scale factors that control the export of DOM to aquatic ecosystems is, therefore, pivotal if the effects of DOM flux to fresh waters are to be fully understood. In this study we present data from a national sampling campaign across the United Kingdom in which we explore the variability in DOM composition in three broad landscape types defined by similar precipitation, geology, land use and management, hydrology, and nutrient enrichment status. We characterised samples from fifty-one sites, grouping them into one of three major underlying classifications: circumneutral streams underlain by clay and mudstone (referred to as ‘clay’), alkaline streams underlain by Cretaceous Chalk or by Carboniferous or Jurassic Limestone (‘limestone’), and acidic streams in peatland catchments underlain by a range of low permeability lithologies (‘peat’). DOM composition was assessed through organic matter stoichiometry (organic carbon: organic nitrogen; organic carbon: organic phosphorus; C/N(P)DOM) and metrics derived from ultra-violet (UV)/visible spectroscopic analysis of DOM such as specific UV absorption (a254 nm; SUVA254). We found similar SUVA254, C/NDOM and DOM/a254 relationships within classifications, demonstrating that despite a large degree of heterogeneity within environments, catchments with shared environmental character and anthropogenic disturbance export DOM with a similar composition and character. Improving our understanding of DOM characterisation is important to help predict shifts in stream ecosystem function, and ecological responses to enrichment or mitigation efforts and how these may result in species composition shifts and biodiversity loss in freshwater ecosystems.
Similar content being viewed by others

Introduction
Landscape derived dissolved organic matter (DOM) has been found to exert a fundamental control on the productivity and ecological status of freshwater ecosystems (Jaffe et al. 2008; McDowell 2003). While in-stream production through carbon fixation and subsequent exudation by stream biota can be an important autochthonous source of DOM, the majority of studies investigating DOM sources at catchment scale have found allochthonous DOM to dominate fluxes (McGlynn and McDonnell 2003; Toming et al. 2013). The degree of nutrient enrichment (Mackay et al. 2020; Mueller et al. 2016), rates of photochemical processing (Cory et al. 2015; Judd et al. 2007; Spencer et al. 2009) and microbial kinetics (Brailsford et al. 2019a, b) all mediate DOM metabolism in streams and stream sediments, and its ultimate availability as a nutrient and energy resource for stream biota, but the key control on its composition is primarily defined by its origins.
Catchment characteristics, including land cover (Williams et al. 2016; Wilson and Xenopoulos 2009), hydrology (Fasching et al. 2016; Heppell et al. 2017), and soil properties (Yates et al. 2019a) have all been observed to play an important role in determining the nature of DOM delivered to fresh waters from their catchments. Others have shown that the structural complexity of DOM decreases as the ratio of continuous cropland to wetland in the catchment increases (Wilson and Xenopoulos 2009), with the dominance of agricultural land use shifting DOM composition towards a more microbial, less plant-derived composition (Graeber et al. 2012, 2015), as determined by DOM fluorescence spectroscopy. An analysis of samples from across a European geoclimatic gradient conducted by Mattsson et al. (2009) found samples from catchments with a greater proportion of agricultural land to have lower dissolved organic carbon to dissolved organic nitrogen ratios (DOC:DON), close to the Redfield OC:ON ratio (molar) of 6.6:1, indicative of microbially sourced DOM. Similarly, a recent global analysis of dissolved organic nitrogen (DON) concentrations relative to total dissolved N concentrations in over 2000 streams from 3.66° S to 69.7° N across arctic, boreal, temperate and tropical biomes identified critical thresholds in DOM composition, with increases in nutrient enrichment linked to lower DOC:DON ratios (Wymore et al. 2021).
Much less is known about dissolved organic P (DOP) in fresh waters, but a recent paper by George et al. (2017) presents a valuable synthesis of knowledge on organic P in terrestrial systems. They conclude that concentrations are strongly linked to soil character and to land use and management, with elevated concentrations observed in agricultural soils, particularly those used for livestock production. In our own work we also see DOP concentrations in streams linked to landscape character and elevated in agricultural streams versus those draining through natural or semi-natural catchments (Yates et al. 2019a; Lloyd et al. 2019; Brailsford et al. 2021), and both microbial and plant uptake of both DOP and DON and as a nutrient resource (Brailsford et al. 2019a, b; Mackay et al. 2020). Such elevated nutrient concentrations are commonly observed in UK agricultural catchments (Yates et al. 2019a). Contradictory observations from Graeber et al. (2012) and others (Sachse et al. 2005) suggest catchments with a high proportion of agricultural activity export structurally complex DOM, characterised by elevated DOC concentrations and a dominance of humic-like fluorescent DOM, though other studies (Lloyd et al. 2021; Mena-Rivera et al. 2021) indicate DOM in agricultural catchments has a high proportion of lower molecular weight compounds resulting from microbial processing of terrestrially derived DOM. Similarly, both Yates et al. (2019a) and Williamson et al. (2021) in a pan-UK study concluded that higher DOC, higher Specific UV Absorbance (SUVA) and higher DOC:DOC ratios were consistently observed in peaty catchments than in lowland intensively farmed agricultural catchments. This inconsistency in findings from different studies suggests the link between land cover and DOM composition may be catchment-specific, reflective of variations in landscape scale processes such as hydrologic connectivity, the nature of nutrient sources in the catchment, its underlying geology, and soil type, but that commonalities in these catchment characteristics might then yield generalisable DOM characteristics for other catchments with similar properties.
Similar, but less pronounced variability in stream DOM composition has been observed in peatland environments with soil-pore water interaction time a major driver of compositional variability between base- and storm-flow conditions (Austnes et al. 2010; Nguyen et al. 2010; Wagner et al. 2019). These variations may be explained, in part, by a combination of differing land management practices that impact upon flow routing through the landscape and residence times within soils (Yates et al. 2016). Irrigation (Hernes et al. 2008), riparian land management (Pisani et al. 2020), alterations to aquatic microbial processing, and fertiliser use and livestock grazing which generates variations in the soil organic matter pool (Heinz et al. 2015; Yates et al. 2019a), have all been shown to impact DOM concentration and composition in fresh waters. Catchment hydrology has also been found to exert control on DOM composition with storm events mobilising pools of organic material not readily accessed under baseflow conditions, including the flushing of organic rich soil horizons (Austnes et al. 2010; Inamdar et al. 2011; Nguyen et al. 2010; Wagner et al. 2019). In contrast, organic material with a low abundance of higher molecular weight compounds such as in lignin-derived phenols, with optical signatures matching that of microbially degraded DOM, have been reported widely in permeable catchments with complex sub-surface drainage networks (Benk et al. 2019; McDonough et al. 2020), most likely showing a response to diagenetic processing in groundwater aquifers with long residence times.
In order to fully understand the impact of catchment characteristics on the delivery of DOM to freshwater ecosystems, and derive an understanding of the generic properties of DOM draining from similar landscape types, a comprehensive knowledge of the interactions between land cover, land management, and catchment hydrology is required. However, this understanding cannot be based on measurements of DOC concentration/composition alone. While studies investigating the impact of land use on organic matter composition are increasing, the majority of studies investigating the source and transport of DOM focus mainly on DOC concentration, and more recently, composition. Global estimates suggest riverine fluxes of DOC to oceans are substantial, accounting for ~ 0.25 Pg C year−1 (Battin et al. 2009; Drake et al. 2019). While no such global estimates exist for DON and DOP, their concentrations in global rivers have found to be equal, if not greater than their inorganic counterparts (Perakis and Hedin 2002; Stanley and Maxted 2008; Yates et al. 2019a; Wymore et al. 2021) particularly in natural and semi-natural ecosystems, and as such should routinely be included in studies of aquatic DOM.
In this paper we explore the variability in stream DOM composition in relation to catchment character, through a detailed analysis of samples collected from a range of UK catchments with marked variations in climate, geology, soils, land use and management, hydrology, and degree of nutrient enrichment. Similarities in DOM composition relative to nutrient stoichiometry are investigated to reveal common characteristics of DOM linked to environmental character, and the impact of hydrology, geology and land use on rates and composition of DOC, DON, and DOP are explored.
Materials and methods
Catchment characteristics
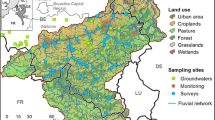
A total of 51 catchments were sampled across the UK (Fig. 1) with catchment area ranging between 0.6 and 416 km2. Sites were selected with three major underlying lithologies and soil types: 18 circumneutral streams underlain by clay and mudstone (hereafter referred to as ‘clay’), 18 alkaline streams dominated by calcium carbonate (CaCO3), underlain by Cretaceous Chalk or by Carboniferous or Jurassic Limestone (hereafter referred to as ‘limestone’), and 15 acidic streams in peatland catchments underlain by a range of low permeability lithologies (hereafter referred to as ‘peat’). Land cover classification data were supplied by the UK Centre for Ecology and Hydrology and were manipulated using geographical information system software (ArcGIS; ESRI 2018. Version 10 Redlands, CA). All sites had previously been the focus of Natural Environment Research Council or UK Government (Defra) funded research on stream DOM, C, N and/or P flux, with long-term or archived high resolution observational data available in open access data archives, providing contextual data to support our spatial sampling programme. Details on the key published literature and open access data archives for each site are provided in Table S1, Supplementary Information. The sites sampled varied considerably in their land cover and soils classification, with the proportion of agricultural land ranging between 0 and 98%, and peatland ranging between 0 and 100% of catchment area in the selected sites. The majority of peatland catchments were located in the UK’s upland areas and were located on blanket bog that has been modified to varying extents by sheep grazing, drainage, grouse moor burning, wildfire and air pollution. By contrast, the intensively farmed agricultural catchments were located in lowland areas, with oilseed rape, barley, and wheat dominating crop distributions in our predominantly arable-dominated chalk catchments, and improved grassland supporting intensive dairy production in our clay catchments. Woodland covered an average of 6.63% of land, ranging between 0.0 and 35% while urban areas only accounted for an average of 1.32%, ranging between 0.0 and 10%, reflecting isolated farmsteads and villages in these largely rural catchments.

Map of a a map of the geoclimatic regions established for the UK by Greene et al. (2015), reproduced with permission, and b the fifty-one sampling locations included in this study, colour coded to indicate their major site classification
Baseflow indices (BFI; range 0–1), which reflect the proportion of stream flow delivered along baseflow (throughflow plus groundwater flow) pathways, ranged between 0.13 and 0.98, encapsulating geologies such as Silurian sedimentary deposits of the central lowlands of Scotland to the Cretaceous Chalk prevalent across much of south-eastern England (BFI from the closest UK Hydrology of Soil Types (HOST) classification). Site average annual rainfall values (1961–1990) were taken from the nearest known gauging station extracted from the UK National River Flow Archive (NRFA: https://nrfa.ceh.ac.uk) or from the published literature for each site. All sites were subsequently separated into three categories for analysis, best describing a combination of their (a) hydrology and geoclimatic features (BFI, rainfall, runoff), (b) land cover classification and (c) dominant soil type and agricultural production system. Site classifications along with mean group attributes are provided in Table 1, below, with a site-specific breakdown and supporting publications presented in Supplementary Information, Table S1.
Sample collection
All samples were collected between June and September 2017. Additional data were collated from published literature (see Supplementary Information) to enable a comparison between spot samples collected in this study and long-term data, which will consider seasonal patterns. All samples were collected between June and September 2017. Samples were collected using acid-washed (10% HCl, v/v) high-density polyethylene sample bottles, refrigerated at 4 °C in the dark and couriered overnight for filtration and analysis at the University of Bristol. All samples were filtered through pre-washed 0.45 µm cellulose-nitrate membrane filters (Whatman GF/C) refrigerated at 4 °C in the dark and analysed within 24 h of being returned to the laboratory.
Analytical methods
Dissolved organic carbon
DOC concentrations were analysed using a Shimadzu TOC-L series analyser (Shimadzu Corp.), determined as non-purgeable organic carbon by high temperature catalytic oxidation, following sample acidification (1% HCl). The mean of three to five replicate injections is presented here, where the coefficient of variance was < 2%. Instrument accuracy was calculated as − 0.9% based on analysis of replicate standards (n = 5). The instrument limit of detection (LoD) following blank replication (n = 60) was calculated as 115 µg C l−1.
Nitrogen species and phosphorus fractions
Total oxidised nitrogen (TON, hereafter referred to as NO3-N, where NO2-N was consistently < 0.1% of TON), soluble reactive phosphorus (SRP; measured as PO4-P), and total ammonium (NH3-N + NH4-N; hereafter referred to as NH4-N) were determined using a Skalar San++ multichannel continuous flow autoanalyser (Skalar Analytical B.V.). Total dissolved nitrogen (TDN) and phosphorus (TDP) were determined following digestion of filtered samples with potassium persulphate (K2S2O8), using the protocol modified from Johnes and Heathwaite (1992), as reported in Yates et al. (2016). Total nitrogen (TN) and phosphorus (TP) were determined following digestion of an unfiltered aliquot. DON and DOP, and particulate organic nitrogen and phosphorus (PON, PP) were determined by difference where:
For detailed method description see Yates et al. (2019a). Mean precision for NO3-N, PO4-P, and NH4-N, based on analysis of replicate standards (n per run = 5) was found to be ± 1.0%, ± 0.8%, and ± 1.0% with accuracy of + 5.0%, + 2.2%, and − 1.4% respectively. Instrument LoD was calculated for each method as 27 µg l−1, 4 µg l−1, and 8 µg l−1, respectively, following blank replication (n = 10).
DOM composition
Absorbance spectra were scanned using a Varian Cary 60 UV/Vis spectrometer (Agilent Technologies) on each sample between 200 and 800 nm at 1 nm intervals, with samples analysed at laboratory ambient temperature (~ 20 °C). Dissolved organic matter composition was examined through calculation of the specific ultraviolet absorbance (SUVA254; a proxy for DOM aromatic content as discussed by Weishaar et al. (2003)) and was calculated by dividing the decadic absorbance at 254 nm by DOC concentration (mg l−1). All absorbance data presented here are as absorption coefficients calculated as given in Eq. 5.
where A(λ) is the measured absorbance and l is the pathlength of the cuvette used for analysis in metres. DOM composition was also inferred using organic matter stoichiometry. All elemental stoichiometry (DOC:DON and DOC:DOP), hereafter referred to as C/NDOM and C/PDOM, are presented as molar ratios. To check the wavelength scale of the instrument was within the manufacturer’s tolerance a sealed holmium perchlorate (holmium oxide 4% m/v in 10% v/v perchloric acid; Starna Scientific) reference was analysed daily, prior to sample analysis.
Watershed delineation and statistical analysis
Watershed delineation and catchment size were determined using the spatial analysis and hydrology toolboxes in ArcGIS, using digital elevation models sourced from EDINA Digimap [OS Terrain 5 DTM (Shape geospatial data), scale 1:10 000, Ordnance Survey, using the EDINA Digimap Ordnance Survey Service]. Land cover data were sourced from the UK Centre for Ecology and Hydrology (Land Cover Map 2007 Great Britain, Centre for Ecology & Hydrology, using the EDINA Environment Digimap Service). The land cover data were further classified with the following categories: arable, improved grassland, woodland, fresh water, peatland and urban.
All statistical analyses were conducted using SPSS (IBM SPSS Statistics for Windows, version 25.0; IBM Corp., Armonk, NY) with plots generated using SigmaPlot (version 14.0; Systat Software, San Jose, CA). In order to visualise statistical differences between land classification variables, principal components analysis (PCA) was conducted with each measurement variable z-score normalised prior to analysis. All data are presented ± 1 standard deviation. Correlation coefficients and correlation matrices were produced using R Studio. Kruskal–Wallis H tests were used to determine statistical significance in land classification data with subsequent pairwise comparisons adjusted by the Bonferroni correction for multiple tests.
Results
Multiple pairwise comparisons using the Kruskal–Wallis H test demonstrate significant differences between the relative distribution of land classifications across geoclimatic groups (Fig. 2a). The proportion of improved grasslands differ significantly between study catchments (χ2 (2) = 32.98, p ≤ 0.001) with limestone and clay differing significantly from peat (χ2 (2) = 25.3, p ≤ 0.001 and χ2 (2) = 31.6, p ≤ 0.001) and a higher proportion of grassland in the clay catchments which support intensive dairy production. There are stark differences in the dominant farming type in these lowland, intensively farmed catchments (Fig. 2b). Catchments within the limestone sub-group are dominated by arable farming, with a higher proportion of land cultivated for oilseed rape (χ2 (2) = 13.1, p ≤ 0.001), field beans (χ2 (2) = 6.49, p ≤ 0.05), barley (χ2 (2) = 5.82, p ≤ 0.05), and wheat (χ2 (2) = 4.95, p ≤ 0.05). By contrast clay catchments have a higher proportion of grassland on heavy clay soils, used to support intensive dairy cattle production. Improved grassland in the peatland catchments, by contrast, is used to support sheep production, but is a minor proportion of land cover in these blanket bog-dominated peatland systems.

Principal components analysis bi-plot with vector loadings for a land use classification (principal components 1 and 2) including all data grouped by site classification, and b crop type (principal components 1 and 2) for limestone and clay classifications (absent in peatland catchments)
Across all sites, mean NO3-N (3.65 ± 3.37 mg N l−1), PO4-P (56.2 ± 91.6 µg P l−1) and NH4-N (56.2 ± 126 µg N l−1) concentrations demonstrated the greatest variability due to the geospatial gradient farming intensity. Sites underlain by peat soils exhibited the lowest range of nitrate concentrations (Table 2; 0.335 ± 0.926 mg N l−1) followed by clay (3.70 ± 2.09 mg N l−1), with the highest concentrations observed in catchments underlain by predominantly chalk geology (6.99 ± 2.62 mg N l−1). A similar pattern is observed for TN concentrations, and while PON comprises a small fraction of TN, DON concentrations were found to be elevated in limestone (0.98 ± 0.34 mg N l−1) and clay (0.89 ± 0.36 mg N l−1) catchments compared to peat (0.68 ± 0.33 mg N l−1) systems. DON decreases as a percentage of TN with increasing TN concentration, accounting for 37% of TN across all sites, representing the dominant component of TN in peat catchments (limestones = 16%, clay = 19%, and peat = 78%). PO4-P concentrations showed a similar pattern and were lowest in the peat catchments (14.3 ± 42.2 µg P l−1), and highest in clay catchments (97.3 ± 92.3 µg P l−1) when compared to the limestone (52.4 ± 104 µg P l−1) (Fig. 3a, b). Unlike with N species, PP comprises a large proportion of TP at all sites, making the relationship between DOP as a percentage of TP less clear in relation to the nutrient gradients in this study.

Scatter plot of a DON as a percentage of TDN and b DOP as a percentage of TDP. Data grouped by geoclimatic classification. Transparent circles represent raw data, solid filled circles represent mean value ± 1 standard deviation
DOM stoichiometry (DOC/DON/DOP) is variable between site typologies. Peat catchments with low inorganic N and P concentrations, predominantly underlain by impermeable geologies demonstrate C-rich, N-poor stoichiometry with high SUVA254 values (mean C/NDOM ratio = 18.5 ± 11.3; SUVA254 = 6.0 ± 0.9 mg C l−1 m−1). This range in both C/NDOM and SUVA254 is commonly associated with forms of organic matter derived from the degradation of terrestrial material such as woody debris and leaf litter, supporting the idea that soil C:N acts as a dominant control on DOM composition in these landscapes (Yates et al. 2019a). Chalk sites, those most enriched in N, demonstrate lower C/NDOM ratios, with sites being C depleted/N enriched (mean C/NDOM ratio = 2.2 ± 1.6; SUVA254 = 2.9 ± 0.7 mg C l−1 m−1), and display values similar to those observed from autochthonous DOM production, and in treated human sewage and animal manures (Yates et al. 2019b). While this difference between categories may be driven by the variability in DOC concentrations across environments, DON concentrations also correlated significantly with the predominance of agricultural land (p < 0.001; Fig. 4a).

a Spearman’s rank correlation matrix including catchment descriptors, land cover and chemical variables. Values are significant at p-values of 0.001***, 0.01**, and 0.05*, colour bar represents correlation coefficient. b Example correlation between SUVA254 and BFI, with mean centroids ± 1 standard deviation grouped by site classification
Land cover classification across geoclimatic groups correlated well with chemical variables. Increases in percent agricultural land in each catchment were associated with increases in nitrate, and TN concentrations (correlation coefficient = 0.76, 0.71, and 0.72 respectively; p < 0.001), while correlating negatively with both DOC concentrations and SUVA254 (− 0.41; p < 0.01 and − 0.65; p < 0.001). Significant correlations were also observed between the proportion of flow derived from groundwater flow or deep throughflow pathways (as reflected in the BFI for each site typology) which correlated negatively with DOC, C/NDOM, and SUVA254 (− 0.70, − 0.78, and − 0.65; p < 0.001), while demonstrating a significant positive correlation with nitrate (0.74; p < 0.001). A heat map of correlation coefficient data is presented in Fig. 4.
In this study, absorption coefficients (λ = 254 nm) correlated well with DOC concentrations within all site types (Fig. 5a), with the strongest association observed in peatland sites (r2 = 0.96, p < 0.001), followed by clay (r2 = 0.95, p < 0.001), then limestone catchments (r2 = 0.89, p < 0.001). Sites with the strongest relationship between DOC and a254 also demonstrate the greatest increase in absorbance per unit DOC, along with the highest SUVA254 values (6.0 ± 0.9 mg C l−1 m−1), consistent with DOM of an allochthonous origin. By contrast, limestone sites, showed a shallower regression slope, and the lowest SUVA254 values (2.9 ± 0.7 mg C l−1 m−1).

Relationship between a DOC and a254. Inset (i) shows the low DOC concentration range for limestone sites only. b Presents the results of a linear regression analysis between DOC and DON across all sites sampled. Data are grouped by site classification. There was no statistically significant relationship generated for limestone catchments
Regression analysis conducted between DOC and DON concentration data, with sites split by classification, is presented in Fig. 5b. Regression equations are assumed to be representative of the mean change in DON per unit DOC across each site classification (Table 3). Peat sites demonstrate the strongest relationship between DOC and DON (r2 = 0.44, p < 0.05), spanning the largest range of DOC concentrations, and the narrowest range of DON concentrations (DOC range = 1.6–32.4 mg C l−1, DON range = 0.283–1.46 mg N l−1).
Discussion
It is evident from the data presented in this study that fundamental differences in DOM character exist between the environments covered by this analysis, driven by differences in catchment properties, including nutrient enrichment through land management practises. Commonalities in SUVA254, C/NDOM and DOM/a254 relationships exist within group classifications, such that similar properties emerge for all peatland catchments, for all clay catchments, and for all limestone catchments. However, local variations in land use and management also influence, and account for, variations in DOM concentration and composition within each group. In the lowland sites included in this study, the high NO3-N concentrations found in both limestone and clay classifications result in DON making up a smaller proportion of TDN than in less intensively farmed peat catchments. The dataset also reveals N enrichment in the stream DOM pool in lowland, intensively farmed catchments, relative to the upland peatland sites. Meanwhile there are marked differences in the proportion of the TDN pool contributed by DON, and the DOM pool composition between the two classes of intensively farmed lowland catchments studied here. Clay catchments which support intensive dairy cattle production have up to 40% of the TDN pool in the form of DON and a higher DOC:DON ratio compared to an average of 20% of TDN and lowest DOC:DON ratio in streams draining through the limestone catchments which are predominantly used for intensive arable farming with low intensity sheep grazing (Fig. 5, Table 3). Both have a lower proportion of TDN contributed by DON and a lower DOC:DON ratio than in our peatland catchments. Thus, while there are strong commonalities between catchments with similar lithology, hydrology and land use and management, there are nevertheless marked differences in the composition of the stream nutrient and DOM pools between agricultural catchments with differing farming systems and population densities evident in our datasets. An approach which groups all ‘agricultural’ catchments together as one category would be likely to misrepresent both the quantitative significance of DOM in the total N and P load to which aquatic organisms are exposed and the specific molecular composition of the DOM pool and its likely impacts on the freshwater ecosystem.
Our findings support the recent analysis of a global dataset of > 77,000 observations across > 2000 sites presented by Wymore et al. (2021) across catchments ranging from arctic, to boreal, temperate, and tropical biomes. Their analysis of a global N speciation dataset identified two critical TDN concentration thresholds at which the composition of the N pool, in particular the proportion of TDN present as DON shifted significantly, at 1.3 and 2.8 mg N l−1. On a global scale, increases in DON and NH4-N were outpaced by increases in NO3-N concentrations, correlating with increases in anthropogenic disturbance in these catchments. The high degree of variability in the relationship between DON and TDN at lower TN concentrations (< 1.3 mg N l−1) observed in that global analysis is echoed in the data presented in this study, suggesting no single landscape control on the composition of the N pool in streams. Rather, local landscape characteristics such as land management, plant species composition, topography, soil characteristics, and wetland cover in natural or semi-natural catchments, as well as relative rates of N deposition (Helliwell et al. 2007) or recovery from acidification (Rodriguez-Cardona et al. 2021) exert catchment-specific controls on N speciation, shifting C:N ratios in the riverine DOM pool and altering its likely ecosystem functional role. Helliwell et al. (2007), for example, report upward shifts in nitrate leading relative to DOC flux to streams moving from low to high N deposition areas, while Rodriguez-Cardona et al. (2021) report changes in DOC:DON stoichiometry of the stream DOM pool as systems recover from atmospheric acid deposition.
Wymore et al. (2021) also report instances where systems approaching a near natural state may be in fact dominated by oxidised N species rather than elevated DON contributions. Our study only included peatland catchments with low intensity sheep grazing, with the data showing a similar pattern of N speciation to that observed by Durand et al. (2011) in their data set for 87 European research catchments and similar N enrichment gradient, and by Helliwell et al. (2007) in their earlier work on N speciation in streams draining from upland catchments in the UK. However, if upland, low intensity grazing sites on mineral soils had been included, it is likely those sites would have shown patterns similar to some of those reported by Wymore et al. (2021). Their work also included a selection of tropical catchments, which were the source of these observations of high nitrate proportion at low TDN concentrations, but otherwise the global dataset was dominated by catchments in temperate and boreal biomes, with a high proportion of peatland sites, more similar in geoclimatic character to the spectrum of upland to lowland sites included in this UK study. As such, our current understanding of the relationship between DON and NO3-N may also be skewed by an inherent sampling bias towards temperate and boreal peatland systems, largely under-representing tropical regions. The patterns and conclusions drawn here may therefore only be robustly applicable in catchments in other temperate and boreal regions.
Comparable data for P fractionation collected during this study suggests a less well-defined relationship than exists, in this study, for DON and NO3-N. While previous work has found % DOP and TDP to have a similar relationship to % DON and TDN concentrations (e.g. Yates et al. 2019a), this study, spanning a greater range of TDP concentrations and catchment typologies, does not. Indeed, a wide range of ratios between SRP and TP are reported in the literature, reflecting the particular balance and composition of point and diffuse P sources in each catchment (Jarvie et al. 1998; 2005; Johnes 2007; Jordan et al. 2007; Thompson and Cotner 2018; Stackpoole et al. 2019), few of these earlier studies report the DOP fraction concentrations. The lack of a strong relationship between % DOP and TDP in this study may reflect the larger range of P source areas captured in this sampling programme than in our earlier two-catchment comparison. Some rural sites in this study received diffuse seepage of P-rich effluent from septic tanks, while in more populated catchments, they additionally received point source discharges from sewage treatment works (STW) which have markedly different nutrient speciation and DOM composition signatures (Yates et al. 2019b). The data presented here suggest some systems, with elevated TDP concentrations may in fact demonstrate near complete dominance of DOP over PO4-P in the TP pool, while others, particularly in intensively farmed arable catchments have PP as the dominant fraction, as has been reported in our previous work (Johnes 2007; Lloyd et al. 2019). Where DOP is the dominant P fraction, this occurs mainly in upland peat dominated sites and in some intensive cattle production sites underlain by clay, and may reflect both soil-derived DOP pools, as well as DOP-rich livestock manures and slurries voided to the land by sheep in the peatland catchments, or by cattle in the clay catchments.
The upland peatland catchments, with minimal agricultural activity are the least nutrient enriched sites included in this study. These have high in-stream C/NDOM (molar) ratios (~ 18) with similar C/NDOM ratios to those observed across a wide range of studies which have previously reported these ratios in peatland catchments (Helliwell et al. 2007; Austnes et al. 2010; Yates et al. 2016, 2019a). High variability in both N and P speciation over a narrow TDN/TDP range at these sites suggests local scale factors contributing to the variable dominance of inorganic vs. organic nutrient fractions in the TN and TP pools. While there is a high degree of variability in % DON and % DOP values within these environments, DOM compositional metrics demonstrate a consistent behaviour with a strong relationship between DOC and a254 observed, spanning a wide range of DOC concentrations. DOC dominates over DON and DOP export at these sites and there is also a moderate, but significant relationship between DOC and DON. These are C-rich and relatively N- and P-poor systems. As suggested by Brookshire et al. (2007) the relationships observed between DOC and DON in this study may indicate that DON losses are largely controlled by soil organic matter dissolution and transport with DON bound up in higher molecular weight compounds. The high SUVA254 values observed for these sites in this study support this, suggesting material is highly aromatic (Weishaar et al. 2003) with high C/NDOM ratios.
Consistently low C/NDOM ratios and SUVA254 values were observed in all limestone catchments in this study, characterised as systems with a high proportion of intensively farmed arable land with some sheep production on steeper slopes, in permeable landscapes where the dominant flow pathway (BFI = 0.92 ± 0.05; Table 1) is via throughflow and groundwater flow pathways. Limestone catchments also have the highest stream NO3-N concentrations in this study (6.99 ± 2.62 mg N l−1). This may be ascribed to high rates of nitrate leaching through calcareous brown earth soils, and legacy nitrate accumulation in the groundwater aquifer (Wang et al. 2016) mainly due to a combination of groundwater enrichment by historic agricultural practices (Wang et al. 2016; Ascott et al. 2017) and inputs from wastewater treatment facilities and septic tanks which are commonplace across the rural British landscape (Withers et al. 2012; Yates et al. 2019b). Although uncertainties in DON calculations increase with lower DON:TDN ratios, increased N loading in agricultural catchments has been found to increase DON loading in receiving waterbodies (Pellerin et al. 2006; van Kessel et al. 2009; Durand et al. 2011; Graeber et al. 2015). C/NDOM ratios for limestone catchments were approximately 8 times lower than those observed in the peat catchments and approximately half those of clay catchments, reflecting the higher soil organic matter content in both clay and peat dominated catchments.
Fertilisation of agricultural soils increases both soil organic carbon content and microbial biomass. A global synthesis of the effects of N fertilisation on microbial communities by Geisseler and Scow (2014), comprising 106 data sets from 64 studies, found N fertilisation to increase soil microbial biomass across all studies, with only 17% of data sets showing a lower organic carbon content in fertilised fields. It should be noted that in each case, P would be added in addition to N, and that it is not possible, therefore, to attribute the increase in microbial biomass to N alone. These results contrast with other studies (Jian et al. 2016) which have found that N fertilisation reduces soil microbial biomass, attributed to a resulting shift in species composition, or a reduction in soil pH, which is offset in alkaline limestone-dominated agricultural catchments or by the addition of lime to acidic or circumneutral soils to buffer soil pH. However, there is no consistent hypothesis which can be argued, based on the evidence currently published in the literature. While little is currently known regarding DON production under different agricultural regimes and cropping species, Oelmann et al. (2007) found increased DON losses under legume production. Similarly, in pastoral systems stocking densities will likely impact both DOM input to soils and waters from both urine and manure patches. These have been reported as hotspots of nutrient cycling (Clough et al. 1998) and microbial DOM uptake and biosynthesis pathways in both soils (Reay et al. 2019) and waters (Brailsford et al. 2019a). Meanwhile those studying N speciation and P fractionation in intensive dairy farming catchments underlain by clay report higher DON and DOP concentrations than in nearby limestone catchments with similar topography and climate (Lloyd et al. 2019).
While studies into the quantitative importance of sewage treatment works (STW) discharges of both N and P to streams are now commonplace in the literature (e.g. Bowes et al. 2005), their influence on the rates and composition of DOM flux to streams is understudied. The direct discharge of treated and, under high flow conditions, untreated effluent from sewerage systems may act as a proximal source of a bioavailable N and P-enriched DOM for the stream biota alongside inorganic N and P fractions for which bioavailability is well-established in the literature. While few studies have investigated the discharge of DOP from STW (see Qin et al. 2015 and Yates et al. 2019b for recent examples), DON has been found to account for up to 65% of TDN in effluents at conventional treatment works, and up to 80% of TDN in treatment plants that utilise nitrification–denitrification systems (Pehlivanoglu-Mantas and Sedlak 2006). Compounds commonly found in effluent DOM, as DOC, DON and DOP have been shown to be highly bioavailable to riverine biota (Bronk et al. 2007, 2010; Liu et al. 2012; Qin et al. 2015; Brailsford et al. 2019a; Mackay et al. 2020). However, the C, N and P composition of effluent has also been shown to be highly variable between treatment facility types, even within the same geographic region, reflecting both the nature and efficacy of treatment processes, and the nature of materials sent to each facility (Yates et al. 2019b). Thus, clear trends are not evident in the literature available to date, and broad conclusions about the DOM composition of effluents cannot be robustly inferred. This may explain the lack of any discernible relationship between DOC and DON in limestone catchments: the presence of scattered farmsteads and villages often served by septic tanks and minor STW in these catchments will lead to the discharge of N and P-rich DOM to the stream which reflects the materials sent to and the processes in place at these treatment or storage facilities. This will have bypassed the soil microbial processing that has been found to exert a dominant control on the composition of nutrient fluxes in natural and semi-natural catchments (Wymore et al. 2021; Yates et al. 2019a).
Weak associations between absorbance DOM absorption coefficients and DOC concentration have been reported as more prevalent in more anthropogenically impacted waters (Spencer et al. 2012). This is due to variation in DOM composition between source areas, as allochthonous (highly chromophoric) organic matter is mainly composed of decaying vegetation with a higher aromatic content, (and concurrently high absorbance at lower wavelengths) than autochthonous DOM produced in-stream or discharged from anthropogenic sources, such as sewage discharges or livestock manures and slurries. This material is chemically diverse, of lower molecular weight, and results in an increase in the variability of DOM stoichiometry in catchments with this complexity of contributing sources.
The data presented here suggest the move along a gradient of nutrient enrichment to more N and P enriched systems is accompanied by a shift in DOM composition towards low C/NDOM ratios, low SUVA254 values, and increasing DON/DOP concentrations. This is visualised in the conceptual diagram presented in Fig. 6, representing an important shift in the nature, composition, and concentration of DOM in streams as they undergo nutrient enrichment. This displays a similarly pattern to those reported between DOC and nitrate in a range of other studies (e.g. Evans et al. 2006; Taylor and Townsend 2010; Rodriguez-Cardona et al. 2021) and between DON and TDN in the global analysis of Wymore et al. (2021). Given the range of scales covered in each of these studies, and in the research presented here, these patterns may well be ubiquitous in fresh waters, representing a transition from N (or P) limitation in low nutrient status waters, typically natural or semi-natural, to systems which are highly enriched with N and P where neither N not P are limiting to production. System stoichiometry then shifts as systems move along this transition in response to external forcing or mitigation efforts in catchments. A comparable theory is proposed for terrestrial ecosystems, linking N saturation to C limitation of soil microbial processes and community composition (Kopacek et al. 2013), and it is logical to argue that a similar impact on stream microbial communities and processing of DOM is likely. Stream DOM pools are not limited to soil-derived DOM, as is evident in this study, simultaneously receiving both inorganic and organic N and P from a mixture of point and diffuse sources as the system enriches. This presents the stream biota with wide range of bioavailable material to support both autotrophic and heterotrophic production (Mackay et al. 2020), which has the potential to induce further mineralisation of previously recalcitrant organic matter through the process of nutrient priming (Ward et al. 2016), stimulating rapid biological uptake and biogeochemical processing of DOM in streams as they undergo enrichment. A negative y intercept value for all DOC/chromophoric DOM relationships also suggests that not all DOM is chromophoric (Pereira et al. 2014; Zhang et al. 2020).

Conceptual diagram of the influence of nutrient enrichment on DOM composition in catchments of differing geoclimatic and landscape character. Letters and colours represent dominant geoclimatic groups a Peat, b Clay, and c Limestone
While all relationships between DOC and a254 observed in this study are significant, limestone sites demonstrate the lowest r2 value of all classifications. Below 2 mg C l−1, the relationship between DOC and a254 appears to weaken. Significant relationships were found in this study between BFI and DOM compositional metrics such as C/NDOM, and SUVA254, particularly in clay and peat catchments, but with a weaker relationship evident in limestone catchments which are driven more strongly by anthropogenic nutrient inputs than soil DOM character. Low C/NDOM, and low SUVA254 values may indicate catchment delivery of N rich material from potentially flow-independent proximal sources such as sewage works, and diffuse runoff from agricultural land, driving a de-coupling of DOC and DON concentrations in-stream similar to those reported for other groundwater-fed headwater streams (Bernal et al. 2018). This decoupling may also reflect the use of DOC primarily as an energy resource (Battin et al. 2009; Bernal et al. 2018) and DON as both an energy and nutrient resource for stream biota (Kaushal and Lewis 2005; Wymore et al. 2015). Clay systems, by contrast, appear to reflect both proximal sewage discharge, the application of N, P and C-rich manures and slurries to grassland and diffuse runoff from these enriched sources, and soil DOM character in their signal, which is intermediate in slope between peat (soil source-dominated signal) and limestone (anthropogenic source-dominated signal). What is clear in this data set is that combining measurements of organically bound N, P and C with DOM compositional metrics allows identification of the key source areas contributing nutrient loading to streams, and the drivers of DOM composition in catchments as this varies according to environmental character.
Lastly, sampling was conducted during this study across the summer months as key biological controls on the export of differing sources of DOM will be less important during the winter months, as will the impact of land management practises e.g., key agricultural practices. While this study focussed solely on site specific spatial differences between locations, there may also be considerable variability in temporal patterns owing to differing in-stream communities controlling nutrient uptake. While temporal variability across and between sites was not monitored in this study, this has been the subject of prior investigation by the project team (e.g., Outram et al. 2014; Lloyd et al. 2016, 2019; Yates et al. 2019a, b), and may be of interest to address in future work delving into temporal variation in DOM character and composition as this varies between a wider range of environment types.
Conclusion
Despite a large degree of heterogeneity in the fresh water DOM pool, we demonstrate that catchments with similar geoclimatic character, land use and management export DOM with similar composition and character to the fresh waters that drain these landscapes. We demonstrate that the total N and P pools in temperate streams draining natural or semi-natural peatland catchments tend to exhibit low inorganic N and P concentrations and are dominated by high proportions, but low absolute concentrations, of both DON and DOP. The DOM exported from these systems also tends to display high DOC concentrations and optical indices indicative of stream DOM of allochthonous origin, reflective of soil DOM pools and the breakdown of plant matter. By contrast, in anthropogenically-disturbed catchments, enriched with N and P from fertiliser use and the excretion of wastes by farm livestock and people, a de-coupling of C/NDOM ratios is evident as systems move from nutrient-limited to nutrient-saturated. This is particularly so for DOP where composition of the DOP is strongly influenced by the discharge of effluent from sewerage systems in most intensively farmed and populated catchments. The composition of the stream nutrient pool is therefore strongly linked to landscape character and management, and the ecosystem functional role of the DOM compounds that comprise these pools will vary accordingly. The challenge for future research is to identify the specific molecular signature of this DOM, as this varies according to anthropogenic disturbance and environmental character, and to pinpoint the functional mechanisms by which the biota access this material. Without this understanding we will be unable to predict shifts in stream ecosystem function, and ecological responses to enrichment or to mitigation efforts, including species composition shifts and biodiversity loss in freshwater ecosystems.
Data availability
Enquiries about data availability should be directed to the authors.
References
Ascott MJ, Gooddy DC, Wang L, Stuart ME, Lewis MA, Ward RS, Binley AM (2017) Global patterns of nitrate storage in the vadose zone. Nat Commun 8:1416. https://doi.org/10.1038/s41467-017-01321-w
Austnes K, Evans CD, Eliot-Laize C, Naden PS, Old GH (2010) Effects of storm events on mobilisation and in-stream processing of dissolved organic matter (DOM) in a Welsh peatland catchment. Biogeochemistry 99(1–3):157–173. https://doi.org/10.1007/s10533-009-9399-4
Battin TJ, Luyssaert S, Kaplan LA, Aufdenkampe AK, Richter A, Tranvik LJ (2009) The boundless carbon cycle. Nat Geosci 2(9):598–600. https://doi.org/10.1038/ngeo618
Benk SA, Yan L, Lehmann R, Roth V-N, Schwab VF, Totsche KU, Kuesel K, Gleixner G (2019) Fueling diversity in the subsurface: composition and age of dissolved organic matter in the critical zone. Front Earth Sci 7:296. https://doi.org/10.3389/feart.2019.00296
Bernal S, Lupon A, Catalan N, Castelar S, Marti E (2018) Decoupling of dissolved organic matter patterns between stream and riparian groundwater in a headwater forested catchment. Hydrol Earth Syst Sci 22:1897–1910. https://doi.org/10.5194/hess-22-1897-2018
Bowes MJ, Hilton J, Irons GP, Hornby DD (2005) The relative contribution of sewage and diffuse phosphorus sources in the River Avon catchment, southern England: implications for nutrient management. Sci Total Environ 344(1–3):67–81. https://doi.org/10.1016/j.scitotenv.2005.02.006
Brailsford FL, Glanville HC, Golyshin PN, Johnes PJ, Yates CA, Jones DL (2019a) Microbial uptake kinetics of dissolved organic carbon (DOC) compound groups from river water and sediments. Sci Rep-Uk 9:11229. https://doi.org/10.1038/s41598-019-47749-6
Brailsford FL, Glanville HC, Golyshin PN, Marshall MR, Lloyd CE, Johnes PJ, Jones DL (2019b) Nutrient enrichment induces a shift in dissolved organic carbon (DOC) metabolism in oligotrophic freshwater sediments. Sci Total Environ 690:1131–1139. https://doi.org/10.1016/j.scitotenv.2019.07.054
Brailsford FL, Glanville HC, Marshall MR, Yates CA, Owen AT, Golyshin P, Johnes PJ, Jones DL (2021) Land cover and nutrient enrichment regulates low molecular weight dissolved organic matter turnover in freshwater ecosystems. Limnol Oceanogr. https://doi.org/10.1002/lno.11852
Bronk DA, See JH, Bradley P, Killberg L (2007) DON as a source of bioavailable nitrogen for phytoplankton. Biogeosciences 4(3):283–296. https://doi.org/10.5194/bg-4-283-2007
Bronk DA, Roberts QN, Sanderson MP, Canuel EA, Hatcher PG, Mesfioui KC, Mulholland MR, Love NG (2010) Effluent organic nitrogen (EON): bioavailabity and photochemical and salinit-mediated release. Environ Sci Technol 44(15):5830–5835. https://doi.org/10.1021/es101115g
Brookshire ENJ, Valett HM, Thomas SA, Webster JR (2007) Atmospheric N deposition increases organic N loss from temperate forests. Ecosystems 10(2):252–262. https://doi.org/10.1007/s10021-007-9019-x
Clough TJ, Ledgard SF, Sprosen MS, Kear MJ (1998) Fate of N-15 labelled urine on four soil types. Plant Soil 199(2):195–203. https://doi.org/10.1023/A:1004361009708
Cory RM, Harrold KH, Neilson BT, Kling GW (2015) Controls on dissolved organic matter (DOM) degradation in a headwater stream: the influence of photochemical and hydrological conditions in determining light-limitation or substrate-limitation of photo-degradation. Biogeosciences 12(22):6669–6685. https://doi.org/10.5194/bg-12-6669-2015
Drake TW, Van Oost K, Barthel M, Bauters M, Hoyt AM, Podgorski DC, Six J, Boeckx P, Trumbore SE, Ntaboba LC, Spencer RGM (2019) Mobilization of aged and biolabile soil carbon by tropical deforestation. Nat Geosci 12(7):541–546. https://doi.org/10.1038/s41561-019-0384-9
Durand P, Breuer L, Johnes PJ, Billen G, Butturini A, Pinay G, Grinsven HV, Garnier J, Rivett M, Reay DS, Curtis C, Siemens J, Maberly S, Kaste O, Humborg C, Loeb R, de Klein J, Hejzlar J, Skoulikidis N, Kortelainen P, Lepisto A, Wright R (2011) Nitrogen processes in aquatic ecosystems. Cambridge University Press, pp 126–146
Evans CD, Reynolds B, Jenkins A, Helliwell RC, Curtis CJ, Goodale CL, Ferrier RC, Emmett BA, Pilkington MG, Caporn SJM, Carroll JA, Norris D, Davies J, Coull MC (2006) Evidence that soil carbon pool determines susceptibility of semi-natural ecosystems to elevated nitrogen leaching. Ecosystems 9:453–462. https://doi.org/10.1007/s10021-006-0051-z
Fasching C, Ulseth AJ, Schelker J, Steniczka G, Battin TJ (2016) Hydrology controls dissolved organic matter export and composition in an Alpine stream and its hyporheic zone. Limnol Oceanogr 61(2):558–571. https://doi.org/10.1002/lno.10232
Geisseler D, Scow KM (2014) Long-term effects of mineral fertilizers on soil microorganisms—a review. Soil Biol Biochem 75:54–63. https://doi.org/10.1016/j.soilbio.2014.03.023
George TS, Giles CD, Menezes-Blackburn D et al (2018) Organic phosphorus in the terrestrial environment: a perspective on the state of the art and future priorities. Plant Soil 427:191–208. https://doi.org/10.1007/s11104-017-3391-x
Graeber D, Gelbrecht J, Pusch MT, Anlanger C, von Schiller D (2012) Agriculture has changed the amount and composition of dissolved organic matter in Central European headwater streams. Sci Total Environ 438:435–446. https://doi.org/10.1016/j.scitotenv.2012.08.087
Graeber D, Boechat IG, Encina-Montoya F, Esse C, Gelbrecht J, Goyenola G, Guecker B, Heinz M, Kronvang B, Meerhoff M, Nimptsch J, Pusch MT, Silva RCS, von Schiller D, Zwirnmann E (2015) Global effects of agriculture on fluvial dissolved organic matter. Sci Rep 5:16328. https://doi.org/10.1038/srep16328
Greene S, Johnes PJ, Bloomfield JP, Reaney SM, Lawley R, Elkhatib Y, Freer J, Odoni N, Macleod CJA, Percy B (2015) A geospatial framework to support integrated biogeochemical modelling in the United Kingdom. Environ Model Softw 68:219–232. https://doi.org/10.1016/j.envsoft.2015.02.012
Heinz M, Graeber D, Zak D, Zwirnmann E, Gelbrecht J, Pusch MT (2015) Comparison of organic matter composition in agricultural versus forest affected headwaters with special emphasis on organic nitrogen. Environ Sci Technol 49(4):2081–2090. https://doi.org/10.1021/es505146h
Helliwell RC, Coull MC, Davies JJL, Evans CD, Norris D, Ferrier RC, Jenkins A, Reynolds B (2007) The role of catchment characteristics in determining surface water nitrogen in four upland regions in the UK. Hydrol Earth Syst Sci 11:356–371. https://doi.org/10.5194/hess-11-356-2007
Heppell CM, Binley A, Trimmer M, Darch T, Jones A, Malone E, Collins AL, Johnes PJ, Freer JE, Lloyd CEM (2017) Hydrological controls on DOC: nitrate resource stoichiometry in a lowland, agricultural catchment, southern UK. Hydrol Earth Syst Sci 21(9):4785–4802. https://doi.org/10.5194/hess-21-4785-2017
Hernes PJ, Spencer RGM, Dyda RY, Pellerin BA, Bachand PAM, Bergamaschi BA (2008) The role of hydrologic regimes on dissolved organic carbon composition in an agricultural watershed. Geochim Cosmochim Acta 72(21):5266–5277. https://doi.org/10.1016/j.gca.2008.07.031
Inamdar S, Singh S, Dutta S, Levia D, Mitchell M, Scott D, Bais H, McHale P (2011) Fluorescence characteristics and sources of dissolved organic matter for stream water during storm events in a forested mid-Atlantic watershed. J Geophys Res Biogeosci. https://doi.org/10.1029/2011JG001735
Jaffe R, McKnight D, Maie N, Cory R, McDowell WH, Campbell JL (2008) Spatial and temporal variations in DOM composition in ecosystems: the importance of long-term monitoring of optical properties. J Geophys Res Biogeosci. https://doi.org/10.1029/2008JG000683
Jarvie HP, Neal C, Withers PJA, Wescott C, Acornley RA (2005) Nutrient hydrochemistry for a groundwater-dominated catchment: the Hampshire Avon, UK. Sci Total Environ 344(1–3):143–158. https://doi.org/10.1016/j.scitotenv.2005.02.012
Jian SY, Li JW, Chen J, Wang GS, Mayes MA, Dzantor KE, Hui DF, Luo YQ (2016) Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: a meta-analysis. Soil Biol Biochem 101:32–43. https://doi.org/10.1016/j.soilbio.2016.07.003
Johnes PJ (2007) Uncertainties in annual riverine phosphorus load estimation: impact of load estimation methodology, sampling frequency, baseflow index and catchment population density. J Hydrol 332(1–2):241–258. https://doi.org/10.1016/j.jhydrol.2006.07.006
Johnes PJ, Heathwaite AL (1992) A procedure for the simultaneous determination of total nitrogen and total phosphorus in fresh-water samples using persulfate microwave digestion. Water Res 26(10):1281–1287. https://doi.org/10.1016/0043-1354(92)90122-K
Jordan P, Arnscheidt A, McGrogan H, McCormick S (2007) Characterising phosphorus transfers in rural catchments using a continuous bank-side analyser. Hydrol Earth Syst Sci 11:372–381. https://doi.org/10.5194/hess-11-372-2007
Judd KE, Crump BC, Kling GW (2007) Bacterial responses in activity and community composition to photo-oxidation of dissolved organic matter from soil and surface waters. Aquat Sci 69(1):96–107. https://doi.org/10.1007/s00027-006-0908-4
Kaushal SS, Lewis WM (2005) Fate and transport of organic nitrogen in minimally disturbed montane streams of Colorado, USA. Biogeochemistry 74:303–321. https://doi.org/10.1007/s10533-004-4723-5
Kopacek J, Cosby BJ, Evans CD, Hruska J, Moldan F, Oulehle F, Santruckova H, Tahovska K, Wright RF (2013) Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry 115(1–3):33–51. https://doi.org/10.1007/s10533-013-9892-7
Liu H, Jeong J, Gray H, Smith S, Sedlak DL (2012) Algal uptake of hydrophobic and hydrophilic dissolved organic nitrogen in effluent from biological nutrient removal municipal wastewater treatment systems. Environ Sci Technol 46(2):713–772. https://doi.org/10.1021/es203085y
Lloyd CEM, Freer JE, Johnes PJ, Collins AL (2016) Using hysteresis analysis of high-resolution water quality monitoring data, including uncertainty, to infer controls on nutrient and sediment transfer in catchments. In: Science of the total environment, vol 543, pp 388–404
Lloyd CEM, Johnes PJ, Freer JE, Carswell A, Jones JI, Stirling MW, Hodgkinson RA, Richmond C, Collins AL (2019) Determining the origins of nutrient flux to waters in catchments: examining the nutrient speciation balance to inform the targeting of mitigation measures. Sci Total Environ 648:1179–1200. https://doi.org/10.1016/j.scitotenv.2018.08.190
Lloyd CEM, Mena-Rivera L, Pemberton J, Johnes PJ, Jones D, Yates CA, Brailsford FL, Glanville HC, McIntyre C, Evershed RP (2021) Characterisation of riverine dissolved organic matter using a hierarchy of mass spectrometric methods. Biogeochemistry. https://doi.org/10.1007/s10533-022-00913-z
Mackay EB, Feuchtmayr H, De Ville MM, Thackeray SJ, Callaghan N, Marshall M, Rhodes G, Yates CA, Johnes PJ, Maberly SC (2020) Dissolved organic nutrient uptake by riverine phytoplankton varies along a gradient of nutrient enrichment. Sci Total Environ 722:137837. https://doi.org/10.1016/j.scitotenv.2020.137837
Mattsson T, Kortelainen P, Laubel A, Evans D, Pujo-Pay M, Raike A, Conan P (2009) Export of dissolved organic matter in relation to land use along a European climatic gradient. Sci Total Environ 407(6):1967–1976. https://doi.org/10.1016/j.scitotenv.2008.11.014
McDonough LK, O’Carroll DM, Meredith K, Andersen MS, Brugger C, Huang H, Rutlidge H, Behnke MI, Spencer RGM, McKenna A, Marjo CE, Oudone P, Baker A (2020) Changes in groundwater dissolved organic matter character in a coastal sand aquifer due to rainfall recharge. Water Res 169:115201. https://doi.org/10.1016/j.watres.2019.115201
McDowell WH (2003) Dissolved organic matter in soils—future directions and unanswered questions. Geoderma 113(3–4):179–186. https://doi.org/10.1016/S0016-7061(02)00360-9
McGlynn BL, McDonnell JJ (2003) Role of discrete landscape units in controlling catchment dissolved organic carbon dynamics. Water Resour Res. https://doi.org/10.1029/2002WR001525
Mena-Rivera L, Lloyd CEM, Reay MK, Goodall T, Read DS, Johnes PJ, Evershed RP (2021) Tracing carbon and nitrogen microbial assimilation in suspended particles in freshwaters. Biogeochemistry. https://doi.org/10.1007/s10533-022-00915-x
Mueller B, den Haan J, Visser PM, Vermeij MJA, van Duyl FC (2016) Effect of light and nutrient availability on the release of dissolved organic carbon (DOC) by Caribbean turf algae. Sci Rep. https://doi.org/10.1038/srep23248
Nguyen HVM, Hur J, Shin HS (2010) Changes in spectroscopic and molecular weight characteristics of dissolved organic matter in a river during a storm event. Water Air Soil Pollut 212(1–4):395–406. https://doi.org/10.1007/s11270-010-0353-9
Oelmann Y, Kreutziger Y, Temperton VM, Buchmann N, Roscher C, Schumacher J, Schulze ED, Weisser WW, Wilcke W (2007) Nitrogen and phosphorus budgets in experimental grasslands of variable diversity. J Environ Qual 36(2):396–407. https://doi.org/10.2134/jeq2006.0217
Outram FN, Lloyd CEM, Jonczyk J, McW H Benskin C, Grant F, Perks MT, Deasy C, Burke SP, Collins AL, Freer J, Haygarth PM, Hiscock KM, Johnes PJ, Lovett AL (2014) High-frequency monitoring of nitrogen and phosphorus response in three rural catchments to the end of the 2011–2012 drought in England. In: Hydrology and earth system sciences, vol 18, 9, pp 3429–3448
Pehlivanoglu-Mantas E, Sedlak DL (2006) Wastewater-derived dissolved organic nitrogen: analytical methods, characterization, and effects—a review. Crit Rev Environ Sci Technol 36(3):261–285. https://doi.org/10.1080/10643380500542780
Pellerin BA, Kaushal SS, McDowell WH (2006) Does anthropogenic nitrogen enrichment increase organic nitrogen concentrations in runoff from forested and human-dominated watersheds? Ecosystems 9(5):852–864. https://doi.org/10.1007/s10021-006-0076-3
Perakis SS, Hedin LO (2002) Nitrogen loss from unpolluted South American forests mainly via dissolved organic compounds. Nature 415:416–419. https://doi.org/10.1038/415416a
Pereira R, Bovolo CI, Spencer RGM, Hernes PJ, Tipping E, Vieth-Hillebrand A, Pedentchouk N, Chappell NA, Parkin G, Wagner T (2014) Mobilization of optically invisible dissolved organic matter in response to rainstorm events in a tropical forest headwater river. Geophys Res Lett 41(4):1202–1208. https://doi.org/10.1002/2013GL058658
Pisani O, Bosch DD, Coffin AW, Endale DM, Liebert D, Strickland TC (2020) Riparian land cover and hydrology influence stream dissolved organic matter composition in an agricultural watershed. Sci Total Environ 717:137165. https://doi.org/10.1016/j.scitotenv.2020.137165
Qin C, Liu H, Liu L, Smith S, Sedlak DL, Gu AZ (2015) Bioavailability and characterization of dissolved organic nitrogen and dissolved organic phosphorus in wastewater effluents. Sci Total Environ 511:47–53. https://doi.org/10.1016/j.scitotenv.2014.11.005
Reay M, Charteris AF, Jones DL, Evershed RP (2019) 15N-amino sugar stable isotope probing (15N-SIP) to trace the assimilation of fertiliser-N by soil bacterial and fungal communities. Soil Biol Biochem 138:107599. https://doi.org/10.1016/j.soilbio.2019.107599
Rodriguez-Cardona BM, Wymore AS, Argerich A, Bernal S, Brookshire ENJ, Fazekas HM, Helton AM, Argerich A, Barnes RT, Bernal S, Coble AA, Dodds WK, Fazekas HM, Helton AM, Johnes PJ, Johnson SL, Jones JB, Kaushal SS, Kortelainen P, Lopez-Lloreda C, Spencer RGM, McDowell WH (2021) Shifting stoichiometry: long-term trends in stream dissolved organic matter reveal altered C:N ratios due to history of atmospheric acid deposition. Glob Change Biol. https://doi.org/10.1111/gcb.15965
Sachse A, Henrion R, Gelbrecht J, Steinberg CEW (2005) Classification of dissolved organic carbon (DOC) in river systems: influence of catchment characteristics and autochthonous processes. Org Geochem 36(6):923–935. https://doi.org/10.1016/j.orggeochem.2004.12.008
Spencer RGM, Stubbins A, Hernes PJ, Baker A, Mopper K, Aufdenkampe AK, Dyda RY, Mwamba VL, Mangangu AM, Wabakanghanzi JN, Six J (2009) Photochemical degradation of dissolved organic matter and dissolved lignin phenols from the Congo River. J Geophys Res Biogeosci. https://doi.org/10.1029/2009JG000968
Spencer RGM, Butler KD, Aiken GR (2012) Dissolved organic carbon and chromophoric dissolved organic matter properties of rivers in the USA. J Geophys Res Biogeosci. https://doi.org/10.1029/2011JG001928
Stackpoole SM, Stets EG, Sprague LA (2019) Variable impacts of contemporary versus legacy agricultural phosphorus on US river water quality. PNAS 116(41):20562–20567. https://doi.org/10.1073/pnas.1903226116
Stanley EH, Maxted JT (2008) Changes in the dissolved nitrogen pool across land cover gradients in Wisconsin streams. Ecol Appl 18(7):1579–1590. https://doi.org/10.1890/07-1379.1
Taylor P, Townsend A (2010) Stoichiometric control of organic carbon–nitrate relationships from soils to the sea. Nature 464:1178–1181. https://doi.org/10.1038/nature08985
Thompson SK, Cotner JB (2018) Bioavailability of dissolved organic phosphorus in temperate lakes. Front Environ Sci. https://doi.org/10.3389/fenvs.2018.00062
Toming K, Tuvikene L, Vilbaste S, Agasild H, Viik M, Kisand A, Feldmann T, Martma T, Jones RI, Noges T (2013) Contributions of autochthonous and allochthonous sources to dissolved organic matter in a large, shallow, eutrophic lake with a highly calcareous catchment. Limnol Oceanogr 58(4):1259–1270. https://doi.org/10.4319/lo.2013.58.4.1259
van Kessel C, Clough T, van Groenigen JW (2009) Dissolved organic nitrogen: an overlooked pathway of nitrogen loss from agricultural systems? J Environ Qual 38(2):393–401. https://doi.org/10.2134/jeq2008.0277
Wagner S, Fair JH, Matt S, Hosen JD, Raymond P, Saiers J, Shanley JB, Dittmar T, Stubbins A (2019) Molecular hysteresis: hydrologically driven changes in riverine dissolved organic matter chemistry during a storm event. J Geophys Res Biogeosci 124(4):759–774. https://doi.org/10.1029/2018JG004817
Wang L, Stuart ME, Lewis MA, Ward RS, Skirvin D, Naden PS, Collins AL, Ascott MJ (2016) The changing trend in nitrate concentrations in major aquifers due to historical nitrate loading from agricultural land across England and Wales from 1925 to 2150. Sci Total Environ 542:694–705. https://doi.org/10.1016/j.scitotenv.2015.10.127
Ward ND, Bianchi TS, Sawakuchi HO, Gagne-Maynard W, Cunha AC, Brito DC, Neu V, Valerio AD, da Silva R, Krusche AV, Richey JE, Keil RG (2016) The reactivity of plant-derived organic matter and the potential importance of priming effects along the lower Amazon River. J Geophys Res Biogeosci 121(6):1522–1539
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K (2003) Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37(20):4702–4708. https://doi.org/10.1021/es030360x
Williams CJ, Frost PC, Morales-Williams AM, Larson JH, Richardson WB, Chiandet AS, Xenopoulos MA (2016) Human activities cause distinct dissolved organic matter composition across freshwater ecosystems. Glob Change Biol 22(2):613–626. https://doi.org/10.1111/gcb.13094
Williamson JL, Tye A, Lapworth DJ, Monteith D, Sanders R, Mayor DJ, Barry C, Bowes M, Burden A, Callaghan N, Farr G, Felgate S, Fitch A, Gibb S, Gilbert P, Hargreaves G, Keenan P, Kitidis V, Juergens M, Martin A, Mounteney I, Nightingale PD, Pereira MG, Olszewska J, Pickard A, Rees A, Spears B, Stinchcombe M, White D, Williams P, Worrall F, Evans C (2021) Landscape controls on riverine export of dissolved organic carbon from Great Britain. Biogeochemistry. https://doi.org/10.1007/s10533-021-00762-2
Wilson HF, Xenopoulos MA (2009) Effects of agricultural land use on the composition of fluvial dissolved organic matter. Nat Geosci 2(1):37–41. https://doi.org/10.1038/srep16328
Withers PJA, May L, Jarvie HP, Jordan P, Doody D, Foy RH, Bechmann M, Cooksley S, Dils R, Deal N (2012) Nutrient emissions to water from septic tank systems in rural catchments: uncertainties and implications for policy. Environ Sci Policy 24:71–82. https://doi.org/10.1016/j.envsci.2012.07.023
Wymore AS, Rodriguez-Cardona B, McDowell WH (2015) Direct response of dissolved organic nitrogen to nitrate availability in headwater streams. Biogeochemistry 126:1–10. https://doi.org/10.1007/s10533-015-0153-9
Wymore A, Johnes PJ, Bernal S, Brookshire ENJ, Fazekas HM, Helton AM, Argerich A, Barnes RT, Coble AA, Dodds WK, Haq S, Johnson SL, Jones JB, Kaushal SS, Kortelainen P, Lopez-Lloreda C, Rodriquez-Cardona B, Spencer RGM, Sullican PL, Yates CA, McDowell WH (2021) Gradients of anthropogenic nutrient enrichment alter N composition and DOM stoichiometry in freshwater ecosystems. Glob Biogeochem Cycles. https://doi.org/10.1029/2021GB006953
Yates CA, Johnes PJ, Spencer RGM (2016) Assessing the controls on dissolved organic matter export from two contrasting lowland catchments. Sci Total Environ 569–570:1330–1340. https://doi.org/10.1016/j.scitotenv.2016.06.211
Yates CA, Johnes PJ, Owen AT, Brailsford FL, Glanville HC, Evans CD, Marshall MR, Jones DL, Lloyd CEM, Jickells T, Evershed RP (2019a) Variation in dissolved organic matter (DOM) stoichiometry in UK freshwaters: assessing the influence of land cover and soil C:N ratio on DOM composition. Limnol Oceanogr 64(6):2328–2340. https://doi.org/10.1002/lno.11186
Yates CA, Johnes PJ, Spencer RGM (2019b) Characterisation of treated effluent from four commonly employed wastewater treatment facilities: a UK case study. J Environ Manag 232:919–927. https://doi.org/10.1016/j.jenvman.2018.12.006
Zhang ZY, Qin JH, Sun H, Yang JY, Liu YY (2020) Spatiotemporal dynamics of dissolved organic carbon and freshwater browning in the Zoige Alpine Wetland, Northeastern Qinghai-Tibetan Plateau. Water 12(9):2453. https://doi.org/10.3390/w12092453
Acknowledgements
This study was undertaken as part of the Natural Environment Research Council DOMAINE Large Grant programme (NE/K010689/1), Characterising the nature, origins, and ecological significance of dissolved organic matter in freshwater ecosystems. The authors wish to acknowledge the local landowners for their co-operation in permitting access to sampling sites, and our many colleagues who have researched and published on these sites previously and who provided advice and access to data and sampling infrastructure for this study.
Funding
This study was supported by Natural Environment Research Council (Grant No. NE/K010689/1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors have not disclosed any competing interests.
Additional information
Responsible Editor: Stephen Maberly
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yates, C.A., Johnes, P.J., Brailsford, F.L. et al. Determining patterns in the composition of dissolved organic matter in fresh waters according to land use and management. Biogeochemistry 164, 143–162 (2023). https://doi.org/10.1007/s10533-022-00964-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-022-00964-2